function of the vertebrate kidney, especially by a study of the
TRANSCRIPT

433
J. Physiol. (I940) 97, 433-452 6I2.463
THE FORMATION OF URINE IN THE AMPHIBIANAND MAMMALIAN KIDNEY
BY P. ELLINGERFrom the Lister Institute, London
(Received 31 December 1937)
THE present communication is concerned with the function of theamphibian and the mammalian kidney as revealed by intravital micro-scopic examination of its responses to certain dyes and drugs, The aimof the experiments has been to discover whether the tubules have asecretory function in addition to their reabsorptive activity, and if so,whether this is a normal function of the proximal convoluted tubulesor one which comes into play only under unusual conditions.
The mechanism of glomerular filtration and tubular reabsorption,postulated by the theory of Ludwig and Cushny, has been largely re-cognized [cf. Smith, 1937]. It has been confirmed in recent research on thefunction of the vertebrate kidney, especially by a study of the chemicalcomposition and the physico-chemical properties of the urine during itspassage through different parts of the urinary tract, which has been carriedout by Richards and his co-workers [Richards, 1929, 1938] and by theintravital microscopic investigations on the frog's kidney by Ellinger &Hirt [1929a, b, c, 1930a, b, 1931] and Ellinger [1934a, b, c, 1935]. Theevidence in favour of the secretory function is far less complete. Apartfrom some investigations on the excretion of dyestuffs which yieldedcontradictory results, it rests mainly on the classical experiments ofNussbaum on the frog's kidney [1878, 1879] and is supported by thecorresponding experiment of Ghiron on the mouse's kidney [1912, 1913]and by the observation that injected acid is excreted through the epi-thelium of the proximal tubules of winter frogs [Ellinger & Hirt, 1930 a].
The results of Nussbaum's experiment showing the cessation of urineformation after ligation of the renal arteries in frogs and its restorationby injection of urea has been confirmed by many workers, but inter-preted in different ways. Some investigators believed that the urea
PH. xCvII. 28

P. ELLINGER
opens collaterals between the renal portal vein and the glomerularcapillaries and so restores the glomerular circulation and filtration. Ifurea acts in this way the value of the experiment for the theory of urineformation would be restricted, while if it stimulates an excretion of urinein the proximal tubules the experiment would lend strong support to thetheory of Bowman and Heidenhain. It was felt that a repetition ofNussbaum's experiment, using intravital microscopy, might decide whichof the two interpretations is correct, since this method allows directvisual observation of all parts of the frog's kidney during the wholecourse of the experiment.
A similar re-examination might also elucidate the mechanism ofGhiron's experiment. Ghiron abolished urine formation in mice bysevering the spinal cord and thus lowering the blood pressure. Urineformation was restored by injection of urea. His technique has beencriticized, but his original observations have never been fully repeated.
This paper presents the results of an investigation of Nussbaum'sand Ghiron's experiments by intravital microscopy and includes furtherobservations on the elimination of certain dyes, urea and acid from thenormal amphibian and mammalian kidney. The normal function of themammalian kidney has not yet been studied by intravital microscopy.Since their experiments gave evidence for the existence of an excretoryfunction of the proximal tubules, the question arose whether it is to beregarded as part of the normal mechanism of urine formation.
METHODSThe experiments were carried out on frogs and rats. Those on frogs
with abolished glomerular function were made in May, June and July,the others at various times during three consecutive years. For thestudy of the elimination of dyes fluorescein and acriflavin were used.They were injected in 0.1% concentration in Ringer's solution. Ureawas injected in 5% solution and acid as sodium bisulphate in 1-65%concentration. In order to examine repeated stimulation of tubularexcretion two different methods were adopted. In the one the effectsof either bisulphate or urea injected repeatedly at half-hourly or hourlyintervals were studied on the elimination of previously injected fluore-scein. In the other the excretion of fluorescein was examined after acourse of daily injections, for ten to twenty days, of either bisulphate orurea.
