economia da natureza ricklefs - 6ªed - cap. 16
TRANSCRIPT

Competição
O botânico britânico A. G. Tansley (1917) proporcionou a primeira demonstração dacompetição entre espécies num estudo experimental. Tansley prefaciou seu relatóriocom a observação de que espécies de plantas proximamente aparentadas vivendo
na mesma região frequentemente crescem em habitats diferentes ou em tipos diferentes desolo. A observação de Tansley não era nova, nem sua sugestão de que uma tal segregaçãoecológica poderia ser o resultado da competição por recursos, levando à exclusão de umaespécie ou da outra. Contudo, ninguém tinha testado experimentalmente esta hipótese, ousua alternativa - isto é, de que as duas espécies tinham tais exigências ecológicas diferentesque cada uma não poderia crescer onde a outra florescia.
Tansleyselecionou duas espécies do gênero Ga/ium, que são pequenas plantas herbácease perenes. Uma espécie, G. saxotile, normalmente vive em solos ácidos e turfosos; a outra,G. sy/vestre, habita os solos alcalinos de morros de calcário e pastos. Tansley plantou semen-tes para cada espécie, tanto separadamente quanto juntas, em áreas com solos tirados dasáreas onde cada espécie crescia naturalmente. Como as sementes foram plantadas juntasnum jardim comum, as únicas diferenças nas áreas eram o tipo de solo e a presença ou au-sência da outra espécie (Fig. 16.1).
Como muitos outros estudos ecológicos, os experimentos de Tansley foram atrapalhadospor problemas técnicos, como germinação fraca e falta de água. Seus resultados, contudo,foram bastante claros. Quando plantadas sozinhas, cada uma das espécies cresceu e semanteve em ambos os tipos de solo, embora a germinação e o crescimento fossem mais ri-gorosos no tipo de solo no qual as espécies crescem naturalmente. Quando as duas espéciesforam cultivadas juntas em solos calcá rios, as plantas de G. sy/vestre crescer9m muito maise sombrearam G. saxati/e. O inverso ocorreu no solo turfoso e mais ácido típico do habitatde G. saxati/e.
Tansley concluiu que G. saxati/e está em desvantagem nos solos calcários, e assim é inca-paz de competir eficientemente com G. sy/vestre naquele tipo de solo. Analogamente, G.sy/vestre não cresce tão bem na turfa e consequentemente é um competidor inferior para G.saxati/e naquele tipo de solo. Ambas as espécies, contudo, foram capazes de se estabelecerem ambos os solos. Estes resultados sugeriram a Tansley que as espécies estão geralmenterestritas aos tipos de solo mais favoráveis quando espécies competidoras estão presentes,mas poderiam estar mais amplamente distribuídas em outros tipos de solo na ausência decompetição. Uma espécie que cresce fracamente num determinado solo - como no caso de
291

292 Competição
G. sylvestrecresce melhor emsolos calcá rios,como observadona natureza.
Quando as duassão plantadasjuntas, G. sylvestreprevalece emsolo calcário.
Jardim comum experimental(sol, chuva e temperatura iguais
para cada grupo de plantas)
G. saxatile.díltivada sozinhaG. saxatile crescemelhor em solos ácidose turfosos, comoobservado na natureza.
Solo calcário Solo ácido
FIG. 16.1 O projeto experimental básico de Tansley é ainda usado na maioria dos estudosmodernos de competição. Tansley cultivou duas espécies de Golium, sozinhos e juntos, em doistipos diferentes de solos num jardim comum.
Quando as duas sãoplantadas juntas,G. saxatile prevaleceem solo ácido.
G. saxatile em solos calcários - provavelmente não sobreviveria à competição, e assim es-taria ausente daquele tipo de solo em toda a sua abrangência.
Em seu curto artigo, Tansley registrou que (1) a presença ou ausência de uma espécie po-de ser determinada pela competição com outra espécie; (2) as condições do ambiente afe-taram o resultado da competição; e (3) a presença da atual segregação ecológica da espéciepode ser o resultado de uma competição no passado. Embora os ecólogos não tenham execu-tado estudos 'de competição novamente por mais de 15 anos, a abordagem de Tansley, oualguma modificação dela, é usada nos estudos mais modernos de competição entre espécies.
CONCEITOS DO CAPíTULO• Os consumidores competem por recursos• A falha das espécies em coexistir nas culturas de laboratório
levou ao princípio da exclusão competitiva• A teoria da competição e coexistência é uma extensão dos
modelos de crescimento logístico• A competição assimétrica pode ocorrer quando diferentes
fatores limitam as populações de competidores
Acompetição é qualquer uso ou defesa de um recurso por umindivíduo que reduz a disponibilidade daquele recurso para
outros indivíduos. A competição é uma das formas mais impor-tantes pelas quais as atividades dos indivíduos afetam o bem-estar de outros, pertençam eles à mesma espécie ou a espéciesdiferentes. A competição entre indivíduos da mesma espécie échamada de competição íntraespecífíca, e a competição entre
• A produtividade do habitat pode influenciar a competiçãoentre as espécies de plantas
• A competição pode ocorrer através da interferência direta• Os consumidores podem influenciar o resultado da competição
indivíduos de espécies diferentes é chamada de competição in-terespecífica.
Como vimos no Capítulo 14, quanto mais densa uma popu-lação, mais fortes serão os efeitos da competição entre os indi-víduos. A competição intraespecífica, como as interações con-sumidor-recurso, regula o crescimento populacional de um mo-do dependente da densidade. Além do mais, quando os fatores

A espécie B aumentaaté esta densidade edepleciona os recursosabaixo do nívelnecessário parasustentar a espécie A.Assim, B sobrepuja A.
Níveis derecursos
Espécie B ------"" ••••••- -----I necessários parasustentar asspécies A e B.
Espécie A---"'III..,---";-- ~
KA KBDensidade populacional
FIG. 16.2 Os competidores superiores podem persistirem níveismais baixos de recurso. À medido que os recursossão consumidos,eles declinam paro níveis que não mais sustentamo crescimento adi-cional do população de consumidores, e o população pode atingirum tamanho de equilíbrio (I<). Se a espécie A pode continuar o cres-cer no nível de recurso que abrevio o crescimento do espécie B, aespécie A sobrepujoró e por fim substituirá o espécie B.
genéticos influenciam a eficiência do uso do recurso, a evoluçãotende a aumentar a capacidade competitiva numa população.
A competição entre indivíduos de espécies diferentes podedeplecionar as populações de ambas; neste caso, cada espéciecontribui para a regulação da outra população, assim como paraa sua própria. Sob algumas condições, particularmente quandoa competição interespecífica é intensa, a população de uma es-pécie pode declinar e finalmente desaparecer. Por causa destepotencial, a competição é um fator importante na determinaçãode quais espécies podem coexistir num habitat.
(a)
Competição 293
o resultado da competição entre espécies depende de quãoeficientemente os indivíduos dentro de cada espécie exploramos recursos compartilhados. Os indivíduos em todas as popula-ções consomem recursos. Quando os recursos são escassos emrelação à demanda por eles, cada ato de consumo por um indi-víduo toma um recurso menos disponível para outros, assimcomo para si próprio. À medida que o consumo continua, os re-cursos declinam para níveis que não mais sustentam um cresci-mento adicional da população consumidora, e esta pode atingirum estado de equilíbrio. Quando uma população pode continuara crescer no nível do recurso que limita o crescimento de umasegunda população, a primeira acabará por substituir a segunda(Fig. 16.2). Assim, a competição e seus diversos resultados de-pendem da relação dos consumidores com seus recursos.
Neste capítulo, consideraremos alguns dos princípios geraisda competição entre espécies, ilustraremos os efeitos potenciaisda competição examinando os resultados de experimentos delaboratório e de campo, e demonstraremos a importância da com-petição dos sistemas naturais.
Os consumidores competem por recursoso ecólogo David Tilman, um professor da Universidade de Min-nesota e um pesquisador líder das interações entre espécies ve-getais, definiu um recurso como qualquer substância ou fatorque é tanto consumido por um organismo quanto sustenta taxasde crescimento populacionais crescentes à medida que sua dis-ponibilidade no ambiente aumenta. Três coisas são chaves paraesta definição. Primeiro, um recurso é consumido, e sua quanti-dade ou disponibilidade é portanto reduzida. Segundo, um con-sumidor usa um recurso para sua própria manutenção e cresci-mento. Assim, o alimento é sempre um recurso, e a água umrecurso para plantas e animais terrestres. Terceiro, quando a dis-ponibilidade do recurso é reduzida, os processos biológicos sãoafetados de tal forma a reduzir o crescimento da população deconsumidores.
FIG. 16.3 Para os animais sésseis, o espaço é um recurso importante. (o) Quando as cracas nos costões rochosos do Maine estãovivendo acima de suo abrangência ótimo na zona entremaré, sua densidade é baixo, e as larvas podem se assentar nos áreas nuas dis-poníveis. (b) Mais baixo na zona entremaré, os larvas podem se assentar somente sobre um indivíduo mais velho, e o adensamento impe-de um crescimento adicional do população. Cortesiado AmericanMuseumof Natural History.

294 Competição
o consumo inclui mais do que somente comer. Os animaissésseis - aqueles permanentemente anexados a um substrato- precisam de lugares disponíveis e abertos, porque para aque-les animais o espaço é um recurso. As cracas (Cirripedia) ane-xadas às rochas dentro da zona entremaré, por exemplo, precisamde espaço para crescer, e suas larvas exigem espaço para se as-sentarem e se desenvolverem em adultas (Fig. 16.3). O adensa-mento aumenta a mortalidade dos adultos e reduz a fecundidade,limitando o crescimento de adultos e o recrutamento de larvas.O espaço aberto promove a reprodução e o ecru ento. Assim,as cracas individuais "consomem" lugare aberto à medida quecolonizam e crescem sobre eles.
Os refúgios e outros lugares seguros são um outro tipo derecurso. Cada habitat tem um número limitado de buracos, fen-das ou fragmentos de cobertura densa, nas quais um indivíduopode escapar da predação ou buscar refúgio do clima inclemen-te. À medida que alguns indivíduos ocupam os melhores lugares,outros devem se assentar em lugares menos favoráveis; conse-quentemente, aqueles indivíduos têm um risco mais alto de seprejudicar ou morrer.
Que tipos de fatores não são recursos? A temperatura não éum recurso. As temperaturas mais altas podem elevar as taxasde reprodução, mas os indivíduos não consomem a temperatura,e um indivíduo não muda a temperatura de um ambiente poroutra. A temperatura e outros fatores físicos e biológicos nãoconsumíveis podem limitar o crescimento das populações, natu-ralmente, mas eles devem ser considerados separadamente dosrecursos.
A competição entre espécies proximamenteaparentadas e distantemente aparentadasCharles Darwin enfatizou que a competição deve ser mais inten-sa entre espécies intimamente aparentadas. Em On the Origin ofSpecies ("A Origem das Espécies") ele observou, "Como as es-pécies do mesmo gênero têm normalmente, embora nunca inva-riavelmente, alguma similaridade em hábitos e constituição, esempre em estrutura, a disputa será geralmente mais severa entreas espécies do mesmo gênero, quando elas entrarem em compe-tição umas com as outras, do que entre espécies de gêneros dis-tintos". Darwin raciocinou que uma estrutura semelhante indicauma função semelhante, especialmente em relação aos recursosconsumidos. Este comentário inspirou Tansley a examinar a com-petição entre duas espécies intimamente aparentadas no mesmogênero de plantas.
