epstein-barr virus infection induces indoleamine 2,3-dioxygenase
TRANSCRIPT

Epstein-Barr Virus Infection Induces Indoleamine 2,3-DioxygenaseExpression in Human Monocyte-Derived Macrophages through p38/Mitogen-Activated Protein Kinase and NF-�B Pathways: Impairmentin T Cell Functions
Wan-li Liu,a,b Yue-hao Lin,a,b Han Xiao,c Shan Xing,a,b Hao Chen,a,b Pei-dong Chi,a,b Ge Zhangc
State Key Laboratory of Oncology in Southern China, Collaborative Innovation Center of Cancer Medicine, Sun Yat-sen University Cancer Center,a Department of ClinicLab, Sun Yat-sen University Cancer Center,b and Department of Microbial and Biochemical Pharmacy, School of Pharmaceutical Sciences, Sun Yat-sen University,c
Guangzhou, China
ABSTRACT
Epstein-Barr virus (EBV) infection has been observed in tumor-infiltrated macrophages, but its infection effects on macrophageimmune functions are poorly understood. Here, we showed that some macrophages in the tumor stroma of nasopharyngeal car-cinoma (NPC) tissue expressed the immunosuppressive protein indoleamine 2,3-dioxygenase (IDO) more strongly than did tu-mor cells. EBV infection induced mRNA, protein, and enzymatic activity of IDO in human monocyte-derived macrophages(MDMs). Infection increased the production of tumor necrosis factor alpha (TNF-�) and interleukin-6 (IL-6), whereas the neu-tralizing antibodies against TNF-� and IL-6 inhibited IDO induction. EBV infection also activated the mitogen-activated proteinkinase (MAPK) p38 and NF-�B, and the inhibition of these two pathways with SB202190 and SN50 almost abrogated TNF-� andIL-6 production and inhibited IDO production. Moreover, the activation of IDO in response to EBV infection of MDMs sup-pressed the proliferation of T cells and impaired the cytotoxic activity of CD8� T cells, whereas the inhibition of IDO activitywith 1-methyl-L-tryptophan (1-MT) did not affect T cell proliferation and function. These findings indicate that EBV-inducedIDO expression in MDMs is substantially mediated by IL-6- and TNF-�-dependent mechanisms via the p38/MAPK and NF-�Bpathways, suggesting that a possible role of EBV-mediated IDO expression in tumor stroma of NPC may be to create a microen-vironment of suppressed T cell immune responses.
IMPORTANCE
CD8� cytotoxic T lymphocytes (CTLs) play an important role in the control of viral infections and destroy tumor cells. Activa-tion of the tryptophan-catabolizing enzyme indoleamine 2,3-dioxygenase (IDO) in cancer tissues facilitates immune escape bythe impairment of CTL functions. IDO expression was observed in some macrophages of the tumor stroma of nasopharyngealcarcinoma (NPC) tissue, and IDO could be induced in Epstein-Barr virus (EBV)-infected human monocyte-derived macro-phages (MDMs). NPC cells and macrophages have been found to produce IDO in a gamma interferon (IFN-�)-dependent man-ner. Instead, EBV-induced IDO expression in MDMs is substantially mediated by IL-6- and TNF-�-dependent mechanisms viathe p38/MAPK and NF-�B pathways, which suppressed the proliferation of T cells and impaired the cytotoxic activity of CD8� Tcells. This finding provides a new interpretation of the mechanism of immune escape of EBV and shows the immunosuppressiverole of EBV-mediated IDO expression in tumor stroma of NPC.
Epstein-Barr virus (EBV) is a ubiquitous human virus of theherpesvirus family that is found in �90% of the world’s pop-
ulation. Infection with EBV is associated with infectious mononu-cleosis and human malignancies including Burkitt’s lymphomaand nasopharyngeal carcinoma (NPC) (1, 2).
EBV can infect monocytes/macrophages, intraepithelial mac-rophages, and Langerhans cells (3, 4). Moreover, EBV expressionin macrophages infiltrating NPC, Burkitt’s lymphoma, and pri-mary lung lymphoma has also been observed (5, 6). The interac-tion of EBV with monocytes has been demonstrated to suppress itsphagocytic activity and inhibit its potent antiviral activity (7, 8).EBV infection inhibits the development of dendritic cells by pro-moting the apoptosis of their monocyte precursors (9). Con-versely, one study reported that EBV infection of monocytes en-hanced their survival and rapidly induced their maturation intomacrophages with the characteristics of potent antigen-present-ing cells (APCs) (10). However, the effects of EBV infection onmacrophage immune functions are poorly understood.
An immunomodulatory role for the enzyme indoleamine 2,3-dioxygenase (IDO) in macrophage functions has been suggested(11). IDO catalyzes the conversion of tryptophan into kynure-nine, and altered IDO activity is often associated with pathologyincluding neoplasia and autoimmunity (12). Several studies havedescribed IDO-dependent T cell suppression by APCs in manyinfectious and inflammatory conditions, indicating that biochem-ical changes due to tryptophan catabolism have a profound effect
Received 15 December 2013 Accepted 24 March 2014
Published ahead of print 2 April 2014
Editor: R. M. Longnecker
Address correspondence to Ge Zhang, [email protected].
W.L. and Y.L. contributed equally to this work.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.03678-13
6660 jvi.asm.org Journal of Virology p. 6660 – 6671 June 2014 Volume 88 Number 12
on March 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

on T cell proliferation and effector functions in tissue microenvi-ronments (13–15). IDO-mediated tryptophan metabolism, notonly in APCs but also in tumor cells, represents a vital mechanismfor potential T cell suppression during tumor growth (16). Ourprevious study indicated that exposure to the milieu created by anIDO-positive NPC cell line significantly impaired lymphocyte cy-totoxicity against target tumor cells (17).
IDO expression is induced in macrophages and several othercell types under various physiological conditions, such as inflam-mation induced by viral and bacterial infections (18). Infectionwith dengue virus, HIV, poliovirus, and hepatitis C virus is asso-ciated with IDO induction in various tissues and cell types, both invivo and in vitro (19–22). Previously reported evidence suggeststhat EBV infection increases the expression level of IDO in B cellsand inhibits NK cell cytotoxicity (23). To date, whether IDO canbe induced by EBV-infected macrophages and what effects EBVinduction of IDO may have on macrophage immune functionshave not been investigated.
To elucidate the potential role of IDO in macrophages duringEBV infection, we investigated the signaling mechanisms bywhich EBV infection induces IDO expression in human mono-cyte-derived macrophages (MDMs) and the possible role of EBV-mediated IDO induction in creating a microenvironment of sup-pressed T cell immune responses in MDMs.
MATERIALS AND METHODSCell culture. Peripheral blood mononuclear cells (PBMCs) were isolatedby Ficoll-Paque Plus gradient centrifugation of leukopacks derived from25 healthy adult volunteers. Informed consent was obtained from thevolunteers prior to participation, in accordance with the human experi-mentation guidelines of the Institute Research Ethics Committee of theCancer Centre, Sun Yat-Sen University. CD14� monocytes were isolatedfrom PBMCs by using magnetic CD14 microbeads (Miltenyi Biotech)according to the manufacturer’s instructions. CD14� cells were maturedinto MDMs by 8 to 12 days in medium with 25 ng/ml macrophage colony-stimulating factor (M-CSF) (R&D Systems). The isolation of CD4� andCD8� T lymphocytes from PBMCs was performed by means of immu-nomagnetic bead separation using a Dynal CD4- or CD8-positive isola-tion kit (Invitrogen Dynal). The purity of CD4� and CD8� cells was�98%, as determined by flow cytometry. CNE2 (human NPC cell line)and B95-8 (EBV-positive B cell line) cells were maintained in our labora-tory. All of these cells were grown in complete RPMI 1640 medium sup-plemented with 10% heat-inactivated fetal bovine serum (HyClone), 100�g/ml streptomycin, and 100 U/ml penicillin under a humidified 5% CO2
atmosphere at 37°C in a CO2 incubator.EBV preparation and cell treatment. EBV was purified from B95-8
cell culture supernatants as previously described (24). In brief, virus waspropagated by inducing B95-8 cells with 30 ng/ml phorbol myristate ac-etate (PMA) and 4 mM butyric acid (both from Sigma) for 3 days. Thevirus was then pelleted by ultracentrifugation at 150,000 rpm for 90 min,filtered through a 0.8-mm pore size filter, and stored at �80°C. Viral titerswere evaluated and adjusted to 2 � 107 transforming units (TFU)/ml.Where indicated, virus stocks were inactivated by pretreatment with 9 mJof UV radiation with a Stratalinker instrument, which completely abol-ished their ability to infect cells.