In frogs all observations were carried out on the left kidney whiledyes and drugs were injected into the lymph sac of the right leg. In rats
434

URINE IN AMPHIBIAN AND MAMMALIAN KIDNEY 435
either of the kidneys was used for observation and all injections weremade into the peritoneal cavity. The special techniques employed wereas follows:
Experiments on frogs. R. esculenta, of both sexes, from Hungary, weighing 70-100 g.were kept cold in the dark with access to running water but without food. For intravitalmicroscopic examination they were anaesthetized with urethane percutaneously. Theabdominal vein was exposed and cut between ligatures. The abdominal muscles wereincised and removed and the peritoneum was opened along the lateral edge of the leftkidney, which was thus exposed to the objective. The method has been described in detailby Ellinger & Hirt [1929b]. In the experiments in which the glomerular function wasabolished by arterial ligation (Nussbaum's experiment), care was taken to ensure the cuttingoff of all possible sources of arterial supply. Under urethane the dorsal part of the portalvein was ligated and divided. In female frogs the ovaries were first removed. The kidneyswere turned so as to bring the aorta and iliac arteries to the surface. Under a dissectingmicroscope all branches from these vessels to both kidneys as well as the mesenteric arterieswere cut between ligatures. The kidney was turned back and any still pulsating branchesreaching the kidney from the vesical or the ureteric arteries were looked for. Only in twoof eleven experiments were such branches found. They came from the iliac artery along theureter and were also cut. All glomeruli were then examined under the intravital microscopeand re-examined an hour later. Four frogs, in which a few glomeruli were seen to be active,were discarded. In the remaining seven cases (five males, two females) all glomeruli wereinactive. The circulation in the veins was not affected.
Experiments on rats. Rats of 150-200 g. of both sexes from the Lister Institute's stockwere used. The surface of the rat's kidney shows only proximal and occasionally a few distaltubules. The glomeruli and all other distal tubules are situated more deeply so that, inorder to expose them, the surface of the kidney must be shaved off. This operation causesserious bleeding. According to a suggestion of Hirt, it can be stopped by employing a heatedknife or by applying boiling Ringer's solution. The following operative technique wasdeveloped: in a preliminary operation under ether the capsule of the exposed kidney isincised, a shaving taken from the surface with a hot scalpel, and the wound closed withsilk. Two or more days later, 1 ml. of a 3% chloral hydrate solution per 100g. rat is givenintraperitoneally, the kidney is fixed on a plate with a slit for the hilum and the plate thenfixed to a heated stage which allows the air round the rat to be heated to 370 C. The plateprevents respiratory movements affecting the kidney. Ringer's solution at 38-40' C. isused for irrigation.
For Ghiron's experiment rats of 200 g. weight were used. A few days after the surfaceof the kidney had been shaved off the rats were anaesthetized with chloral hydrate andether. The cervical vertebrae were exposed and identified, the head was bent forward asfar as possible and a fine double-edged scalpel inserted into the appropriate intervertebralspace in order to cut the cord at the fifth or sixth segment. Four of ten animals survivedthe operation, the others dying from respiratory failure from shock which might have beenavoided by adequate narcosis. The best anaesthetic is gas, but this was unfortunately notavailable. The wound was closed, the animal wrapped in cotton-wool and kept in a hot roomat 370 C. Next day microscopic examination of the kidney revealed a very slow circulationin the intertubular capillaries. No glomeruli were found to be active in three animals, inone there was a slow circulation in a few glomerular capillaries. The glomerular capsuleswere empty and only a few greenish lyochromes were visible.
28-2

P. ELLINGER
RESULTSExperiments on frogs
In frogs with intact arterial supply to the kidney only a certain pro-portion of the glomeruli were active. Greenish fluorescent lyochromes[Ellinger, 1938] were seen only in the capsules of active glomeruli andin the lumen of tubules derived from them. Spontaneous changes inactivity of the glomeruli occurred only occasionally. The elimination offluorescein and acriflavin and the staining of the different cells of thekidney with these dyes did not vary in the experiments carried out insummer and winter. Seasonal differences in the kidney function noted inHeidelberg [Ellinger & Hirt, 1929c, 1930a, 1931] were not observed inthis country.
When the arterial supply to the kidney was abolished the glomerularcapsules did not contain lyochromes and none was seen in the tubularlumen.
