enhanced heat shock protein 25 immunoreactivity in cranial
TRANSCRIPT

ORIGINAL ARTICLE
Enhanced heat shock protein 25 immunoreactivity in cranial nervemotoneurons and their related fiber tracts in rats prenatally-exposedto X-irradiation
Kazuhiko Sawada1, Shigeyoshi Saito3,4, Miwa Horiuchi-Hirose2, and Kenya Murase3,4
Departments of 1Physical Therapy, 2Nutrition, Faculty of Medical and Health Sciences, Tsukuba International University, Tsuchiura and3Department of Medical Engineering, Division of Health Sciences, and 4Center for Information and Neural Networks (CiNet), NationalInstitute of Information and Communications Technology, Osaka University Graduate School of Medicine, Osaka, Japan
ABSTRACT Alterations in histoarchitecture of the brain-stem were examined immunohistochemically in 4-week-oldrats with a single whole body X-irradiation at a dose of 0.5, 1.0,or 1.5 Gy on embryonic day (ED) 15 using anti-heat shockprotein 25 (HSP25). HSP25 immunostaining was seen in theneuronal perikarya of cranial nerve motoneurons, that is, themotor and mesencephalic nuclei of the trigeminal nerve, facialnucleus, abducens nucleus and accessory facial nucleus inthe pons, and the ambiguous nucleus, dorsal nucleus of vagusnerve and hypoglossus nucleus in the medulla oblongataof intact controls. In 0.5 to 1.5 Gy-irradiated rats, HSP25immunostaining in those neurons was more intense than incontrols, while the most intense immunostaining was marked in1.5 Gy-irradiated rats. HSP25 immunostaining was also appar-ent in the spinal tract of the trigeminal nerve and facial nervetracts in 0.5 to 1.5 Gy-irradiated rats, but was faint in controls.Interestingly, HSP25 immunostaining was aberrantly enhancedin dendritic arbors in the magnocellular region of medial ves-tibular nucleus of 0.5–1.5 Gy-irradiated rats. Those arbors wereidentified as excitatory secondary vestibulo-ocular neurons bydouble immunofluorescence for HSP25 and SMI-32. The resultssuggest an increase of HSP25 expression in cranial nervemotoneurons and their related fiber tracts from prenatal expo-sure to ionizing irradiation. This may be an adaptive responseto chronic hypoxia due to malformed brain arteries caused byprenatal ionizing irradiation.
Key Words: brainstem, cranial nerves, immunohistochmistry,ionizing irradiation, medulla oblongata, pons
INTRODUCTION
Prenatal exposure to irradiation causes severe harmful effects onthe development of various organs. The central nervous system isone of the organ systems most vulnerable to prenatal irradiation.Epidemiological studies have reported a high incidence of themicrocephalus brain with gray matter heterotopias in prenatally-exposed atomic bombing survivors of Hiroshima and Nagasaki(Blot and Miller 1973). Effects of prenatal radiation on variousbrain regions have often been reported in mice and rats, although
such studies focused mainly on the cerebral cortex (Fukui et al.1991; Miki et al. 1995; Saito et al. 2011, 2012), hippocampus (Mikiet al. 1999) and cerebellum (Li et al. 2002; Sawada et al. 2013).In terms of the brainstem, prenatal radiation induced increasedcalbindin D-28k immunostaining and/or astrocyte hypertrophywith increased expression of glial fibril acidic protein (GFAP) inthe nucleus of solitary tracts and hypoglossal nucleus (Jacquinet al. 2000; Kaneto et al. 2000). However, anatomical andcytoarchitectural alterations in the other brainstem regions have notbeen fully addressed.
Heat shock protein 25 (HSP25) is a human homolog of HSP27that is known as a pleiotropic inhibitor that mainly acts on stressedcells (Garrido et al. 1997, 1998, 1999; Park et al. 2000). In thecentral nervous system of rats and/or mice, constitutive expressionof HSP25 is seen in neurons in the cranial nerve motor nuclei andspinal cord ventral horn, the median preoptic nucleus, and in asubset of cerebellar Purkinje cells (Plumier et al. 1997; Armstrongand Hawkes 2000; Armstrong et al. 2000, 2001). Due to limiteddistributions of HSP25-expressing neurons, HSP25 is useful forinvestigating the histoarchitecture of the brainstem based on cranialnerve motoneurons. The present study aimed to clarify abnormali-ties in the lower brainstem (the pons and medulla oblongata)characterized by HSP25 immunostaining, using rats with prenatalionizing irradiation.
