fast imaging in two dimensions resolves extensive … · gustavo brum, adom gonzález, juliana...
TRANSCRIPT

2000;528;419-433 J. Physiol.
Gustavo Brum, Adom González, Juliana Rengifo, Natalia Shirokova and Eduardo Ríos
skeletal muscle sparks in frog2+Fast imaging in two dimensions resolves extensive sources of Ca
This information is current as of January 9, 2007
publication unless article is open access. This version of the article may not be posted on a public website for 12 months after
http://jp.physoc.org/cgi/content/full/528/3/419
This is the final published version of this article; it is available at:
[email protected]. No part of this article may be reproduced without the permission of Blackwell Publishing:
articles are free 12 months afterThe Journal of Physiology Online. http://jp.physoc.org/subscriptions/ go to: The Journal of Physiology Onlinepublished continuously since 1878. To subscribe to
is the official journal of The Physiological Society. It has beenThe Journal of Physiology Online
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

The process of Ca¥ release for contractile activation in
striated muscle requires that dihydropyridine receptors in
the transverse tubules sense the action potential, and
transduce it to opening of the sarcoplasmic reticulum Ca¥
channels (review by Leong & MacLennan, 1999). In
amphibian skeletal muscle, Ca¥ sparks (Cheng et al. 1993;Tsugorka et al. 1995) constitute a distinctive, and apparently
dominant form of Ca¥ release (Klein et al. 1996). Althoughsparks require Ca¥ as mediator (Klein et al. 1996; Shirokova& R� úos, 1997; Gonz�alez et al. 2000a), little else is known
about their mechanism (e.g. Niggli, 1999; Shirokova et al.
1999; Schneider, 1999). Our recent work (Gonz�alez et al.2000a,b) provided evidence that spark sources involve
multiple channels, presumably some or all of the channels in
a couplon (junctional array of release channels; Stern et al.1997; Franzini-Armstrong et al. 1999). If this is the case,
then spark sources must be extensive, rather than punctual,
and sparks may be asymmetric, not spherically symmetric
as they are often assumed to be (e.g. R� úos et al. 1999).
Until now, two-dimensional spatial properties of sparks
have been explored by line scanning successively in two
directions (Cheng et al. 1996), a method that does not yield
Journal of Physiology (2000), 528.3, pp.419—433 419
Fast imaging in two dimensions resolves extensive sources of
Ca¥ sparks in frog skeletal muscle
Gustavo Brum, Adom Gonz�alez *, Juliana Rengifo*, Natalia Shirokova*
and Eduardo R� úos *
*Department of Molecular Biophysics and Physiology, Rush University, 1750 W. HarrisonStreet, Chicago, IL 60612, USA and Departamento de Biof� úsica, Facultad de Medicina,
G. Flores 2125, Montevideo, Uruguay
(Received 2 May 2000; accepted after revision 7 July 2000)
1. Ca¥ sparks were monitored by confocal laser-scanning microscopy of fluo-3 at video rates,
in fast twitch muscle fibres, stimulated by exposure to caffeine andÏor low [Mg¥]. Scanning
was in two spatial dimensions (‘2D’) or 2D plus time, at 4 ms per image frame. Sparks were
identified in 2D images of normalized fluorescence by an automatic procedure, which also
evaluated the event’s location and morphometric parameters.
2. Most sparks were circular, but some were elongated, especially in caffeine. Separation of the
spark from circular symmetry was quantified by its eccentricity (lengthÏwidth − 1).
3. In an internal solution with 0·4 mÒ [Mg¥], sparks (989 events in 4 cells) had amplitude 0·73,
width 1·94 ìm, length 2·12 ìm and eccentricity not significantly different from 0. Upon
application of 1 mÒ caffeine, length (of 2578 events in the same cells) increased significantly
(by 0·41 ìm, or 19%), width increased by 0·18 ìm (9%) and amplitude decreased slightly.
The eccentricity became significantly different from 0, and the sparks’ long axis
predominantly oriented parallel to the plane of the Z disks.
4. More than 10% of the events in caffeine had length greater than 4 ìm, a relatively flat top,
and a sharp termination at both ends of the major axis. Additionally, there was only a weak
correlation between eccentricity and amplitude. These properties suggest that the elongated
events are produced by simultaneous opening of multiple channels within a junction, rather
than anisotropic diffusion of Ca¥ or random overlap of round sparks.
5. Elongated events often increased in eccentricity early in their evolution. Then, most
remained elongated during their rise and decay, while others spread spatially in the plane of
the Z disks. In 1—2% of the events, the centre of mass migrated in space, over time, at
•0·1 ìm ms¢.
6. These spatio-temporal features require the involvement of multiple release channels, at
spatially resolvable locations. Because sources often spread over distances greater than 1 or
2 ìm, and arrays of junctional elements (couplons) are at most 1·2 ìm long, it must be
possible for activation of release to propagate between neighbouring couplons, especially
under the influence of caffeine andÏor low [Mg¥].
11050
Keywords:
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

