evolution of the woody growth habit - land colonization...
TRANSCRIPT

Evolution of the Woody Growth Habit -Land Colonization and Adaptation
• Trees or tree forms plants are classified into
• Ferns
• Gymnosperms
• Angiosperms
• Plants apparently first emerged on land from their forerunner algal ancestors during the mid-Ordovician period, some 460 Mya.
• Remarkable evolution changes: generation of specialized cell types, such as lignified (reinforced) secondary cell walls and vascular tissues; formation of protective wood and bark tissues, and other specialized cell types within; the ability to continuously orient/reorient massive photosynthetic canopies; elaboration of a plethora of often species-specific distinct biochemical pathways leading to, for example, chemical defense systems; evolution of distinct plant pollination/reproductive strategies and adaptations, and a myriad of related regulatory processes at the genomic/proteomic and metabolic levels.
Early phases of Land Colonization-Turgor-Based Stem Support Systems
• The earliest terrestrial plants had features similar to today’s bryophytes (mosses, liverworts, and hornworts), and were thus nonwoody.
• Small in stature
• Transporting water by hydroid cell types
• Covered by cuticle layers
• Significant changes in initial cell wall properties and assemblies of these early plant forms then followed, including that of secondary wall formation/ thickening in the late Silurian period around 415 Mya.
Cooksonia sp.several cm in height
Turgor-based stem support system:e.g., Rhynia sp.
cpx
Stem cross-section
13–22 cmin height
Turgor-based stem support system:Helical thickenings
Modern tracheid withhelical secondary
thickenings (internalcut-away view)
Douglas fir(Pseudotsuga menziesii)
Self-supporting stem system:Helical thickenings
Self-supporting stem system:e.g., Psilophyton sp.
Stem cross-section
75–200 cmin height
xp
c
s
1 µm
Cooksonia pertonii tracheid
10 µm
r
t
pw
ht
(b)(a)
(d)(c)
Figure 13 Early and late tracheophyte developments that ultimately lead to woody tissue. These progressions also reflectthe biophysical/mechanical properties of the polymeric constituents conferred within the tracheophyte cell wall composite(s).Cooksonia spp. were among the earliest tracheophytes, whose tracheids had decay-resistant helical secondary cell wallthickenings (red arrow) with anatomical structures that allowed for improved overall water transport (a). Extant tracheids, suchas in Douglas fir (P. menziesii) wood, are variations on the basic theme of a primary wall (pw) (i.e., dark background) withinternal helical secondary cell wall thickenings (ht) for structural reinforcement (lighter raised surfaces) (b). Mechanical stemstrength of early tracheophytes diversified from a system dependent on turgor pressure as in Rhynia (c) to a systemcomplemented by a hypodermal sterome fortified by sclerenchyma cells (s) with thick secondary cell walls as in Psilophyton(d). Circle diagrams indicate stem structure in cross-section. Abbreviations: c, cortical parenchyma; ht, helical thickening; p,phloem; pw, primary wall; r, ray parenchyma; s, sterome; t, tracheid lumen; x, xylem/tracheids. Reproduced from Le Mondedes Vegetaux, copyright 1997 (http://www.creaweb,fr/bv/banque/cormo-cooksonia.html) and with permission of WileyInter Science from D. Edwards, New Phytol. 1993, 125, 225–247, copyright 1993 (a). Reproduced with permission ofSyracuse University Press from H. A. Core; W. A. Cote; A. C. Day, Wood Structure and Identification, 2nd ed.; SyracuseUniversity Press: Syracuse, 1979, copyright 1979, with whole tree photograph from L. B. Davin (b). Reproduced withpermission of Elsevier fromN. Rowe; T. Speck, Hydraulics andMechanics of Plants: Novelty, Innovation and Evolution. In TheEvolution of Plant Physiology: FromWhole Plants to Ecosystems; A. R. Hemsley; I. Poole, Eds.; Elsevier: Amsterdam, 2004; pp297–325, copyright 2004; (c, d).
Trees: A Remarkable Biochemical Bounty 1185
Early and late tracheophyte developments that ultimately lead to woody tissue. These progressions also reflect the biophysical/mechanical properties of the polymeric constituents conferred within the tracheophyte cell wall composite(s). Cooksonia spp. were among the earliest tracheophytes, whose tracheids had decay-resistant helical secondary cell wall thickenings (red arrow) with anatomical structures that allowed for improved overall water transport
Turgor-based stem support system: Helical thickenings

Early phases of Land Colonization-Turgor-Based Stem Support Systems
• Following Cooksonia emergence, however, the early plant tracheophytes subsequently diversified, resulting in numerous independent early lineages, as evidenced by the fossil records of Rhynia Sennicaulis, and Gossilingia.
• Within these plant groups, the tracheid secondary cell walls thus essentially became thicker with control of lateral water movement being improved throughout via defined lateral cell wall pits.
Turgor-based stem support system:Rhynia sp.
Cooksonia sp.several cm in height
Turgor-based stem support system:e.g., Rhynia sp.
cpx
Stem cross-section
13–22 cmin height
Turgor-based stem support system:Helical thickenings
Modern tracheid withhelical secondary
thickenings (internalcut-away view)
Douglas fir(Pseudotsuga menziesii)
Self-supporting stem system:Helical thickenings
Self-supporting stem system:e.g., Psilophyton sp.
Stem cross-section
75–200 cmin height
xp
c
s
1 µm
Cooksonia pertonii tracheid
10 µm
r
t
pw
ht
(b)(a)
(d)(c)
Figure 13 Early and late tracheophyte developments that ultimately lead to woody tissue. These progressions also reflectthe biophysical/mechanical properties of the polymeric constituents conferred within the tracheophyte cell wall composite(s).Cooksonia spp. were among the earliest tracheophytes, whose tracheids had decay-resistant helical secondary cell wallthickenings (red arrow) with anatomical structures that allowed for improved overall water transport (a). Extant tracheids, suchas in Douglas fir (P. menziesii) wood, are variations on the basic theme of a primary wall (pw) (i.e., dark background) withinternal helical secondary cell wall thickenings (ht) for structural reinforcement (lighter raised surfaces) (b). Mechanical stemstrength of early tracheophytes diversified from a system dependent on turgor pressure as in Rhynia (c) to a systemcomplemented by a hypodermal sterome fortified by sclerenchyma cells (s) with thick secondary cell walls as in Psilophyton(d). Circle diagrams indicate stem structure in cross-section. Abbreviations: c, cortical parenchyma; ht, helical thickening; p,phloem; pw, primary wall; r, ray parenchyma; s, sterome; t, tracheid lumen; x, xylem/tracheids. Reproduced from Le Mondedes Vegetaux, copyright 1997 (http://www.creaweb,fr/bv/banque/cormo-cooksonia.html) and with permission of WileyInter Science from D. Edwards, New Phytol. 1993, 125, 225–247, copyright 1993 (a). Reproduced with permission ofSyracuse University Press from H. A. Core; W. A. Cote; A. C. Day, Wood Structure and Identification, 2nd ed.; SyracuseUniversity Press: Syracuse, 1979, copyright 1979, with whole tree photograph from L. B. Davin (b). Reproduced withpermission of Elsevier fromN. Rowe; T. Speck, Hydraulics andMechanics of Plants: Novelty, Innovation and Evolution. In TheEvolution of Plant Physiology: FromWhole Plants to Ecosystems; A. R. Hemsley; I. Poole, Eds.; Elsevier: Amsterdam, 2004; pp297–325, copyright 2004; (c, d).