MATERIALS AND METHODS
AnimalsAll experimental procedures were conducted in accordance with theguidelines of the National Institutes of Health (NIH) for the Careand Use of Laboratory Animals (No. 80-23, revised 1996). TheInstitutional Animal Care and Use Committee of the University ofOsaka approved the procedures, and all efforts were made to mini-mize the number of animals used and their attendant suffering.Pregnant Sprague–Dawley rats (n = 4, 250–280 g) were obtainedfrom Japan SLC. Animals had free access to food and water, andwere kept under standard laboratory conditions of 22–23°C roomtemperature at around 50% humidity, and a 12/12 h light/darkcycle. Awake pregnant rats were exposed to a single whole-bodyX-irradiation at 0.5 Gy (n = 1), 1.0 Gy (n = 1) or 1.5 Gy (n = 1) onday 15 of pregnancy in a hand-made plastic box. The X-ray wasgiven at a dose rate of 0.88 Gy/min (120 kV, 15 mA with a 1-mm-thick aluminum filter, Rigaku Radioflex 350 X-ray Generator). Anintact pregnant rat (n = 1) was used as the control. The male off-spring at 4 weeks of age (control, n = 2; 0.5 Gy, n = 2; 1.0 Gy,n = 2; 1.5 Gy, n = 2) were deeply anesthetized with an overdose of
Correspondence: Kazuhiko Sawada, PhD, Department of Physical Therapy,Faculty of Medical and Health Sciences, Tsukuba International University,6-8-33 Manabe, Tsuchiura, Ibaraki 300-0051, Japan. Email: [email protected]
Received August 1, 2013; revised and accepted September 1, 2013.
bs_bs_banner
doi:10.1111/cga.12028 Congenital Anomalies 2014; 54, 87–93 87
© 2013 The AuthorsCongenital Anomalies © 2013 Japanese Teratology Society

pentobarbital (Dainippon Sumitomo Pharma Co., Ltd, Japan), andwere perfused intracordially with 0.9% NaCl (50 mL) followedby 4% paraformaldehyde in a 10 mM phosphate-buffered saline,pH 7.4 (200 mL).
ImmunohistochemistryThe brainstem including the pons and medulla oblongata wasembedded in OCT compound, and sectioned in the coronal planeat 40 μm by a Retoratome (REM-700; Yamato Koki Industrial,Asaka, Japan) with a refrigeration unit (Electro Freeze MC-802A,Yamato Kohki Industrial). Sections were reacted with a rabbitpolyclonal anti-HSP25 (1:1000, Stressgen, Victoria, BC, Canada:SPA-801, lot #B111411) containing 10% normal goat serum at4°C overnight. Anti-HSP25, which was raised against full-lengthrecombinant mouse HSP25 protein, recognizes both thephosphorylated and non-phosphorylated forms of HSP25. Anti-body absorption controls using full-length recombinant mouseHSP25 protein abolished all immunostaining (Armstrong et al.2000). After incubation, the sections were rinsed with PBS andreacted with biotinylated anti-mouse IgG. The immunoreactiveproducts were visualized by a Vectastain ABC elite kit (VectorLabs, Burlingame, CA, USA) using 0.01% 3,3′-diaminobenzidinetetrachloride (Sigma, St. Louis, MO, USA) in 0.03% H2O2 as achromogen.
For double immunostaining, the sections were reacted with amixture of anti-HSP25 (1:500) with a mouse anti-SMI-32 mono-clonal antibody (1:1000, Covance, Princeton, NJ, USA), or a mouseanti-calretinin monoclonal antibody (1:1000, Millipore, Temecula,CA, USA) containing 10% normal goat serum at 4°C. Anti-SMI-32recognized the non-phosphorylated form of the neurofilamentheavy chain. It gave a highly specific immunostaining pattern in therat, consistent with that reported previously (Murashov et al. 1998).Anti-calretinin produced highly specific immunostaining in the rattissues in a previous report (Liu et al. 2009). After washing withPBS, the sections were reacted with a mixture of an Alexa 594-labeled donkey anti-rabbit IgG antibody (1:500, Molecular Probes,Eugene, OR, USA) and an Alexa 488-labeled donkey anti-mouseIgG (1:500, Molecular Probes). Images of double-immunostainedsections were acquired with a fluorescence microscope (Axioimager A1; Zeiss, Gottingen, Germany).
Evaluation of the immunostaining intensityEvaluation of the immunostaining intensity was performed using amodified protocol designed by Sawada et al. (2008). To enable acomparison of the intensity of immunostaining, all sections wereprocessed under identical staining conditions. The immunostainingintensity was evaluated by its optical density (OD) on capturedimages using ImageJ software (National Institutes of Health,Bethesda, MD, USA). The captured images were converted into agrayscale digital image consisting of a 640 × 480 grid of pixels. Thebrightness level of each pixel ranged from 0 to 255 gray levels.HSP25-immunoreactive neurons were outlined manually using thePolygon tool of ImageJ software, after which the OD was meas-ured. The threshold of the OD value of each section was determinedby averaging two random polygons (area of POLYGON = 900 μm2)of the unstained neuropil.
RESULTS
Immunohistochemistry for HSP25Representative coronal images of HSP25-immunostained brainstemsections in control, 0.5 Gy-irradiated, 1.0 Gy-irradiated and 1.5 Gy-
irradiated rats are shown in Figure 1. In control rats, definite HSP25immunostaining appeared in neurons of major cranial nerve nuclei,that is, the motor and mesencephalic nuclei of the trigeminal nerve(Figs 1A,2A), facial nucleus (Fig. 1E), abducens nucleus (Fig. 2E)and accessory facial nucleus (Fig. 2I) in the pons, and in the ambigu-ous nucleus (Fig. 1M), dorsal nucleus of vagus nerve (Fig. 1M) andhypoglossus nucleus (Figs 1M,2M), in the medulla oblongata.