For the preparation of conditioned medium (CM), MDM cellswere seeded at 5 � 105 cells/ml for 24 h. Next, cells were infected withEBV or UV-irradiated EBV (UV-EBV) at various TFU/ml values forthe indicated times. In certain experiments, 100 �M 1-methyl-L-tryp-tophan (1-MT) (Sigma-Aldrich) was added to the medium. Subse-quently, the culture supernatants were harvested as CM and stored inaliquots at �80°C before use.
For antibody-blocking experiments, MDMs were preincubated with
10 �g/ml of neutralizing antibodies against tumor necrosis factor alpha(TNF-�) (mouse IgG1 [mIgG1]; eBioscience) or interleukin-6 (IL-6)(mIgG1; R&D systems) or isotype-matched control mIgG1 antibodies(eBioscience) for 1 h at 37°C before EBV infection.
For inhibitor-blocking experiments, MDMs were preincubated for 1 hat 37°C with 10 �M U0126 (MEK/extracellular signal-regulated kinase[ERK] inhibitor), 10 �M SB203580 (p38 inhibitor), 10 �M SP600125(Jun N-terminal protein kinase [JNK] inhibitor), 0.3 �M wortmannin(phosphatidylinositol 3-kinase [PI3K] inhibitor), or 30 �M SN50 (NF-�Binhibitor). NF-�B SN50M (30 �M) (the inactive control peptide) wasused as a negative control for SN50. Dimethyl sulfoxide (DSMO) was usedas a vehicle control. U0126 was purchased from Cell Signaling Technol-ogy, and other reagents were obtained from Calbiochem. MDMs weresubsequently infected with EBV at 0.2 � 105 TFU for 48 h.
Immunohistochemistry and immunofluorescence. Immunohisto-chemistry and immunofluorescence were performed by using paraffin-embedded NPC tissues or EBV-infected MDMs (EBV-MDMs). For im-munohistochemistry, a rabbit polyclonal anti-IDO antibody (1:200;homemade) (17) and a mouse monoclonal anti-CD68 antibody (1:100;BD Pharmingen) were incubated with tissue sections overnight at 4°C.After washing, tissue sections were treated with biotinylated anti-mouseor anti-rabbit antibody (Zymed), followed by further incubation with thestreptavidin-horseradish peroxidase (HRP) conjugate. For immunofluo-rescence, tissue sections or MDM cells were incubated with antibodiesagainst IDO (1:200) or CD68-phycoerythrin (PE) (1:100; BD Pharmin-gen) or the serum of a NPC patient who was positive for the EBV antibody(1:100) (viral capsid antigen [VCA]-IgA titer of 1:1,280, as determined byan immunofluorescence assay) at 4°C overnight. After washing, the slideswere incubated with fluorescein isothiocyanate (FITC)-conjugated sec-ondary antibodies (1:1,000; Abcam) or Alexa Fluor 594-conjugated anti-human IgG1 antibodies (1:1,000; BD Biosciences), and the cell nuclei werestained with 4=,6-diamidino-2-phenylindole (DAPI) (Sigma). The num-ber of CD68� and IDO� cells in the tumor stroma was counted. The meanvalues were derived from a visual analysis of 10 high-power fields (HPFs)(magnification, �400), using a Nikon Eclipse E-200 microscope, whichcorresponded to an area of 0.5 mm2 per HPF.
Flow cytometry. CD14� cells were separated from PBMCs by the useof magnetic CD14 microbeads (Miltenyi Biotech). MDMs were generatedfrom CD14� cells in the presence of M-CSF. MDMs were infected withEBV at 0.2 � 105 TFU/ml, infected with UV-irradiated EBV, or not in-fected with EBV for 48 h. Cells were fixed for 10 min in 1% paraformal-dehyde and then permeabilized with 1% saponin (BD Pharmingen). AFACScalibur instrument (Becton, Dickinson) and CellQuest software(Becton, Dickinson) were used for flow cytometric analysis. Monoclonalantibodies (MAbs) were obtained from BD Pharmingen, including anti-CD3-FITC, -CD4-FITC, -CD8-PE, -CD14-FITC, -CD19-PE, -CD21-FITC, -CD68-PE, -CD163-PE, and CD206-PE antibodies. The serum of aNPC patient who was positive for the EBV antibody (1:100) (VCA-IgAtiter of 1:1,280, as determined by an immunofluorescence assay) and Al-exa Fluor 594-conjugated goat anti-human IgG1 antibodies (BD Biosci-ences) were used to detect EBV infection. A rabbit polyclonal anti-IDOantibody (homemade) and goat anti-rabbit IgG1-FITC (Abcam) wereused to detect IDO expression. Negative controls included isotype-matched antibodies (BD Pharmingen) and normal human serum (VCA-IgG negative, as determined by an immunofluorescence assay), which wastreated as a control for detection of EBV infection.
Immunoblotting. Total protein extracts were prepared from MDMcells after treatment for the indicated time intervals. IDO was detectedby using an anti-IDO polyclonal antibody (1:1,000). Mouse monoclo-nal antibody to EBV latent membrane protein 1 (LMP1) (1:1,000;Abcam), BZLF1 antibody (1:2,000; Millipore), and EBV nuclear anti-gen 1 (EBNA1) antibody (1:1,000; Santa Cruz Biotech) were used todetect EBV protein expression in MDMs. An anti--tubulin monoclo-nal antibody (1:5,000; Abcam) was used to confirm equal loading.
To examine the activation of mitogen-activated protein kinase
EBV Induces IDO Expression in Human Macrophages
June 2014 Volume 88 Number 12 jvi.asm.org 6661
on March 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

(MAPK), PI3K, and NF-�B pathways, cell lysates were prepared fromMDMs infected with EBV at 2 � 105 TFU/ml or without EBV infec-tion and probed with specific antibodies against individual compo-nents of these pathways (all antibodies were obtained from Cell Sig-naling). The active forms of the three MAPKs (p38, JNK, and ERK),Akt (a downstream target of PI3K), and the p65 and I�B� subunits ofthe NF-�B complex were detected with rabbit monoclonal antibodiesto phosphorylated p38 (Thr180/Tyr1824), JNK (Thr183/Tyr185), ERK(Thr202Tyr204), Akt (Ser473), I�B� (Ser32), and NF-�B p65 (Ser536).The total amounts of MAPK, Akt, and NF-�B proteins were analyzed withrabbit monoclonal antibodies to JNK, Akt, and NF-�B p65; rabbit poly-clonal antibodies to p38 and ERK; and a mouse monoclonal antibody toI�B�.
Quantitative PCR (qPCR). The total mRNA of the cells was extractedafter treatment for the indicated times. First-strand cDNA synthesis wascarried out by using 500 ng of total RNA. The quantification of target and
reference (glyceraldehyde-3-phosphate dehydrogenase [GAPDH]) geneswas performed in triplicate by using a LightCycler 480 II instrument(Roche, Applied Science). The primers used for real-time PCR were asfollows: forward primer 5=-GGCAAAGGTCATGGAGATGT-3= and re-verse primer 5=-CTGCAGTCTCCATCACGAAA-3= for IDO and forwardprimer 5=-GCACCGTCAAGGCTGAGAAC-3= and reverse primer 5=-TGGTGAAGACGCCAGTGGA-3= for GAPDH.