Embora o insight de Darwin esteja de modo geral correto, osorganismos distantemente aparentados podem também usar mui-tos dos mesmos recursos. As cracas e os mexilhões, assim comoas algas, as esponjas, os briozoários, os tunicados e outros, ocu-pam espaço na zona entremaré e ativamente competem um como outro pela apropriação e domínio do espaço. Tanto os peixesquanto as aves aquáticas predam sobre os invertebrados aquáti-cos. O krill antártico (Euphausia superba), um crustáceo seme-lhante ao camarão que abunda as águas subantárticas (Fig. 16.4),alimenta virtualmente todos os tipos de grandes animais mari-nhos, incluindo peixes, lulas, aves mergulhadoras, focas e baleias.As populações de focas e pinguins no Oceano do Sul recente-mente aumentaram, aparentemente porque a exploração comer-cial dizimou as populações de um grande grupo competidor, asbaleias. Nos habitats terrestres, as aranhas, besouros-de-solo,salamandras e aves consomem invertebrados que vivem na se-rapilheira da floresta. Nos ecos sistemas de deserto, as aves e os
FIG. 16.4 Fontes de alimento comuns fazem organismos distan-temente aparentados competir. No Oceano do Sul, o krill (Euphou-sio superbo) supre alimento paro uma amplo gomo de espéciesanimais marinhos. Cortesia de Dr. Uwe Kils.
lagartos comem muitas das mesmas espécies de insetos, e asformigas, roedores e aves consomem as sementes de muitas dasmesmas plantas. Estes exemplos ilustram o forte potencial paracompetição entre organismos de parentesco distante e nos lem-bram uma vez mais quão extensa e complexa é a teia das intera-ções entre as espécies numa comunidade biológica.
Recursos renováveis e não renováveisOs recursos podem ser classificados de acordo com o fato de elespoderem ser regenerados ou não. Os recursos não renováveis,tais como o espaço, não são regenerados. Uma vez ocupado, oespaço se torna indisponível; ele é "reposto" somente quando oconsumidor o abandona. Por outro lado, os recursos renováveissão constantemente regenerados, ou renovados. Os nascimentosnuma população de presas continuamente suprem itens de ali-mentos para os predadores, assim como a contínua decomposiçãode detritos orgânicos no solo proporciona um suprimento frescode nitrato para as raizes das plantas.
Como os competidores consomem recursos compartilhados,as interações entre os consumidores e seus recursos influenciamas relações de competição. Isto é especialmente evidente quandoconsideramos as relações consumidor-recurso no contexto maisamplo de todo o sistema ecológico, incluindo as respostas evo-lutivas entre suas espécies constituintes. Por exemplo, algunsrecursos renováveis se originam fora do sistema, além da influ-ência dos consumidores. A luz do Sol atinge a superfície da Ter-ra, a despeito de as plantas a "consumirem", e a precipitaçãolocal é amplamente independente da assimilação de água pelasplantas. Assim, os consumidores reduzem a disponibilidade ime-diata de tais recursos para outros, mas não seu suprimento. Maisainda, tais recursos não respondem de qualquer forma ao con-sumo.
Um segundo tipo de recurso renovável é gerado dentro dosistema ecológico. Os consumidores diretamente deplecionama abundância destes recursos. A maioria das interações preda-
dor-presa, herbívoro-planta e parasita-hospedeiro envolve os re-cursos renováveis, porque o suprimento de presas, plantas e hos-pedeiros é constantemente regenerado. Contudo, ao reduzir aspopulações e seus recursos, os consumidores potencialmente re-duzem a taxa de renovação de seu próprio suprimento alimentar.
Alguns recursos renováveis regenerados dentro do ecossiste-ma estão conectados somente indiretamente a seus consumidores,seja através de outras conexões numa cadeia alimentar ou deprocessos abióticos. No ciclo do nitrogênio de uma floresta, porexemplo, as plantas assimilam nitratos de solo. Os herbívoros eos detritívoros consomem plantas ou restos de plantas, devol-vendo grandes quantidades de compostos nitrogenados orgânicospara o solo. Estes compostos nitrogenados são ainda mais de-compostos por micro-organismos que liberam o nitrogênio comonitrato, uma forma que as plantas podem utilizar. A assimilaçãodo nitrato pelas plantas, contudo, tem pouco efeito direto sobresua renovação pelos detritívoros e micro-organismos. Analoga-mente, o consumo de detritos pelos detritívoros não influenciade imediato a produção das plantas. Claramente, contudo, osdetritívoros e os micro-organismos de fato influenciam a produ-ção das plantas indiretamente através da taxa na qual eles liberamnitrogênio no solo.
Recursos limitantesAo diminuir seus recursos, os consumidores limitam seu própriocrescimento populacional. À medida que a população cresce, suademanda global por recursos cresce também. Quando suas exi-gências aumentam ao ponto de os suprimentos decrescentes deum recurso não poderem mais preencher suas necessidades, otamanho da população se estabiliza, ou mesmo começa a dimi-nuir. Contudo, enquanto os consumidores reduzem tanto os re-cursos renováveis quanto os não renováveis, nem todos os recur-sos limitam as populações de consumidores. Todos os animaisterrestres demandam oxigênio, por exemplo, mas eles não de-plecionam perceptivelmente seu nível na atmosfera antes quealgum outro recurso, como suprimento de comida, limite seucrescimento populacional.
Já houve tempo em que os ecólogos acreditavam que as po-pulações eram limitadas pelo único recurso que era mais escas-so relativo à demanda. Este princípio foi chamado de lei do mí-nimo de Liebig, em homenagem a Justus von Liebig, um quí-mico alemão que articulou a ideia em 1840. De acordo comesta lei, cada população cresce até que o suprimento de algumrecurso - o recurso Iimitante - não mais satisfaça às neces-sidades da população por ele.
O suprimento de um recurso necessário para sustentar o cres-cimento de uma população sobre um dado conjunto de condiçõesé único para cada recurso. Por exemplo, David Tilman descobriuque quando a diatornácea Cyclotella meneghiniana é cultivadacom suprimentos limitados de silicato e fosfato numa cultura delaboratório, o crescimento populacional cessa quando os níveisde fosfato são reduzidos a 0,2 milimolar (mM) ou os níveis desilicato são reduzidos a 0,6 mM. De acordo com a lei de Liebigdo mínimo, qualquer destes recursos que for reduzido a este va-lor limitante regula em primeira instância o crescimento da po-pulação de Cyclotella.
A lei de Liebig se aplica estritamente apenas aos recursos quetêm uma influência independente na população de consumidores.Em muitos casos, contudo, dois ou mais recursos interagem pa-ra determinar a taxa de crescimento da população de um consu-midor, isto é, a taxa de crescimento de uma população num de-terminado nível de um recurso depende do nível de um ou mais
Competição 295
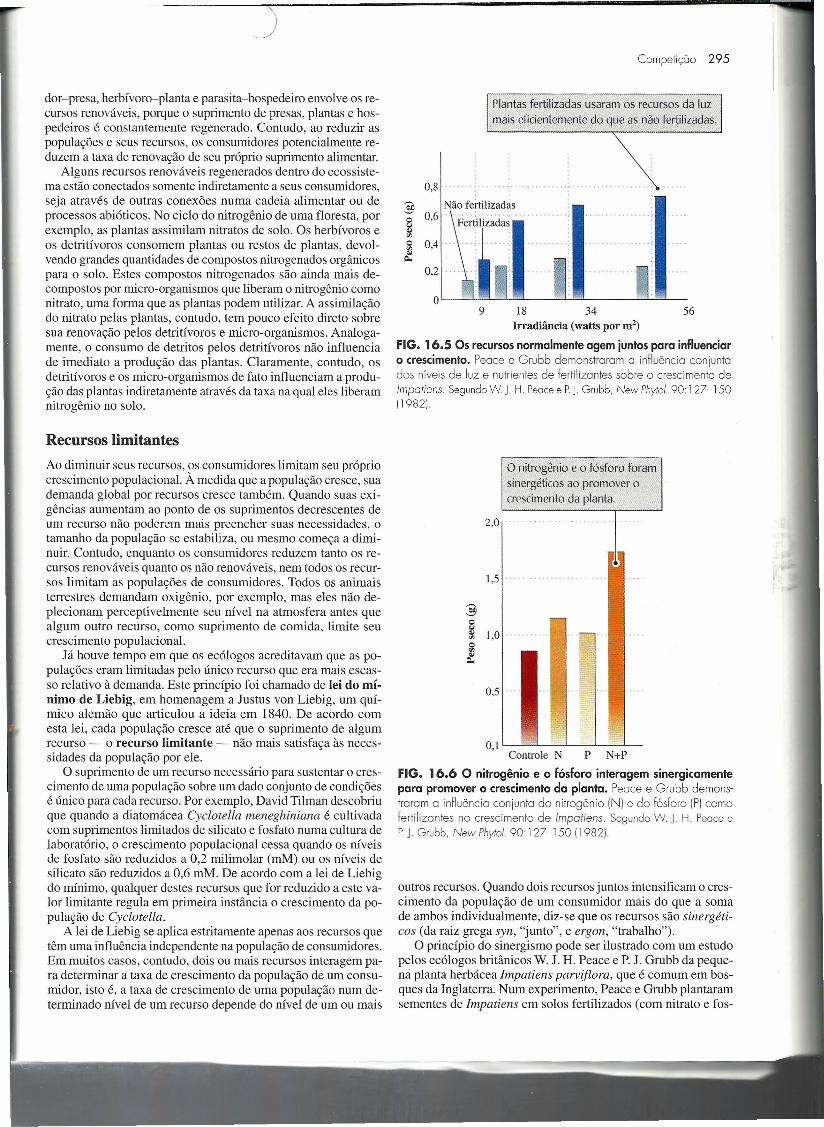
Plantas fertilizadas usaram os recursos da luzmais eficientemente do que as não fertilizadas.
0,8 ......•.....
:§ Não fertilizadasQ 0,6~Q 0,4~
0,2
9 18 34 56Irradiância (watts por m2)
FIG. 16.5 Os recursos normalmente agem juntos para influenciaro crescimento. Peace e Grubb demonstraram a influência conjuntodos níveis de luz e nutrientes de fertilizantes sobre o crescimento de/mpotiens. SegundoW. J H. Peocee PJ Grubb, New Phyto/. 90 127-15011982)
o nitrogênio e o fósforo foramsinergéticos ao promover ocrescimento da planta.
2,0 .
1,5 . r ,
38~ 1,0' .
~
0,5 ...
O, I '-----'-----'-----'----'--Controle N P N+P
FIG. 16.6 O nitrogênio e o fósforo interagem sinergicamentepara promover o crescimento da planta. Peace e Grubb demons-traram a influência conjunto do nitrogênio (NI e do fósforo (PIcomofertilizantes no crescimento de /mpotiens. SegundoW. J H. PeoceePJ Grubb, New Phyto/. 90 127-15011982)
outros recursos. Quando dois recursos juntos intensificam o cres-cimento da população de um consumidor mais do que a somade ambos individualmente, diz-se que os recursos são sinergéti-cos (da raiz grega syn, "junto", e ergon, "trabalho").