two-dimensional information on individual events. (Such
information is present, but not systematically analysed, in
images of Tanaka et al. 1997.) We reported a sizable increase
in spark width under caffeine, using line scanning parallel
to the fibre axis (Gonz�alez et al. 2000a). This direction is
roughly perpendicular to the long axis of the couplon, and
hence especially unsuitable for exploring spatial aspects of
the sources. To test the possible involvement of extended
arrays of channels in spark generation it is necessary to
simultaneously monitor the events in two spatial directions.
We now take advantage of the speed of a video-rate laser-
scanning confocal system to study for the first time the
properties of skeletal muscle Ca¥ sparks in two dimensions
of space, and follow the evolution of such images in time. It
becomes apparent that caffeine, which increases the
sensitivity of release channels to stimulation by Ca¥
(e.g. Herrmann-Frank et al. 1999), causes sparks to grow in
the direction of the junctional arrays of release channels.
METHODS
Experiments were carried out in cut skeletal muscle fibres from the
semitendinosus muscle of Rana pipiens. Following a protocol
approved by our Institutional Animal Care and Use Committee,
adult frogs were anaesthetized in 15% ethanol, then killed by
pithing. Fibres were mounted on the stage of an inverted
microscope with a video-rate confocal scanner (RCM8000, Nikon
Inc., Melville, NY, USA; Tsien, 1990; Kawanishi et al. 1994;Lacampagne et al. 1999).
The experiments were carried out in permeabilized cells, using
methods derived from those of Lacampagne et al. (1998), describedin detail by R� úos et al. (1999). Cells were stretched to between 3·2
and 3·6 ìm per sarcomere and immersed in a solution with low
[Mg¥], which elicited sparks at a frequency of •15 per image. The
solution, adjusted to pH 7 and 260 mosmol kg¢, contained (mÒ):
100 caesium glutamate, 1 EGTA, 5 glucose, 5 phosphocreatine, 10
Hepes, 5 Mg-ATP, 0·1 fluo-3 (Molecular Probes, Eugene, OR,
USA), with added calcium for a nominal [Ca¥] of 100 nÒ, 0·25 mÒ
MgClµ (0·41 mÒ nominal [Mg¥]), plus 8% (40000 kDa) dextran.
Temperature was 17—19°C.
Excitation light of an argon-ion laser (Enterprise 621, Coherent,
Santa Clara, CA, USA) was selected by a 505 nm dichroic mirror
and emitted light by a 515 nm long pass filter. Imaging used a ²40,
1·15 NA water immersion objective (CF UV, Nikon). Scanning, in
two dimensions, proceeded first along what we define as the
y direction, presented vertically in the figures, at a rate of one line
of 512 pixels (spaced 0·167 ìm) every 64 ìs; 481 lines stacked from
left to right in the x direction constitute an image. As lines are
scanned at distances of 0·138 ìm, imaging in the x direction
proceeds at •2·2 ìm ms¢, so that the 15 ìm horizontal calibration
bar in every image corresponds also to about 7 ms. All images
illustrated are presented at equal spacing (of 0·167 ìm), by linear
interpolation in the x direction.
Space- and time-resolved images (termed ‘3D’ for short) were
obtained by scanning repeatedly a 5·3 ìm ² 85 ìm band in the
object, at 4 ms intervals.
Normalization to resting fluorescence
Images of fluorescence F(x, y) or F(x, y, t), and normalized
fluorescence, FÏF0, which provides a first estimate of increase in
local [Ca¥] relative to its resting value, are shown.
Single 2D images do not contain resting fluorescence information in
those locations where sparks occur. Therefore we developed a
procedure, illustrated in Fig. 1, that uses signal averages. Figure 1Ais a full individual 2D image. Up to 30 images could be obtained at
the same location without deterioration or movement. In the
example, 19 images were obtained, each containing several sparks.
Figure 1B is their pixel-to-pixel sum divided by 19. This ‘rough’
average was used as divisor to obtain a rough normalized version of
every individual image. On each the automatic locator identified
spark areas (those exceeding the value 1 by a set amount), and
excised them (zeroing the corresponding pixels) from the original
non-normalized records. Figure 1C is the boxed part of individual
image A after zeroing the spark regions (in red). These excised
images were then added, to obtain the sum image D. To obtain
resting fluorescence, the sum was divided by a (pixel-dependent)
‘counter’ E. The counter is an array, with the dimensions of the
image, whose value at any pixel is the number of images not zeroed
there, and therefore may range between 1 and 19. The ratio of the
sum D and the counter E is the resting fluorescence, F0(x, y),illustrated in Fig. 1F. Normalized images were obtained dividing
F(x, y) by F0(x, y), pixel to pixel. Figure 1G is the image in Anormalized to resting fluorescence. Especially in the presence of
caffeine, among many circular, symmetric sparks, there are some
obviously elongated events. Shown at the bottom is the profile of
normalized fluorescence as a function of coordinate ‘s’ along the
axis of the large event. It made sense to determine the orientation
of this axis relative to the plane of the Z disks.
The direction of Z disks
2D scanning proceeded first in the longitudinal or y direction
(parallel to fibre axis) and rastered transversely, in the x direction.
While the x direction was generally parallel to the plane of the
Z disks, these could be tilted as much as 30 deg, as shown in Fig. 1F.An automatic method was devised to find locally the direction of
Z disks. The method, illustrated in Fig. 2, uses the clear striation
visible in the averaged image of resting fluorescence (Fig. 2A). When
the event locator described below finds a spark in an individual
image, say, at the position marked by crossbars, it then analyses the
surrounding sub-array of approximately 30 ìm ² 30 ìm. This sub-
array is rotated, relative to the cartesian axes, by 1 deg increments
between −45 and 45 deg. Figure 2B shows the sub-array after
rotation by −25 deg (i.e. clockwise). At every rotation the sub-array
is then averaged along the x coordinate, yielding a function F(y)represented in Fig. 2B. (To avoid edge effects produced by rotation
of a square array, only a central 15 ìm ² 15 ìm box in the sub-
array is averaged.) The root mean square (r.m.s.) value of the
average is high at this particular rotation, because the sarcomeric
striation becomes nearly perpendicular to the y axis. Figure 2Cshows the dependence of the r.m.s. on the rotation angle. The
position of its maximum (−28 deg) determines the angle of the
Z disks relative to the y axis.
Figure 2D shows as short segments the direction of the Z disks at
every position where a spark was found in any of the 19 images
from the same region, demonstrating good agreement between the
method and the direction appraised by eye.
G. Brum, A. Gonz�alez, J. Rengifo, N. Shirokova and E. R� úos J. Physiol. 528.3420
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

Morphometry of events
A computer procedure described earlier for line scans (Cheng et al.1999), which locates events automatically, was modified to work on
2D images. Events were detected — on images normalized and
filtered — by a relative criterion, fluorescence exceeding F0 by 2·5
standard deviations of the non-spark regions. Figure 3A is one
individual image in the presence of caffeine, after normalization.
At the bottom is the profile of normalized fluorescence along the
slanted axis s. Figure 3B illustrates automatic measurements
performed by the locator on the events detected. For each event a
contour was traced (the border of the region where the detection
threshold is exceeded; in blue). On this contour the program located
the two pixels farthest apart from each other. These points
determine the event’s major axis (central line in red). The spark’s
length is the length of this segment, while its width is the distance
between the two tangents to the contour parallel to the major axis
(in red). These two parameters were used to define an absolute
eccentricity, evaluating separation of the event from circular
Ca¥ sparks in three dimensionsJ. Physiol. 528.3 421
Figure 1. Normalization of sparks in two dimensions
A, fluorescence image (after 9-point median filtering), in a cell exposed to 1 mÒ caffeine. The longitudinal
axis of the fibre is represented vertically (y coordinate); scanning proceeds by vertical lines of pixels and
the area is swept from left to right. The grey scale spans 0 to 255 units of fluorescence. B, rough reference
image; average of A plus 18 other images from the same location (portion shown is boxed in A). The colourscale, in E, spans 0 to 63 units of fluorescence. Normalization of image A by reference B was followed by
application of the automatic event detector, which identified spark areas above a threshold level
(mean + 1·3 s.d.). C, the original image in A with spark areas excised (set to 0) and marked in red. D, sumof all 19 images with spark areas excised. The darker areas are those that had sparks in a greater number of
individual images. E, a counter of sparks, which starts at 19 everywhere and is reduced by one in every
pixel where a spark is excised. The colour table spans the range 0—19. F, the resting fluorescence image
F0(x, y), the result of dividing D by E. Note the clear sarcomeric structure. G, image A divided by resting
fluorescence F0(x, y). At bottom is the profile of normalized fluorescence (averaged for a 0·8 ìm band of 5
pixels) along abscissa s. Identifier: 1203n2, image 69 and sum of images 65 to 83.
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