HSP25-immunoreactive neurons were distributed in the samecranial nerve nuclei of 0.5–1.5 Gy-irradiated rats, while HSP25staining was more intense than that in controls. The most intenseimmunostaining was marked in 1.5 Gy-irradiated rats. In controlrats, the facial nucleus, which mainly consisted of HSP25-immunoreactive neurons, was located on the ventral region of thebrainstem at the level of the inferior cerebellar peduncle, appearingdorsally to the spinal tracts of the trigeminal nerve (Fig. 1I). Thatnucleus was not seen at a more rostral level (the level of facial nervetracts) in the coronal section (Fig. 1E). In irradiated rats, the facialnucleus was more rostrally situated than in controls. The facialnucleus was seen through the level of the facial nerve tracts to thelevel of the inferior cerebellar peduncle (Fig. 1J–K). In addition, theventral spinocerebellar tract, which presents ventrally on the rostralregion of facial nucleus, was seen at the level of the inferior cer-ebellar peduncle in control rats (Fig. 1I). In irradiated rats, theventral spinocerebellar tract was seen ventrally in respect of thefacial nucleus at the level of facial nerve tracts (Fig. 1F–H), but notat the level of the inferior cerebellar peduncle (Fig. 1K). Thisrevealed that the ventral region of the pons, including the ventralspinocerebellar tract, in irradiated rats was shifted anteriorly byprenatal exposure to irradiation.
Figure 2 shows high magnification images of HSP25-immunoreactive neurons in mesencephalic nuclei of the trigeminalnerve, abducens nucleus, accessory facial nucleus, and ambiguousnucleus. HSP25 staining in those neurons was more intense inirradiated rats than in control rats, and strong HSP25 stainingfilled dendritic arbors in 1.5 Gy-irradiated rats (Fig. 2D,H,L). Inthe mesencephalic nuclei, HSP25 immunoreactive fibers extend-ing from the motor nucleus of the trigeminal nerve were seen(arrowheads in Fig. 2D). HSP25 immunostaining further appearedin the spinal tract of the trigeminal nerve and facial nerve tracts,as well as the cranial nerve motor nuclei in irradiated rats(Fig. 1B–D,F–H,J–L,M–P). In 1.5 Gy-irradiated rats, a number ofHSP25-immunoreactive fibers were extended from the facialnucleus through the genu of facial nerve tracts (arrowheads inFig. 1L).