Trees: A Remarkable Biochemical Bounty 1185
Self-supporting stem system:Psiophyton sp.
Cooksonia sp.several cm in height
Turgor-based stem support system:e.g., Rhynia sp.
cpx
Stem cross-section
13–22 cmin height
Turgor-based stem support system:Helical thickenings
Modern tracheid withhelical secondary
thickenings (internalcut-away view)
Douglas fir(Pseudotsuga menziesii)
Self-supporting stem system:Helical thickenings
Self-supporting stem system:e.g., Psilophyton sp.
Stem cross-section
75–200 cmin height
xp
c
s
1 µm
Cooksonia pertonii tracheid
10 µm
r
t
pw
ht
(b)(a)
(d)(c)
Figure 13 Early and late tracheophyte developments that ultimately lead to woody tissue. These progressions also reflectthe biophysical/mechanical properties of the polymeric constituents conferred within the tracheophyte cell wall composite(s).Cooksonia spp. were among the earliest tracheophytes, whose tracheids had decay-resistant helical secondary cell wallthickenings (red arrow) with anatomical structures that allowed for improved overall water transport (a). Extant tracheids, suchas in Douglas fir (P. menziesii) wood, are variations on the basic theme of a primary wall (pw) (i.e., dark background) withinternal helical secondary cell wall thickenings (ht) for structural reinforcement (lighter raised surfaces) (b). Mechanical stemstrength of early tracheophytes diversified from a system dependent on turgor pressure as in Rhynia (c) to a systemcomplemented by a hypodermal sterome fortified by sclerenchyma cells (s) with thick secondary cell walls as in Psilophyton(d). Circle diagrams indicate stem structure in cross-section. Abbreviations: c, cortical parenchyma; ht, helical thickening; p,phloem; pw, primary wall; r, ray parenchyma; s, sterome; t, tracheid lumen; x, xylem/tracheids. Reproduced from Le Mondedes Vegetaux, copyright 1997 (http://www.creaweb,fr/bv/banque/cormo-cooksonia.html) and with permission of WileyInter Science from D. Edwards, New Phytol. 1993, 125, 225–247, copyright 1993 (a). Reproduced with permission ofSyracuse University Press from H. A. Core; W. A. Cote; A. C. Day, Wood Structure and Identification, 2nd ed.; SyracuseUniversity Press: Syracuse, 1979, copyright 1979, with whole tree photograph from L. B. Davin (b). Reproduced withpermission of Elsevier fromN. Rowe; T. Speck, Hydraulics andMechanics of Plants: Novelty, Innovation and Evolution. In TheEvolution of Plant Physiology: FromWhole Plants to Ecosystems; A. R. Hemsley; I. Poole, Eds.; Elsevier: Amsterdam, 2004; pp297–325, copyright 2004; (c, d).
Trees: A Remarkable Biochemical Bounty 1185
Mechanical stem strength of early
tracheophytes diversified from a
system dependent on turgor pressure
as in Rhynia to a system
complemented by a hypodermal
sterome fortified by sclerenchyma cells
(s) with thick secondary cell walls as in
Psilophyton.
Early phases of Land Colonization-Turgor-Based Stem Support Systems
• Present-day plants. For example, Douglas fir (Pseudotsuga menziesii) has helical-shaped secondary thickenings in its tracheids (ht) partially covering the internal surfaces of its primary cell walls.
• Such modifications, in turn, resulted in changes in plant cell wall biopolymer composition and organization, thereby facilitating their functional properties, for example, in terms of being better able to stand upright and for water conduction, and so forth.
Self-supporting stem support system:Helical thickenings
Cooksonia sp.several cm in height
Turgor-based stem support system:e.g., Rhynia sp.
cpx
Stem cross-section
13–22 cmin height
Turgor-based stem support system:Helical thickenings
Modern tracheid withhelical secondary
thickenings (internalcut-away view)
Douglas fir(Pseudotsuga menziesii)
Self-supporting stem system:Helical thickenings
Self-supporting stem system:e.g., Psilophyton sp.
Stem cross-section
75–200 cmin height
xp
c
s
1 µm
Cooksonia pertonii tracheid
10 µm
r
t
pw
ht
(b)(a)
(d)(c)
Figure 13 Early and late tracheophyte developments that ultimately lead to woody tissue. These progressions also reflectthe biophysical/mechanical properties of the polymeric constituents conferred within the tracheophyte cell wall composite(s).Cooksonia spp. were among the earliest tracheophytes, whose tracheids had decay-resistant helical secondary cell wallthickenings (red arrow) with anatomical structures that allowed for improved overall water transport (a). Extant tracheids, suchas in Douglas fir (P. menziesii) wood, are variations on the basic theme of a primary wall (pw) (i.e., dark background) withinternal helical secondary cell wall thickenings (ht) for structural reinforcement (lighter raised surfaces) (b). Mechanical stemstrength of early tracheophytes diversified from a system dependent on turgor pressure as in Rhynia (c) to a systemcomplemented by a hypodermal sterome fortified by sclerenchyma cells (s) with thick secondary cell walls as in Psilophyton(d). Circle diagrams indicate stem structure in cross-section. Abbreviations: c, cortical parenchyma; ht, helical thickening; p,phloem; pw, primary wall; r, ray parenchyma; s, sterome; t, tracheid lumen; x, xylem/tracheids. Reproduced from Le Mondedes Vegetaux, copyright 1997 (http://www.creaweb,fr/bv/banque/cormo-cooksonia.html) and with permission of WileyInter Science from D. Edwards, New Phytol. 1993, 125, 225–247, copyright 1993 (a). Reproduced with permission ofSyracuse University Press from H. A. Core; W. A. Cote; A. C. Day, Wood Structure and Identification, 2nd ed.; SyracuseUniversity Press: Syracuse, 1979, copyright 1979, with whole tree photograph from L. B. Davin (b). Reproduced withpermission of Elsevier fromN. Rowe; T. Speck, Hydraulics andMechanics of Plants: Novelty, Innovation and Evolution. In TheEvolution of Plant Physiology: FromWhole Plants to Ecosystems; A. R. Hemsley; I. Poole, Eds.; Elsevier: Amsterdam, 2004; pp297–325, copyright 2004; (c, d).