symmetry, as (lengthÏwidth − 1). A relative eccentricity (with sign)
is defined later.
The orientation of the spark was quantified, relative to the plane of
the Z disk, by an orientation angle. For this purpose, after locating
a spark, the program reverted to the resting fluorescence image to
determine the local direction of the Z disks as described above. For
the environment of the boxed spark in Fig. 3B, this direction is
represented by the black arrow in the enlargement. The orientation
angle á is measured (counterclockwise) between the Z disk direction
and the long axis of the spark (red arrow in the enlargement).
Statistics
Following every parameter average, in parentheses, is the standard
error of the mean. Significance of differences between averages, X1
and Xµ, of samples of sizes N1 and Nµ, was established by ANOVA
or by two-tailed tests of likelihood of the conventionally defined tvariable:
(X1 − Xµ)�{[N1Nµ (N1 + Nµ − 2)]Ï[(N1 + Nµ)(N1S1 + NµSµÂ)]},
where Sj are the sample variances.
RESULTS
Morphology of sparks in two dimensions
2D images of events obtained in the presence of 1 mÒ
caffeine are illustrated in Figs 1 and 3, while images from
the same fibre in the absence of caffeine (reference) are in
Fig. 4. To the eye, most sparks in reference were symmetric,
and approximately circular. This is confirmed by the
contours drawn for every detected event by the locator
program (right side panels on Fig. 4). By contrast, in
caffeine there were many highly asymmetric, oval or
elongated events. This asymmetry might reflect an actual
feature of the local increase in [Ca¥], or trivially result from
anisotropic dye distribution. The asymmetries introduced
by an inhomogeneous distribution of dye were corrected to a
first approximation normalizing F(x, y) by F0(x, y), the
fluorescence at rest, as described in Methods. Figure 1Fshows the resting fluorescence of the boxed portion of
G. Brum, A. Gonz�alez, J. Rengifo, N. Shirokova and E. R� úos J. Physiol. 528.3422
Figure 2. Determination of direction of the Z disks
A, portion of the resting fluorescence image made as described in Fig. 1. The procedure determines the
direction of Z disks at or near the location of a detected spark (marked by the crossbar). A sub-array of
30 ìm ² 30 ìm around this point is rotated incrementally. B, the sub-array after rotation by −25 deg.
Plotted on the right side is F(y), average over the x dimension of the rotated image (to avoid edge effects
clearly visible at the corners of the sub-array, only the portion within the box is used). F(y) varies widelyaround its mean, because the structural striations are almost parallel to the x axis at this rotation angle.
C, root mean square value of the function F(y)-mean, plotted as a function of the rotation angle. Note the
sharp maximum at −28 deg, which is taken to be the local angle of the Z disks. D, resting fluorescence
image with vectors marking the Z disk direction, placed by the spark detector at all locations where events
were found in any of the 19 images. The array of vectors follows correctly the orientation of the Z disks.
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

Fig. 1A, obtained using 19 images from the same location.
Note that the periodic variation in F0, probably due to dye
accumulation (Tsugorka et al. 1995; Klein et al. 1996),outlines the sarcomeric structure and demonstrates that the
preparation was steady during imaging.
Normalized images in caffeine are shown in Figs 1G and 3A(the lower panels in Fig. 3 are example events from other
images). The periodic pattern of fluorescence present in the
raw images F(x, y) is removed in the normalized images, butthe sparks that were oval originally maintain their shape
upon normalization. Elongated sparks are therefore not a
consequence of uneven dye distribution.
Four spark parameters, defined in Methods, were analysed.
For 13 images in reference (four of which are illustrated in
Fig. 4) and 19 images in caffeine (illustrated in Figs 1 and 3)
the histogram of absolute eccentricity (lengthÏwidth − 1) is
Ca¥ sparks in three dimensionsJ. Physiol. 528.3 423
Figure 3. 2D properties and analysis of sparks in caffeine
A, normalized fluorescence image, in a cell exposed to 1 mÒ caffeine. The colour table spans the range
0—2·3. At the bottom is the profile of fluorescence along the abscissa s. B, automatic location and
morphometry of events. The locator finds a contour (blue, see enlargement), a major axis, its length, event
width (distance between tangents parallel to major axis) and orientation angle á between the local direction
of the Z disks (black arrow) and that of the event’s axis (red). Identifier: 1203n2, image 78. C, D and E,example images of large sparks and profiles along their major axis. Identifiers: C, 1203n2.96; D, 0313d.104;E, 1203n2.74.
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

plotted in Fig. 5A. In both reference and caffeine the
frequency peaks at a small non-zero value (because of noise,
sparks are seldom perfectly symmetric). There is an excess
of events of large eccentricity in caffeine; the difference of
averages is small but significant (see figure legend).
More impressive is the effect of caffeine on the orientation of
sparks relative to the plane of the Z disks. The distribution
of orientation angle á, in Fig. 5B, is nearly isotropic in
reference (dashed line) but becomes concentrated near the
value 0 in caffeine.
A working hypothesis is that the increase in frequency of
elongated events is due to the increase in the number of
channels involved in individual sparks. In this hypothesis,
the increase in event length should occur along junctions,
that is, in the direction of the Z planes. Therefore, the
increase in eccentricity should be accompanied by an
alignment — an increase in the frequency of small angles á.
We combined the two predicted effects, increase in
eccentricity and orientation, into a single indicator, (relative)
eccentricity, with a sign defined as positive when the
orientation is aligned with the Z plane (−45 deg < á
G. Brum, A. Gonz�alez, J. Rengifo, N. Shirokova and E. R� úos J. Physiol. 528.3424
Figure 4. 2D properties of sparks in reference
Left, normalized fluorescence in the fibre of Fig. 1, before exposure to caffeine. Portions represented are
from different images, among 13 obtained in the same location. Right, the locator finds sparks that are
generally circular — the ‘major axis’ is randomly oriented. Identifier: 1203n2, images 7—9, 11.
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