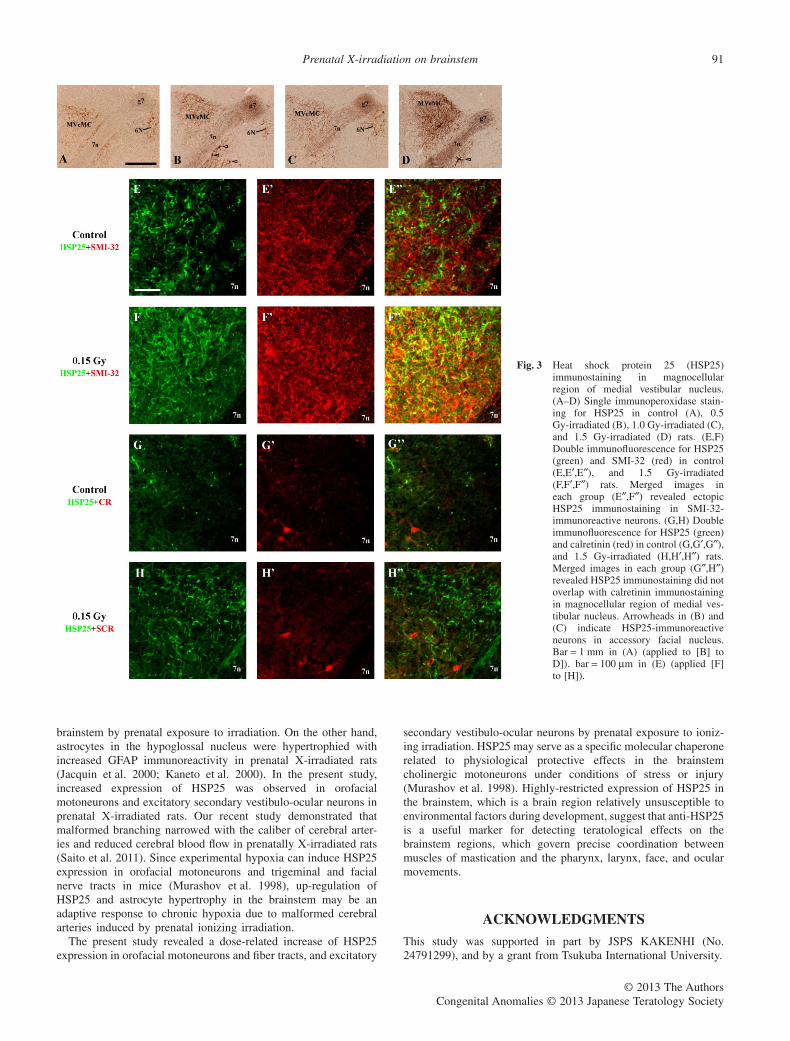
The most noticeable results in the present study are thatneuronal perikarya with dendritic arbors in the magnocellularregion of the medial vestibular nucleus ectopically expressedHSP25 immunostaining in irradiated rats (Figs 1F–H,3B–D). Aswith the other neurons and fiber tracts, the most intense HSP25staining in those neurons was obtained in 1.5 Gy-irradiated rats(Fig. 3D). Two distinct neuronal populations are known to bepresent in the magnocellular region of the medial vestibularnucleus: SMI-32-immunoreactive excitatory secondary vestibulo-ocular neurons; and calretinin-immunoreactive neurons belongingto the excitatory upgaze pathways for eye movements (Ahlfeldet al. 2011). In control and irradiated rats, calretinin- and SMI-32-immunoreactive neurons were present in the magnocelllular region(Fig. 3E,G). Double immunofluorescence revealed that increasedHSP25 immunostaining appeared in SMI-32-immunoreactiveneurons, but not in calretinin-immunoreactive neurons in irradiatedrats (Fig. 3F,H). Therefore, HSP25-expressing neurons in themagnocellular region of the medial vestibular nucleus were iden-tified as excitatory secondary vestibulo-ocular neurons.
88 K. Sawada et al.
© 2013 The AuthorsCongenital Anomalies © 2013 Japanese Teratology Society
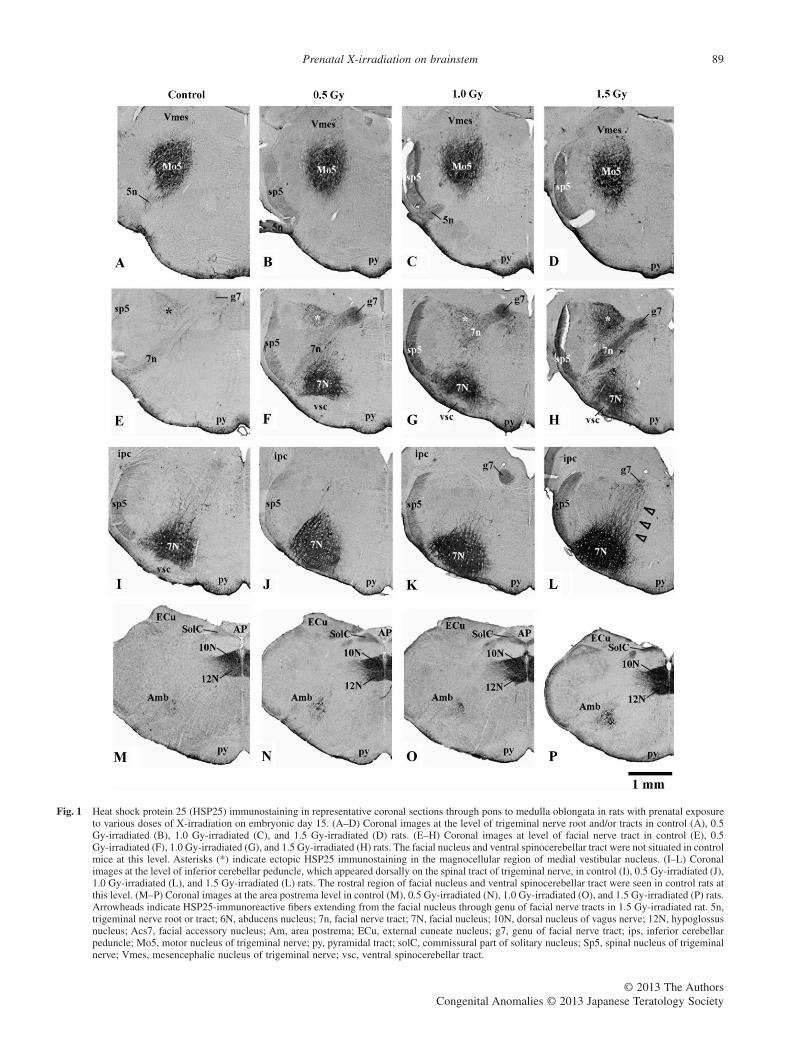
Fig. 1 Heat shock protein 25 (HSP25) immunostaining in representative coronal sections through pons to medulla oblongata in rats with prenatal exposureto various doses of X-irradiation on embryonic day 15. (A–D) Coronal images at the level of trigeminal nerve root and/or tracts in control (A), 0.5Gy-irradiated (B), 1.0 Gy-irradiated (C), and 1.5 Gy-irradiated (D) rats. (E–H) Coronal images at level of facial nerve tract in control (E), 0.5Gy-irradiated (F), 1.0 Gy-irradiated (G), and 1.5 Gy-irradiated (H) rats. The facial nucleus and ventral spinocerebellar tract were not situated in controlmice at this level. Asterisks (*) indicate ectopic HSP25 immunostaining in the magnocellular