O princípio do sinergismo pode ser ilustrado com um estudopelos ecólogos britânicos W. J. H. Peace e P. J. Grubb da peque-na planta herbácea Impatiens parviflora, que é comum em bos-ques da Inglaterra. Num experimento, Peace e Grubb plantaramsementes de Impatiens em solos fertilizados (com nitrato e fos-
296 Competição
fato) e não fertilizados (controle), e então expuseram as plântu-Ias resultantes a diferentes níveis de iluminação por 5 semanasapós a germinação da semente. Uma luz adicionada aumentouo crescimento das plantas fertilizadas mais do que as do contro-le (Fig. 16.5). Assim, a capacidade de Impatiens em usar a luzdepende da disponibilidade de outros recursos. O crescimentoda planta exige tanto o carbono assimilado pela fotossíntese,como fonte de energia e para carboidratos estruturais, quantonitrogênio e fósforo, que são necessários para a síntese de pro-teínas, fosfolipídios e ácidos nucleicos. Nas intensidades de luzmais altas usadas no experimento, o nitrogênio e o fósforo tam-bém mostraram que interagem sinergicamente para aumentar ocrescimento da planta (Fig. 16.6).
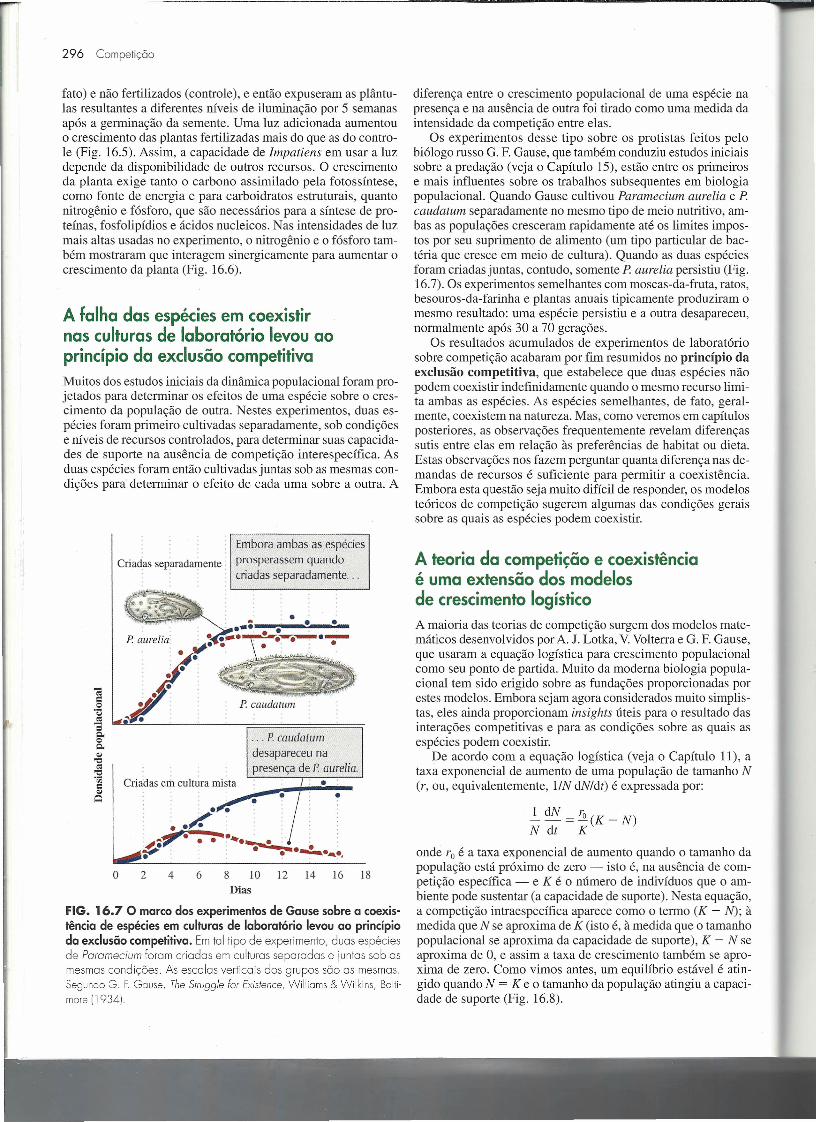
A falha das espécies em coexistirnas culturas de laboratório levou aoprincípio da exclusão competitivaMuitos dos estudos iniciais da dinâmica populacional foram pro-jetados para determinar os efeitos de uma espécie sobre o cres-cimento da população de outra. Nestes experimentos, duas es-pécies foram primeiro cultivadas separadamente, sob condiçõese níveis de recursos controlados, para determinar suas capacida-des de suporte na ausência de competição interespecífica. Asduas espécies foram então cultivadas juntas sob as mesmas con-dições para determinar o efeito de cada uma sobre a outra. A
Embora ambas as espéciesprosperassem quandocriadas separadamente ...
Criadas separadamente.
••• : • • e_P. aure/lia: : : •••• _:~r::;•.•.r.:.__ .-:-..
· : .· '.'•.i'· .
~jY .P. caudatum .
. P caudatum
desapareceu napresença de P aurelio.
•Criadas em cultura mista•
: : /.'.~: ..· . . . .1i".. ......•... ..- -._._. .~.. .~ .ti' . • •••••••.•.•.•,
6 8 10 12 14 16 18o 2 4Dias
FIG. 16.7 O marco dos experimentos de Gause sobre a coexis-tência de espécies em culturas de laboratório levou ao princípioda exclusão competitiva. Em tal tipo de experimento, duas espéciesde Paramecium foram criadas em culturas separadas e iuntas sob asmesmas condições. As escalas verticais dos grupos são as mesmas.Segundo G. F.Gouse, The Stru99/e for Existence, Willioms & vvtlkíns. Bolti-more 11934).
diferença entre o crescimento populacional de uma espécie napresença e na ausência de outra foi tirado como uma medida daintensidade da competição entre elas.
Os experimentos desse tipo sobre os protistas feitos pelobiólogo russo G. F. Gause, que também conduziu estudos iniciaissobre a predação (veja o Capítulo 15), estão entre os primeirose mais influentes sobre os trabalhos subsequentes em biologiapopulacional. Quando Gause cultivou Paramecium aurelia e P.caudatum separadamente no mesmo tipo de meio nutritivo, am-bas as populações cresceram rapidamente até os limites impos-tos por seu suprimento de alimento (um tipo particular de bac-téria que cresce em meio de cultura). Quando as duas espéciesforam criadas juntas, contudo, somente P. aurelia persistiu (Fig.16.7). Os experimentos semelhantes com moscas-da-fruta, ratos,besouros-da-farinha e plantas anuais tipicamente produziram omesmo resultado: uma espécie persistiu e a outra desapareceu,normalmente após 30 a 70 gerações.
Os resultados acumulados de experimentos de laboratóriosobre competição acabaram por fim resumidos no princípio daexclusão competitiva, que estabelece que duas espécies nãopodem coexistir indefinidamente quando o mesmo recurso limi-ta ambas as espécies. As espécies semelhantes, de fato, geral-mente, coexistem na natureza. Mas, como veremos em capítulosposteriores, as observações frequentemente revelam diferençassutis entre elas em relação às preferências de habitat ou dieta.Estas observações nos fazem perguntar quanta diferença nas de-mandas de recursos é suficiente para permitir a coexistência.Embora esta questão seja muito difícil de responder, os modelosteóricos de competição sugerem algumas das condições geraissobre as quais as espécies podem coexistir.
A teoria da competição e coexistênciaé uma extensão dos modelosde crescimento logísticoA maioria das teorias de competição surgem dos modelos mate-máticos desenvolvidos por A. J. Lotka, V. Volterra e G. F. Gause,que usaram a equação logística para crescimento populacionalcomo seu ponto de partida. Muito da moderna biologia popula-cional tem sido erigido sobre as fundações proporcionadas porestes modelos. Embora sejam agora considerados muito simplis-tas, eles ainda proporcionam insights úteis para o resultado dasinterações competitivas e para as condições sobre as quais asespécies podem coexistir.
De acordo com a equação logística (veja o Capítulo 11), ataxa exponencial de aumento de uma população de tamanho N(r, ou, equivalentemente, I/N dN/dt) é expressada por:
.l dN = !l(K - N)N dt K
onde ro é a taxa exponencial de aumento quando o tamanho dapopulação está próximo de zero - isto é, na ausência de com-petição específica - e K é o número de indivíduos que o am-biente pode sustentar (a capacidade de suporte). Nesta equação,a competição intraespecífica aparece como o termo (K - N); àmedida que N se aproxima de K (isto é, à medida que o tamanhopopulacional se aproxima da capacidade de suporte), K - N seaproxima de O, e assim a taxa de crescimento também se apro-xima de zero. Como vimos antes, um equilíbrio estável é atin-gido quando N = K e o tamanho da população atingiu a capaci-dade de suporte (Fig. 16.8).
~ . . - ~ -::-

Uma taxa de crescimento per capitadecrescente expressa a influênciada competição intraespecífica.
Quando N =K, a taxa decrescimento da população é O.
KDensidade populacional (N)
FIG. 16.8 A taxa exponencial de aumento de uma populaçãoque cresce logisticamente é uma função de sua densidade. Mesmoquando a competição introespecillco depleciona r, a populaçãocontinua a aumentar até que seu tamanho (Nj seja igual à capaci-dade de suporte (I<].
Ao modelar a interação entre duas espécies cujas populaçõescrescem logisticamente, distinguimos cada uma das populaçõesusando o subscrito 1 para a espécie 1 e o subscrito 2 para a es-pécie 2. Agora podemos incluir o efeito da competição da espé-cie 2 sobre o crescimento populacional da espécie 1 subtraindoo termo al,2N2 da equação entre parênteses. Assim,
onde ri é a taxa exponencial de aumento da espécie 1 quando otamanho de sua população está próximo de O, N2 é o número deindivíduos da espécie 2 e al,2 é o coeficiente de competição - is-to é, o efeito de um indivíduo da espécie 2 sobre a taxa exponen-cial de aumento da espécie l. Você pode pensar no coeficientede competição al,2 como o grau no qual cada indivíduo da espé-cie 2 usa os recursos da espécie 1 em relação a um indivíduo daespécie l. Assim, al,2N2 representa o efeito da população 2 sobrea taxa de crescimento da população 1 (Fig. 16.9).
Como cada espécie competindo exerce um efeito sobre a ou-tra, a relação mútua entre elas exige duas equações. Uma, apre-sentada acima, incorpora o efeito da espécie 2 sobre a espécie 1.A segunda equação incorpora o efeito da espécie 1 sobre a es-pécie 2 e é semelhante à primeira, mas com os subscritos 1 e 2revertidos.