Ca¥ sparks in three dimensionsJ. Physiol. 528.3 425
Figure 5. Distributions of eccentricity and orientation angle
A, histograms of absolute eccentricity, defined as lengthÏwidth − 1. In caffeine (continuous trace) the
distribution is displaced to higher values. The average eccentricity went from 0·36 (s.e.m. = 0·019) in
reference, to 0·43 (s.e.m. = 0·011) in caffeine. The difference is significant by ANOVA (P < 0·03).
B, distribution of orientation angle, which is almost random in reference (dashed), but is concentrated
around 0 in caffeine. C, distribution of relative eccentricity, with sign defined as positive when |á| < 45 deg,
and negative otherwise. In caffeine there are many events with high positive values of eccentricity,
resulting in a clear shift in the positive direction. There are 213 events in reference (detected in 13 images
from the same location), and 409 in caffeine (19 images from the same location). Identifier 1203n2.
––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
Table 1. Effects of caffeine on spark morphology
––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
Events Amplitude Width (ìm) Length (ìm) Eccentricity Angle (deg)––––– –––––––– –––––––– –––––––– ––––––––– –––––––––Ref. Caff. Ref. Caff. Ä Ref. Caff. Ä Ref. Caff. Ä Ref. Caff. Ä Ref. Caff. Ä
––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
0312b 289 1463 0·49 0·44 −0·1 1·79 1·98 0·19 2·30 2·67 0·37 −0·16 −0·02 0·14 53·9 42·5 −11·4
0313d 206 320 0·59 0·64 0·05 1·83 2·19 0·36 2·31 3·05 0·74 0·00 0·19 0·19 43·0 32·8 −10·2
1203n 281 386 0·81 0·88 0·07 2·20 2·25 0·05 2·94 3·15 0·21 −0·09 0·10 0·19 48·2 35·6 −12·6
1203n2 213 409 1·01 0·82 −0·19 1·92 2·05 0·13 2·59 2·92 0·33 0·04 0·22 0·18 40·6 29·9 −10·7
Total 989 2578 – – – – – – – – – – – – – – –
Mean – – 0·73 0·70 −0·03 1·94 2·12 0·18 2·54 2·95 0·41* −0·05 0·12 0·18* 46·43 35·20 −11·2*
s.e.m. – – 0·12 0·10 0·06 0·09 0·06 0·07 0·15 0·10 0·11 0·05 0·05 0·01 2·95 2·70 0·52
––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
Under amplitude, width and length are listed averages of parameter values (determined as described in
Methods). Ref. and Caff., respectively, list averages in reference condition and in the presence of 1 mÒ
caffeine. Angle is the absolute value of the orientation angle á (see Methods). Ä is paired difference of
averages (Caff. − Ref.). Means are averages over all experiments (average of paired differences in bold type).
*Significantly different from 0 (see Methods). Among caffeine effects, the increase in length and
eccentricity and the decrease in angle were statistically significant in every individual experiment.
––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from
< 45 deg) and negative otherwise. The distribution of this
parameter is shown in Fig. 5C. In reference, it was roughly
bell shaped and centred at 0 (mean 0·00, s.e.m. 0·03) as onewould expect from roughly circular sparks with no preferred
orientation. In caffeine the distribution, still bell shaped,
moved positive (mean 0·22 ± 0·02).
In four experiments analysed, nearly 1000 events in
reference and 2600 in caffeine were detected. The average
parameter values are listed in Table 1. Caffeine caused a
significant increase in length, of 16%, and eccentricity,
from an average value of −0·05 in reference, to 0·18. It
increased width by 9%, and it reduced the absolute value of
the orientation angle by 25%. The changes in length,
eccentricity and angle were also significant when examined
in every individual experiment.
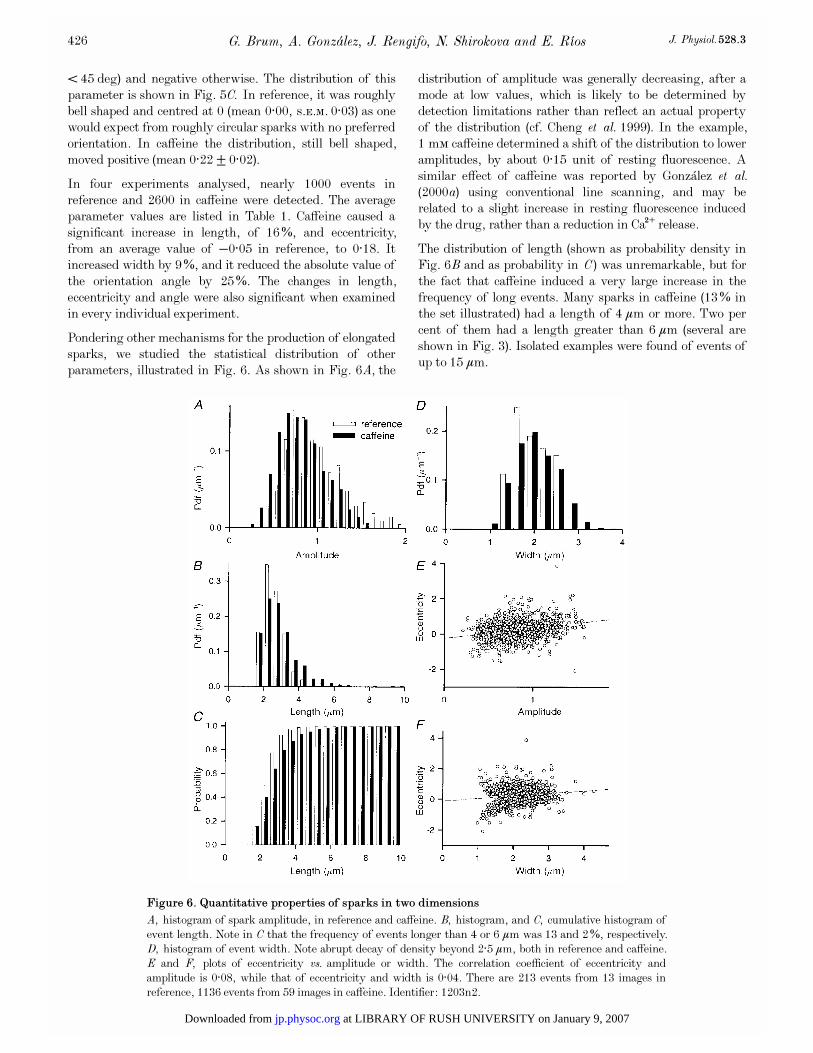
Pondering other mechanisms for the production of elongated
sparks, we studied the statistical distribution of other
parameters, illustrated in Fig. 6. As shown in Fig. 6A, the
distribution of amplitude was generally decreasing, after a
mode at low values, which is likely to be determined by
detection limitations rather than reflect an actual property
of the distribution (cf. Cheng et al. 1999). In the example,
1 mÒ caffeine determined a shift of the distribution to lower
amplitudes, by about 0·15 unit of resting fluorescence. A
similar effect of caffeine was reported by Gonz�alez et al.(2000a) using conventional line scanning, and may be
related to a slight increase in resting fluorescence induced
by the drug, rather than a reduction in Ca¥ release.
The distribution of length (shown as probability density in
Fig. 6B and as probability in C) was unremarkable, but forthe fact that caffeine induced a very large increase in the
frequency of long events. Many sparks in caffeine (13% in
the set illustrated) had a length of 4 ìm or more. Two per
cent of them had a length greater than 6 ìm (several are
shown in Fig. 3). Isolated examples were found of events of
up to 15 ìm.
G. Brum, A. Gonz�alez, J. Rengifo, N. Shirokova and E. R� úos J. Physiol. 528.3426
Figure 6. Quantitative properties of sparks in two dimensions
A, histogram of spark amplitude, in reference and caffeine. B, histogram, and C, cumulative histogram of
event length. Note in C that the frequency of events longer than 4 or 6 ìm was 13 and 2%, respectively.
D, histogram of event width. Note abrupt decay of density beyond 2·5 ìm, both in reference and caffeine.
E and F, plots of eccentricity vs. amplitude or width. The correlation coefficient of eccentricity and
amplitude is 0·08, while that of eccentricity and width is 0·04. There are 213 events from 13 images in
reference, 1136 events from 59 images in caffeine. Identifier: 1203n2.
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