Elimination of fluorescein. 1-2 min. after the injection of fluorescein (0-1-2 0 mg. perfrog) the dye was seen in the arterioles. Almost simultaneously it appeared in the glomerularcapsules with a greenish fluorescence which according to Ellinger & Hirt [1930b] corre-sponds to pH 7.0. 3-5 min. after injection, the lumen and the epithelium of the proximaltubules became brighter with a greenish yellow fluorescence (pH 6-5) and after another5 min. a rather yellow fluorescence (pH 6-0) appeared in the distal tubules spreading witha slightly greener tint (pH 6.5) into the epithelium there. 15 min. after the injection abright yellow fluorescence (pH 5-5) was seen in the collecting tubules and ureter, and in theepithelium of the former the nuclei were stained and the dye separated out in the cyto-plasm. The increased fluorescence in the lumen and cells of the convoluted tubules lasted1-3 hr. and that in the cells of the collecting tubules for about 10 hr. These changes wereseen only in tubules connected with active glomeruli. Therefore the dye must have comefrom the glomeruli and entered the epithelium only from the lumen (Figs. 1, 2). After deathof the cell the dye spread through the whole tissue.
These observations show that in frogs with intact arterial supply tothe kidney the dye is eliminated in the glomeruli in about the same con-centration and at the same pH as it is present in the plasma and thatthere is a slight concentration and acidification in the proximal and astronger one in the distal tubules. Compared with the results obtainedin Heidelberg [Ellinger & Hirt, 1929c, 1930a, 1931] concentration andacidification were less pronounced than in summer frogs and far more sothan in winter frogs.
In five frogs whose arterial supply to the kidney had been abolished, injected fluorescein(0-2-1-0 mg. per frog) was seen after 2 min. in the renal portal blood and almost at the sametime in the epithelium of the proximal tubules with a rather yellow fluorescence (pH 6-0).15 min. after the injection a fainter yellowish fluorescence (pH 5-0) appeared in the lumenof the proximal tubules. The glomerular capsules and the lumen as well as the epitheliumof the distal tubules remained completely unstained. This picture lasted for at least 3-4 hr.
436

URINE IN AMPHIBIAN AND MAMMALIAN KIDNEY 437
These results indicate that a small amount of acid fluid containingdye is excreted in the proximal tubules, but that there is not sufficientexcretion of water to push forward the dye into the lumen of the lowerurinary tract. A similar behaviour has been observed in experimentsmade in Heidelberg on summer frogs with abolished arterial supply tothe kidney.
Elimination of acriflavin. In frogs with intact arterial supply to thekidney the dye was eliminated in the glomeruli in low concentrationand partly reabsorbed and concentrated in the tubules.
Acriflavin (0-2-2-0 mg. per frog) was seen in the blood about 5 min. after its injection.The fluorescence was not so pronounced as that observed after fluorescein injection. A littlelater the nuclei of the blood corpuscles of the glomerular epithelium, and of the vascularmuscle cells were stained. The glomerular capsules brightened distinctly but not intensely.The lymph spaces bordering the epithelium of the proximal tubules fluoresced very brightly,those bordering the distal tubules to a much less degree. The cells were stained at firstdiffusely, then their nuclei took up the stain temporarily and during the next hour or twothe stain was gradually aggregated into discrete granules in the cytoplasm. In the mean-time the pericellular spaces had become darkened. While the lumen of the whole tubularsystem down to the ureter was fluorescent the epithelium of the collecting tubules was notstained as when fluorescein was injected.
Acriflavin was not so markedly concentrated in its passage downwardsas fluorescein. It was seen in the lumen only oftubules derived from activeglomeruli, but was contained in the epithelium of all convoluted tubulesalike. This fact and the appearance of fluorescence in the pericellularlymph spaces before it reached the cells suggested that the dye hadentered the cells from the lymph and blood and not from the lumen.
The fluorescence of the lumen lasted 10-24 hr., but acriflavin particles were present inthe cells many days later, even after the death of the cell. The mechanism of staining of thetubular epithelium, observed in these experiments, has also been noted in experimentsmade in Heidelberg with acriflavin on summer frogs.
When the arterial supply to the kidney had been abolished noacriflavin was excreted.