Trees: A Remarkable Biochemical Bounty 1185
Extant tracheids, such as in Douglas fir (P. menziesii) wood, are
variations on the basic theme of a primary wall (pw) (i.e., dark
background) with internal helical secondary cell wall thickenings (ht)
for structural reinforcement (lighter raised surfaces).
• Presence of phenolic-based components, including the structural cell-wall polymers, lignins, to account for the apparent decay-resistant nature of their secondary wall thickenings.
• However, it is yet unclear as to when lignins proper arose.• Early tracheid-containing organisms had enhanced resistance to
implosion that helped maintain turgor pressure, and a means of improved water delivery.

Stem-Thickening Systems- Further Adaptations
• To diversify into drier environments and compete for sunlight and gamete dispersal, some plant forms shifted from mainly having a turgor-based stem strength to a more self-supporting system.
• by the mid-Devonian period, around 390 Mya, several independent lineages had begun to achieve ‘self-support’ by development of a hypodermal sterome at the periphery of the stem.This consisted of rows of cells with very thick, decay-resistant, secondary cell walls called sclerenchyma.
• Continued evolution and diversification of secondary cell walls within water-conducting and structurally reinforcing sclerenchyma cells later improved water transport and stem support, respectively.
• Several other stem-thickening strategies subsequently arose, including the development of specialized primary thickening as in the extant tree ferns, numerous monocot angiosperms, and cycads. This was also achieved through independent emergence of unifacial and bifacial secondary growth in several very different taxa.
• Of these adaptations resulting in tree forms, however, only bifacial secondary growth succeeded to the extent of giving rise to the diversity of woody species in existence that we so greatly value today.
Several other stem-thickening strategies subsequently arose, including the development of specializedprimary thickening (Figure 14(a)) as in the extant tree ferns, numerous monocot angiosperms, and cycads.23,24
This was also achieved through independent emergence of unifacial (Figure 15(a)) and bifacial (Figure 16(a))secondary growth in several very different taxa.25 Of these adaptations resulting in tree forms, however, onlybifacial secondary growth succeeded to the extent of giving rise to the diversity of woody species in existencethat we so greatly value today.
The earliest tree form with a perennial trunk emerged during the mid-Devonian period, 397–385Mya. Thiswas the now extinct Wattieza (Figure 14(c)), a giant !8–16m cladoxylopsid, which represents a lineage thatwas ancestral to major lineages, such as Pteridophyta (i.e., the ferns) and Sphenophyta (the horsetails).26 Unlikepresent-day trees, Wattieza apparently lacked a taproot, permanent branches, and secondary growth.Nevertheless, using a primary thickening strategy (see below), it successfully achieved arborescence andcomprised the earliest known forest.26
1 m
Root mass
PithXylem/phloem
Leaf traces
Sclerenchyma
Xylem/phloem
Pith
Tree fern trunk
SAMPTMBases of
leaves
Leafbases
Root mass
Primary thickening meristem (PTM)in monocot angiosperms
Extinct tree fern:Wattieza
Extant tree fern Extent palm(angiosperm, monocot)
(b)(a)
(e)(d)(c)
Figure 14 Primary stem-thickening systems. Stem thickening by primary tissues evolved independently in several taxa,including some extinct lineages, ferns, andmonocot angiosperms. The primary thickening meristem (PTM) is derived from theshoot apical meristem (SAM), where the PTM produces leaves that add to stem thickness (a). Mechanical strength in treeferns is derived from placement of xylem and sclerenchyma to the stem periphery, and accumulation of leaf bases along theentire stem and root growth (sometimes from the leaf bases) to form a root mass along the lower portion of the stem (b). Thecladoxylopsid, Wattiezia, preceded both ferns and horsetails and reached at least 8m in height (c). In extant tree ferns, oldleaves abscise (white arrows) leaving behind prominent leaf scars (yellow arrowheads), with the remnant leaf basescontributing to primary thickening. Some present-day tree ferns (d) can reach !20m in height (not shown). Tall extantmonocots, such as palms, have similar primary thickening systems (e). Reproduced with permission of Botanical Society ofAmerica from D. A. DeMason, Am. J. Bot. 1983, 70, 955–962, copyright 1983 (a). Reproduced with permission of M. F. Largefrom M. F. Large; J. E. Braggins, Eds.; Tree Ferns; Timber Press: Portland, 2004, copyright 2004 (b). Reproduced withpermission of Nature Publishing Group (a division of Macmillan Publishers Ltd.) from W. E. Stein; F. Mannolini; L. V. Hernick;E. Landing; C. M. Berry, Nature 2007, 446, 904–907, copyright 2007 (c). Photos from M. J. Kato, Instituto de Quimica,Universidade de Sao Paulo, Sao Paulo, Brazil (d) and D. G. Vassao, Washington State University (e).
1186 Trees: A Remarkable Biochemical BountyPrimary stem-thickening systems
Stem thickening by primary tissues evolved independently in several taxa, including some extinct lineages, ferns, and monocot angiosperms. The primary thickening meristem (PTM) is derived from the shoot apical meristem (SAM), where the PTM produces leaves that add to stem thickness (a).
Mechanical strength in tree ferns is derived
from placement of xylem and sclerenchyma
to the stem periphery, and accumulation of
leaf bases along the entire stem and root
growth (sometimes from the leaf bases) to
form a root mass along the lower portion of
the stem (b). The cladoxylopsid, Wattiezia,
preceded both ferns and horsetails and
reached at least 8 m in height (c). In extant
tree ferns, old leaves abscise (white arrows)
leaving behind prominent leaf scars (yellow
arrowheads), with the remnant leaf bases
contributing to primary thickening. Some
present-day tree ferns (d) can reach around
20 m in height (not shown). Tall extant
monocots, such as palms, have similar
primary thickening systems (e).