The distribution of widths was interesting. In three of four
fibres examined in reference and caffeine, it showed a
noticeable drop in density at 2·5 to 3 ìm, which could be
taken to indicate a physical barrier to diffusion (the
sarcomere length in all fibres was greater than 3 ìm).
Such a restriction could provide a mechanism for the
production of elongated sparks: Ca¥ coming from a point
source first diffuses isotropically, then reaches an obstacle
and starts to diffuse anisotropically, as water flowing in a
channel or ‘gutter’. This and related mechanisms predict for
elongated sparks a high correlation between eccentricity and
amplitude, and, perhaps most characteristically, a spatial
dependence of fluorescence that peaks at or near the centre
and ‘spills’ to the sides according to the laws of diffusion.
The profiles of fluorescence along the spark axis, shown in
Figs 1 and 3, were not consistent with this hypothesis.
Occasionally, long sparks showed a peak at their centre
(Fig. 3A). Most long sparks, however, had a flat or slanted
top (as in Figs 1G and 3C, D and E). Figure 6E plots
eccentricity vs. amplitude for the sparks in caffeine. Against
the prediction, the variables were not correlated (the
correlation coefficient r  was on average 0·09, s.e.m. = 0·08,
n = 4 experiments in caffeine). Figure 6F shows that in the
example there was no correlation between width and
eccentricity either (r  = 0·04, s.e.m. = 0·09, n = 4).
Elongated sparks could also result from random overlap of
circular ones, but this should occur infrequently (as shown
in Discussion). In conclusion, the elongated events
correspond either to activation of an extended source — a
group of channels in an extended area — or to non-random
Ca¥ sparks in three dimensionsJ. Physiol. 528.3 427
Figure 7. Sparks in three dimensions
A, normalized fluorescence in reference, imaged at 4 ms intervals in a 5·3 ìm-wide band. B, representativesparks characterized by the automatic locator are circular, and their position is steady. C, circular and oval
sparks coexisting in the same image. D, oval sparks remain elongated during their evolution, although the
earliest image is often more symmetric. For elongated sparks, the major axis is stably oriented, within
Z planes. Identifiers: n1129, images 31 and 41.
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

overlap of sparks from point sources. Both alternatives
require that multiple channels contribute to the elongated
events in caffeine.
Mobile sparks
The conclusion above is consistent with images, illustrated
in Figs 7—9, obtained with the time series facility of the
RCM8000 system. In this mode a narrow region in the
object is sampled repeatedly at 4 ms intervals, yielding
snapshots of the same spark at different times. These band
images were processed in essentially the same way as full
2D images. Figure 7A illustrates a succession of normalized
images from the same band in reference solution, and Fig. 7Bthe analysis by the locator program. The images feature the
typical evolution of round sparks. As shown in Fig. 7B, thesparks at top and bottom remain roughly circular from
beginning to end; their ‘major axis’, which is only slightly
longer than other diameters, assumes variable orientations,
indicative of basically symmetric events. The sparks remain
in the same location (confirmed by the evolution of their
centre of mass, not shown). Figure 7C, from the same fibre
in reference, features an oval spark with typical evolution.
G. Brum, A. Gonz�alez, J. Rengifo, N. Shirokova and E. R� úos J. Physiol. 528.3428
Figure 8. Propagating sparks
A and D, examples of sparks that invade an adjacent region as they grow (black arrows). B and E, theautomatic locator confirms the expansion, and shows other events that do not move (red arrows),
demonstrating a steady preparation. C and F, evolution of the abscissa of the centres of signal mass of two
sparks in each frame (plots colour-coded to arrows). The event in A appears to grow by recruiting a discrete
spark at time 16 ms, while the one in D grows more continuously. Images are normalized to F0. Identifiers:
0317b, image 130, and n1129, 52.
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

It starts small and circular, then grows rapidly in length
and remains elongated during most of its development and
decay, with the long axis pointing steadily in the direction
of the Z disk.
A different type of evolution of elongated sparks is
illustrated in Fig. 8. The sparks marked with black arrows
in Fig. 8A and D originate at the edge of the band, then
invade it progressively. The movement is demonstrated by
the evolution of the abscissa of the centre of mass, plotted in
black in Fig. 8C and F. Propagation was real, not an artifactof preparation drift or errant sampling, as witnessed by
‘landmark’ events in the same band (red arrows), which
were steady or moved slowly in the opposite direction.
Because the spreading events were only partially imaged,
whether they actually migrated or simply grew in length is
unclear.
Examples in Fig. 9 demonstrate actual migration of sparks.
The abscissa of the centre of mass of three events in Fig. 9Ais plotted in Fig. 9B, colour-coded to the arrows. ‘Black’ and‘green’ moved at •0·1 ìm ms¢, while other sparks in the
image remained steady. The migrating sparks became
elongated transiently as they moved and grew. The spark
marked by a black arrow in Fig. 9C first invaded the band
from the right edge. Later the peak of the spark moved into
the band as it grew. This evolution is represented three-
dimensionally in the graph of Fig. 9E, plotting the averagein the five central y pixels as a function of x (‘space’) and
time.
Propagating sparks were relatively infrequent. In two
experiments with caffeine, 500 frames with bands were
recorded and 120 were kept for further examination. There
were 410 sparks in the images kept. Twenty-one of those
propagated and had a landmark in the same frame.
Assuming that the records not kept had the same frequency
of events but no propagating sparks, the ratio of
propagatingÏsteady sparks would be 1%. This estimate
increases to about 2% if the requirement for a landmark in
the same frame is waived. We did not attempt to quantify
the frequency of mobile sparks in reference, but it appeared
to be lower, which is consistent with the fact that mobile
sparks tended to be elongated, and with the scarcity of
elongated events in reference.
DISCUSSION
The combination of rapid confocal scanning in two spatial
dimensions and time with automatic detection and
morphometry of large numbers of sparks allowed us to
characterize their spatio-temporal properties without the
limitations imposed by traditional line scanning. Several
new properties of sparks are documented.
In ‘reference’ conditions (which included a low [Mg¥]é, to
elicit a workable frequency of events) sparks were generally
circular in the image plane, reflecting presumably a
spherical three-dimensional shape. This resulted in an
essentially random distribution of the orientation angle, and
a low average eccentricity (Figs 4 and 5, and Table 1). The
average diameter in reference, 1·94 ìm, is substantially
greater than the full width at half-magnitude (FWHM)
usually reported in line scan studies. This is because in the
present work diameters were measured on suprathreshold
contours (Fig. 3) rather than at half-magnitude. Moreover,
in line scans FWHM is measured at the time of peak
fluorescence, while in the present work sparks were caught
at different times, most often after the peak, when spatial
spread continues to increase.
In the presence of caffeine sparks grew in spatial extent (in
agreement with previous observations with line scanning),
but the growth occurred largely within the plane of the
Z disks. Accordingly, the orientation angle concentrated
sharply around the value 0, and the eccentricity increased
and became ‘positive’ — reflecting the increased alignment
of the major axis.
The significance of elongated sparks
Three explanations were considered for elongated sparks.
One, first proposed by Gonz�alez et al. (2000a) in their studyof caffeine effects, is that the increase in frequency of
elongated events is due to a recruitment of multiple
channels to participate together in individual sparks. This
hypothesis justifies the effect of caffeine (as this drug
sensitizes channels to activation by Ca¥, thus facilitating
recruitment) and explains the alignment with the Z planes
that accompanies the increase in eccentricity.
The second alternative is a mechanical constraint that retains
Ca¥ in a ‘gutter’, or, more generally, any anisotropy, localized
or distributed, in the diffusion or binding properties (as
invoked by Cheng et al. 1996, to explain asymmetric sparks
in cardiac muscle). A distributed anisotropy, however, is
against the fact that most sparks are quite symmetric, even
in the presence of caffeine, and that symmetric and
elongated sparks occur some times at the same location in
successive images.
In support of the existence of a discrete barrier to diffusion,
we found an asymmetric distribution of widths, with a
sudden reduction in density at about 2·8 ìm. If this barrier
was the main cause of the elongation of sparks, amplitude
and width should be correlated with eccentricity. Indeed,
eccentricity should increase only upon reaching this
obstacle, which predicts a strong, albeit non-linear
correlation with event width. Amplitude should also be
correlated, because only sparks of a certain amplitude may
reach the barriers. Finally, the profile of concentration
(fluorescence) along the gutter should be characteristic of
diffusion away from an open source.
These predictions were contradicted by the results. There
was no correlation between amplitude and eccentricity
(consistent with the modest decay of spark amplitude with
caffeine), or between width and eccentricity. Perhaps most
significantly, the spatial dependence of fluorescence along
Ca¥ sparks in three dimensionsJ. Physiol. 528.3 429
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