Measurement of IDO activity. IDO activity was assayed according toa method described previously (17). The cell culture medium was mixedwith trichloroacetic acid and then centrifuged. Next, the supernatant wasinjected into a C18 column and eluted with KH2PO4. The concentrationsof tryptophan and kynurenine were analyzed by high-performance liquidchromatography (HPLC) (Waters). Tryptophan was measured by the de-tection of its native fluorescence at 285-nm excitation and 365-nm emis-sion wavelengths. Kynurenines were detected by UV absorption at the360-nm wavelength in the same chromatographic run, and the results
FIG 1 High IDO expression levels in macrophages from nasopharyngeal carcinoma (NPC) tissue. (A and B) Representative IDO expression in NPC tissuesections detected by immunohistochemistry (magnifications, �200 [A] and �400 [B]). The arrow indicates strong IDO expression in some stromal cells. (C andD) Representative CD68 expression in NPC tissue sections detected by immunohistochemistry (magnifications, �200 [C] and �400 [D]). (E) Representativestaining of IDO (green, FITC), CD68 (red, Alexa Fluor 594), and nuclei (blue, DAPI) in NPC tumor stroma detected by coimmunofluorescence(magnification, �1,000).
Liu et al.
6662 jvi.asm.org Journal of Virology
on March 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

were processed by using Waters Breeze software, version 3.30. IDO activ-ity was determined by calculating the kynurenine-to-tryptophan (Kyn/Trp) ratio (�M/�M).
Cell proliferation. CD4� and CD8� T cell proliferation was assessedby a standard thymidine incorporation assay. Briefly, 1 � 105 cells werecultured in various CMs and stimulated with plate-bound anti-CD3 MAb(OKT3; ATCC) and soluble anti-CD28 MAb (BD Bioscience). After 72 hof culture, 1 �Ci [3H]thymidine was added, and incorporation was mea-sured after 24 h with a -Counter (Wallac).
EBV-specific cytotoxic T lymphocyte (CTL) assays against autologousEBV-transformed lymphoblastoid cell lines (EBV LCLs) were performedas described previously (25). Briefly, 5 � 106 to 10 � 106 PBMCs fromhealthy donors were used for the establishment of an EBV LCL by infec-tion of PBMCs with concentrated supernatants from the B95-8 workingcell banks. EBV LCLs were used as the target cells. To generate EBV-specific CTLs, 2 � 106 PBMCs/ml were cocultured with -irradiated(40 Gy) autologous EBV LCLs. After 3 consecutive restimulations, thecultures were expanded in RPMI 1640 medium containing 20 U/ml re-combinant human IL-2 (rhIL-2) (R&D Systems) and 10% fetal calf serum(FCS) medium. CD8� T lymphocytes, separated from the expanded bulkCTLs by anti-CD8 MAb-precoated Dynabeads (Invitrogen Dynal), wereincubated in different CMs at 37°C for 5 h just before the cell-mediatedcytotoxicity assay was performed and were used as the effector cells. Thecytotoxic activity of CD8� CTLs in different CMs was evaluated in astandard 4-h 51Cr release assay. The effector and target cells were adjustedto effector/T cell (E/T) ratios of 40:1, 20:1, and 10:1. Cytotoxicity wasquantified by the measurement of 51Cr release with a gamma counter(Beckman 9000; Beckman Coulter). The percentage of specific lysis wascalculated as follows: 100 � (total count/min in experimental wells �count/min of spontaneous release)/(count/min of maximum release �count/min of spontaneous release).
Cytokine analysis. The qualification of cytokines in CM was per-formed using a BD Cytometric Bead Array panel kit (BD Biosciences).The analytes included in the 8-plex kit were IL-1, IL-2, IL-4, IL-6, IL-8,IL-10, gamma interferon (IFN-), and TNF-�. The 8 cytokines were mea-sured by flow cytometry according to the manufacturer’s instructions.
Statistical analysis. Paired t tests were used to analyze data, unlessotherwise indicated. We report the nominal P value for each comparisonwithout adjusting for multiple testing. Thus, these results should be inter-preted cautiously due to the possible inflation of type 1 errors. A P valueof �0.05 was considered to be statistically significant.
RESULTSIDO expression in macrophages of NPC tissue. To investigatethe exact expression level of IDO in vivo, IDO protein was detectedby immunohistochemistry of paraffin-embedded archived NPCtissues. High IDO immunoreactivity was observed in NPC tissue(Fig. 1A and B). IDO localization was observed in the cytoplasm ofsome tumor cells. In addition, IDO staining was also observed incertain cells within the tumor stroma, and these cells exhibitedstronger staining than did the cancer cells. To classify these IDO-positive cells in the tumor stroma, CD68 was detected by immu-nohistochemistry of NPC tissues. High CD68 expression levelswere observed at the tumor stroma region (Fig. 1C and D).Furthermore, NPC tissue sections were coimmunostained withanti-IDO and anti-CD68 antibodies and visualized by immuno-fluorescence. As shown in Fig. 1E, in the tumor stroma, someIDO-positive cells were colocalized with CD68-positive cells bydouble staining. IDO-positive cells in the tumor stroma were ob-served in all 23 tissue samples examined. About 71.5% of CD68�
cells expressed IDO protein (range, 21.2% to 89.7%) (Table 1).These assays revealed that some macrophages in the tumor stromaregion of NPC tissue express IDO.
EBV infection induces IDO expression in MDMs. As macro-
phages in NPC tissues express IDO, we investigated whether EBVinfection altered IDO expression in macrophage cells. MDMswere generated from CD14� cells, in which CD3 T cells and CD19B cells were not detectable (Fig. 2A to C). Figure 2D to G show thatthe phenotype of MDMs was CD14high CD163high CD206medium
CD68positive. The level of CD21 expression was very low and wasobserved for only 2.3% of MDMs (Fig. 2H). The lytic infectionprotein BZLF1 and the latent infection proteins EBNA1 andLMP1 were expressed in MDMs infected with EBV, whereas noneof the three proteins were found in MDMs infected with UV-irradiated EBV (UV-EBV) as well as in those without EBV infec-tion (Fig. 2I). IDO expression in MDMs infected with EBV andUV-EBV was assessed by qPCR and Western blot analysis, respec-tively. The IDO mRNA and protein assays showed that IDO can beinduced in MDMs by infection with live EBV but not UV-EBV(Fig. 3A to D). Infection with low-dose EBV (0.01 � 105 TFU) for48 h was able to induce IDO expression. The IDO level was furtherincreased in a time-dependent manner, peaking at 48 h (Fig. 3Aand B), and in a dose-dependent manner, peaking at 0.2 � 105
TFU (Fig. 3C and D). The enzymatic activity of IDO was alsoinvestigated by HPLC. The enzymatic activity of IDO was almostundetectable in the supernatant of MDMs infected with UV-EBVbut was observed in cells infected with EBV (Fig. 3E and F). Theseresults are consistent with the qPCR and Western blotting results.Using an EBV-positive serum (Fig. 3G), about 54% (range, 52.1%to 56.7%) of MDMs were detected to be infected by EBV, and 47%of MDMs showed EBV infection and IDO expression by flow cy-tometric analysis. EBV infection and IDO expression were notdetectable in MDMs infected with UV-irradiated EBV (Fig. 3H).