Se duas espécies devem coexistir, as populações de ambasdevem atingir um estado de equilíbrio com um tamanho maiordo que zero. Isto é, tanto dN/Nldt e dN/N2dt devem igualar-sea zero em alguma combinação de valores positivos de NI e N2•
Da equação anterior, vimos que dN/Nldt = O quando
N2 = K2 - a2•JNJ
O pequeno chapéu nsobre os Ns indica que eles são valores deequilíbrio. Na ausência de competição interespecífica (ai 2 = O),o tamanho da população de equilíbrio N[ é igual a KI, uma me-dida dos recursos disponíveis para a espécie 1. A competição
Competição 297
CHAVE Competição[=::J Intraespecífica[=::J Interespecífica baixa_ Interespecífica alta
Densidade populacional (N I)
FIG. 16.9 A competição interespecífica reduz o nível de equilí-brio de uma população abaixo de sua capacidade de suporte.A taxa exponencial de aumento da espécie 1 em função de suadensidade populacional (Nd é mostrada sob uma competição pu-ramente intraespecífica e sob níveis baixos e altos de competiçãointerespecífica.
interespecífica reduz a capacidade de suporte efetiva do ambien-te para a espécie 1pela quantidade al,2 N 2' Assim, a capacidade desuporte para a espécie 1 é reduzida proporcionalmente ao tama-nho da população e do coeficiente de competição da espécie 2(veja a Fig. 16.9).
Em geral, estas equações nos dizem que a coexistência é maisprovável (N1 e N2 > O) quando a competição interespecífica érelativamente fraca comparada com a competição intraespecífi-ca - isto é, quando os coeficientes de competição al•2 e a2,1 sãomenores do que 1. Em outras palavras, para coexistir, as espéciesdevem se limitar mais do que elas limitam uma à outra. Qualquernúmero de espécies pode coexistir enquanto este critério for aten-dido para todos os pares delas. Os coeficientes de competiçãoprovavelmente são menores do que 1 quando os competidoressão especializados para usar diferentes recursos de forma maiseficiente. Note que o resultado da competição não depende dastaxas de crescimento exponencial das populações quando elassão pequenas.
MAISNA
REDE
Vá para Gráficos Interativos. Use o tutorial interativo so-bre competição para entender melhor a dinâmica entreas espécies.
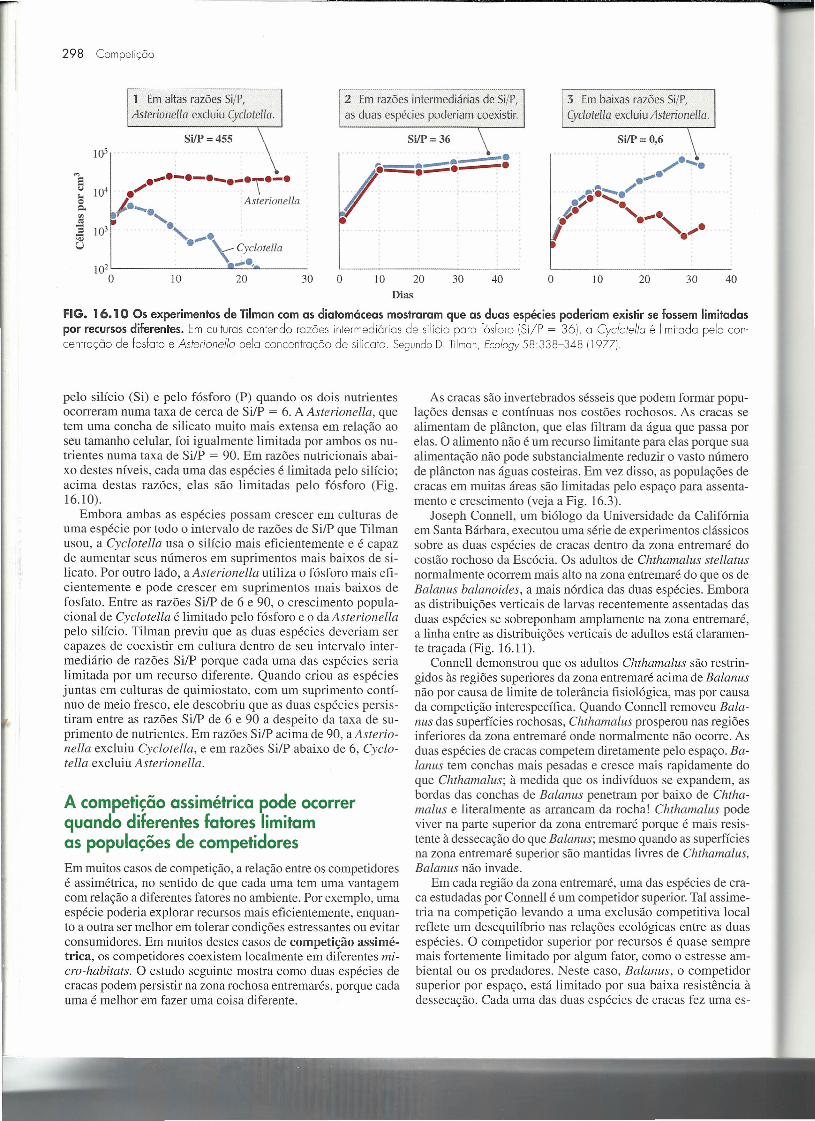
A coexistência sobre múltiplos recursosNos experimentos de Gause sobre a competição com Parame-dum, duas espécies foram forçadas a usar um recurso limitante(um certo tipo de alimento). Quando duas espécies competempor dois recursos limitantes necessários em diferentes quantida-des para cada espécie, certas combinações de abundância derecurso possibilitam que as duas espécies coexistam. David Til-man foi quem primeiro demonstrou este princípio numa série deexperimentos elegantes sobre competição entre duas espécies dediatomáceas do gênero Cyclotella e Asterionella. Ambas as es-pécies demandam silício, para produzir suas conchas externasvítreas, e fósforo, para sintetizar os fosfolipídios da membrana,DNA e outras moléculas, mas suas demandas relativas por estesrecursos diferem. Os experimentos iniciais demonstraram que ocrescimento populacional de Cyclotella foi igualmente limitado
----~~4!'I.- .,- _ .~...,. .,. ~.. -- --- -- - ---
298 Competição
1 Emaltas razões Si/P, 2 Emrazões intermediárias de SijP, 3 Embaixas razões Si/P,Asterionella excluiuCyclotella. as duas espécies poderiam coexistir. Cyclotella excluiuAsterionella.
SiIP = 455 SiIP = 36 SiIP = 0,6105 _. / •.....•--- •.., ·.,...·-·-·-·-·r·-· /=;_. •••••5 •104 ./ ,'"'--.. /10 ,..,•......•
o I. Asterionella .~.e, -. ri· ' •.,....,Vl '.~:2 10' ' .......'c ;.",. •U Cyclotella
102..-.,
O 10 20 30 O 10 20 30 40 O 10 20 30 40Dias
FIG. 16.10 Os experimentos de Tilman com as diatomáceas mostraram que as duas espécies poderiam existir se fossem limitadaspor recursos diferentes. Em culturas contendo razões intermediárias de silício para fósforo (Si/P = 36), a Cye/otello é limitada pela con-centração de fosfoto e Asterionello pela concentração de silicato. SegundoD. Tilman,Ec%gy 58338-348119771.
pelo silício (Si) e pelo fósforo (P) quando os dois nutrientesocorreram numa taxa de cerca de SilP = 6. A Asterionella, quetem uma concha de silicato muito mais extensa em relação aoseu tamanho celular, foi igualmente limitada por ambos os nu-trientes numa taxa de Si/P = 90. Em razões nutricionais abai-xo destes níveis, cada uma das espécies é limitada pelo silício;acima destas razões, elas são limitadas pelo fósforo (Fig.16.10).
Embora ambas as espécies possam crescer em culturas deuma espécie por todo o intervalo de razões de Si/P que Tilmanusou, a Cyclotella usa o silício mais eficientemente e é capazde aumentar seus números em suprimentos mais baixos de si-licato. Por outro lado, a Asterionella utiliza o fósforo mais efi-cientemente e pode crescer em suprimentos mais baixos defosfato. Entre as razões Si/P de 6 e 90, o crescimento popula-cional de Cyclotella é limitado pelo fósforo e o da Asterionellapelo silício. Tilman previu que as duas espécies deveriam sercapazes de coexistir em cultura dentro de seu intervalo inter-mediário de razões Si/P porque cada uma das espécies serialimitada por um recurso diferente. Quando criou as espéciesjuntas em culturas de quimiostato, com um suprimento contí-nuo de meio fresco, ele descobriu que as duas espécies persis-tiram entre as razões Si/P de 6 e 90 a despeito da taxa de su-primento de nutrientes. Em razões Si/P acima de 90, a Asterio-nella excluiu Cyclotella, e em razões Si/P abaixo de 6, Cyclo-tella excluiu Asterionella.
A competição assimétrica pode ocorrerquando diferentes fatores limitamas populações de competidoresEm muitos casos de competição, a relação entre os competidoresé assimétrica, no sentido de que cada uma tem uma vantagemcom relação a diferentes fatores no ambiente. Por exemplo, umaespécie poderia explorar recursos mais eficientemente, enquan-to a outra ser melhor em tolerar condições estressantes ou evitarconsumidores. Em muitos destes casos de competição assimé-trica, os competidores coexistem localmente em diferentes mi-cro-habitats. O estudo seguinte mostra como duas espécies decracas podem persistir na zona rochosa entremarés, porque cadauma é melhor em fazer uma coisa diferente.
As cracas são invertebrados sésseis que podem formar popu-lações densas e contínuas nos costões rochosos. As cracas sealimentam de plâncton, que elas filtram da água que passa porelas. O alimento não é um recurso limitante para elas porque suaalimentação não pode substancialmente reduzir o vasto númerode plâncton nas águas costeiras. Em vez disso, as populações decracas em muitas áreas são limitadas pelo espaço para assenta-mento e crescimento (veja a Fig. 16.3).
Joseph Connell, um biólogo da Universidade da Califórniaem Santa Bárbara, executou uma série de experimentos clássicossobre as duas espécies de cracas dentro da zona entremaré docostão rochoso da Escócia. Os adultos de Chthamalus stellatusnormalmente ocorrem mais alto na zona entremaré do que os deBalanus balanoides, a mais nórdica das duas espécies. Emboraas distribuições verticais de larvas recentemente assentadas dasduas espécies se sobreponham amplamente na zona entremaré,a linha entre as distribuições verticais de adultos está claramen-te traçada (Fig. 16.11).
Connell demonstrou que os adultos Chthamalus são restrin-gidos às regiões superiores da zona entremaré acima de Balanusnão por causa de limite de tolerância fisiológica, mas por causada competição interespecífica. Quando Connell removeu Bala-nus das superfícies rochosas, Chthamalus prosperou nas regiõesinferiores da zona entremaré onde normalmente não ocorre. Asduas espécies de cracas competem diretamente pelo espaço. Ba-lanus tem conchas mais pesadas e cresce mais rapidamente doque Chthamalus; à medida que os indivíduos se expandem, asbordas das conchas de Balanus penetram por baixo de Chtha-malus e literalmente as arrancam da rocha! Chthamalus podeviver na parte superior da zona entremaré porque é mais resis-tente à dessecação do que Balanus; mesmo quando as superfíciesna zona entremaré superior são mantidas livres de Chthamalus,Balanus não invade.