G. Brum, A. Gonz�alez, J. Rengifo, N. Shirokova and E. R� úos J. Physiol. 528.3430
Figure 9. For legend see facing page.
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

the spark axis was generally flat or slanted rather than
symmetrically decaying from a central maximum.
The third explanation considered was random overlap of
circular sparks, resulting in images with the appearance of
elongated sparks. A first calculation of probability of such
random overlap can be made assuming that all sparks are
equal, so that the occurrence of sparks in contiguous locations
will be sufficient to generate the impression of an elongated
spark. Every image covers about 30 sarcomeres and 62 ìm
in the x direction. The overall frequency of events in caffeine,
•20 per image in the example of Figs 1—4, corresponds to •1
per image per 100 ìm of junction. The relative frequency of
events longer than 4 ìm (requiring overlap of at least two
sparks in a 4 ìm region) is less than the ratio: (conditional
probability of one event occurring in the 4 ìm region
surrounding another)Ï(probability of one event). If events
are independent, then this is equal to the (unconditional)
probability of an event occurring within a 4 ìm region, or
0·04 (1 ² 4 ìmÏ100 ìm), while their actual frequency was
0·13. That of a 6 ìm event (requiring overlap of at least
three sparks) was 0·0016, while their actual frequency was
0·02.
The above calculation may be refined by recognizing that spark
amplitudes and widths are distributed. For example, the long
sparks in Figs 1 and 3 could occur by superposition of round sparks,
of amplitudes between 0·8 and 1·3, and widths between 2 and
3 ìm. These conditions are satisfied by about 15% of the entire
population (e.g. Fig. 6). Taking this condition into account reduces
7-fold the probability of superposition giving rise to 4 ìm sparks,
and 50-fold that of 6 ìm sparks. Further reductions in probability
are required if it is recognized that more than two sparks should
overlap within 4 ìm to really appear as a single 4 ìm event.
When a spark was imaged at different times, radically
different evolutions were found for elongated and circular
ones. While the latter remained circular during their rise
and ebb, the elongated ones had various possibilities. Some
remained elongated, and when this happened, the long axis
remained essentially fixed in the transverse direction. Of
those that remained elongated, some kept growing in
length, invading adjacent areas at •0·1 ìm ms¢. Yet other
events actually moved in space, displacing their peak at a
similar rate.
Therefore, the possible mechanisms at play in the generation
of elongated sparks must involve the activation of multiple
sources (channels or their arrays). The transverse orientation
of their axis suggests recruitment of channels along
junctional arrays. Two chief modalities of recruitment
appear to be at play: a continuum mechanism, whereby
multiple channels are incorporated smoothly to the group
that generates the event (see Figs 7A and 8C and E), and a
discontinuous recruitment of individual channels, or their
(small) groups, each contributing a discrete, more symmetric
spark (see Fig. 8A).
The mechanism of propagation is likely to involve Ca¥-
induced Ca¥ release (CICR) (Endo et al. 1970; Fabiato,
1984). Interestingly, an earlier stochastic model of
propagation of release within couplons, in which a mobile
‘head’ of elevated Ca¥ provides the activation signal, yields
velocities of about 0·1 ìm ms¢, similar to those observed
here (Stern et al. 1997).
The present observations are also relevant to the mechanism
of termination of sparks. Some sparks migrate, implying
that the regions that activate first are also the first to close
(an inactivation, Sham et al. 1998). They, however,
constitute a minority. Sparks that are elongated usually
remain elongated during their decay phase (as in Fig. 7Cand D), which suggests that, if the spark involves a group ofchannels, then the whole activated array must inactivate
more or less simultaneously. This agrees with the conclusion
that release in sparks terminates abruptly, as if coming
from a single channel (Lacampagne et al. 1999). Such rapid
closure of channels was not reproduced by the couplon
model of Stern et al. (1997), in which release stopped by a
Ca¥-mediated inactivation. Allosteric interactions like
those reported by Marx et al. (1998) might provide a faster
mechanism of channel closure.
The increase in length induced by caffeine, or the extent of
observed propagation, should contain information regarding
the spatial extent of the sources. On average, caffeine
increases length of events by 0·41 ìm, a value that is well
within the length of most frog muscle couplons (Franzini-
Armstrong et al. 1999), hence requiring no more than
increased propagation within a couplon. In many cases,
Ca¥ sparks in three dimensionsJ. Physiol. 528.3 431
Figure 9. Migrating sparks
A, migrating sparks (green and black arrows), at both ends of a band. B, evolution of abscissa (x) of the
centre of mass for the sparks at the ends of the band (black and green) and one other near the centre (red),
which provides a landmark. The velocity reaches •0·1 ìm ms¢. Identifier: n1129, image 50. C, a spark
(black arrow) first grows towards the centre of the band as its intensity increases; later a local peak becomes
visible and moves in the same direction. A second spark (red arrow) is relatively steady. D, evolution of
abscissa of the spark’s centre of mass. Note that its movement accelerates (to 0·1 ìm ms¢) when the peak
separates from the edge of the band, probably because the centre of mass is better determined when the
whole event is taken into account. E, three-dimensional representation of the evolution of the spark in D.The central five pixels of the spark are averaged at every x position and in every band, then plotted vs. xand time. The plot shows spatial growth first, then movement of the local peak of fluorescence. Identifier
0317b, image 143. Images are normalized to F0.
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