region of medial vestibular nucleus. (I–L) Coronalimages at the level of inferior cerebellar peduncle, which appeared dorsally on the spinal tract of trigeminal nerve, in control (I), 0.5 Gy-irradiated (J),1.0 Gy-irradiated (L), and 1.5 Gy-irradiated (L) rats. The rostral region of facial nucleus and ventral spinocerebellar tract were seen in control rats atthis level. (M–P) Coronal images at the area postrema level in control (M), 0.5 Gy-irradiated (N), 1.0 Gy-irradiated (O), and 1.5 Gy-irradiated (P) rats.Arrowheads indicate HSP25-immunoreactive fibers extending from the facial nucleus through genu of facial nerve tracts in 1.5 Gy-irradiated rat. 5n,trigeminal nerve root or tract; 6N, abducens nucleus; 7n, facial nerve tract; 7N, facial nucleus; 10N, dorsal nucleus of vagus nerve; 12N, hypoglossusnucleus; Acs7, facial accessory nucleus; Am, area postrema; ECu, external cuneate nucleus; g7, genu of facial nerve tract; ips, inferior cerebellarpeduncle; Mo5, motor nucleus of trigeminal nerve; py, pyramidal tract; solC, commissural part of solitary nucleus; Sp5, spinal nucleus of trigeminalnerve; Vmes, mesencephalic nucleus of trigeminal nerve; vsc, ventral spinocerebellar tract.
Prenatal X-irradiation on brainstem 89
© 2013 The AuthorsCongenital Anomalies © 2013 Japanese Teratology Society

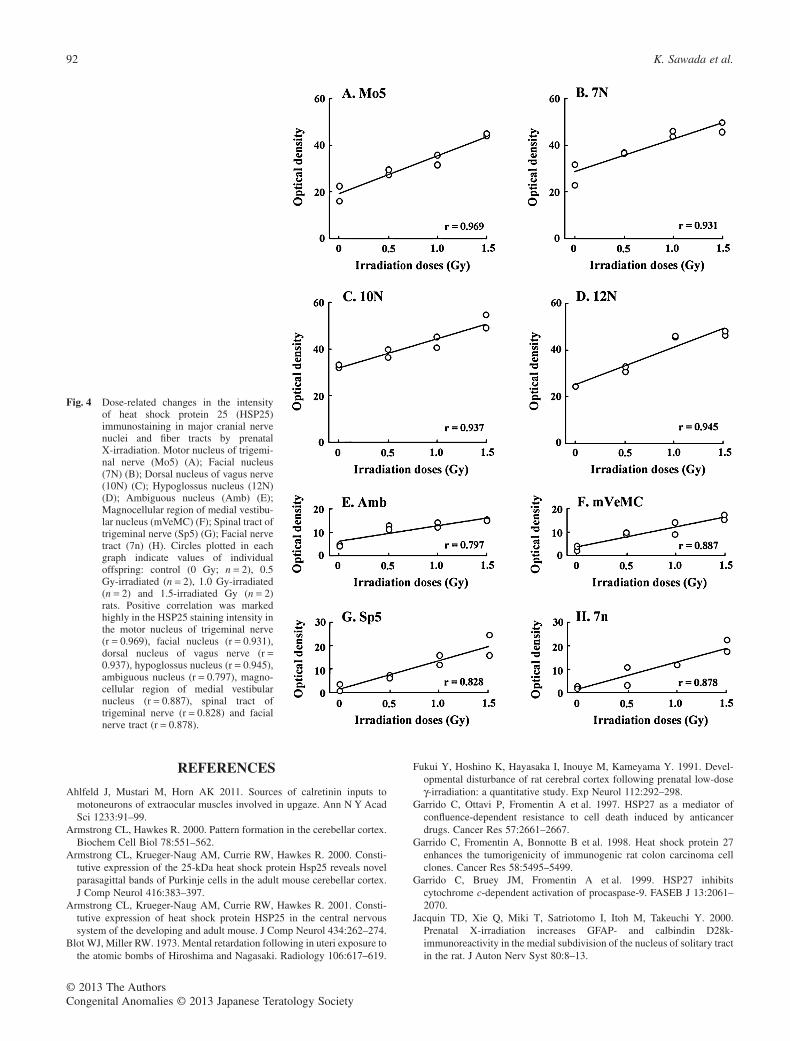
Evaluation of immunostaining intensityThe intensity of HSP25 immunostaining was evaluated in majorcranial nerve nuclei and nerve fiber tracts of control and irradiatedrats with the results shown in Figure 4. The staining intensity wasvariable among the brainstem regions examined, and enhancedwith increasing doses of prenatal X-irradiation. Correlations ofirradiation doses with the staining intensity were examined. Apositive correlation was very marked in the HSP25 staining inten-sity in the motor nucleus of trigeminal nerve (r = 0.969), facialnucleus (r = 0.931), dorsal nucleus of vagus nerve (r = 0.937),hypoglossus nucleus (r = 0.945), ambiguous nucleus (r = 0.797),magnocellular region of medial vestibular nucleus (r = 0.887),spinal tract of trigeminal nerve (r = 0.828) and facial nerve tract(r = 0.878).