LETTERS
Giant cladoxylopsid trees resolve the enigma of theEarth’s earliest forest stumps at GilboaWilliam E. Stein1, Frank Mannolini2, Linda VanAller Hernick2, Ed Landing2 & Christopher M. Berry3
The evolution of trees of modern size growing together in forestsfundamentally changed terrestrial ecosystems1–3. The oldest treesare often thought to be of latest Devonian age (about 380–360Myrold) as indicated by the widespread occurrence of Archaeopteris(Progymnospermopsida)4. Late Middle Devonian fossil treestumps, rooted and still in life position, discovered in the 1870sfrom Gilboa, New York5, and later named Eospermatopteris, arewidely cited as evidence of the Earth’s ‘oldest forest’6,7. However,their affinities and significance have proved to be elusive becausetheaerial portionof theplanthas beenunknownuntil now.Herewereport spectacular specimens from Schoharie County, New York,showing an intact crown belonging to the cladoxylopsid Wattieza(Pseudosporochnales)8 and its attachment to Eospermatopteristrunk and base. This evidence allows the reconstruction of a tall(at least 8m), tree-fern-like plant with a trunk bearing large bran-ches in longitudinal ranks. The branches were probably abscisedas frond-likemodules. Lower portions of the trunk show longitud-inal carbonaceous strands typical of Eospermatopteris, and a flatbottom with many small anchoring roots. These specimens pro-vide new insight into Earth’s earliest trees and forest ecosys-tems. The tree-fern-like morphology described here is the oldestexample so far of an evolutionarily recurrent arborescent bodyplan within vascular plants. Given their modular construction,these plants probably produced abundant litter, indicating thepotential for significant terrestrial carbon accumulation and adetritus-based arthropod fauna by the Middle Devonian period.
Although the fossil record provides evidence of increasing size andcomplexity of plants through the Devonian, little is known abouthow the origin of tree-sized individuals changed the evolutionarydynamics of terrestrial ecosystems. The famous ‘earliest forest’ occur-rence of upright and rooted sandstone casts called Eospermatopterisfound at three stratigraphic levels (Middle Devonian, upper Givetianto about theMiddle/Upper Devonian boundary, about 385Myr ago)at Gilboa, New York, displays typical difficulties encountered inmaking morphological and ecological interpretations. Each cast,invariably broken 50–150 cm above the base, is typically 50–100 cmin circumference, flaring proximally to as much as 330 cm at thebase. A distinct pattern of longitudinally oriented carbonaceousstrands occurs on the trunk, often showing proximal anastomosis.Eospermatopteris was initially thought to be associated with isolatedbranch systems and was reconstructed as a pteridosperm (early seedplant)9. However, subsequent work has provided little support forthis reconstruction10. Thus, evidence so far allows wide latitude inphylogenetic and ecological interpretations of the plant, includinganeurophytalean progymnosperm11, lepidosigillarioid lycopsid1,12 orcladoxylopsid13.
Among these possibilities, it is becoming increasingly evident thatmembers of the class Cladoxylopsida, ranging at least from the early
Middle Devonian (Eifelian) into the Carboniferous, were majorcontributors to floras worldwide14. Traditionally considered inter-mediate between Lower Devonian vascular plants and ferns orsphenopsids, we do not yet understand these plants well enough toreconstruct their overall architecture or phylogeny accurately. Recentwork indicates that the group may have had considerable morpho-logical and anatomical diversity. Several forms were probably quitelarge and some had significant secondary tissues15–19. WithinCladoxylopsida, the order Pseudosporochnales, represented espe-cially by the genus Pseudosporochnus from Goe, Belgium (latestEifelian), are among the best-studied20,21. Compressions are com-monly found of digitately branched systems bearing sterile andfertile non-laminar appendages, all having a highly distinctive‘speckled’ pattern caused by sclereid nests in the cortex. On thebasis of the association of typical branches with trunk fragments,Pseudosporochnus has been reconstructed as a small tree22. How-ever, only one specimen shows direct insertion of a single fragment-ary branch base on the trunk. Well-defined surfaces at the proximalend of branches clearly match the regular pattern of attachment scarson the trunks. From this, regular abscission of the branches from thetrunk has been inferred22. The genus Wattieza from Belgium andVenezuela shows a similar construction of branch systems, includingprobable basal abscission surfaces and speckled texture, but bearsmore complex appendages, some with recurved tips bearing spor-angia8. However, the trunk remains unknown.
The material described here is derived from a New York StateDepartment of Environmental Conservation quarry on the north-west slope of South Mountain, Schoharie County, New York(42u 239N, 74u 169W). This site is located about 13 km east of thehighest in situGilboa stump horizon atManorkill Falls7 and lies closeto inferred palaeo-shoreline23. Long-term collecting at this localityhas produced a diverse flora24.
The quarry exposes 7.5m of rock in the Oneonta Formation, thelowest formation of the eastern terrestrial facies of the GeneseeGroup. Although the dating of the terrestrial sediments remains im-precise, palynostratigraphy of nearby samples yields latest Givetianto earliest Frasnian age25. The lower 3.3-m exposure includes terrest-rial red/brown siltshales, sandstones with root structures, and severalsoil horizons. The highest soil horizon is overlain by a sandstone unitwith west-dipping foresets (3.3–6.0m), which is consistent with asmall fluvial delta. Abundant plant debris occurs low in these foresets,including shoot systems and logs ranging from a few centimetresto several metres in length. The logs mostly lie in near north–southorientation perpendicular to the palaeocurrent direction and parallelwith small wave ripples. Some specimens show attachment of finesterile or fertile appendages to branches. These data indicate minimaltransport and rapid deposition. We describe two exceptionally com-plete specimens here.
1Department of Biological Sciences, Binghamton University, Binghamton, New York 13902-6000, USA. 2New York State Museum, Albany, New York 12230, USA. 3School of Earth,Ocean and Planetary Sciences, Cardiff University, Cardiff CF10 3YE, UK.