however, sparks propagate over a micrometre or more (Fig. 9),
and events of several micrometres are easy to find (Figs 1
and 3). Because the length of frog couplons is at most
1·3 ìm, these cases require propagation over multiple
couplons.
The idea that propagation is restricted within one couplon
stems from simulations (Stern et al. 1997), and has
theoretical importance, as it explains why release activation,
an intrinsically self-sustaining process, remains always under
voltage control. The present observations, however, do not
require radically changing this concept. Sparks of multi-
couplon length are infrequent, and it would not be difficult,
just by increasing Ca¥ sensitivity in the model, to simulate
events that occasionally jump between couplons. In keeping
with this idea, there is other evidence of transmission of
activation over micrometre distances, especially in cardiac
muscle (e.g. Parker et al. 1996; Blatter et al. 1997). Bycomparison, skeletal muscle appears much more ‘uptight’ as
regards Ca¥ release. Interestingly, however, loss of control of
Ca¥ release by voltage in the presence of caffeine was
reported (Simon et al. 1989), reflecting perhaps uncontrollablecontagion of CICR under special circumstances.
The effect of event frequency
In the present experiments, the so-called reference
condition was actually non-physiological, altered to have a
finite, albeit low, frequency of spontaneous sparks. This was
done by lowering the free [Mg¥] to about 0·4 mÒ
(Lacampagne et al. 1998). The mechanism of this effect is
thought to be a relief from the inhibition exerted by Mg¥ on
the release channels. Such relief renders the channels more
susceptible to opening, which presumably occurs under the
influence of local Ca¥. On the other hand, caffeine is
believed to increase the sensitivity of channels to activation
by Ca¥, hence their proclivity to open ‘spontaneously’, or be
recruited into a spark. Therefore, both caffeine and low
[Mg¥] should increase the length and eccentricity of sparks,
as both should promote activation, and recruitment of
additional channels.
The study that will test whether the effects of caffeine and
low [Mg¥] are similar, a determination of spark
morphology at different [Mg¥], has not been done. In
support of this idea, however, it appeared that spark length,
eccentricity and orientation grew in parallel with the overall
frequency of events per image. This was seen best when the
frequency of events varied substantially in the course of
time. In one experiment that had eccentricity of 0·00 (0·01)
in reference, while a series of 20 images was being recorded
in the presence of caffeine, the frequency changed
monotonically from 36 to 19 events per image. For the
events in the first 10 images, when frequency averaged 32
events per image, eccentricity was 0·19 (0·02), while in the
second half of the sequence (when the frequency was 23 per
image) the eccentricity was 0·08 (0·02). When the set of
events was divided into four groups, each gathered during
five successive images, eccentricity correlated well with
frequency (r  was 0·85) but amplitude was essentially
constant (r  = 0·22).
In conclusion, the parallel evolution of eccentricity and
frequency during uncontrolled changes suggests that the
increase in eccentricity and event length associated with the
presence of caffeine is not specific for this agonist, but a
consequence of an increase in the channels’ propensity to
open.
The nature of ‘standard’ sparks
Sparks that are elongated, or grow in length or move during
their evolution, require the involvement of multiple
channels, opening simultaneously or sequentially. While in
some cases propagation of sparks appears to occur by
discrete jumps, and could be explained by recruitment of a
few channels (or their groups), in others (as in Fig. 9E) it iscontinuously graded, suggesting the involvement of tens of
channels. That the sparks studied here were elicited by non-
physiological stimulants of channel activity may suggest
that the involvement of multiple channels requires such
artificial enhancers, and is not a feature of normal or
standard sparks. On the other hand, a previous study
(Gonz�alez et al. 2000a) showed that caffeine modifies sparks
quantitatively, increasing for instance the frequency of
sparks of large width, rather than inducing qualitatively
different events that do not occur in reference. This suggests
that standard sparks (i.e. those induced by normal stimuli in
physiological internal solutions) should also involve multiple
channels. The same conclusion was reached by us, on
entirely different grounds, in two other publications (R� úos etal. 1999; Gonz�alez et al. 2000b).
Because membrane depolarization also operates by
increasing the propensity of release channels to open (an
effect compared to the removal of Mg¥ inhibition; Lamb,
2000), a similar cooperative activation of channel arrays
might be taking place when the membrane is depolarized,
either experimentally or physiologically during an action
potential. A study of the effect of membrane voltage on the
spatio-temporal morphology of sparks is clearly called for,
and is in progress.
Blatter, L. A., H�user, J. & R�úos, E. (1997). Sarcoplasmic reticulumCa¥ release flux underlying Ca¥ sparks in cardiac muscle.Proceedings of the National Academy of Sciences of the USA 94,4176—4181.
Cheng, H., Lederer, W. J. & Cannell, M. B. (1993). Calciumsparks: elementary events underlying excitation-contractioncoupling in heart muscle. Science 262, 740—744.
Cheng, H., Lederer, M. R., Xiao, R. P., Gomez, A. M., Zhou, Y. Y.,Ziman, B., Spurgeon, H., Lakatta, E. G. & Lederer, W. J.(1996). Excitation-contraction coupling in heart: new insights fromCa¥ sparks. Cell Calcium 20, 129—140.
Cheng, H., Song, L. S., Shirokova, N., Gonz �alez, A., Lakatta,E. G., R� úos, E. & Stern, M. D. (1999). Amplitude distribution ofcalcium sparks in confocal images. Theory and studies with anautomatic detection method. Biophysical Journal 76, 606—617.
G. Brum, A. Gonz�alez, J. Rengifo, N. Shirokova and E. R� úos J. Physiol. 528.3432
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