The dye (2 mg. per frog) appeared in the blood 7-10 min. after its injection and stainedthe nuclei of the blood corpuscles, of the vascular muscle cells, and of the glomerularepithelium. Almost at the same time the edges of the epithelium of the proximal tubulesbecame fluorescent and a little later the dye passed from the circumference into the interiorof the cells, became diffused in the cytoplasm and stained the nuclei temporarily. About2 hr. later it had formed large particles in the cytoplasm. No dye was seen in any portionof the lumen, a result contrary to that obtained when the glomeruli were active. Only inthe proximal tubules were the cells stained.
Effect of a single injection of urea or sodium bisulphate. In frogs withintact arterial supply to the kidney, urea activated all inactive glomeruliand stimulated both mechanisms of urine formation, e.g. glomerular

P. ELLINGER
dFig. 1 Fig. 2
It
Fig. 3 Fig. 4
b
AWa .C>$
~I '212
Fig. 5 Fig. 6
Fig. I Fig.2
Key to Plate I.

THE JOURNAL OF PHYSIOLOGY, VOL. 97, No. 4 PLATE I
Fig. 1.
Fig. 3.
Fig. 5.
Fig. 2.
Fig. 4.
Fig. 6.
To face p. 438
THE JOURNAL OF PHYSIOLOGY, VOL. 97, No. 4
Fig. 7.
Fig. 9.
Fig.3-
Fig. 10.
Fig. 1I.
PLATE II
Fig. 12,

URINE IN AMPHIBIAN AND MAMMALIAN KIDNEY 439
a
bb Q
~~~2(V~~~
Wc X~~Fig. 7 Fig. 8
c ~~~~~122 "1114~a 78
4b 2 1
bFig. 9 Fig. 10
Fig. 11 Fig. 12Key to Plate II.

440 P. ELLINGER
filtration and tubular excretion. With bisulphate there was some glo-merular filtration but the acid was mainly excreted through the proximaltubules. Similar observations have been made on winter frogs in Heidel-berg [Ellinger & Hirt, 1930 a]. The results are illustrated by the followingobservations.
When urea (0.25 g. per frog) was given at the height of elimination of fluorescein (2 mg.per frog previously injected), all inactive glomeruli became instantly active and thefluorescence in the lumen of the proximal tubules became a much lighter yellow (pH 5 0).This change in colour extended rapidly into the lumen of the distal tubules and beyond.The point where the greenish fluorescent fluid (pH 7-0) from the glomerular capsulesmet the more acid fluid in the proximal tubules could be clearly distinguished. In theseexperiments the dye was eliminated in 1-1k hr. in contrast to the customary 5-6 hr. takenin the absence of urea even when only small doses of dye were injected. When a singleinjection of sodium bisulphate (16.5 mg. per frog) was given after fluorescein (2 mg. perfrog) the fluorescence of the glomerular capsules became instantaneously lighter yellow, andat the same time still a lighter yellowish tint was seen in the epithelium of the proximaltubules and passed into their lumen. This light yellow colour moved quickly down intothe lower parts of the urinary tract and after 20-30 min., the tint of the fluorescence inthe whole urinary tract was as it had been before the injection of bisulphate.
In frogs with abolished arterial supply to the kidney the glomeruliwere not activated by urea, but the excretion of acid urine was stimulatedtbrough the epithelium of the proximal tubules. The urine containedfluorescein but no acriflavin when either of these dyes was previouslyinjected.
These results were obtained on five frogs in which urea (0*25 g. per frog) was injected1-2 hr. after the dyes had been given. In the case of acriflavin the injection of urea causedno change of the staining and no dye was visible in the lumen at any time. In the case offluorescein, immediately after the injection of urea, the epithelium of the proximal tubulesbecame brighter with a bright yellowish fluorescence (pH 4.5) in their lumen. This lightyellow colour rapidly passed into the lumen of the distal tubules and more slowly into thecollecting tubules and the ureter. Their epithelium, however, remained unstained. Allglomeruli remained inactive and there was no fluorescence in their capsules. 1-2 hr. afterthe injection of urea the fluorescence began to fade in the epithelium and lumen of theproximal tubules and later from the lumen of the tubules lower down (Figs. 3-7).