TABLE 1 Characteristics of nasopharyngeal carcinoma patientsa
CharacteristicValue for nasopharyngealcarcinoma patients (n � 23)
No. (%) of patients of sexFemale 4 (17.4)Male 19 (82.6)
Median age of patients (yr) (range) 47 (19–76)
No. (%) of patients at clinical stageI–II 3 (13.1)III–IV 20 (86.9)
No. (%) of patients with WHO histologicalclassification
NKUC 17 (73.9)NKDC 6 (26.1)
MacrophagesNo. (%) of patients with CD68�
macrophages isolated23 (100)
Median no. of CD68� macrophages/HPF (range)
37 (14–67)
No. (%) of patients with CD68� IDO�
macrophages isolated23 (100)
Median no. of CD68� IDO�
macrophages/HPF (range)25 (6–58)
Median % IDO�/CD68� macrophages(range)
71.5 (21.2–89.7)
a HPF, high-power field; WHO, World Health Organization; NKUC, nonkeratinizingundifferentiated carcinoma; NKDC, nonkeratinizing differentiated carcinoma.
EBV Induces IDO Expression in Human Macrophages
June 2014 Volume 88 Number 12 jvi.asm.org 6663
on March 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

These results confirmed that EBV infection can induce IDO ex-pression in human MDMs.
Involvement of IL-6 and TNF-� in induction of IDO by EBV-MDMs. To investigate the cytokines that are involved in the in-duction of IDO in EBV-MDMs, cytokine concentrations weremeasured by using a cytokine bead array. As shown in Table 2,IFN-, IL-2, IL-4, and IL-12 concentrations in the supernatants ofEBV-MDMs or MDMs infected with UV-irradiated EBV (UV-
EBV-MDM) were below the limits of detection until 72 h. Lowlevels of IL-1 were detected in the supernatants of both EBV-MDM and UV-EBV-MDM cells, but they were not observed to besignificantly different. TNF-� was detected in the supernatant ofEBV-MDMs but was undetectable in control UV-EBV-MDMcells. Levels of IL-6 and IL-10 were significantly increased in thesupernatant of EBV-MDMs compared to the levels in the super-natant of UV-EBV-MDMs (IL-6, 46.3 5.2 versus 3.4 1.2
FIG 2 Purity of CD14 monocytes separated from PBMCs by magnetic CD14 microbeads. (A to C) Expression levels of CD14 (A), CD3 (B), and CD19 (C) weredetermined by flow cytometry. MDMs were generated in the presence of M-CSF. (D to H) Expression levels of CD14 (D), CD68 (E), CD163 (F), CD206 (G), andCD21 (H) were assessed. Gray, tested markers; white, isotype controls. Data are representative of three independent experiments. (I) Representative Westernblots for EBV proteins expressed in EBV-infected MDMs. The expression levels of BZLF, LMP1, and EBNA1 in MDMs infected with EBV at 0.2 � 105 TFU/ml(EBV), infected with UV-irradiated EBV (UV-irradiated EBV), or without EBV infection (RPMI 1640 [1640]) for 48 h were assessed by Western blotting.-Tubulin expression served as the control. Data are representative of three independent experiments, each with cells derived from a different donor.
Liu et al.
6664 jvi.asm.org Journal of Virology
on March 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

pg/ml at 48 h; IL-10, 9.3 1.8 versus 2.3 1.1 pg/ml at 48 h [P �0.001]). TNF-� levels (13.2 2.4 pg/ml) and IL-6 levels (46.3 5.2 pg/ml) reached their peak at 48 h post-EBV infection; the IL-6level was substantially elevated (Fig. 4A and B). Given that TNF-�and IL-6 are potent inducers of IDO expression, we examined thepossible involvement of these two cytokines in IDO expressioninduced by EBV infection using neutralizing antibody assays.Neutralizing antibodies against IL-6 and TNF-� significantlyblocked the increased IDO expression level and enzyme activityinduced by EBV-MDMs, and the combination of both antibodiesalmost completely blocked the induction of IDO protein and itsenzyme activity (Fig. 4C and D). These data indicate that EBV
infection is able to induce several proinflammatory cytokines inMDMs, with the exception of IFN-, and the expression of IL-6and TNF-� mediated by EBV infection is involved in the induc-tion of IDO.
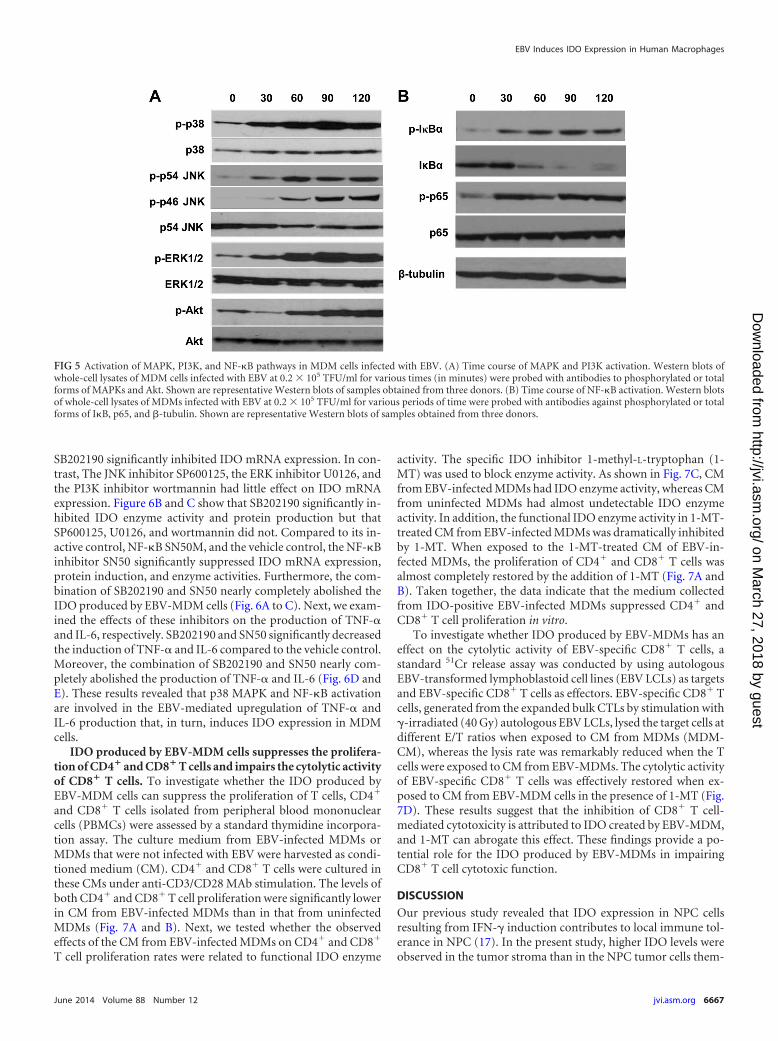
Role of MAPK and NF-�B activation in expression of EBV-induced IDO, IL-6, and TNF-�. Activation of the MAPK, PI3K,and NF-�B signal transduction pathways is critical for IFN--independent induction of IDO. We investigated whether EBV in-fection activates the MAPK (p38, JNK, and ERK), PI3K, andNF-�B pathways in MDMs by Western blot analysis using anti-bodies against the phosphorylated forms of MAPK, Akt, andNF-�B subunits. As shown in Fig. 5, live EBV rapidly induced the
FIG 3 EBV infection induces IDO expression in monocyte-derived macrophages (MDMs). (A) IDO mRNA expression in MDMs infected with UV-irradiatedEBV (UV-EBV) (white) and live EBV (EBV) (black) at 0.2 � 105 TFU/ml for 0 h, 24 h, 48 h, and 72 h, assessed by qPCR. (B) IDO mRNA expression in MDMsinfected with UV-EBV or EBV for 48 h at different dosages, assessed by qPCR. (C) Representative Western blots for IDO protein expression in MDM cells infectedwith UV-EBV or EBV at 0.2 � 105 TFU/ml for the indicated time points. (D) Representative Western blots for IDO protein expression in MDMs infected withUV-EBV or EBV for 48 h at various dosages. (E) Induction of IDO enzymatic activity in the supernatants of MDMs infected with UV-EBV or EBV at 0.2 � 105
TFU/ml for the indicated time points. (F) Induction of IDO enzymatic activity in the supernatants of MDMs infected with UV-EBV or EBV for 48 h at variousdosages. (G) Total proteins of purified EBV virions were analyzed by using a Western blot assay with EBV-positive and -negative human MW, molecular weight.(H) IDO and EBV infection were detected in MDMs infected with EBV or UV-EBV at 0.2 � 105 TFU/ml for 48 h by flow cytometry with rabbit anti-IDOantibodies and EBV-positive human serum. Rabbit IgG1 (Control-FITC) and an EBV-negative human serum (Control-Alexa Fluor 594) were used as controls.Data are representative of three independent experiments. Bars represent the means standard deviations of results from six donors.