Endo, M., Tanaka, M. & Ogawa, Y. (1970). Calcium induced releaseof calcium from the sarcoplasmic reticulum of skinned skeletalmuscle fibres. Nature 228, 34—36.
Fabiato, A. (1984). Dependence of the calcium-induced release formthe sarcoplasmic reticulum of skinned skeletal muscle fibres from thefrog semitendinosus on the rate of change of free Ca¥ at the outersurface of the sarcoplasmic reticulum. Journal of Physiology 353,56P.
Franzini-Armstrong, C., Protasi, F. & Ramesh, V. (1999). Shape,size, and distribution of Ca¥ release units and couplons in skeletaland cardiac muscles. Biophysical Journal 77, 1528—1539.
Gonz�alez, A., Kirsch, W. G., Shirokova, N., Pizarro, G., Stern,M. D. & R�úos, E. (2000a). The spark and its ember. Separately gatedlocal components of Ca¥ release in skeletal muscle. Journal ofGeneral Physiology 115, 139—158.
Gonz�alez, A., Shirokova, N., Kirsch, W. G., Pessah, I. N., Cheng,H., Stern, M. D., Pizarro, G., Brum, G. & R�úos, E. (2000b).Involvement of multiple channels in calcium sparks of skeletalmuscle. Proceedings of the National Academy of Sciences of theUSA 97, 4380—4385.
Herrmann-Frank, A., L�uttgau, H.-C. & Stephenson, D. G. (1999).Caffeine and excitation-contraction coupling in skeletal muscle: astimulating story. Journal of Muscle Research and Cell Motility 20,223—237.
Kawanishi, T., Asou, H., Kato, T., Uneyama, C., Toyoda, K.,Ohata, H., Momose, K. & Takahashi, M. (1994). Ratio-imaging ofcalcium waves in cultured hepatocytes using rapid scanningconfocal microscope and indo-1. Bioimages 2, 7—14.
Klein, M. G., Cheng, H., Santana, L. F., Jiang, Y. H., Lederer,W. J. & Schneider, M. F. (1996). Two mechanisms of quantizedcalcium release in skeletal muscle. Nature 379, 455—458.
Lacampagne, A., Klein, M. G. & Schneider, M. F. (1998).Modulation of the frequency of spontaneous sarcoplasmic reticulumCa¥ release events (Ca¥ sparks) by myoplasmic [Mg¥] in frogskeletal muscle. Journal of General Physiology 111, 207—224.
Lacampagne, A., Ward, C. W., Klein, M. G. & Schneider, M. F.(1999). Time course of individual voltage-activated Ca¥ sparksrecorded at ultra-high time resolution in frog skeletal muscle.Journal of General Physiology 113, 187—198.
Lamb, G. (2000). Excitation-contraction coupling in skeletal muscle.Comparison with cardiac muscle. Clinical and ExperimentalPharmacology and Physiology 27, 216—224.
Leong, P. & Maclennan, D. H. (1998). Complex interactions betweenskeletal muscle ryanodine receptor and dihydropyridine receptorproteins. Journal of Biochemistry and Cell Biology 76, 681—694.
Marx, S. O., Ondrias, K. & Marks, A. R. (1998). Coupled gatingbetween individual skeletal muscle Ca release channels (ryanodinereceptors). Science 281, 818—821.
Niggli, E. (1999). Localized intracellular calcium signaling in muscle:calcium sparks and calcium quarks. Annual Review of Physiology61, 311—335.
Parker, I., Zang, W. J. & Wier, W. G. (1996). Ca¥ sparks involvingmultiple Ca¥ release sites along Z-lines in rat heart cells. Journal ofPhysiology 497, 31—38.
R�úos, E., Stern, M. D., Gonz �alez, A., Pizarro, G. & Shirokova, N.(1999). Calcium release flux underlying Ca¥ sparks of frog skeletalmuscle. Journal of General Physiology 114, 31—48.
Schneider, M. F. (1999). Perspective: Ca¥ sparks in frog skeletalmuscle: generation by one, some, or many SR Ca¥ release channels?Journal of General Physiology 113, 365—372.
Sham, J. S., Song, L. S., Chen, Y., Deng, L. H., Stern, M. D.,Lakatta, E. G. & Cheng, H. (1998). Termination of Ca¥ release bya local inactivation of ryanodine receptors in cardiac myocytes.Proceedings of the National Academy of Sciences of the USA 95,15096—15101.
Shirokova., N., Gonzalez, A., Kirsch, W. G., Rios, E., Pizarro,G., Stern, M. D. & Cheng, H. (1999). Calcium sparks: releasepackets of uncertain origin and fundamental role. Journal ofGeneral Physiology 113, 377—384.
Shirokova, N. & R�úos, E. (1997). Small event Ca¥ release: a probableprecursor of Ca¥ sparks in frog skeletal muscle. Journal ofPhysiology 502, 3—11.
Simon, B. J., Klein, M. G. & Schneider, M. F. (1989). Caffeine slowsturn-off of calcium release in voltage clamped skeletal muscle fibers.Biophysical Journal 55, 793—797.
Stern, M. D., Pizarro, G. & R�úos, E. (1997). Local control model ofexcitation-contraction coupling in skeletal muscle. Journal ofGeneral Physiology 110, 415—440.
Tanaka, H., Nishimaru, K., Sekine, T., Kawanishi, T.,Nakamura, R., Yamagaki, K. & Shigenobu, K. (1997). Two-dimensional millisecond analysis of intracellular Ca¥ sparks incardiac myocytes by rapid scanning confocal microscopy: increasein amplitude by isoproterenol. Biochemical and Biophysical ResearchCommunications 233, 413—418.
Tsien, R. Y. (1990). Laser scanning confocal fluorescence microscopyat video rate (30 framesÏsec) with dual-wavelength emissionrationing for quantitative imaging of intracellular messages.Proceedings of the Royal Microscopical Society 25, S53.
Tsugorka, A., R� úos, E. & Blatter, L. A. (1995). Imagingelementary events of calcium release in skeletal muscle cells. Science269, 1723—1726.
Acknowledgements
We thank Dr Gonzalo Pizarro for stimulating discussions. This
project was supported by grants from National Institutes of Health
to E.R. (R01-AR32808) and N.S. (R01-AR45690).
Corresponding author
E. R� úos: Department of Molecular Biophysics and Physiology, Rush
University, 1750W. Harrison Street, Chicago, IL 60612, USA.
Email: [email protected]
Author’s present address
N. Shirokova: Department of Pharmacology and Physiology,
UMDNJ, New Jersey Medical School, Newark, NJ 07103, USA.
Ca¥ sparks in three dimensionsJ. Physiol. 528.3 433
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from

2000;528;419-433 J. Physiol.
Gustavo Brum, Adom González, Juliana Rengifo, Natalia Shirokova and Eduardo Ríos skeletal muscle
sparks in frog2+Fast imaging in two dimensions resolves extensive sources of Ca
This information is current as of January 9, 2007
& ServicesUpdated Information
http://jp.physoc.org/cgi/content/full/528/3/419including high-resolution figures, can be found at:
Permissions & Licensing
http://jp.physoc.org/misc/Permissions.shtmlits entirety can be found online at: Information about reproducing this article in parts (figures, tables) or in
Reprints http://jp.physoc.org/misc/reprints.shtml
Information about ordering reprints can be found online:
at LIBRARY OF RUSH UNIVERSITY on January 9, 2007 jp.physoc.orgDownloaded from