Em cada região da zona entremaré, uma das espécies de era-ca estudadas por Connell é um competidor superior. Tal assime-tria na competição levando a uma exclusão competitiva localreflete um desequilíbrio nas relações ecológicas entre as duasespécies. O competidor superior por recursos é quase sempremais fortemente limitado por algum fator, como o estresse am-biental ou os predadores. Neste caso, Balanus, o competidorsuperior por espaço, está limitado por sua baixa resistência àdessecação. Cada uma das duas espécies de cracas fez uma es-

~~--- ~ ~A dessecação limita Balanus nesta zona, +possibilitando a Chthamalus prosperar. t ~'-'--~"'--' ••..•...."-Balanus limita Chthamalus nesta zona, 1dominando-a por ocupação espacial.
Marés maisaltas Zona entrernaré superior
Zona entrernaré inferior
Marés maisbaixas
Competiçõo 299
FIG. 16.11 As duas espécies de cracas apresentam competição assimétrica. As distribuições verticais das espécies de cracas Ba/anusba/anoides e Chthama/us stel/atus na zona entremaré diferem porque Ba/anus é um competidor superior por espaço, mas nõo pode suportaro estressedo dessecação tão bem quanto Chthamalus. SegundoJ. H. Connell, Ecology 42:720-723 119611e Eco/. Monogr. 3161-104119611
colha diferente entre a tolerância ao estresse e a capacidade com-petitiva.
Os resultados de experimentos de campo sobre as espéciesque coexistem na natureza sugerem que a competição assimé-trica é comum. Estes experimentos são geralmente conduzidoscom indivíduos de cada espécie que crescem na presença e naausência do outro. A competição interespecífica é avaliada pordiferenças no crescimento dos indivíduos, no sucesso reprodu-tivo, na densidade populacional ou em outras medidas relacio-nadas com o crescimento populacional. De acordo com um le-vantamento de interações entre 98 pares de espécies, nenhumadas espécies foi afetada negativamente pela outra em 44 dos ca-sos (sem competição), ambas as espécies foram afetadas emgraus semelhantes em 21 casos (competição recíproca) e somen-te uma espécie foi afetada em 33 das interações (competiçãoassimétrica). A grande proporção de casos na última categoriaindica que a competição assimétrica é mais comum do que a re-cíproca e que as espécies normalmente coexistem na naturezaporque são limitadas por diferentes fatores.
.NJ:Ãl,# MÓDULO DE ANÁLISE DE DADOS A Assimetria na Com-I -y\ petição. Use os dados para cracas e algas para explorarDE DADOS os efeitos das condicões ambientais do resultado da com-petição entre as duas espécies. Você encontrará este módulo emhttp://www.whfreeman.com/ricklefs6e.
A produtividade do habitat pode influenciara competição entre as espécies de plantasAs plantas podem competir tanto por água quanto por nutrientesno solo e luz do Sol acima do solo; todos estes fatores aumentama produtividade da vegetação. Diversos ecólogos sugeriram queo resultado da competição entre plantas depende dos níveis denutrientes nos solos, porque os níveis de nutrientes controlam ocrescimento dos brotos das plantas e sua capacidade em som-brear os competidores. De acordo com uma hipótese, proposta
pelos ecólogos de plantas P. J. Grubb e David Tilman, as plantascompetem mais intensamente quando os nutrientes minerais sãomenos abundantes no solo. Altos níveis de nutrientes têm menosprobabilidade de limitar as populações de plantas, e portanto acompetição interespecífica deve ser mais fraca. O ponto de vis-ta oposto, defendido por J. P. Grime e Paul Keddy, sugere que acompetição é menos intensa quando a água e os nutrientes mi-nerais são menos abundantes. A razão, de acordo com Grime eKeddy, é que a competição por luz é mais importante do que acompetição por nutrientes no solo. A ausência de água e nutrien-tes deveria limitar as populações até o ponto em que cada plan-ta estivesse suficientemente distante das outras e não competis-se por luz. A diferença entre as hipóteses de Grubb- Tilman eGrime-Keddy jaz na relativa importância colocada sobre a com-petição por recursos abaixo do solo e acima do solo - isto é,por nutrientes e luz, respectivamente.
Estas duas hipóteses podem ser testadas por experimentos decompetição em ambientes de alta e baixa produtividade, mas osresultados normalmente dependem da espécie usada e do proje-to do experimento. Por exemplo, um estudo experimental, con-duzido por S. D. Wilson e David Tilman num habitat de prada-ria mésico em Minnesota com três espécies de gramíneas depradaria, encontrou pouca correlação entre os níveis de nutrien-tes e competição. Os pesquisadores estabeleceram áreas de pro-dutividade baixa, média e alta, adicionando como fertilizador onitrato de amônia ao solo. A biomassa acima do solo variou deduas a três vezes entre as áreas, mas a intensidade de competiçãonão variou significativamente entre as áreas. Wilson e Tilmansugeriram que as plantas competiram abaixo do solo principal-mente por nutrientes em áreas de poucos nutrientes e acima dosolo principalmente por luz em áreas ricas em nutrientes, resul-tando em uma intensa competição em todos os níveis de nutrien-tes.
Em outro estudo, Nancy Emery e seus colaboradores PatrickEwanchuk e Mark Bertness manipularam os níveis de nutrientesao longo de um gradiente de salinidade e saturação de solo (am-bas condições estressantes para plantas terrestres) num alagadosalobro em Rhode Island. As espécies de plantas estavam forte-
-
":,. ( '1 ~ "" • ~~~~ - '-:a:1_"~~ . ~. -; " 1111')" .
~
~-
300 Competição
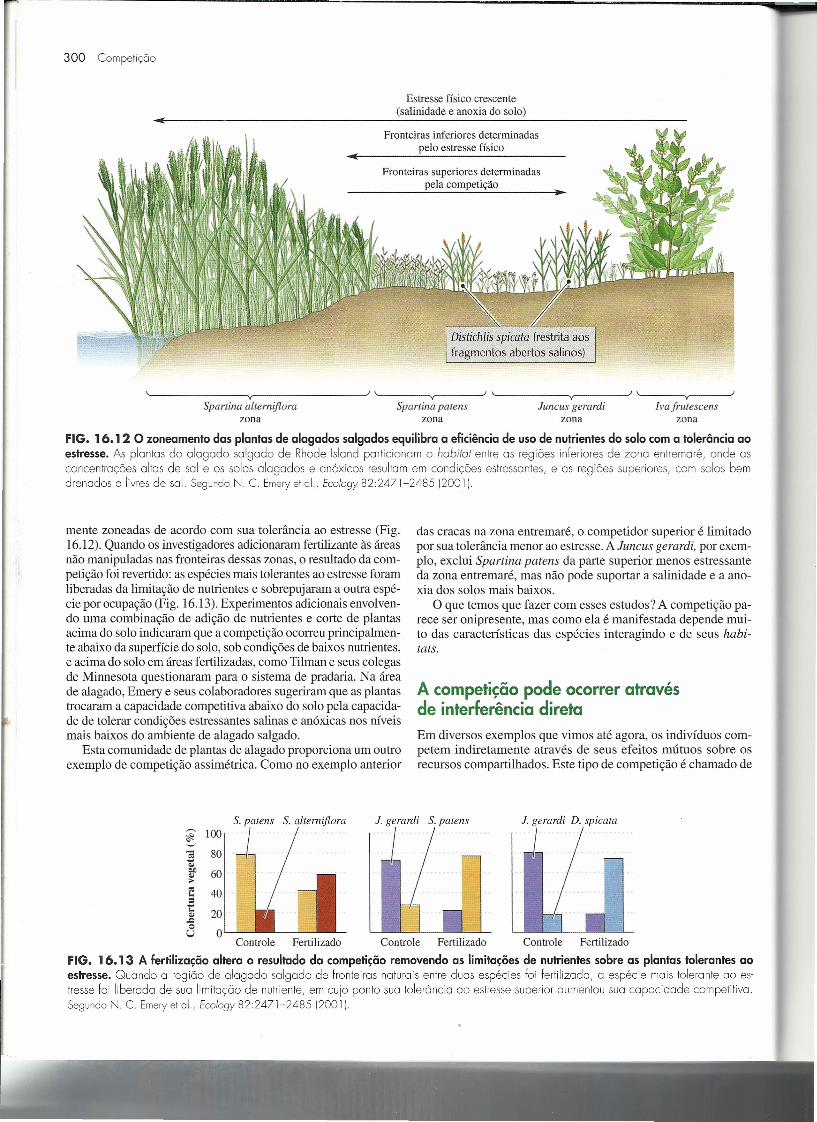
Estresse físico crescente(salinidade e anoxia do solo)
Fronteiras inferiores determinadaspelo estresse físico
Fronteiras superiores determinadaspela competição
Distichlis spicata (restrita aosfragmentos abertos salinos)
~ ~y~ --JJ'~ -Vy ~J '~----------~y __ ----~J'~---- __ VY ~
Spartina altemiflora Spartina patens Juncus gerardi Iva frutescenszona zona zona zona
FIG. 16.12 O zoneamento das plantas de alagados salgados equilibra a eficiência de uso de nutrientesdo solo com a tolerância aoestresse. As plantas do alagado salgado de Rhode lslcnd particionam o habitat entre as regiões inferiores de zona entremaré, onde asconcentrações altas de sal e os solos alagados e anóxicos resultam em condições estressantes, e as regiões superiores, com solos bemdrenados e livres de sal. SegundoN. C. Emeryet 0[., Ec%gy 82:2471-2485 (2001 I.
mente zoneadas de acordo com sua tolerância ao estresse (Fig.16.12). Quando os investigadores adicionaram fertilizante às áreasnão manipuladas nas fronteiras dessas zonas, o resultado da com-petição foi revertido: as espécies mais tolerantes ao estresse foramliberadas da limitação de nutrientes e sobrepujaram a outra espé-cie por ocupação (Fig. 16.13). Experimentos adicionais envolven-do uma combinação de adição de nutrientes e corte de plantasacima do solo indicaram que a competição ocorreu principalmen-te abaixo da superfície do solo, sob condições de baixos nutrientes,e acima do solo em áreas fertilizadas, como Tilman e seus colegasde Minnesota questionaram para o sistema de pradaria. Na áreade alagado, Emery e seus colaboradores sugeriram que as plantastrocaram a capacidade competitiva abaixo do solo pela capacida-de de tolerar condições estressantes salinas e anóxicas nos níveismais baixos do ambiente de alagado salgado.
Esta comunidade de plantas de alagado proporciona um outroexemplo de competição assimétrica. Como no exemplo anterior
das cracas na zona entremaré, o competidor superior é limitadopor sua tolerância menor ao estresse. A Juncus gerardi, por exem-plo, exclui Spartina patens da parte superior menos estressanteda zona entremaré, mas não pode suportar a salinidade e a ano-xia dos solos mais baixos.
O que temos que fazer com esses estudos? A competição pa-rece ser onipresente, mas como ela é manifestada depende mui-to das características das espécies interagindo e de seus habi-tats.
A competição pode ocorrer atravésde interferência diretaEm diversos exemplos que vimos até agora, os indivíduos com-petem indiretamente através de seus efeitos mútuos sobre osrecursos compartilhados. Este tipo de competição é chamado de
S. alterniflora J. gerardi D. spicata
20O
Controle Fertilizado Controle Fertilizado Controle Fertilizado
FIG. 16.13 A fertilização altera o resultado da competição removendo as limitações de nutrientes sobre as plantas tolerantes aoestresse. Quando a região de alagado salgado de fronteiras naturais entre duas espécies foi fertilizada, a espécie mais tolerante ao es-tresse foi liberada de sua limitação de nutriente, em cujo ponto sua tolerância ao estressesuperior aumentou sua capacidade competitiva.SegundoN. C. Emeryet 01, Ec%gy 822471-2485 (2001 I.