E;ffect of repeated injections of urea or sodium bisulphate. Repeatedinjections of either sodium bisulphate or urea at short intervals lead todamage of the epithelium of the proximal tubules which lose theirexcretory function and further injections of either of these drugs maythen be fatal.
For instance, if, after fluorescein (2 mg. per frog), bisulphate (1.65 mg. per frog) isinjected every hour, or half hour the tubular excretion of acid stops after the third or fourthinjection and the frog soon dies. With urea the excretion through the proximal tubules failsafter the sixth or seventh dose (0.25 g. per frog), but the animal may not be killed even byone or two further injections.

URINE IN AMPHIBIAN AND MAMMALIAN KIDNEY 441
When the arterial supply to the glomeruli is abolished even 0-025 g. of urea causesexcretion through the proximal tubules up to the fourth and fifth dose. Then excretionstops and the animal dies if a further dose is given showing that when there is no othermeans of elimination and tubular function is stimulated the injection even of urea maybe lethal.
Daily injections of either urea or bisulphate also produce destructionof the epithelium of the proximal tubules. They lose their ability forselective absorption of water, alkali and dyes and also their excretoryfunction.
This was revealed when fluorescein was injected. Otherwise urea appeared to be harm-less, whereas daily injections of bisulphate caused local damage at the site of injection, i.e.oedema, haemorrhage, epidermolysis and paralysis of the sciatic nerve, and led usually lateron to general oedema. The intravital microscopic examination of the kidneys showed aconsiderable reduction in the amount of green and yellow fluorescent lyochromes. Theeffect was especially pronounced after bisulphate. After daily injections of this substanceoccasionally a white bright fluorescence was seen in the epithelial cells of the proximaltubules while the intercellular lymph spaces were not fluorescent (Fig. 8). A similar whitefluorescence has often been observed in glomeruli destroyed by parasites and could beattributed to a high concentration of calcium salts. The same explanation accounted forthe finding in these experiments as shown by micro-calcium determination. When kidneysof frogs, after a course of daily injections of bisulphate, did not show definite signs of calci-fication a tendency for such form of damage could be demonstrated by the following pro-cedure. The kidneys were removed in a sterile operation, kept for 2-4 days at 370 C. in a saltsolution containing 8 mg. Ca-ions and 5 mg. P as inorganic phosphate per 100 ml. [Robison& Rosenheim, 1934] and then stained with silver nitrate or orange G; they showed markedcalcification of the epithelium of the proximal tubules and sometimes of the walls of therenal arteries. This was never found in kidneys of untreated control frogs.
When fluorescein was injected into frogs which had been given daily injections ofbisulphate the dye was visible within a few minutes in the blood and in the glomerularcapsules with the greenish tint ofpH 7-0 as seen in untreated animals. When the dye ap-peared in the lumen of the proximal tubules it had the same tint and concentration andremained so to the end of the experiment. In this case there was no fluorescence in the cells.The fluorescein reached the distal tubules only a little concentrated, their epithelium andthat of the tubules beyond being stained as in normal frogs. The urine in the ureter showedalso the greenish tint ofpH 7 0, but the increased intensity of the fluorescence indicatedthat the dye was present in a higher concentration than in the lumen of the proximaltubules. After the daily administration of urea the subsequent injection of fluoresceinshowed a similar but less pronounced abnormality in the elimination of the dye. Littleacidification or concentration was seen in the proximal tubules. If in such experiments theusual dose of urea was given at the height of elimination of the dye some staining of theepithelium occurred but no excretion through the epithelium into the lumen ofthe proximaltubules. Glomeruli previously inactive became active and the dye in the tubules was moredilute.
In three frogs which had been given daily injections of bisulphate the glomerularfunction was abolished by arterial ligation. The subsequent injection of fluorescein causedonly slight diffuse staining of the epithelium of the proximal tubules and a little greenishfluorescence (pH 7 0) was seen in their lumen. The yellow fluorescence in the lumen of theproximal tubules which was observed in the corresponding experiments on frogs which hadnot undergone treatment with daily injections of bisulphate was absent. The injections of

442 P. ELLINGER
urea caused the formation of very dilute urine containing fluorescein in about the sameconcentration and of the same tint as in the blood. Small amounts of bisulphate (8 mg. perfrog) did not do this, larger amounts (16-5 mg. per frog) caused death.