Vol 446 | 19 April 2007 |doi:10.1038/nature05705
904Nature ©2007 Publishing Group
The first (Fig. 1a, b) represents the only tree-sized member of thePseudosporochnales so far discovered with substantial connection ofthe aerial portions of the plant. More than 2m of trunk was found inthe field, of which the upper 130 cmhas been recoveredmostly intact.The trunk ranges in width from 15 cm at the top, within a well-defined crown region, to 18 cm at the recovered specimen’s proximalend. At least eight digitate branches are attached in the crown, butseveral more were probably attached both above and below theexposed surface of the compression. The branches bear sterile andfertile appendages (Fig. 1c). The appendages have a central axisand whorled dichotomous ultimate units. Fertile appendages bearrecurved tips bearing sporangia. These features allow identificationtoWattieza Stockmans8. Attached branches observed in face-on andedge-on views seem to have been bilaterally symmetrical, giving theplant an overall appearance similar to that of amodern tree-fern withlarge fronds. Within the zone of obvious attachment, the branchesoccur in orthostichous ranks, and a regular pattern of branch scarscontinues down the entire preserved length of trunk (Fig. 1b). Justbelow the crown, isolated branches occur in the matrix along theright side of the trunk aligned as if still attached. This suggests anextensive, mostly ascendant, set of branches borne by the plant at thetime of death. Further down, the trunk shows no evidence of attachedsenescent branches, and branch scars become progressively less dis-tinct. The latter suggests the remodelling of outer tissues of the trunkafter branch fall. The trunk was found within and slightly abovebedding planes showing detached and randomly arranged branches.Detached branches match attached branches in morphology and sizeand thus potentially comprise litter derived from this individual.Some detached branches show likely abscission surfaces generallycomparable to those described previously in Pseudosporochnus andWattieza22.
The second specimen, a large trunk (Fig. 2a), was found about 2maway and seems to be mostly proximal to the first specimen, but withsome overlap. This specimen is a flattened internal cast with both
upper and lower counterparts showing surface features. The trunkis more than 6m in length, with a width of 13 cm at the top andexpanding to 47 cm near the base. The top is truncated but shows anidentical pattern of branch scars and speckled surface to thatobserved about 75 cm below the crown in the first specimen(Fig. 2a, 3a). Below this level, the trunk shows diminution of branchattachment features, as in the first specimen, replaced by a system oflongitudinal coalified strands. The coalified strands are distally nar-row and closely spaced, becoming larger and more separate towardsthe base. Near its proximal end, about 70 cm from the base of thespecimen (Fig. 2a, 3b), the trunk swells to form a flattened cast (8–10 cm thick), with a change in the pattern of the longitudinal strandsindicating anastomosis. Combined evidence leaves little doubt aboutassignment of the specimen to Eospermatopteris. Unlike the uprightEospermatopteris stumps at Gilboa, however, this trunk is preservedalong a bedding plane and is radially compressed. We think thisdifference very probably explains the absence of a bulbous base inthis specimen as opposed to that observed in some, but not all, Gilboastumps. Below the interpreted flat bottom of the trunk, many coali-fied strands extend somewhat sinuously into the matrix, suggestingattached roots.
The union ofWattieza branches with a trunk and base identical tothose of Eospermatopteris provides the first direct evidence of generalbody form including attachment for both genera, and allows newinsight into the enigmatic Gilboa trees. This evidence also providesthe first reasonably complete picture of the size and architecture oflarge pseudosporochnalean cladoxylopsids (Fig. 2b). Evidence fromthe two specimens combined indicates that the height of the tree mayhave exceeded 8m. The basal diameter of our specimen falls withinthe range of stump sizes observed at Gilboa. However, the largeststumps at Gilboa are twice the diameter of ours, suggesting evengreater height.
Cladoxylopsids have long been interpreted as intermediatesbetween the earliest vascular plants and living pteridophytes. It is
a b c
Figure 1 | Wattieza Stockmans from South Mountain, New York, NYSM17039. a, General view of the crown portion, showing longitudinal ranks ofbranch bases on the trunk proximally, and attached branches with digitateramification and speckled surface pattern distally. Scale bar, 20 cm. b, Line
drawing of the specimen as recovered including trunk and crown; the boxshows the portion in a, and the arrow indicates the branch in c. Scale bar,10 cm. c, Close-up of a distal branch showing speckled texture and lateralappendages. Scale bar, 20mm.
NATURE |Vol 446 | 19 April 2007 LETTERS
905Nature ©2007 Publishing Group
therefore interesting to see how instantly recognizable and, in a sig-nificant sense, ‘modern’ the tree-like architecture of Wattieza seemsto be. Indeed, phylogenetically divergent modern forms includingtree-ferns, cycads and palms are fundamentally similar in structure.This body plan now stands unequivocally as the oldest known arbor-escent terrestrial plant form, with trunks of similar diameter (at least13 cm) showing attachment scars known from the lower MiddleDevonian (Goe, Eifelian)22. TheWattieza body plan contrasts sharplywith more diffusely branched Archaeopteris, conifers and most dico-tyledonous angiosperm trees.
In addition, it is evident that this early tree architecture is notmerely primitive but instead has evolved recurrently within vascularplants. It is perhaps significant that arborescence in Wattieza isassociated with trunk bases lacking taproots, seeming instead to beanchored by many roots of nearly equal size. We recognize this syn-drome in the modern analogues, where an extensive root mantle insome contributes significantly to their self-supporting habit. Onepreviously described cladoxylopsid is known to exhibit a rootmantle,although in this instance the plant is apparently not arborescent26. Itremains to be determined whether Eospermatopteris also has a rootmantle at the base. Perhaps this morphology represents a wetlandecological specialization distinguishing pseudosporochnalean cla-doxylopsids of Gilboa from riparian conifer-like Archaeopteris7. Sofar, however, neither kind of root has been adequately circumscribedas a morphological, ecological or biogeochemical entity. Caution is
a b
Figure 3 | Trunk top and base of Wattieza Stockmans from SouthMountain, New York; details of the large trunk specimen, NYSM 17040.a, Counterpart of the distal portion of the trunk indicated by arrows inFig. 2a, having speckled texture and prominent branch scars. Scale bar,10 cm. b, Lower counterpart of the proximal cast region of trunk indicatedby arrows in Fig. 2a, with the cast removed. The specimen shows longitudinalcarbonaceous strands that anastomose at the base of the trunk (arrow);below, roots appear converging and loose in the matrix. Scale bar, 50 cm.
a
1 m
b
Figure 2 | Large trunk and reconstruction of plant from South Mountain,New York. a, Composite image of large trunk specimen, a cast with upperand lower counterparts, NYSM 17040. Arrows at the distal end (top)correspond to the region in Fig. 3a; arrows at the proximal end (bottom)correspond to the region in Fig. 3b. b, Line drawing showing the architectureof Wattieza attached to Eospermatopteris. The length of the trunk is notfirmly established, so the minimum tree height is shown. Light branchesright, also in Fig. 1a right, appear in life position but are not definitivelyattached. Scale bar, 1m for both panels.