-
.. '\t' .'" ".,," '~'i\!~."" ----
;,' ~ri1"~:~_J'lr":~~1'
" }i!IIii;~~::ldIi.~~~tJ" _
competição explorativa. Menos frequentemente, os competi-dores interagem diretamente através da defesa agressiva dos re-cursos. Este comportamento é denominado de competição deinterferência, embora a distinção entre as duas seja às vezesdifícil. Os beija-flores perseguem outros beija-flores, para nãomencionar abelhas e mariposas, para mantê-los longe dos arbus-tos florescentes. As esponjas incrustadoras usam venenos quí-micos para sobrepujar outras espécies de esponja à medida quese expandem para preencher o espaço nas superfícies rochosas.Muitos arbustos liberam químicos tóxicos no solo para inibir ocrescimento de competidores. Até mesmo as bactérias usam equi-pamento químico de guerra umas com as outras para fazer abalança pender para o seu lado em suas interações competiti-vas.
A competição de interferência é normalmente evidente emmanipulações experimentais de espécies animais competidoras.Por exemplo, duas espécies de rato-do-campo (pequeno roedorsemelhante a um rato do gênero Microtus) estão ambas presentesem algumas áreas das Montanhas Rochosas na América do Nor-te: no oeste de Montana, o rato-de-pradaria (M. pennsylvanicus)normalmente vive em habitats úmidos circundando pequenoslagos e cursos de água, enquanto o rato-de-montanha (M. mon-tanus) fica restrito aos habitats secos. Os ecólogos acreditavamque esta divisão espacial resultava da competição entre as duasespécies, mas a natureza da interação não era conhecida. Con-tudo, quando os ratos-de-pradaria foram experimentalmente cap-turados e removidos de uma área de habitat úmido, os ratos-de-montanha imediatamente começaram a se mover para lá vindodas vizinhanças de habitats secos. Este resultado sugeriu que osratos-de-pradaria excluem os ratos-de-montanha das áreas maisúmidas por encontros agressivos e diretos - um caso de terri-torialidade interespecífica. O interessante é que, quando os ra-tos-de-campo de montanha foram capturados e removidos de umhabitat seco que eles normalmente ocupavam exclusivamente,os ratos-de-pradaria começaram a se mover para lá. Assim, cadaespécie é comportamentalmente dominante no seu habitat pre-ferido, ilustrando o princípio da vantagem de campo domésticonos roedores.
AlelopatiaA competição química, ou alelopatia, tem sido relatada maisfrequentemente em plantas terrestres, nas quais tais interaçõespodem assumir diversas formas. Na maioria dos casos, uma subs-tância tóxica causa danos (-patia) a outros (alelo-) indivíduosdiretamente. Por exemplo, a nogueira-negra (Juglans nigra) pro-duz juglone, um composto orgânico aromático que inibe certasenzimas em outras plantas. Em consequência, poucas espéciesde plantas são capazes de germinar e se estabelecerem sob essasárvores. Um mecanismo um tanto quanto diferente de ação foisugerido para explicar os abundantes óleos nas folhas e na cascado tronco das árvores do eucalipto da Austrália - isto é, queelas promovem frequentes incêndios na serapilheira, que mataas plântulas dos competidores (Fig. 16.14).
Nos habitats de arbusto no sul da Califómia, diversas espéciesde sálvia (gênero Salvia) produzem terpenos voláteis, uma clas-se de compostos orgânicos que inclui a cânfora e dá aos alimen-tos temperados com a salva parte de seu sabor distinto. Os ter-penos inibem o crescimento de outras vegetações no laboratório,e assim os investigadores propõem uma função alelopática paraestes compostos na natureza. Aglomerações de Salvia arbustivasão normalmente circundadas por zonas abertas separando a sál-via das áreas de gramíneas circundantes (Fig. 16.15). Quando
Competição 301
FIG. 16.14 Algumas plantas competem por meios químicos. Asfolhas e a casca das árvores de eucalipto da Austrália têm um altoconteúdo de óleo, que promove incêndios e mata as plôntulas depotenciais competidores, mas deixa o eucalipto intacto. Fotografia deR. E. Ricklefs.
observada por longos períodos, a Salvia pode ser vista se expan-dindo na direção de áreas gramadas.
A despeito de a Sal via liberar compostos inibidores, sua in-teração com os competidores potenciais é mais complicada. Emexperimentos subsequentes, os investigadores cercaram partesde áreas abertas envolvendo os fragmentos de Salvia para excluirherbívoros como coelhos e marmotas. Sem herbívoros presentes,outras plantas se estabeleceriam logo na borda dos fragmentosde sálvia. Contagens de fezes nas zonas abertas mostraram queos coelhos raramente se aventuram mais do que um metro paralonge do paraíso seguro sob a cobertura dos arbustos de Salvia.Estas observações implicam que os coelhos devem estar se ali-mentando dos competidores mais nutritivos da sálvia, mas nãoda sálvia propriamente dita. A observação de que as zonas aber-tas também circundam fragmentos de outras espécies de arbustos,tais como Baccharis, que não produz químicos voláteis tóxicos,sugere que os herbívoros medeiam a competição entre as espé-cies de plantas. Poder-se-ia considerar as zonas abertas como oresultado de um tipo de alelopatia no qual as plantas usam osherbívoros, em vez de químicos tóxicos, para travar suas batalhascom os competidores.
A competição de interferência pode ser um fator importanteno sucesso de espécies invasoras. Por exemplo, as árvores depau-ferro da Austrália (Casuarina equisetifolia) foram introdu-zidas em muitas regiões tropicais e subtropicais do mundo, in-cluindo a Flórida e as Ilhas Havaianas, onde elas rapidamenteinvadiram e excluíram outra vegetação. A quase completa au-sência de germinação ou estabelecimento de vegetação nativanos solos cobertos pelas folhas de Casuarina aciculada sugereum efeito alelopático (Fig. 16.16).

302 Competição
(a) (b)
FIGo 16015 Algumas plantas competem a distância produzindo toxinas aéreas ou abrigando consumidores. (o) A zona aberto nofronteira de um aglomerado de sálvia inclui uma faixa de 2 m de largura sem plantas (A até B) e uma área mais amplo de gramíneas ini-bidos (B até C) sem carvalhos e gramíneas, que são encontrados com outras espécies à direita de C na área não afetado. (b) Uma vistoaérea mostra a sálvia e o arbusto do Colilórn.o invadindo um campo anual no Vale de Santa Ynez da Califórnia. Cortesia de C. H. Muller,de C. H. Muller, Buli. Torrey Bot. C/ub 93:332-351 (1966)
munidades biológicas influenciando o resultado das interaçõescompetitivas entre as espécies de presas.
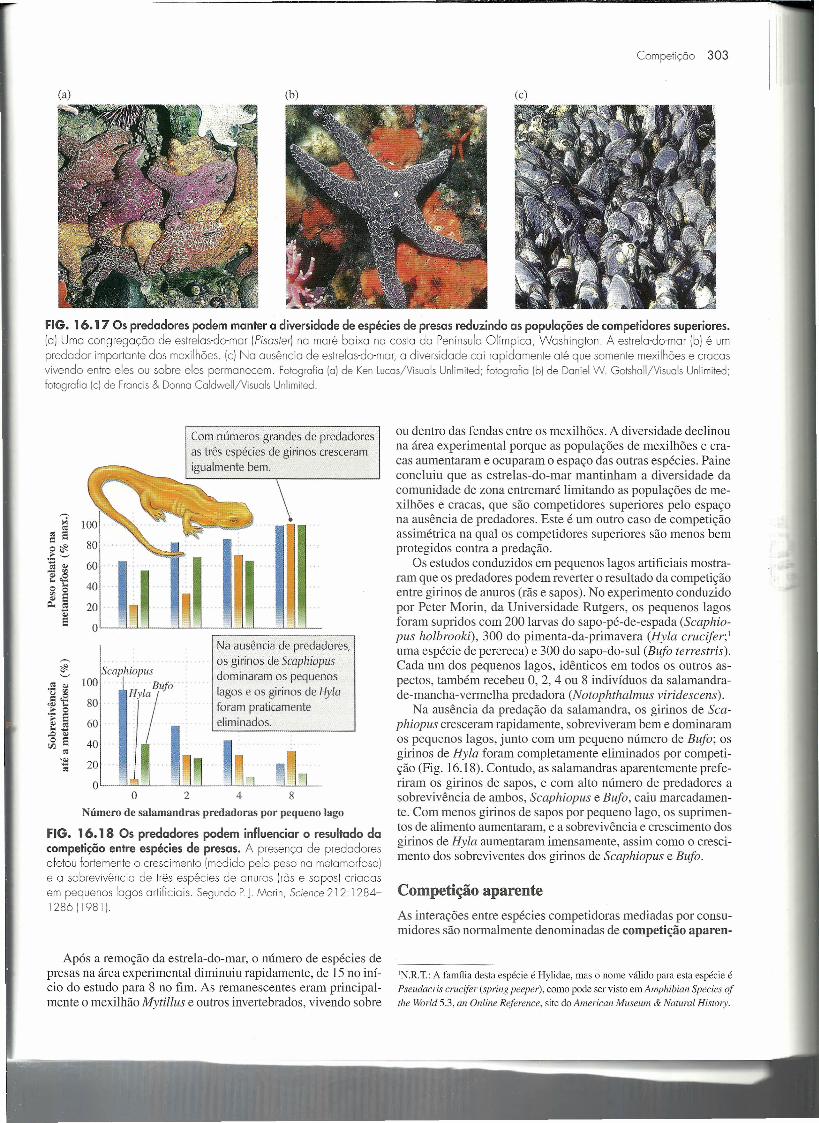
O ecólogo Robert Paine, da Universidade de Washington, foium dos primeiros investigadores a demonstrar isso experimental-mente. Sobre um costão rochoso exposto de Washington, a zonaentremaré abriga diversas espécies de moluscos, cracas-de-pesco-ço-de-ganso, mexilhões, lapas e quítons (um tipo de molusco pas-tador). Todos estes animais são predados pela estrela-do-mar Pi-saster (Fig. 16.17). Paine removeu as estrelas-do-mar de uma áreade rocha de 8 m de largura por 2 m de altura; uma área adjacentede tamanho semelhante foi deixada intacta como controle.
FIGo 16016 A serapilheira embaixo de pau-ferro introduzido (Casuarina equisefifo!ia) su-prime a germinação e o crescimento das es-pécies nativas. Este bosque foi fotografado emKauai, nos Ilhas Havaianas, onde a espécie éinvasora. Cortesia de Eric Guinther.
Os consumidores podem influenciaro resultado da competiçãoCharles Darwin foi o naturalista de quintessência. Entre suasmuitas observações seminais, ele notou que a pastagem podemanter uma alta diversidade de plantas nos campos. Na ausênciade pastadores, os competidores dominantes crescem rapidamen-te e excluem as outras espécies. Resultados semelhantes têmsido obtidos em experimentos em comunidades algais marinhaspastadas por lapas, caracóis e ouriços. Estes estudos indicam quea predação tem uma mão forte na formação da estrutura das co-
(a) (b)
Com números grandes de predadoresas três espécies de girinos cresceramigualmente bem.