Experiments on ratsIn normal rats greenish fluorescent lyochromes were observed only
in the lumen of capsules of active glomeruli and of tubules derived fromthem [cf. Ellinger, 1938]. A much smaller proportion of the glomeruliwas active than in frogs and spontaneous changes were infrequent. Inprinciple the reaction of the rat's kidney to injected dyes and drugs wassimilar to that of the frog's kidney. This was true for the experimentswith intact and abolished glomerular circulation.
Elimination offluore8cein. Fluorescein (0-25-2 mg./100 g.) injected into the peritonealcavity immediately caused the blood in the kidney to become bright and in less than 1 min.the active glomeruli showed an increased greenish fluorescence (pH 7 0) in the capsules.Almost at the same time the proximal tubules connected with them became brighter andtheir greenish fluorescence became yellower (pH 6 5). Their epithelium showed the samecolour. The dye was here more concentrated than in the capsules and soon became more soin their epithelium than in their lumen. After 2-5 min. the dye appeared in still moreconcentrated form in the lumen of the distal tubules, and also, in a fainter degree, in theirepithelium. This picture lasted j-5 hr. according to the dose given. Then the glomerularcapsules grew darker, followed by the lumen of the convoluted tubules, the epithelium ofthe distal, and finally of the proximal tubules (Figs. 9, 10).
These observations show that the dye is eliminated in the activeglomeruli, slightly concentrated, acidified, and partly reabsorbed in theproximal tubules. In the distal tubules it is seen in a much higher con-centration and with a yellower tint. The urine is, however, less acidifiedthan in frogs.
In rats whose glomerular circulation was abolished, fluorescein (2 mg./100 g.) appearedin the renal vessels 20-30 min. after its peritoneal injection. At about the same time thelymph spaces bordering the cells of the proximal tubules became gradually brighter andfrom thence the dye passed into the cells. About 1 hr. later the dye was distributed diffuselythroughout the cytoplasm, both here and in the cells of the distal tubules. There was nonein the lumen and no dye was found in the bladder at the end of the experiment.
These results show that when the glomerular circulation is abolishedno urine is formed and the tubular epithelium takes up the stain fromthe lymph spaces.
Elimination of acriftavin. As in frogs acriflavin stains the tubularepithelium independently of its elimination. Entering the cells from thelymph spaces the dye is eliminated in active glomeruli in low concen-tration and slightly concentrated during its passage through the tubules.This is shown by the following experiments.
Acriflavin (0.5-4 mg./100 g.) was not visible in the blood of the normal rat until 10-S15min. after its injection, when the nuclei of the leucocytes and of the arteriolar muscle cells

URINE IN AMPHIBIAN AND MAMMALIAN KIDNEY 443
were also stained. A few minutes later the dye was seen in the capsule, in the nuclei of theglomerular epithelium and also in the lymph spaces bordering the epithelium of theproximal tubules. From the lymph spaces it entered the cells, at first staining the cytoplasmdiffusely, after j-3 hr. it stained temporarily the nuclei, and 2-5 hr. later it became aggre-gated into relatively large particles in the cytoplasm. When it was entering the epitheliumfrom the lymph spaces the dye could also be seen coming down from the glomeruli in thelumen of the proximal tubules. Some may have been reabsorbed there, but the greater partreached the lumen of the distal tubules in but slightly greater concentration. Here theepithelium took up the stain only slowly and not intensely, either from the lumen or fromthe lymph or blood vessels. Only the epithelium of those distal tubules which had pre-viously shown green fluorescence, and were connected with active glomeruli, contained thedye, whereas the epithelium of all the proximal tubules was stained. This picture lasted formany hours, even if small doses of dye had been injected (Figs. 11, 12).
In one rat with abolished glomerular circulation acriflavin (4 mg./100 g.) was seen about30 min. after its injection in the epithelium of the proximal and later in that of the distaltubules. This picture remained unchanged for many hours and no dye could be observedin the tubular lumen or in the bladder.