EBV Induces IDO Expression in Human Macrophages
June 2014 Volume 88 Number 12 jvi.asm.org 6665
on March 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

phosphorylation of p38, JNK, ERK, and Akt, which generallypeaked at approximately 30 to 90 min postinfection (Fig. 5A). Inresting cells, the p50 and p65 heterodimers of the transcriptionfactor NF-�B are sequestered in the cytoplasm by the I�B protein.NF-�B activation involves the phosphorylation and subsequentdegradation of I�B and the phosphorylation of p65. EBV infection
of MDMs led to an increase in the phosphorylation of I�B� andp65 within 30 min. Synchronously, total I�B� was degraded atbetween 30 and 120 min (Fig. 5B). The requirement of these path-ways for the EBV-induced expression of IDO was also assessed byusing specific pharmacological inhibitors. As shown in Fig. 6A,compared to the DMSO vehicle control, the p38 MAPK inhibitor
TABLE 2 Levels of cytokines in supernatants of MDMs infected with EBV or UV-EBV
Cytokine
Mean cytokine concn (pg/ml) SDa
UV-EBV EBV
0 h 12 h 24 h 48 h 72 h 0 h 12 h 24 h 48 h 72 h
IL-1 ND ND 0.5 0.2 1.2 0.5 0.9 0.3 ND ND 0.4 0.2 0.9 0.3 1.1 0.4IL-2 ND ND ND ND ND ND ND ND ND NDIL-4 ND ND ND ND ND ND ND ND ND NDIL-6 0.3 0.2 1.1 0.4 2.3 0.9 3.4 1.2 6.1 1.7 0.3 0.1 9.1 1.4* 19.3 3.1* 46.3 5.2* 39.3 3.7*IL-10 ND 0.5 0.3 1.1 0.5 2.3 1.1 3.1 1.7 ND 1.6 0.3* 4.1 0.7* 9.3 1.8* 8.3 1.9*IL-12 ND ND ND ND ND ND ND ND ND NDIFN- ND ND ND ND ND ND ND ND ND NDTNF-� ND ND ND ND ND ND 1.9 0.3 3.9 0.4 13.2 2.4 4.2 0.2a UV-EBV, UV-irradiated EBV; ND, not detected. Asterisks indicate a P value of �0.001.
FIG 4 Involvement of IL-6 and TNF-� in induction of IDO by EBV-infected MDMs. (A and B) IL-6 (A) and TNF-� (B) levels in supernatants of EBV- orUV-EBV-infected MDMs at 0.2 � 105 TFU/ml for the indicated times. (C and D) Representative Western blots for IDO and -tubulin in cell lysates (C) and IDOenzymatic activity in supernatants (D) of EBV-infected MDMs at 0.2 � 105 TFU/ml in the presence of neutralizing antibodies to IL-6 or TNF-� or isotype-matched mouse IgG1 (mIgG1) control antibodies for 48 h. Bars represent the means standard deviations of results from triplicate determinations. �, belowthe detection limit; �, P � 0.01; ��, P � 0.001.
Liu et al.
6666 jvi.asm.org Journal of Virology
on March 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
SB202190 significantly inhibited IDO mRNA expression. In con-trast, The JNK inhibitor SP600125, the ERK inhibitor U0126, andthe PI3K inhibitor wortmannin had little effect on IDO mRNAexpression. Figure 6B and C show that SB202190 significantly in-hibited IDO enzyme activity and protein production but thatSP600125, U0126, and wortmannin did not. Compared to its in-active control, NF-�B SN50M, and the vehicle control, the NF-�Binhibitor SN50 significantly suppressed IDO mRNA expression,protein induction, and enzyme activities. Furthermore, the com-bination of SB202190 and SN50 nearly completely abolished theIDO produced by EBV-MDM cells (Fig. 6A to C). Next, we exam-ined the effects of these inhibitors on the production of TNF-�and IL-6, respectively. SB202190 and SN50 significantly decreasedthe induction of TNF-� and IL-6 compared to the vehicle control.Moreover, the combination of SB202190 and SN50 nearly com-pletely abolished the production of TNF-� and IL-6 (Fig. 6D andE). These results revealed that p38 MAPK and NF-�B activationare involved in the EBV-mediated upregulation of TNF-� andIL-6 production that, in turn, induces IDO expression in MDMcells.
IDO produced by EBV-MDM cells suppresses the prolifera-tion of CD4� and CD8� T cells and impairs the cytolytic activityof CD8� T cells. To investigate whether the IDO produced byEBV-MDM cells can suppress the proliferation of T cells, CD4�
and CD8� T cells isolated from peripheral blood mononuclearcells (PBMCs) were assessed by a standard thymidine incorpora-tion assay. The culture medium from EBV-infected MDMs orMDMs that were not infected with EBV were harvested as condi-tioned medium (CM). CD4� and CD8� T cells were cultured inthese CMs under anti-CD3/CD28 MAb stimulation. The levels ofboth CD4� and CD8� T cell proliferation were significantly lowerin CM from EBV-infected MDMs than in that from uninfectedMDMs (Fig. 7A and B). Next, we tested whether the observedeffects of the CM from EBV-infected MDMs on CD4� and CD8�
T cell proliferation rates were related to functional IDO enzyme
activity. The specific IDO inhibitor 1-methyl-L-tryptophan (1-MT) was used to block enzyme activity. As shown in Fig. 7C, CMfrom EBV-infected MDMs had IDO enzyme activity, whereas CMfrom uninfected MDMs had almost undetectable IDO enzymeactivity. In addition, the functional IDO enzyme activity in 1-MT-treated CM from EBV-infected MDMs was dramatically inhibitedby 1-MT. When exposed to the 1-MT-treated CM of EBV-in-fected MDMs, the proliferation of CD4� and CD8� T cells wasalmost completely restored by the addition of 1-MT (Fig. 7A andB). Taken together, the data indicate that the medium collectedfrom IDO-positive EBV-infected MDMs suppressed CD4� andCD8� T cell proliferation in vitro.
To investigate whether IDO produced by EBV-MDMs has aneffect on the cytolytic activity of EBV-specific CD8� T cells, astandard 51Cr release assay was conducted by using autologousEBV-transformed lymphoblastoid cell lines (EBV LCLs) as targetsand EBV-specific CD8� T cells as effectors. EBV-specific CD8� Tcells, generated from the expanded bulk CTLs by stimulation with-irradiated (40 Gy) autologous EBV LCLs, lysed the target cells atdifferent E/T ratios when exposed to CM from MDMs (MDM-CM), whereas the lysis rate was remarkably reduced when the Tcells were exposed to CM from EBV-MDMs. The cytolytic activityof EBV-specific CD8� T cells was effectively restored when ex-posed to CM from EBV-MDM cells in the presence of 1-MT (Fig.7D). These results suggest that the inhibition of CD8� T cell-mediated cytotoxicity is attributed to IDO created by EBV-MDM,and 1-MT can abrogate this effect. These findings provide a po-tential role for the IDO produced by EBV-MDMs in impairingCD8� T cell cytotoxic function.