80 .
604020O~~~~~======~~~
Scaphiopus100 Bufo
Hyla
Na ausência de predadores,os girinos de Scaphiopusdominaram os pequenoslagos e os girinos de Hylaforam praticamenteeliminados.
80
o 2 4
604020
OL-~~~ __~~ __~~-L __ -L~L-
8Número de salamandras predadoras por pequeno lago
FIG. 16.18 Os predadores podem influenciar o resultado dacompetição entre espécies de presas. A presença de predadoresafetou fortemente o crescimento (medido pelo peso na metamorfose)e a sobrevivência de três espécies de anuros (rãs e sapos) criadasem pequenos lagos artificiais. Segundo P.J Morin, Science 2121284-1286 (1981)
Após a remoção da estrela-do-mar, o número de espécies depresas na área experimental diminuiu rapidamente, de 15 no iní-cio do estudo para 8 no fim. As remanescentes eram principal-mente o mexilhão Mytillus e outros invertebrados, vivendo sobre
Competição 303
FIG. 16.1 7 Os predadores podem manter a diversidade de espécies de presas reduzindo as populações de competidores superiores.(a) Uma congregação de estrelas-do-mar (Pisoster) na maré baixo na costa do Península Olímpica, Washington. A estrela-da-mar (b) é umpredador importante dos mexilhões. (c) Na ausência de estrelas-da-mar, a diversidade cai rapidamente até que somente mexilhões e cracasvivendo entre eles ou sobre eles permanecem. Fotogrofia (a) de Ken luccs/Visuols Unlimited; fotografia [b] de Daniel W. GotshalljVisuals Unlimited;fotografia (e) de Froncis & Donno ColdwelljVisuols Unlimited.
(c)
ou dentro das fendas entre os mexilhões. A diversidade declinouna área experimental porque as populações de mexilhões e era-cas aumentaram e ocuparam o espaço das outras espécies. Paineconcluiu que as estrelas-do-mar mantinham a diversidade dacomunidade de zona entremaré limitando as populações de me-xilhões e cracas, que são competidores superiores pelo espaçona ausência de predadores. Este é um outro caso de competiçãoassimétrica na qual os competidores superiores são menos bemprotegidos contra a predação.
Os estudos conduzidos em pequenos lagos artificiais mostra-ram que os predadores podem reverter o resultado da competiçãoentre girinos de anuros (rãs e sapos). No experimento conduzidopor Peter Morin, da Universidade Rutgers, os pequenos lagosforam supridos com 200 larvas do sapo-pé-de-espada (Scaphio-pus holbrooki), 300 do pimenta-da-primavera (Hyla crucifer; 1
uma espécie de perereca) e 300 do sapo-da-sul (Bufo terrestris).Cada um dos pequenos lagos, idênticos em todos os outros as-pectos, também recebeu O, 2, 4 ou 8 indivíduos da salamandra-de-mancha-vermelha predadora (Notophthalmus viridescens).
Na ausência da predação da salamandra, os girinos de Sca-phiopus cresceram rapidamente, sobreviveram bem e dominaramos pequenos lagos, junto com um pequeno número de Bufo; osgirinos de Hyla foram completamente eliminados por competi-ção (Fig. 16.18). Contudo, as salamandras aparentemente prefe-riram os girinos de sapos, e com alto número de predadores asobrevivência de ambos, Scaphiopus e Bufo, caiu marcadamen-te. Com menos girinos de sapos por pequeno lago, os suprimen-tos de alimento aumentaram, e a sobrevivência e crescimento dosgirinos de Hyla aumentaram imensamente, assim como o cresci-mento dos sobreviventes dos girinos de Scaphiopus e Bufo.
Competição aparenteAs interações entre espécies competidoras mediadas por consu-midores são normalmente denominadas de competição aparen-
'N.R.T.: A família desta espécie é Hylidae, maso nome válido para esta espécie éPseudacris crucifer (spring peeper), como pode ser visto em Amphibian Species ofthe World 5.3, an Online Reference, site do American Museum & Natural History.
pl..-s-.
304 Competição
te - aparente porque o efeito depressor de uma espécie de com-petidor sobre a outra se assemelha à competição de exploraçãoou de interferência mas representa a ação de um mecanismo di-ferente. Vimos a competição aparente nas interações entre a sál-via e as gramíneas e entre os mexilhões e outros organismosrochosos da entremaré: em ambos os casos, os consumidoresdeterminam o equilíbrio competitivo. O resultado da competiçãodepende menos da capacidade de competidores em utilizar oalimento ou outros recursos eficientemente do que de sua habi-lidade para evitar ou tolerar seus próprios consumidores. Nocaso da interação sálvia-grarnínea, os arbustos proporcionamuma cobertura protetora para os herbívoros que preferem comeras gramíneas, mais nutritivas.
Os patógenos podem representar um papel semelhante aodesequilibrar a balança entre os competidores. Certamente, àmedida que os europeus colonizaram outros continentes, parti-cularmente a América do Norte e do Sul, trouxeram doenças comeles, como o sarampo, que suas próprias populações podiam maisou menos tolerar, mas que foram devastadoras para os america-nos nativos. Muitas plantas e animais invasores podem dever seusucesso a mecanismos semelhantes. Por exemplo, a mostarda-do-alho eurasiana (Alliaria petiolata) tem se espalhado na Amé-rica do Norte, onde resiste à herbivoria pelos insetos e cervosamericanos. Por causa de sua resistência aos consumidores quese alimentam da vegetação nativa, a mostarda-da-alho provavel-mente ganha sua vantagem através de uma competição aparente,em vez de qualquer capacidade que possa ter para utilizar os re-cursos de forma eficiente.
~ECÓLOGOSEM CAMPO
A competição aparente entre corais e algasmediada pelos micróbios. A competição apa-rente mediada por consumidores ilustra a
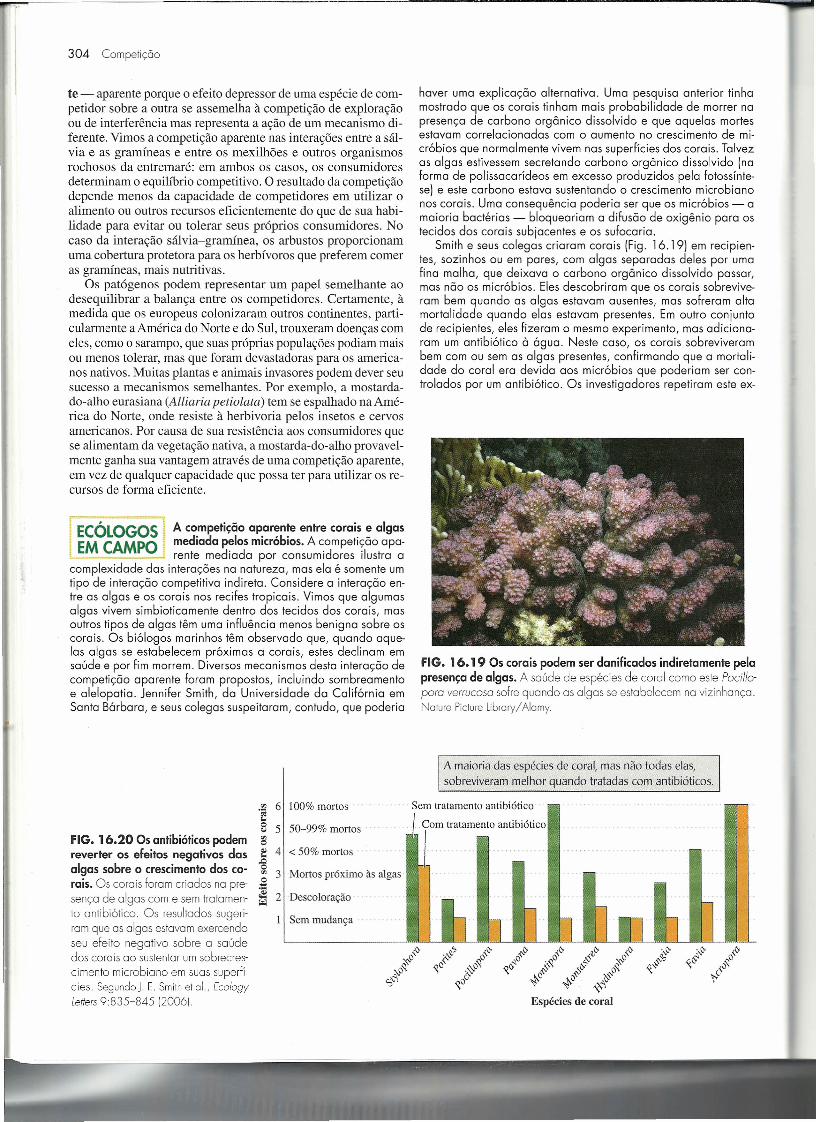
complexidade das interações na natureza, mas ela é somente umtipo de interação competitiva indireta. Considere a interação en-tre as algas e os corais nos recifes tropicais. Vimos que algumasalgas vivem simbioticamente dentro dos tecidos dos corais, masoutros tipos de algas têm uma influência menos benigna sobre oscorais. Os biólogos marinhos têm observado que, quando aque-las algas se estabelecem próximas a corais, estes declinam emsaúde e por fim morrem. Diversos mecanismos desta interação decompetição aparente foram propostos, incluindo sombreamentoe alelopatia. Jennifer Smith, da Universidade da Califórnia emSanta Bárbara, e seus colegas suspeitaram, contudo, que poderia
6 100% mortos
5 50-99% mortos
4 < 50% mortos
3 Mortos próximo às algas
2 Descoloração
Sem mudança
CJl·e8
FIGo 16020 Os antibióticos podem ~reverter os efeitos negativos das ~algas sobre o crescimento dos co- ~rais. Os corais foram criados na pre- S
d I ~sença e a gas com e sem tratamen- r.<to antibiótico. Os resultados sugeri-ram que as algas estavam exercendoseu efeito negativo sobre a saúdedos corais ao sustentarum sobrecres-cimento microbiano em suas superfí-cies. Segundo], E. Smithet ol., Eco/ogyLellers 9835-845 (20061
haver uma explicação alternativa. Uma pesquisa anterior tinhamostrado que os corais tinham mais probabilidade de morrer napresença de carbono orgânico dissolvido e que aquelas mortesestavam correlacionadas com o aumento no crescimento de mi-cróbios que normalmente vivem nas superfícies dos corais. Talvezas algas estivessem secretando carbono orgânico dissolvido (naforma de polissacarídeos em excesso produzidos pela fotossínte-se) e este carbono estava sustentando o crescimento microbianonos corais. Uma consequência poderia ser que os micróbios - amaioria bactérias - bloqueariam a difusão de oxigênio para ostecidos dos corais subjacentes e os sufocaria.