DISCUSSION
Heat shock protein 25 is expressed constitutively in limited motorand sensory neurons of the brainstem and spinal cord, andneurons in the preoptic area in rats and mice (Plumier et al. 1997;Armstrong et al. 2001), and zebrin II-expressing Purkinje subsetsin the cerebellum of mice (Armstrong and Hawkes 2000;Armstrong et al. 2000). In particular, high levels of HSP25expression are seen in cranial nerve motoneurons in the mousebrainstem (Plumier et al. 1997; Armstrong et al. 2001). HSP25 isthought to have a protective effect on brain injury, becauseexcitotoxic damage to the immature brain induces HSP25 expres-sion in surviving reactive astrocytes, which co-expresses cleavedcaspase-3 (Villapol et al. 2008). In our recent study, HSP25
expressing Purkinje cell subsets in the cerebellum of Cav2.1mutant mice (rolling mice) did not express tyrosine hydroxylase(Sawada et al. 2010), which is known to appear ectopically inzebrin II-expressing Purkinje cell populations preceding thedegeneration of those neurons (Sarna et al. 2006). Thus, constitu-tive expression of HSP25 in the central nervous system may haveprotective effects against stress or injury.
The present study revealed an increase of HSP25 expression inthe same brainstem regions by prenatal exposure to X-irradiation inrats. Furthermore, SMI-32-immunoreactive dendritic arbors in themagnocellular region of the medial vestibular nucleus enhancedHSP25 immunostaining in prenatally-irradiated rats. Since thoseSMI-32-immunoreactive neurons are identified as excitatory sec-ondary vestibulo-ocular neurons in that region (Ahlfeld et al. 2011),prenatal exposure to ionizing irradiation may be involved inup-regulation of HSP25 in orofacial motoneurons and fiber tracts,and excitatory secondary vestibulo-ocular neurons.
The developing brainstem is relatively unsusceptible to envi-ronmental factors including ionizing irradiation, compared to theother developing brain regions. A single exposure of X-irradiationduring mid-gestation causes certain brain abnormalities such asderanged architecture of the cerebral cortex (Sun et al. 1999) andhippocampus (Miki et al. 1999), and cortical hypoplasia with lossof neurons in the cerebrum (Miki et al. 1995) and cerebellum (Liet al. 2002; Sawada et al. 2013) in rats and mice. However,obvious histoarchitectural alterations in the brainstem have notbeen reported. In the present study, the ventral region of the pons,including the facial nucleus and ventral spinocerebellar tract, inirradiated rats was shifted anteriorly, suggesting hypoplasia of the
Fig. 2 High-magnification images of heatshock protein 25 (HSP25)-immunoreactive neurons in mesen-cephalic nucleus, abducens nucleus,accessory facial nucleus and ambiguousnucleus. (A–D) HSP25-immunoreactiveneurons in mesencephalic nucleus ofcontrol (A), 0.5 Gy-irradiated (B), 1.0Gy-irradiated (C), and 1.5 Gy-irradiated(D) rats. Arrowheads indicate HSP25-immunoreactive fibers from the motornucleus of trigeminal nerve. (E–H)HSP25-immunoreactive neurons inabducens nucleus of control (E), 0.5Gy-irradiated (F), 1.0 Gy-irradiated (G),and 1.5 Gy-irradiated (H) rats. (I–L)HSP25-immunoreactive neurons inaccessory facial nucleus of control (I),0.5 Gy-irradiated (J), 1.0 Gy-irradiated(K), and 1.5 Gy-irradiated (L) rats.(M–P) HSP25-immunoreactive neuronsin ambiguous nucleus of control (M),0.5 Gy-irradiated (N), 1.0 Gy-irradiated(O), and 1.5 Gy-irradiated (P) rats.Bar = 10 μm in (A) (applied to [B] to[P]).
90 K. Sawada et al.
© 2013 The AuthorsCongenital Anomalies © 2013 Japanese Teratology Society
brainstem by prenatal exposure to irradiation. On the other hand,astrocytes in the hypoglossal nucleus were hypertrophied withincreased GFAP immunoreactivity in prenatal X-irradiated rats(Jacquin et al. 2000; Kaneto et al. 2000). In the present study,increased expression of HSP25 was observed in orofacialmotoneurons and excitatory secondary vestibulo-ocular neurons inprenatal X-irradiated rats. Our recent study demonstrated thatmalformed branching narrowed with the caliber of cerebral arter-ies and reduced cerebral blood flow in prenatally X-irradiated rats(Saito et al. 2011). Since experimental hypoxia can induce HSP25expression in orofacial motoneurons and trigeminal and facialnerve tracts in mice (Murashov et al. 1998), up-regulation ofHSP25 and astrocyte hypertrophy in the brainstem may be anadaptive response to chronic hypoxia due to malformed cerebralarteries induced by prenatal ionizing irradiation.