DISCUSSION
Our previous study revealed that IDO expression in NPC cellsresulting from IFN- induction contributes to local immune tol-erance in NPC (17). In the present study, higher IDO levels wereobserved in the tumor stroma than in the NPC tumor cells them-
FIG 5 Activation of MAPK, PI3K, and NF-�B pathways in MDM cells infected with EBV. (A) Time course of MAPK and PI3K activation. Western blots ofwhole-cell lysates of MDM cells infected with EBV at 0.2 � 105 TFU/ml for various times (in minutes) were probed with antibodies to phosphorylated or totalforms of MAPKs and Akt. Shown are representative Western blots of samples obtained from three donors. (B) Time course of NF-�B activation. Western blotsof whole-cell lysates of MDMs infected with EBV at 0.2 � 105 TFU/ml for various periods of time were probed with antibodies against phosphorylated or totalforms of I�B, p65, and -tubulin. Shown are representative Western blots of samples obtained from three donors.
EBV Induces IDO Expression in Human Macrophages
June 2014 Volume 88 Number 12 jvi.asm.org 6667
on March 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

selves. Given that viral infection can induce IDO expression inmacrophages and that EBV-infected macrophages have been ob-served in NPC tissues, we then asked whether EBV infection ofmacrophages can induce IDO expression. Indeed, we found thatEBV infection does induce IDO expression in MDMs but thatirradiated EBV cannot. These data suggested that EBV infection isa potent inducer of IDO expression in MDMs.
In most cell types, IDO is induced in response to specific in-flammatory stimuli. IFN- represents the principal IDO inducer,and other inflammatory stimuli also induce IDO, includingTNF-�, IL-6, IL-1, and lipopolysaccharide (LPS), althoughmarkedly less than IFN- (26–29). In the present study, TNF-�synergized with IL-6 to mediate IDO induction in EBV-MDMsand did not necessarily require the synthesis of IFN-. In line withour results, several microbial infections of MDMs have been re-ported to upregulate IDO expression in a TNF-�-dependent man-ner but not in an IFN--dependent manner, including HIV, West
Nile virus (WNV), Haemophilus ducreyi, and Listeria monocyto-genes infections (20, 30–32). Synergistic activation of IDO by IL-1, TNF-�, and IL-6 in human monocytic THP-1 cells exposed toLPS was reported previously (28). Recently, IL-6 was shown todirectly upregulate IDO expression in both Neuro-2a cells and anorganotypic hippocampal tissue culture (33). In addition, ourdata indicate that a combination of anti-TNF-� and anti-IL-6antibodies did not completely abolish the IDO production in-duced by EBV infection, supporting the possible involvement ofadditional molecules.
TNF-�-mediated activation of IDO through the MAPK/p38and NF-�B pathways has been reported for microbial infections(29–32, 34). Our data indicate that EBV induces IDO expressionin response to TNF-� and IL-6 stimuli in MDMs dominantlythough the activation of the MAPK/p38 and NF-�B pathways.These results are consistent with previous studies that describedHIV-induced IDO expression in dendritic cells and WNV-in-
FIG 6 Role of MAPK and NF-�B activation in expression of EBV-induced IDO, IL-6, and TNF-�. MDM cells were incubated in medium (1640) (mock) orinfected with EBV in the presence of SB203580 (SB) (p38 inhibitor), SP600125 (SP) (JNK inhibitor), U0126 (ERK inhibitor), the PI3K inhibitor wortmannin(WM), the NF-�B inhibitor (SN50), the NF-�B inactive control (SN50M), and the DMSO vehicle control. (A) Effects of MAPK, PI3K, and NF-�B inhibitors onIDO1 mRNA expression, determined by qPCR. (B) Effects of MAPK, PI3K, and NF-�B inhibitors on IDO enzyme activity in supernatants of EBV-infectedMDMs. (C) Representative Western blots of the effects of MAPK, PI3K, and NF-�B inhibitors on IDO protein expression. (D and E) Effects of MAPK and NF-�Binhibitors on production of IL-6 (D) and TNF-� (E). Culture supernatants from MDMs infected with EBV for 12 or 24 h were assessed for the accumulation ofIL-6 (n � 5) and TNF-� (n � 5). Bars represent the means standard deviations of results from three independent experiments. �, P � 0.001 compared to thevehicle control.
Liu et al.
6668 jvi.asm.org Journal of Virology
on March 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

duced IDO expression in MDMs that predominantly dependedon the activation of the MAPK/p38 and NF-�B pathways (20, 30).It has been confirmed that EBV infection activates the PI3K, JNK,and MAPK/p38 pathways and both the canonical and noncanoni-cal NF-�B signaling pathways (35–37). Activation of the p38, JNK,ERK, and NF-�B pathways leads to the production of IDO-induc-ing cytokines such as TNF-� and IL-6, which act in an autocrinemanner to stimulate the MAPK and NF-�B pathways. The MAPKand NF-�B pathways contribute to IDO induction indirectly bypromoting the production of TNF-� and directly by regulatingIDO gene promoter activity, in which transcriptional factors, suchas AP-1, NF-�B, and NF–IL-6, are known to be activated byTNF-� and IL-6 (28).
IDO expression in APCs and tumor cells can potently inhibitthe immune response (38, 39). It is therefore plausible that activeIDO, expressed in MDMs after EBV infection, acts to create animmunosuppressive microenvironment. This mechanism may beone of the mechanisms by which NPC cells evade the immuneresponse. Our data show that exposure to the microenvironmentcreated by IDO-positive MDM cells severely suppressed bothCD4� and CD8� T cell proliferation. These results favor a model
describing the proliferation arrest of T cells by IDO (39). In addi-tion, we have presented evidence that the exposure to IDO-ex-pressing CM from EBV-MDMs dramatically weakened the cyto-lytic function of CD8� T cells, derived from PBMCs, against targetcells. In accordance with our observations, Liu et al. reported thatin an experimental rat lung allograft model, IDO creates a localmicroenvironment that leads to not only a reduction in the num-bers of CD8� tumor-infiltrating lymphocytes (TILs) but also theloss of the cytotoxic activity of the CD8� effector T cells towardtheir target cells (40). The impaired cytotoxic function observedfor IDO-treated CD8� T cells was accompanied by defects in theproduction of granule cytotoxic proteins, including perforin andgranzymes A and B. Moreover, IDO leads to an impaired bioen-ergetic condition in active CD8� T cells via the selective inhibitionof complex I in the mitochondrial electron transfer chain (40).Our previous study also showed that exposure to the milieu cre-ated by IDO-positive tumor cells, including NPC cells and esoph-ageal tumor cells, significantly impaired lymphocytes against tar-get tumor cells (17, 41). Although IDO was induced by tumor cellsand stromal macrophages in NPC tissue through different mech-anisms, the production of IDO from both of them together con-
FIG 7 Effects of IDO in conditional medium (CM) of EBV-MDMs on proliferation of CD4� T cells and CD8� T cells, and cytolytic activity of CD8� T cells. (Aand B) Representative [3H]thymidine ([3H]TdR) incorporation proliferation assay of CD4� T cells (A) and CD8� T cells (B) derived from PBMCs in CM ofMDM cells infected or not with EBV by coactivation with anti-CD3/CD28 antibodies. (C) Concentrations of kynurenine and tryptophan in the supernatants ofMDMs infected with EBV at 0.2 � 105 TFU/ml (EBV-CM), without EBV infection (CM), or infected with EBV and treated with 1-MT (100 �M) (EBV-CM �1MT) for 48 h, as measured by HPLC. (D) Effects of IDO on cytotoxic activity of EBV-specific CD8 CTLs against target cell EBV LCLs. EBV-specific CD8 CTLswere cultured in the CM of EBV-infected MDMs at 0.2 � 105 TFU/ml (EBV-CM), uninfected MDMs (CM), or MDMs infected with EBV and treated with 1-MT(100 �M) (EBV-CM � 1MT) for 48 h. Cytolytic activity against the target cells was evaluated by using a standard 51Cr release assay. The E/T ratios are indicated.Bars represent the means standard deviations of results from three independent experiments. �, P � 0.001.