There is therefore no elimination of acriflavin when the glomerularcirculation is abolished.
Effects of single injections of urea or sodium bisulphate. As in frogs,a single injection of urea activated all visible inactive glomeruli andstimulated simultaneously excretion of acid urine through the epi-thelium of the proximal tubules. Fluorescein or acriflavin previouslyinjected, and staining the epithelial cells of the proximal tubules, leftthe cells while the excretory function was working. Acid (sodiumbisulphate) when injected was also at least partly excreted throughthe proximal tubules. These facts are revealed by the following ob-servations.
When a large dose of urea (0-25 g./100 g.) was injected into normal rats after fluorescein(2 mg./100 g.) had been given, and its elimination had reached its height, instantly all theglomeruli in view and all their capillaries became active and charged with dye. Within afew minutes, the colour in the epithelium of the proximal tubules began to fade and theirlumen as well as that of the distal tubules became bright and of a lighter yellow colour.Occasionally the lymph spaces bordering the cells would suddenly fluoresce intensely fora few seconds. In about an hour the fluorescein had all been eliminated, far quicker thanin the corresponding experiments without urea. When sodium bisulphate (16-5 mg./100 g.)was injected under similar conditions the fluorescence of the capsules became more yellowand that in the lumen of the proximal tubules became suddenly very bright and assumeda pale yellow tint (pH 5.5). This colour moved down quickly into the lumen of the distaltubules and the tubules beyond. An injection of urea (0.25 g./100 g.) given duringthe elimination of acriflavin produced no marked changes other than activation of theglomeruli. The concentration of dye in the lumen of the tubules was not increased, and intwo of the three experiments performed the epithelial cells became free of the dye pre-sumably by way of the capillaries or lymph spaces. Occasionally these flushed brightly fora moment. Contrary to the results obtained with fluorescein the concentration of acriflavinin the urine in the bladder was lower than that found after injection of this dye withouturea.

444 P. ELLINGER
In rats with abolished glomerular circulation the injection of ureadid not activate inactive glomeruli, but stimulated excretion of acidurine through the proximal tubules, containing fluorescein but no acri-flavin, previously injected.
For instance, when urea (0.25 g./100 g.) was injected after the injection of fluoresceinthe glomeruli remained inactive, but the circulation in the capillaries of the tubules im-proved. No dye was seen in the capsules, but the lumen of the proximal tubules rapidlybecame bright yellow (pH 5*0) and the dye passed into the distal tubules. This pictureremained unchanged for several hours until the death of the animal. The bladder alwayscontained small amounts of stained urine. Even injections of 0-025 g. of urea per 100 g.caused an excretion of urine through the proximal tubules. After acriflavin (4 mg./100 g.)the injection of urea (0.25 g./100 g.) caused no change of the staining and no acriflavin wasseen in either tubules or bladder.
Effects of repeated injections of urea or sodium bisulphate. The resultsobtained on rats with normal as well as with abolished glomerular cir-culation are similar to those obtained in frogs.
When, after fluorescein, sodium bisulphate (8-16.5 mg./100 g.) was injected every halfhour or hour, excretion of acid urine through the epithelium of the proximal tubules wasobtained only after the first two or at most the first three injections. The rats may diein coma even after the second injection. The yellowish fluorescence which was observedunder similar conditions with a single injection of urea can still be obtained when urea(0 1 g./100 g.) is repeatedly injected up to the fifth or sixth injection, but not with laterones although even ten injections given at intervals of i or 1 hr. may be survived.
In rats with abolished glomerular circulation excretion through the proximal tubulescan be stimulated by injections of urea (0-025 g./100 g.) repeated every half an hour orevery hour up to the fourth or fifth injection. Then the excretion fails and the rats dieif the injection is continued.
This repeated stimulation of tubular excretion at short intervalsannihilates this function and even urea becomes toxic if tubular excretionis abolished and no other means of elimination are available.