The present study revealed a dose-related increase of HSP25expression in orofacial motoneurons and fiber tracts, and excitatory
secondary vestibulo-ocular neurons by prenatal exposure to ioniz-ing irradiation. HSP25 may serve as a specific molecular chaperonerelated to physiological protective effects in the brainstemcholinergic motoneurons under conditions of stress or injury(Murashov et al. 1998). Highly-restricted expression of HSP25 inthe brainstem, which is a brain region relatively unsusceptible toenvironmental factors during development, suggest that anti-HSP25is a useful marker for detecting teratological effects on thebrainstem regions, which govern precise coordination betweenmuscles of mastication and the pharynx, larynx, face, and ocularmovements.
ACKNOWLEDGMENTS
This study was supported in part by JSPS KAKENHI (No.24791299), and by a grant from Tsukuba International University.
Fig. 3 Heat shock protein 25 (HSP25)immunostaining in magnocellularregion of medial vestibular nucleus.(A–D) Single immunoperoxidase stain-ing for HSP25 in control (A), 0.5Gy-irradiated (B), 1.0 Gy-irradiated (C),and 1.5 Gy-irradiated (D) rats. (E,F)Double immunofluorescence for HSP25(green) and SMI-32 (red) in control(E,E′,E″), and 1.5 Gy-irradiated(F,F′,F″) rats. Merged images ineach group (E″,F″) revealed ectopicHSP25 immunostaining in SMI-32-immunoreactive neurons. (G,H) Doubleimmunofluorescence for HSP25 (green)and calretinin (red) in control (G,G′,G″),and 1.5 Gy-irradiated (H,H′,H″) rats.Merged images in each group (G″,H″)revealed HSP25 immunostaining did notoverlap with calretinin immunostainingin magnocellular region of medial ves-tibular nucleus. Arrowheads in (B) and(C) indicate HSP25-immunoreactiveneurons in accessory facial nucleus.Bar = 1 mm in (A) (applied to [B] toD]). bar = 100 μm in (E) (applied [F]to [H]).
Prenatal X-irradiation on brainstem 91
© 2013 The AuthorsCongenital Anomalies © 2013 Japanese Teratology Society
REFERENCES
Ahlfeld J, Mustari M, Horn AK 2011. Sources of calretinin inputs tomotoneurons of extraocular muscles involved in upgaze. Ann N Y AcadSci 1233:91–99.
Armstrong CL, Hawkes R. 2000. Pattern formation in the cerebellar cortex.Biochem Cell Biol 78:551–562.
Armstrong CL, Krueger-Naug AM, Currie RW, Hawkes R. 2000. Consti-tutive expression of the 25-kDa heat shock protein Hsp25 reveals novelparasagittal bands of Purkinje cells in the adult mouse cerebellar cortex.J Comp Neurol 416:383–397.
Armstrong CL, Krueger-Naug AM, Currie RW, Hawkes R. 2001. Consti-tutive expression of heat shock protein HSP25 in the central nervoussystem of the developing and adult mouse. J Comp Neurol 434:262–274.
Blot WJ, Miller RW. 1973. Mental retardation following in uteri exposure tothe atomic bombs of Hiroshima and Nagasaki. Radiology 106:617–619.
Fukui Y, Hoshino K, Hayasaka I, Inouye M, Kameyama Y. 1991. Devel-opmental disturbance of rat cerebral cortex following prenatal low-doseγ-irradiation: a quantitative study. Exp Neurol 112:292–298.
Garrido C, Ottavi P, Fromentin A et al. 1997. HSP27 as a mediator ofconfluence-dependent resistance to cell death induced by anticancerdrugs. Cancer Res 57:2661–2667.
Garrido C, Fromentin A, Bonnotte B et al. 1998. Heat shock protein 27enhances the tumorigenicity of immunogenic rat colon carcinoma cellclones. Cancer Res 58:5495–5499.
Garrido C, Bruey JM, Fromentin A et al. 1999. HSP27 inhibitscytochrome c-dependent activation of procaspase-9. FASEB J 13:2061–2070.
Jacquin TD, Xie Q, Miki T, Satriotomo I, Itoh M, Takeuchi Y. 2000.Prenatal X-irradiation increases GFAP- and calbindin D28k-immunoreactivity in the medial subdivision of the nucleus of solitary tractin the rat. J Auton Nerv Syst 80:8–13.