LETTERS NATURE |Vol 446 | 19 April 2007
906Nature ©2007 Publishing Group
Among the Pteridophyta, ferns with tree forms began appearing ca. 340Mya. These were quite abundant by245Mya and were represented by several independent lineages.24 Ancient tree ferns were, however, similar tothe extant tree ferns of the Dicksoniaceace and Cyatheaceae that can reach maximum heights of !20m(Figure 14(d)).24 Tree ferns lack secondary growth though, instead deriving their stem strength fromlocalization of xylem and sclerenchyma to the stem periphery, and from the accumulation of leaf bases and/or roots around the periphery (Figure 14(b)).7,24,27 This latter stem-strengthening strategy thickens the stem,allowing for an arborescent form. This is considered a primary thickening strategy because the leaves (and leafbases) are produced from a primary thickening meristem (PTM) (Figure 14(a)). This particular strategyresulted in arborescent forms arising independently in several lineages, including monocot angiosperms,23 suchas the present-day palms (Figure 14(e)), which can reach as much as 58m in height.28 These again reflectedchanges in cell wall biopolymer content, and tissue and cell wall organization.
2 m
~ 5 m
cambial initial
meristem
xylem
phloem
cambial initial
xylemxylem
pith
xylem
Bifacial Cambium
Phloem
Cortex
Secondary stem-thickening systems:bifacial cambium
• Xylem & phloem production• Capacity for stem expansion by
generation of new cambial initials
Extinct: Archaeopteris sp. Extant primitive gymnosperm:Araucaria araucana
(a)
(c)(b)
Figure 16 The most prevalent extant stem secondary thickening system: the bifacial cambium. This meristem not onlyproduces secondary phloem and xylem, but also regenerates its own cambial initial cells to allow for stem expansion (a). Theearliest tree form considered to have bifacial cambium activity was from the genus Archaeopteris (b). This belonged to theprogymnosperms, which in turn gave rise to gymnosperms and later angiosperms. Araucaria araucana (c) is an example of aprimitive extant gymnosperm. Redrawn with permission of Paleontological Society fromM. J. Donoghue, Paleobiology 2005,31, 77–93, copyright 2005 (a). Reproducedwith permission of Botanical Society of America fromC. B. Beck,Am. J. Bot. 1962,49, 373–382, copyright 1962 (b). Photo from L. B. Davin, Washington State University (c).
1188 Trees: A Remarkable Biochemical BountyMost prevalent extant stem secondary
thickening system: the bifacial cambiumThis meristem not only produces
secondary phloem and xylem, but also regenerates its own cambial
initial cells to allow for stem expansion (a). The earliest tree
form considered to have bifacial cambium activity was from the
genus Archaeopteris (b). This belonged to the progymnosperms,
which in turn gave r ise to g y m n o s p e r m s a n d l a t e r
angiosperms. Araucaria araucana (c) is an example of a primitive
extant gymnosperm.
Phloem rays
Vascular cambium
Sapwood Innerbark
Outer bark
Vascularcambium Cork
cambium(phellogen)
Heartwood
Sapwood
Growth rings
Pith
Xylem rays
Vascular cambium(not clearly visible)
PithGrowthrings
Innerbark
Outer barkWoodrays
Peridermlayers
Shootapical
meristemLeafprimordium
Pith Epidermis(outer layer)
Axillarybuds(later = branches)
Primaryxylem &phloem
Procambium
Secondary tissues: Cross section view
Groundtissueparenchyma
Protoderm (outer layer)
Bark and outer sapwood
Primary tissues
(d)(c)
(a) (b)
Heartwood Sapwood
Outerbark
Inner bark
{
(e)
Shootapical
meristem
Rootapical
meristem
Roots
Axillarybuds
Leaf
Figure 17
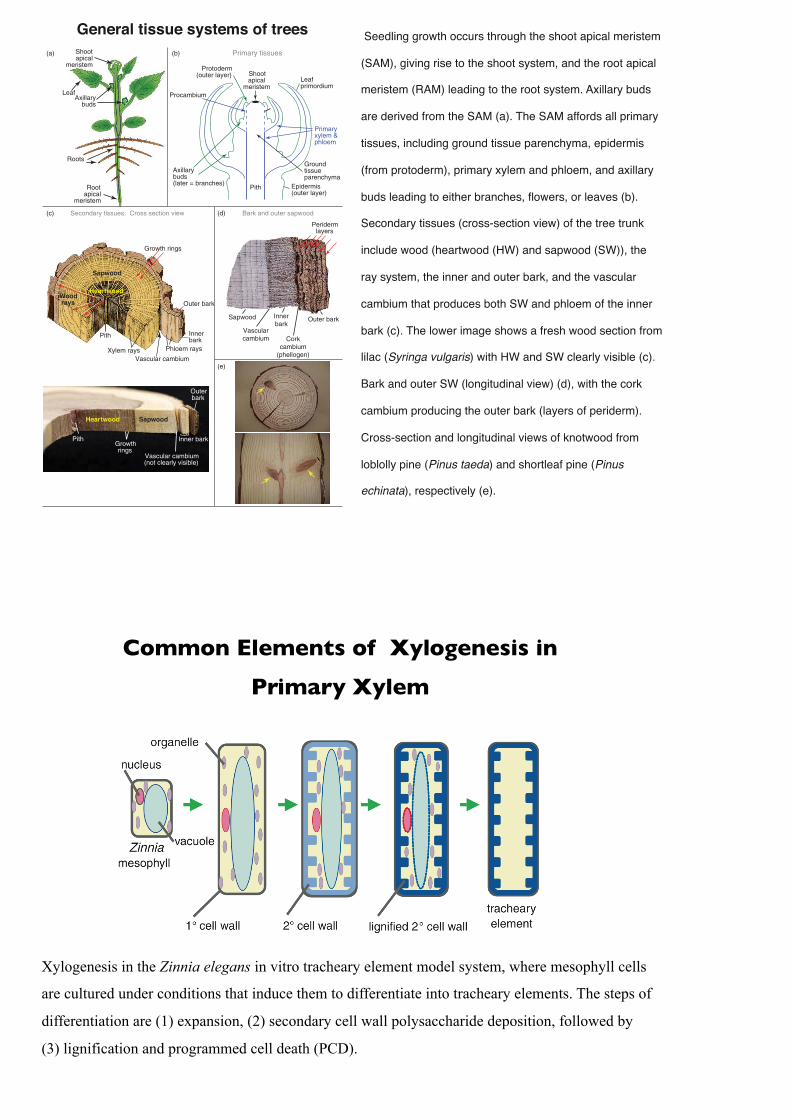
1190 Trees: A Remarkable Biochemical BountyGeneral tissue systems of trees Seedling growth occurs through the shoot apical meristem
(SAM), giving rise to the shoot system, and the root apical
meristem (RAM) leading to the root system. Axillary buds
are derived from the SAM (a). The SAM affords all primary
tissues, including ground tissue parenchyma, epidermis
(from protoderm), primary xylem and phloem, and axillary
buds leading to either branches, flowers, or leaves (b).