Smith e seus colegas criaram corais (Fig. 16.19) em recipien-tes, sozinhos ou em pares, com algas separadas deles por umafina malha, que deixava o carbono orgânico dissolvido passar,mas não os micróbios. Eles descobriram que os corais sobrevive-ram bem quando as algas estavam ausentes, mas sofreram altamortalidade quando elas estavam presentes. Em outro conjuntode recipientes, eles fizeram o mesmo experimento, mas adiciona-ram um antibiótico à água. Neste caso, os corais sobreviverambem com ou sem as algas presentes, confirmando que a mortali-dade do coral era devida aos micróbios que poderiam ser con-trolados por um antibiótico. Os investigadores repetiram este ex-
FIGo 16019 Os corais podem ser danificados indiretamente pelapresença de algas. A saúde de espécies de coral como este Pocillo-para verrucosa sofre quando as algas se estabelecem na vizinhança.Nalure Picturelibrary/ Alamy.
A maioria das espéciesde coral, mas nãa todas elas,sobreviveram melhor quando tratadas com antibióticos.
Sem tratamento antibiótico
Com tratamento antibiótico
j

perimento com um número maior de espécies de corais e algas.Nos recipientes sem tratamento antibiótico 95% das espécies decoral passaram por um declínio de saúde. Com o antibiótico,contudo, somente 20% das espécies de coral foram negativamen-te afetadas pela presença das algas (Fig. 16.20).
Em todos estes experimentos, não foi esclarecido por que osmicróbios que vivem naturalmente nos corais mataram os coraisquando as algas estavam por perto. Os investigadores conduzi-ram experimentos adicionais nos quais mediram o crescimentomicrobiano na superfície dos corais. Eles descobriram que, quan-do as algas secretam polissacarídeos em excesso, os micróbioscrescem mais rápido e consomem o oxigênio na água imediata-mente circundante aos corais, causando o sufocamento dos corais.O fato de que a saúde de algumas espécies de coral, tais comoPociúopora, era restaurada por tratamento antibiótico, enquantoa saúde de outras, como Acropora, não era, indica uma comple-xidade até mesmo maior neste sistema.
Este exemplo mostra que uma associação negativa entre duasespécies pode ocorrer sem competição direta por um recurso com-partilhado. No caso das algas e os corais, a interação negativa é
RESUMO
Competição 305
causada por uma cadeia de eventos iniciada pela emissão de pc-lissacarídeos pelas algas. Aqueles compostos de carbono favorecemo sobrecrescimento dos micróbios associados aos corais, que re-duzem as concentrcções de oxigênio na água a níveis que danifi-cam os últimos. E importante reconhecer que a poluição causadapelas atividades humanas é também uma fonte de compostos decarbono orgânico que promovem o crescimento microbiano nosrecifes de coral e é assim um fator importante na deterioração de-les, que agora vemos ocorrer em muitas partes do mundo .•
Como vimos nos Capítulos 14-16, as populações que cons-tituem as comunidades biológicas estão engajadas em diversasinterações que determinam suas abundâncias e distribuições eco-lógicas relativas, e mesmo sua capacidade em persistir, dentrodo sistema ecológico. Estas interações são também um compo-nente do ambiente que seleciona os atributos genéticos que dãoa cada população uma vantagem em suas diversas interações. Asrespostas evolutivas à seleção natural imposta pelas interaçõesdas espécies são o objeto do próximo capítulo.
1. A competição é qualquer uso ou defesa de um recursopor um indivíduo que reduz a disponibilidade daquele recur-so para outros indivíduos. Quando os indivíduos pertencemà mesma espécie, sua interação é chamada de competição in-traespecífica; quando pertencem a espécies diferentes, de in-terespecífica.
2. Um recurso pode ser definido como qualquer coisa que éconsumida e usada pelo consumidor para sua própria manuten-ção e crescimento, e cuja disponibilidade promove o crescimen-to da população de consumidores. Assim, o alimento, a água, aluz, os nutrientes minerais e o espaço são recursos, mas a tem-peratura e outras condições desse tipo não são.
3. A competição tende a ser mais intensa entre espécies pro-ximamente aparentadas, mas espécies distantemente aparentadastambém competem por recursos.
4. Os recursos podem ser classificados como não renováveis(espaço) ou renováveis (luz, água e alimento). Os recursos reno-váveis podem ser ainda mais distinguidos de acordo com a in-fluência do consumidor sobre sua taxa de suprimento: sem in-fluência, com influência direta ou influência indireta.
5. De acordo com a lei de Liebig, os recursos limitantes sãoaqueles que são mais escassos em relação à demanda. De todosos recursos consumidos por uma população, somente um ou unspoucos normalmente limitam seu crescimento. Dois ou mais re-cursos podem interagir sinergicamente para limitar a populaçãode um consumidor.
6. A diferença entre o crescimento populacional de uma espé-cie na presença e na ausência de outra é uma medida da intensi-dade da competição entre elas.
7. As investigações teóricas e os estudos de laboratório têmlevado à conclusão de que duas espécies competindo podem co-existir quando o mesmo recurso limita ambas as espécies. Estageneralização veio a ser conhecida como o princípio de exclusãocompetitiva.
8. Alguns tratamentos matemáticos da competição são basea-dos na equação logística para crescimento populacional, na qualum termo é adicionado para levar em conta o efeito da compe-tição interespecífica. A intensidade deste efeito é especificadapelo coeficiente de competição.
9. Os tamanhos populacionais de equilíbrio entre duas espéciescompetidoras podem ser descritos por uma equação que incluias capacidades de suporte e os coeficientes de competição paracada uma das espécies. Em termos mais gerais, a coexistênciaexige que o produto dos coeficientes de competição da primeirae da segunda espécie sejam menores do que 1. Em outras pala-vras, para coexistir, as espécies devem se auto limitar mais doque limitar uma à outra.10. Quando recursos diferentes limitam as populações de duasespécies, elas podem coexistir.11. Em alguns casos, duas espécies podem coexistir porque ca-da uma tem uma vantagem em relação a fatores diferentes noambiente, tais como a eficiência no uso de um recurso versus atolerância a um estresse. Tal competição assimétrica é comumna natureza.12. Os experimentos de transplante com plantas sob diversascondições de competição intra e interespecífica ilustram as di-ferenças dos mecanismos de competição entre habitats de alta ebaixa produtividade. Alguns experimentos mostraram que asplantas competem abaixo do solo por nutrientes quando os níveisde nutrientes são baixos, e acima do solo por luz quando os ní-veis de nutrientes são altos.13. Quando os indivíduos competem indiretamente através deseus efeitos sobre os recursos compartilhados, diz-se que a com-petição é exploradora; quando competem diretamente defenden-do os recursos um contra o outro, seu comportamento é conhe-cido como competição de interferência.14. A invasão rápida de um habitat por uma espécie animal mó-vel após a remoção de uma outra demonstra a competição deinterferência através do comportamento agressivo.15. Muitas plantas competem diretamente produzindo subs-tâncias químicas que impedem o crescimento e a sobrevivên-cia dos indivíduos de outras espécies. Este mecanismo é co-nhecido como alelopatia. As plantas podem também reduziras populações de seus competidores sustentando consumido-res de várias formas, como provendo refúgios seguros aeles.16. Os consumidores podem alterar o resultado das interaçõesde competição e promover a coexistência se eles seletivarnen-

306 Competição
te predarem sobre os competidores superiores. As interaçõesentre espécies de consumidores que são mediadas por seus
QUESTÕ-eS DE REVISÃO1. Qual é a diferença entre recursos renováveis e não renová-veis?2. Como a lei do mínimo de Liebig explica os limites sobre ocrescimento de uma população?3. Se duas espécies exigem o mesmo recurso limitante, o quevocê preveria sobre sua capacidade de coexistir?4. Quando duas espécies estão em equilíbrio, por que nós fazemosa taxa de crescimento populacional das espécies igual a zero?5. Compare e confronte as hipóteses de Grubb- Tilman e deGrime-Keddy em relação a como os níveis de nutrientes do so-lo afetam a intensidade da competição interespecífica.
LEITURAS SUGERIDASBartholomew, B. 1970. Bare zone between California shrub and grassland
communities: The role of animais. Science 170:1210-1212.Bruno, J. F., J. J. Stachowicz, and M. D. Bertness. 2003. Inclusion offaci-
litation into ecological theory. Trends in Ecology and Evolution 18:119-125.
Connell, J. H. 1961. The influence of interspecific competition and otherfactors on the distribution of the barnacle Chthamalus stellatus. Eco-logy42:710-723.
Emery, N. C., P. J. Ewanchuk, and M. D. Bertness. 2001. Competition andsalt-marsh plant zonation: Stress tolerators may be dominant competi-tors. Ecology 82:2471-2485.
Goldberg, D. E., and A. M. Barton. 1992. Patterns and consequences ofinterspecific competition in natural communities: A review offield ex-periments with plants. American Naturalist 139:771-801.
Grace,1. B., and D. Tilman (eds.). 1990. Perspectives on Plant Competi-tion. Academic Press, San Diego.
Halligan, 1. P. 1973. Bare areas associated with shrub stands in grasslands:The case of Artemisia californica. BioScience 23:429-432.
Halsey, R. W. 2004. In search of allelopathy: An eco-historical view of the in-vestigation of chemical inhibition in California coastal sage scrub and cha-mise chaparral, Journal ofthe TorreyBotanical Society 131:343-367.
Hardin, G. 1960. The competitive exclusion principie. Science 131:1292-1297.
Kadmon, R. 1995. Plant competition along soil moisture gradients: A fieldexperiment with the desert annual Stipa capensis. Journal of EcoLogy83:253-262.
próprios predadores ou patógenos são denominadas de compe-tição aparente.
6. Por que a alelopatia é considerada uma forma de competiçãode interferência?7. O que poderia ser um argumento para a hipótese de que es-pécies intimamente aparentadas deveriam experimentar umacompetição mais intensa do que as espécies distantemente apa-rentadas?8. Como é que a presença de consumidores pode causar um au-mento na diversidade de espécies em competição?
Keddy, P. 1989. Competition. Chapman & Hall, London.Maksimowich, D. S., and A. Mathis. 2000. Parasitized salamanders are
inferior competitors for territories and food resources. Ethology106:319-329.
Muller, C. H., W. H. Muller, and B. L. Haines, 1964. Volatile growth inhi-bitors produced by aromatic shrubs. Science 143:471-473.
Paine, R. T. 1974. Intertidal community structure: Experimental studies onthe relationship between a dominant competitor and its principal pre-dato r. Oecologia 15:93-120.
Schenk, J. J. 2006. Root competition: Beyond resource depletion. Journalof Ecology 94:725-739.
Schenk, H. J., R. M. Callaway, and B. E. Mahall. 1999. Spatial root segre-gation: Are plants territorial? Advances in. Ecological Research 28: 145-180.
Schoener, T. W. 1983. Field experiments on interspecific competition. Ame-rican Naturalist 122:240-285.
Smith, J. E., et aI. 2006. Indirect effect of algae on coral: Algae-mediatedmicrobe-induced coralmortality. Ecology Letters 9:835-845.
Stachowicz, J. J. 200 I. Mutualism, facilitation, and the structure of ecolo-gical communities. BioScience 51:235-246.
Tilman, D. 1977. Resource competition between planktonic algae: An ex-perimental and theoretical approach. Ecology 58:338-348.
Tilman, D. 1982. Resource Competition and Community Structure. Prin-ceton University Press, Princeton, NJ.
Wilson, S. D., and D. Tilman. 1993. Plant competition and resource availa-bility in response to disturbance and fertilization. Ecology 74:599-611.