Fig. 4 Dose-related changes in the intensityof heat shock protein 25 (HSP25)immunostaining in major cranial nervenuclei and fiber tracts by prenatalX-irradiation. Motor nucleus of trigemi-nal nerve (Mo5) (A); Facial nucleus(7N) (B); Dorsal nucleus of vagus nerve(10N) (C); Hypoglossus nucleus (12N)(D); Ambiguous nucleus (Amb) (E);Magnocellular region of medial vestibu-lar nucleus (mVeMC) (F); Spinal tract oftrigeminal nerve (Sp5) (G); Facial nervetract (7n) (H). Circles plotted in eachgraph indicate values of individualoffspring: control (0 Gy; n = 2), 0.5Gy-irradiated (n = 2), 1.0 Gy-irradiated(n = 2) and 1.5-irradiated Gy (n = 2)rats. Positive correlation was markedhighly in the HSP25 staining intensity inthe motor nucleus of trigeminal nerve(r = 0.969), facial nucleus (r = 0.931),dorsal nucleus of vagus nerve (r =0.937), hypoglossus nucleus (r = 0.945),ambiguous nucleus (r = 0.797), magno-cellular region of medial vestibularnucleus (r = 0.887), spinal tract oftrigeminal nerve (r = 0.828) and facialnerve tract (r = 0.878).
92 K. Sawada et al.
© 2013 The AuthorsCongenital Anomalies © 2013 Japanese Teratology Society

Kaneto T, Xie Q, Ito M, Sakurai T, Satriotomo I, Takeuchi Y. 2000. Theeffects of prenatal X-irradiation on hypoglossal nucleus: a GFAP immu-nohistochemical study. Okajimas Folia Anat Jpn 7:181–188.
Li HP, Miki T, Gu H et al. 2002. The effect of the timing of prenatalX-irradiation on Purkinje cell numbers in rat cerebellum. Brain Res DevBrain Res 139:159–166.
Liu F, You Y, Li XI et al. 2009. Brain injury does not alter the intrinsicdifferentiation potential of adult neuroblasts. J Neurosci 29:5075–5087.
Miki T, Fukui Y, Takeuchi Y, Itoh M. 1995. A quantitative study of theeffects of prenatal X-irradiation on the development of cerebral cortex inrats. Neurosci Res 23:241–247.
Miki T, Sawada K, Sun X-Z, Hisano S, Takeuchi Y, Fukui Y. 1999. Abnor-mal distribution of hippocampal mossy fibers in rats exposed toX-irradiation in utero. Brain Res Dev Brain Res 112:275–280.
Murashov AK, Talebian S, Wolgemuth DJ. 1998. Role of heat shock proteinHsp25 in the response of the orofacial nuclei motor system to physiologi-cal stress. Brain Res Mol Brain Res 63:14–24.
Park SH, Cho HN, Lee SJ et al. 2000. Hsp25-induced radioresistance isassociated with reduction of death by apoptosis: involvement of Bcl2 andthe cell cycle. Radiat Res 154:421–428.
Plumier JCL, Hopkins DA, Robertson HA, Currie RW. 1997. Constitutiveexpression of the 27-kDa heat shock protein HSP27 in sensory and motorneurons of the rat nervous system. J Comp Neurol 384:409–428.
Saito S, Aoki I, Sawada K et al. 2011. Quantitative and noninvasive assess-ment of prenatal X-ray irradiation-induced CNS hypoplasia using mag-netic resonance imaging. Radiat Res 175:1–9.
Saito S, Aoki I, Sawada K, Suhara T. 2012. Quantitative assessment ofCNS disorder induced by prenatal X-ray exposure using diffusion andmanganese-enhanced MRI (MEMRI). NMR Biomed 25:75–83.
Sarna JR, Marzban H, Watanabe M, Hawkes R. 2006. Complementarystripes of phospholipase Cβ3 and Cβ4 expression by Purkinje cell subsetsin the mouse cerebellum. J Comp Neurol 496:303–313.
Sawada K, Hosoi E, Bando M et al. 2008. Differential alterations in expres-sions of ryanodine receptor subtypes in cerebellar cortical neurons of anataxic mutant, rolling mouse Nagoya. Neuroscience 152:609–617.
Sawada K, Sakata-Haga H, Fukui Y. 2010. Alternating array of tyrosinehydroxylase and heat shock protein 25 immunopositive Purkinje cellstripes in zebrin II-defined transverse zone of the cerebellum of rollingmouse Nagoya. Brain Res 1343:46–53.
Sawada K, Saito S, Horiuchi-Hirose M, Mori Y, Yoshioka Y, Murase K.2013. Dose-related cerebellar abnormality in rats with prenatal exposureto X-irradiation by MRI volumetric analysis. Congenit Anom (Kyoto)53:127–130
Sun XZ, Takahashi S, Fukui Y et al. 1999. Different patterns of abnormalneuronal migration in the cerebral cortex of mice exposed to irradiation.Brain Res Dev Brain Res 114:99–108.
Villapol S, Acarin L, Faiz M, Castellano B, Gonzalez B. 2008. Survivin andheat shock protein 25/27 colocalize with cleaved caspase-3 in survivingreactive astrocytes following excitotoxicity to the immature brain. Neu-roscience 153:108–119.
Prenatal X-irradiation on brainstem 93
© 2013 The AuthorsCongenital Anomalies © 2013 Japanese Teratology Society