Secondary tissues (cross-section view) of the tree trunk
include wood (heartwood (HW) and sapwood (SW)), the
ray system, the inner and outer bark, and the vascular
cambium that produces both SW and phloem of the inner
bark (c). The lower image shows a fresh wood section from
lilac (Syringa vulgaris) with HW and SW clearly visible (c).
Bark and outer SW (longitudinal view) (d), with the cork
cambium producing the outer bark (layers of periderm).
Cross-section and longitudinal views of knotwood from
loblolly pine (Pinus taeda) and shortleaf pine (Pinus
echinata), respectively (e).
Common Elements of Xylogenesis in
Primary Xylem
Xylogenesis in the Zinnia elegans in vitro tracheary element model system, where mesophyll cells
are cultured under conditions that induce them to differentiate into tracheary elements. The steps of
differentiation are (1) expansion, (2) secondary cell wall polysaccharide deposition, followed by
(3) lignification and programmed cell death (PCD).

Common Elements of Xylogenesis in Secondary Xylem• For angiosperm vessels and
fibers, arising from the vascular
cambium.
• For vessels of angiosperms, and
to a lesser extent for gymnosperm
tracheids, expansion is strongly in
the radial direction, followed by
secondary cell wall production,
lignification, and protoplast lysis.
• For angiosperm fibers, expansion
is axial as the cells elongate by
intrusive growth, followed by
secondary cell wall synthesis,
lignification, and protoplast lysis.
(A) start of cytokinesis in radial view; (B) later stage of
cytokinesis with extending cell plate and opposing phragmoplasts
in radial view; (C) similar stage of cytokinesis in tangential view;
(D) enlarged diagram of telophase and initiation of the
phragmoplast; (E) beginning of expansion of the cell plate as the
nuclei reform; (F) enlarged view of the expanding cell plate and
phragmoplasts. (G–J) High-pressure frozen dividing cambial cells
of Pinus contorta; Figs. G, H, and I are equivalent to Figs. D, E,
and F, respectively. (G) transmission electron micrographs (TEM)
showing the initiation of the phragmoplast between the recently
separated chromosomes; (H) light micrograph (LM) showing the
formation of the cell plate between the reforming nuclei; (I) light
micrograph showing the expansion of the cell plate by
advancement of the phragmoplast through the central vacuole;
(J) transmission electron micrograph showing the greatly
extended cell plate and, within the phragmoplast, the delicate
newly formed cell plate. Scale bars = 1 µm for Figs. G and J;
Scale bars = 10 µm for Figs. H and I.
Pinus strobus dividing cambial cells

Primary and Secondary Metabolism
Primary metabolism
The biological reactions are essential to maintain life in living organisms
and are known as primary metabolism.
Plant convert sunlight energy to chemical energy, such as ATP, NADPH, by
the mediation of chlorophyll in chloroplasts and synthesize sugars and
starch from CO2 by using ATP and NADPH+.
These carbohydrates are stored and used for differentiation and formation
of plant tissues.
Primary and Secondary Metabolism
Secondary metabolism
The metabolisms which are not directly related to maintaining life, are
known as secondary metabolisms.
The products formed by secondary metabolism are called secondary
metabolites.
Secondary metabolite play a role in reinforcement of tissue and tree body
(e.g. cellulose, lignin, suberin), protection against insects, dieses, and plant
regulation (plant hormones).

Primary and Secondary Metabolism
All organisms need to transform and interconvert a vast
number of organic compounds to enable them to live, grow and
reproduce.
All organisms need to provide themselves with energy in the
form of ATP, and a supply of building blocks to construct their
own tissues.
An integrated network of enzyme-mediated and carefully
regulated chemical reactions in used for this purpose,
collectively referred to as intermediary metabolism, and the
pathways involved are termed metabolic pathway.
Primary and Secondary Metabolism
The pathways for generally modifying and synthesizing
carbohydrates, proteins, fats, and nucleic acids are found to be
essentially to same in all organisms, apart from minor variations.
These processes demonstrate the fundamental unity of all living matter,
and are collectively described as primary metabolism, with the compounds
involved in pathways being termed primary metabolites.

Primary Metabolisms
Degradation of carbohydrates and sugars generally proceeds via
the well characterized pathways, known as glycolysis and the
kerbs / citric acid / tricarboxylic acid cycle, which release energy
from the organic compounds by oxidative reactions.
Oxidation of fatty acids from fats by the sequence called β-
oxidation also provides energy.
Primary Metabolisms
Aerobic organisms are able to optimize these processed by
adding on a further process, oxidative phosphorylation. This
improves the efficiency of oxidation by incorporating a more
general process applicable to oxidation of a wide variety of
substrates rather then having to provide specific process for each
individual substrate.

Primary Metabolisms
Proteins taken in via the diet provide amino acids, but the
proportions of each will almost certainly vary from the
organism’s requirements.
Most organisms can synthesize only a proportion of the amino
acids they actually require for protein synthesis. Those
structures not synthesized, so-called essential amino acids, must
be obtained from external sources.
Secondary Metabolisms
The compounds which synthesized from the secondary metabolisms are so-called secondary metabolites.
Secondary metabolites are formed in only specific organisms, or groups of organisms, and are expression of the individuality of species.
Secondary metabolites are not necessarily produced under all conditions, and in the vast majority of cases the function of these compounds and their benefit to the organism is not yet known.
It is this area of secondary metabolism that provides most of the pharmacologically active natural products.

Secondary Metabolisms
To make such compounds as sugars, waxes, lignin starch,
pigments, or alkaloids, plants utilize very specific enzymes, each
of which catalyzes a specific metabolic reaction.
The enzymes are proteins called organic catalysts.
These enzymes are coded by specific genes in the plants DNA and are
made via processed we call transcription and translation.
When there is a series of enzymatically catalyzed reaction in a well-defined
sequence of step, we have what is termed a metabolic pathway.
Primary and Secondary Metabolism
Primary and secondary metabolites leave a “grey area” at the
boundary, so that some groups of natural products could be
assigned to either divisions.