EBV Induces IDO Expression in Human Macrophages
June 2014 Volume 88 Number 12 jvi.asm.org 6669
on March 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

tributed to the immune-suppressive microenvironment, whichmight downregulate anti-EBV or antitumor T cell responses, lead-ing to EBV persistence or tumor immune evasion. However, fur-ther studies are needed to elucidate the exact mechanisms bywhich IDO expression in EBV-infected MDMs reduces the cyto-toxicity of CD8� T cells.
Taken together, these findings indicate that TNF-� and IL-6are upregulated by MDMs in response to EBV infection and thatthese two cytokines synergistically mediate the induction of IDOexpression. EBV-induced IDO expression in MDMs is mediatedlargely by the p38/MAPK and NF-�B pathways. The activation ofIDO in response to EBV infection of MDMs could be a key eventin creating an immunosuppressive microenvironment by sup-pressing the proliferation of T cells and impairing the cytotoxicfunction of CD8� T cells. Although the precise role of tumoralIDO in EBV-MDMs remains to be elucidated, our findings sug-gest that blocking IDO activity may provide a means of restoringhost antivirus and antitumour immunity in the treatment of NPC.
ACKNOWLEDGMENTS
This work was supported by the National Natural Science Foundation ofChina (grant no. 30972762) and the 973 Program (grant no.2011CB504800).
REFERENCES1. Tao Q, Young LS, Woodman CB, Murray PG. 2006. Epstein-Barr virus
(EBV) and its associated human cancers— genetics, epigenetics, pathobi-ology and novel therapeutics. Front. Biosci. 11:2672–2713. http://dx.doi.org/10.2741/2000.
2. Xu ZJ, Zheng RS, Zhang SW, Zou XN, Chen WQ. 2013. Nasopharyn-geal carcinoma incidence and mortality in China in 2009. Chin. J. Cancer32:453– 460. http://dx.doi.org/10.5732/cjc.013.10118.
3. Tugizov S, Herrera R, Veluppillai P, Greenspan J, Greenspan D, Palef-sky JM. 2007. Epstein-Barr virus (EBV)-infected monocytes facilitate dis-semination of EBV within the oral mucosal epithelium. J. Virol. 81:5484 –5496. http://dx.doi.org/10.1128/JVI.00171-07.
4. Walling DM, Ray AJ, Nichols JE, Flaitz CM, Nichols CM. 2007. Epstein-Barr virus infection of Langerhans cell precursors as a mechanism of oralepithelial entry, persistence, and reactivation. J. Virol. 81:7249 –7268.http://dx.doi.org/10.1128/JVI.02754-06.
5. Shimakage M, Sakamoto H. 2010. Macrophage involvement in Epstein-Barr virus-related tumors. Exp. Ther. Med. 1:285–291. http://dx.doi.org/10.3892/etm_00000044.
6. Shimakage M, Sakamoto H, Harada S, Sasagawa T, Kodama K. 2007.Expression of the Epstein-Barr virus in lymphoproliferative diseases of thelung. Oncol. Rep. 17:1347–1352. http://dx.doi.org/10.3892/or.17.6.1347.
7. Savard M, Bélanger C, Tremblay MJ, Dumais N, Flamand L, Borgeat P,Gosselin J. 2000. EBV suppresses prostaglandin E2 biosynthesis in humanmonocytes. J. Immunol. 164:6467–6473. http://www.jimmunol.org/content/164/12/6467.long.
8. Savard M, Belanger C, Tardif M, Gourde P, Flamand L, Gosselin J.2000. Infection of primary human monocytes by Epstein-Barr virus. J.Virol. 74:2612–2619. http://dx.doi.org/10.1128/JVI.74.6.2612-2619.2000.
9. Li L, Liu D, Hutt-Fletcher L, Morgan A, Masucci MG, Levitsky V. 2002.Epstein-Barr virus inhibits the development of dendritic cells by promot-ing apoptosis of their monocyte precursors in the presence of granulocytemacrophage-colony-stimulating factor and interleukin-4. Blood 99:3725–3734. http://dx.doi.org/10.1182/blood.V99.10.3725.
10. Salek-Ardakani S, Lyons SA, Arrand JR. 2004. Epstein-Barr virus pro-motes human monocyte survival and maturation through a paracrineinduction of IFN-alpha. J. Immunol. 173:321–331. http://www.jimmunol.org/content/173/1/321.long.
11. Johnson TS, Munn DH. 2012. Host indoleamine 2,3-dioxygenase: con-tribution to systemic acquired tumor tolerance. Immunol. Invest. 41:765–797. http://dx.doi.org/10.3109/08820139.2012.689405.
12. Munn DH, Mellor AL. 2007. Indoleamine 2,3-dioxygenase and tumor-induced tolerance. J. Clin. Invest. 117:1147–1154. http://dx.doi.org/10.1172/JCI31178.
13. Munn DH, Mellor AL. 2003. Macrophages and the regulation of self-reactive T cells. Curr. Pharm. Des. 9:257–264. http://dx.doi.org/10.2174/1381612033392026.
14. Fallarino F, Grohmann U, Vacca C, Orabona C, Spreca A, Fioretti MC,Puccetti P. 2003. T cell apoptosis by kynurenines. Adv. Exp. Med. Biol.527:183–190. http://dx.doi.org/10.1007/978-1-4615-0135-0_21.
15. de Souza Sales J, Lara FA, Amadeu TP, de Oliveira Fulco T, da CostaNery JA, Sampaio EP, Pinheiro RO, Sarno EN. 2011. The role of in-doleamine 2,3-dioxygenase in lepromatous leprosy immunosuppression.Clin. Exp. Immunol. 165:251–263. http://dx.doi.org/10.1111/j.1365-2249.2011.04412.x.
16. Katz JB, Muller AJ, Prendergast GC. 2008. Indoleamine 2,3-dioxygenasein T-cell tolerance and tumoral immune escape. Immunol. Rev. 222:206 –221. http://dx.doi.org/10.1111/j.1600-065X.2008.00610.x.
17. Liu P, Xie BL, Cai SH, He YW, Zhang G, Yi YM, Du J. 2009. Expressionof indoleamine 2,3-dioxygenase in nasopharyngeal carcinoma impairs thecytolytic function of peripheral blood lymphocytes. BMC Cancer 9:416.http://dx.doi.org/10.1186/1471-2407-9-416.
18. Zelante T, Fallarino F, Bistoni F, Puccetti P, Romani L. 2009. Indoleam-ine 2,3-dioxygenase in infection: the paradox of an evasive strategy thatbenefits the host. Microbes Infect. 11:133–141. http://dx.doi.org/10.1016/j.micinf.2008.10.007.
19. Becerra A, Warke RV, Xhaja K, Evans B, Evans J, Martin K, de BoschN, Rothman AL, Bosch I. 2009. Increased activity of indoleamine 2,3-dioxygenase in serum from acutely infected dengue patients linked to interferon antiviral function. J. Gen. Virol. 90:810 – 817. http://dx.doi.org/10.1099/vir.0.004416-0.
20. Boasso A, Herbeuval JP, Hardy AW, Anderson SA, Dolan MJ, Fuchs D,Shearer GM. 2007. HIV inhibits CD4� T-cell proliferation by inducingindoleamine 2,3-dioxygenase in plasmacytoid dendritic cells. Blood 109:3351–3359. http://dx.doi.org/10.1182/blood-2006-07-034785.
21. Heyes MP, Saito K, Jacobowitz D, Markey SP, Takikawa O, Vickers JH.1992. Poliovirus induces indoleamine-2,3-dioxygenase and quinolinicacid synthesis in macaque brain. FASEB J. 6:2977–2989.
22. Larrea E, Riezu-Boj JI, Gil-Guerrero L, Casares N, Aldabe R, Sarobe P,Civeira MP, Heeney JL, Rollier C, Verstrepen B, Wakita T, Borrás-Cuesta F, Lasarte JJ, Prieto J. 2007. Upregulation of indoleamine 2,3-dioxygenase in hepatitis C virus infection. J. Virol. 81:3662–3666. http://dx.doi.org/10.1128/JVI.02248-06.
23. Song H, Park H, Kim J, Park G, Kim YS, Kim SM, Kim D, Seo SK, LeeHK, Cho D, Hur D. 2011. IDO metabolite produced by EBV-transformed B cells inhibits surface expression of NKG2D in NK cells viathe c-Jun N-terminal kinase (JNK) pathway. Immunol. Lett. 136:187–193.http://dx.doi.org/10.1016/j.imlet.2011.01.009.
24. Moss DJ, Pope JH. 1972. Assay of the infectivity of Epstein-Barr virus bytransformation of human leucocytes in vitro. J. Gen. Virol. 17:233–236.http://dx.doi.org/10.1099/0022-1317-17-2-233.
25. Smith CA, Ng CY, Heslop HE, Holladay MS, Richardson S, Turner EV,Loftin SK, Li C, Brenner MK, Rooney CM. 1995. Production of geneticallymodified Epstein-Barr virus-specific cytotoxic T cells for adoptive transfer topatients at high risk of EBV-associated lymphoproliferative disease. J. Hema-tother. 4:73–79. http://dx.doi.org/10.1089/scd.1.1995.4.73.
26. Murakami Y, Hoshi M, Imamura Y, Arioka Y, Yamamoto Y, Saito K.2013. Remarkable role of indoleamine 2,3-dioxygenase and tryptophanmetabolites in infectious diseases: potential role in macrophage-mediatedinflammatory diseases. Mediators Inflamm. 2013:391984. http://dx.doi.org/10.1155/2013/391984.
27. Fujigaki S, Saito K, Sekikawa K, Tone S, Takikawa O, Fujii H, Wada H,Noma A, Seishima M. 2001. Lipopolysaccharide induction of indoleamine2,3-dioxygenase is mediated dominantly by an IFN-gamma-independentmechanism. Eur. J. Immunol. 31:2313–2318. http://dx.doi.org/10.1002/1521-4141(200108)31:8�2313::AID-IMMU2313�3.0.CO;2-S.
28. Fujigaki H, Saito K, Fujigaki S, Takemura M, Sudo K, Ishiguro H,Seishima M. 2006. The signal transducer and activator of transcription1alpha and interferon regulatory factor 1 are not essential for the induc-tion of indoleamine 2,3-dioxygenase by lipopolysaccharide: involvementof p38 mitogen-activated protein kinase and nuclear factor-kappaB path-ways, and synergistic effect of several proinflammatory cytokines. J.Biochem. 139:655– 662. http://dx.doi.org/10.1093/jb/mvj072.
29. Prendergast GC, Chang MY, Mandik-Nayak L, Metz R, Muller AJ.2011. Indoleamine 2,3-dioxygenase as a modifier of pathogenic inflam-mation in cancer and other inflammation-associated diseases. Curr. Med.Chem. 18:2257–2262. http://dx.doi.org/10.2174/092986711795656072.
Liu et al.
6670 jvi.asm.org Journal of Virology
on March 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

30. Yeung AW, Wu W, Freewan M, Stocker R, King NJ, Thomas SR. 2012.Flavivirus infection induces indoleamine 2,3-dioxygenase in humanmonocyte-derived macrophages via tumor necrosis factor and NF-�B. J.Leukoc. Biol. 91:657– 666. http://dx.doi.org/10.1189/jlb.1011532.
31. Li W, Katz BP, Spinola SM. 2011. Haemophilus ducreyi lipooligosac-charides induce expression of the immunosuppressive enzyme indoleam-ine 2,3-dioxygenase via type I interferons and tumor necrosis factor alphain human dendritic cells. Infect. Immun. 79:3338 –3347. http://dx.doi.org/10.1128/IAI.05021-11.
32. Popov A, Abdullah Z, Wickenhauser C, Saric T, Driesen J, Hanisch FG,Domann E, Raven EL, Dehus O, Hermann C, Eggle D, Debey S,Chakraborty T, Krönke M, Utermöhlen O, Schultze JL. 2006. Indoleam-ine 2,3-dioxygenase-expressing dendritic cells form suppurative granulo-mas following Listeria monocytogenes infection. J. Clin. Invest. 116:3160 –3170. http://dx.doi.org/10.1172/JCI28996.
33. Kim H, Chen L, Lim G, Sung B, Wang S, McCabe MF, Rusanescu G,Yang L, Tian Y, Mao J. 2012. Brain indoleamine 2,3-dioxygenase con-tributes to the comorbidity of pain and depression. J. Clin. Invest. 122:2940 –2954. http://dx.doi.org/10.1172/JCI61884.
34. Fu X, Lawson MA, Kelley KW, Dantzer R. 2011. HIV-1 Tat activatesindoleamine 2,3 dioxygenase in murine organotypic hippocampal slicecultures in a p38 mitogen-activated protein kinase-dependent manner. J.Neuroinflammation 8:88. http://dx.doi.org/10.1186/1742-2094-8-88.
35. Vaysberg M, Hatton O, Lambert SL, Snow AL, Wong B, Krams SM,Martinez OM. 2008. Tumor-derived variants of Epstein-Barr virus latent
membrane protein 1 induce sustained Erk activation and c-Fos. J. Biol.Chem. 283:36573–36585. http://dx.doi.org/10.1074/jbc.M802968200.
36. Dawson CW, Laverick L, Morris MA, Tramoutanis G, Young LS. 2008.Epstein-Barr virus-encoded LMP1 regulates epithelial cell motility andinvasion via the ERK-MAPK pathway. J. Virol. 82:3654 –3664. http://dx.doi.org/10.1128/JVI.01888-07.
37. Yoshizaki T, Kondo S, Wakisaka N, Murono S, Endo K, Sugimoto H,Nakanishi S, Tsuji A, Ito M. 2013. Pathogenic role of Epstein-Barr viruslatent membrane protein-1 in the development of nasopharyngeal carci-noma. Cancer Lett. 337:1–7. http://dx.doi.org/10.1016/j.canlet.2013.05.018.
38. Uyttenhove C, Pilotte L, Théate I, Stroobant V, Colau D, Parmentier N,Boon T, Van den Eynde BJ. 2003. Evidence for a tumoral immune resistancemechanism based on tryptophan degradation by indoleamine 2,3-dioxygenase. Nat. Med. 9:1269–1274. http://dx.doi.org/10.1038/nm934.
39. Mellor AL, Keskin DB, Johnson T, Chandler P, Munn DH. 2002. Cellsexpressing indoleamine 2,3-dioxygenase inhibit T cell responses. J. Immu-nol. 168:3771–3776. http://www.jimmunol.org/content/168/8/3771.long.
40. Liu H, Liu L, Liu K, Bizargity P, Hancock WW, Visner GA. 2009.Reduced cytotoxic function of effector CD8� T cells is responsible forindoleamine 2,3-dioxygenase-dependent immune suppression. J. Immu-nol. 183:1022–1031. http://dx.doi.org/10.4049/jimmunol.0900408.
41. Zhang G, Liu WL, Zhang L, Wang JY, Kuang MH, Liu P, Lin YH, DaiSQ, Du J. 2011. Involvement of indoleamine 2,3-dioxygenase in impair-ing tumor-infiltrating CD8 T-cell functions in esophageal squamous cellcarcinoma. Clin. Dev. Immunol. 2011:384726. http://dx.doi.org/10.1155/2011/384726.
EBV Induces IDO Expression in Human Macrophages
June 2014 Volume 88 Number 12 jvi.asm.org 6671
on March 27, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from