Primary metabolites → Biochemistry
Secondary metabolites → Natural products Chemistry
Generalized View of a Plant cell and Its Subcellular Compartments
顆粒性內質網:附著其上的小顆粒為核糖體,是負責蛋白質合成及運輸(至高基氏體)
平滑內質網:無核糖體附著,負責脂質的合成及運輸。
液泡植物細胞生長時先行成一些小液泡,這些小液泡逐漸融合成為大液泡,所以成熟的植物細胞中常可見到大型液泡-或稱為「中央液泡」。液泡可貯存物質,如醣類、胺基酸、花青素,也和細胞滲透壓維持有關。
高基氏體: 由許多層扁平囊狀膜構成。內含酵素。為分泌物合成與蛋白質(構成細胞膜的蛋白質和分泌至細胞外的蛋白質)加工的場所。兩側的小囊泡為「分泌小泡」,裝載要分泌至細胞外的物質。
粒線體: 分成內外兩層膜。內膜平行排列上有呼吸酵素,行呼吸作用,製造ATP。粒線體有自己的DNA和核糖體。
Generalized View of a Plant cell and Its Subcellular Compartments

Generalized View of a Plant cell and Its Subcellular Compartments
Cell wall is the primary site for polymerization of cell wall polysaccharides, lignin, and amorphous silica gel in plants that accumulate this polymer.
Nucleus is the information center of the cell It is surrounded by a double lipid membrane and contains genetic
information (DNA) needed to create proteins within the cell. Cytoplasm is the liquid phase of the cell that contain:
Majority of the ribosome involved in protein synthesis. Microtubules and microfilaments that provide a physical skeleton for the cell
and also action cellular trafficking of proteins and organelle.
All the soluble enzymes of the cell not found within organelles or cellular membranes.
Microfilaments
Microfilaments are fine, thread-like protein fibers, 3-6 nm in diameter. They are composed predominantly of a contractile protein called actin, which is the most abundant cellular protein. Microfilaments' association with the protein myosin is responsible for muscle contraction. Microfilaments can also carry out cellular movements including gliding, contraction, and cytokinesis.
Microtubules
Microtubules are cylindrical tubes, 20-25 nm in diameter. They are composed of subunits of the protein tubulin--these subunits are termed alpha and beta. Microtubules act as a scaffold to determine cell shape, and provide a set of "tracks" for cell organelles and vesicles to move on. Microtubules also form the spindle fibers for separating chromosomes during mitosis. When arranged in geometric patterns inside flagella and cilia, they are used for locomotion.
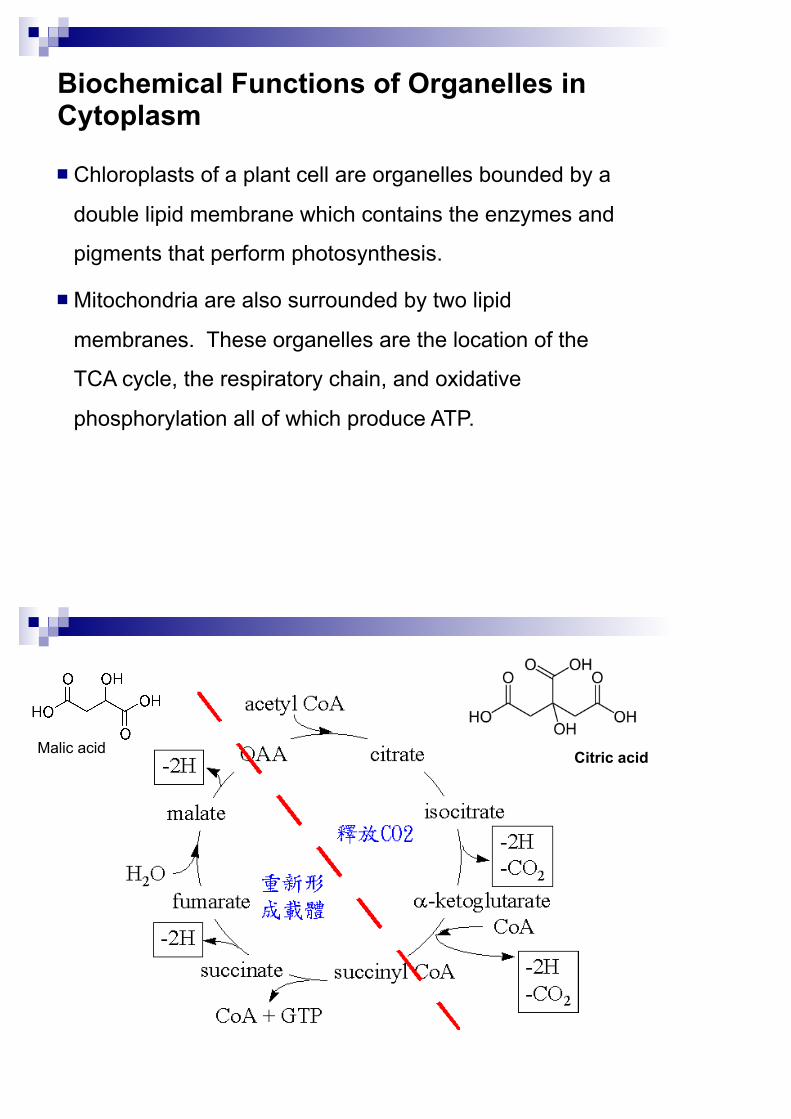
Biochemical Functions of Organelles in Cytoplasm
Chloroplasts of a plant cell are organelles bounded by a
double lipid membrane which contains the enzymes and
pigments that perform photosynthesis.
Mitochondria are also surrounded by two lipid
membranes. These organelles are the location of the
TCA cycle, the respiratory chain, and oxidative
phosphorylation all of which produce ATP.
Citric acidMalic acid

Biochemical Functions of Organelles in Cytoplasm
Endoplasmic reticulum (ER) is a system of membrane-
bound tubes and flattened sacs that spread through the
cell and work in conjunction with dictyosomes (on Golgi
bodies) to produce and secrete various compounds as
well as to deliver specific proteins and membrane lipids to
their proper locations within the cell.
網狀體
Biochemical Functions of Organelles in Cytoplasm Peroxysomes are microbody organelles that have the very
important function of housing the formation of toxic peroxides that are necessary for other metabolic mechanisms but would otherwise kill the cell.
Glyoxysomes are specialized peroxysomes found only in the early stages of plant development. They contain the enzymes necessary for the conversion of stored lipids to carbohydrates in such processed as seed germination where photosynthesis is not yet possible.
過氧化小體
乙醛小體

Biochemical Functions of Organelles in Cytoplasm Vacuoles play a wide variety of roles in cellular metabolic,
some “waste bins”, but they also play a vary important role
as a support structure.
Cyclosis
Most of the cell’s content are in a continuous state of motion, called
cyclosis, and each molecule within the cell is, at a molecular level,
experiencing Browing movement.
胞質環流