Daily injections of urea in doses up to 0-25 g./100 g. did not produce any toxic effectbut sodium bisulphate given daily in small doses (6-8 mg./100 g.) for 10-20 days or inlarger doses (16.5 mg./100 g.) for 2-3 days produced coma and death. At autopsy localinflammation was present in the intestines. As in frogs the number of green and yellowlyochromes in the kidney was considerably reduced, particularly after bisulphate. Aftera course of daily injections of urea or bisulphate the injected fluorescein appeared withina few minutes in the capsules and passed into the proximal tubules in about the sameconcentration and colour (pH 7 0) without staining the epithelium, and into the distaltubules in somewhat greater concentration but still of the same tint. As in correspondingexperiments in frogs the reabsorption of water, dye and alkali in the proximal tubules andthe staining of the epithelium had ceased, but the reabsorption of water in the loops ofHenle was not affected. A subsequent injection of urea caused no marked change, but smalldoses of bisulphate (8 mg./100 g.) temporarily changed the fluorescence in the capsules andin the lumen throughout the tubules to a yellower tint, showing that glomerular filtrationof acid was also not affected. Larger doses of bisulphate killed the animal.

URINE IN AMPHIBIAN AND MAMMALIAN KIDNEY 445
In two rats in which after a course of bisulphate the cord was cut, urineformation stopped, and after an injection of urea (25 mg./100 g.) fluorescein previouslyinjected was seen neither in the capsules nor in the proximal tubules, though theepithelium showed sometimes a faint green fluorescence. Small amounts of bisulphate(up to 8 mg./100 g.) injected under similar conditions had no effect, larger ones killedthe rat.
These experiments give evidence that daily injections of bisulphatedestroy also the excretory function of the proximal tubules. The occa-sional staining of the epithelial cells with fluorescein of the same tintand concentration as in the blood, caused by diffusion of the dye intothe cells, shows also the loss of the selective permeability of the cellmembranes.
DIsCUSSION
The absence in the present experiments of seasonal differences in thefunction of the frog's kidney illustrates that results obtained in differentcountries or on different species of frogs may vary to a great extent. InEngland, in all seasons, most of the partial functions of the kidney behavedin a manner intermediate between that shown by summer and winterfrogs in Heidelberg, but more similar to that of the former. This was truefor the mode of staining of the tubular epithelium with fluorescein andacriflavin, for the pH of this epithelium and of the preliminary urine inthe lumen of the tubules, and for the finding that only a comparativelysmall proportion of the glomeruli was active and that their activityseldom altered spontaneously. The response of the kidney with abolishedarterial supply to fluorescein was equal to that obtained in similar ex-periments during summer in Heidelberg, while the excretion of acidurine through the proximal tubules after the injection of bisulphatecorresponded to the response obtained during winter in Germany.Seasonal differences or the use of different species of frogs may alsoaccount for the conflicting evidence concerning the site of acidification.Montgomery & Pierce [1937] found that in American frogs, R. pipiens,acidification of the urine occurred only in certain parts of the distaltubules. In the present experiments carried out in this country onHungarian frogs, R. esculenta, there was a definite acidification in theproximal as well as in the distal tubules, the effect being stronger inthe latter. Ellinger & Hirt [1929c, 1930a, 1931], however, working inHeidelberg on German frogs, R. esculenta, found that while in summeracidification was not only stronger but also more pronounced in theproximal than in the distal tubules, in winter the urine of these frogswas on the whole much less acidified.

The observation on the frog's kidney that acriflavin first stained thepericellular lymph spaces of the proximal tubules, whether their glo-meruli were active or not, and later stained the cells, shows that this dyereaches them from the blood or lymph and not from the lumen. A similarphenomenon was observed by Ellinger & Lambrechts [1937] usingrhodamin B. The fact that none of the acriflavin passed from the cellinto the lumen illustrates that the presence of a dye in the epitheliumdoes not necessarily imply that it is being excreted there. The taking upof an injected dye by the tubular epithelium and its excretion areprocesses independent of one another and its absence in the lumen whenit is present in the epithelial cells is no evidence in favour of the theoryof Bowman & Heidenhain. Singer [1933], who repeated the experimentsof Ellinger & Hirt on American R. pipiens with results in general agree-ment with ours, believed that he observed injected acriflavin in thetubular lumen before it appeared in the glomerular capsules. He injectedacriflavin only when the elimination of injected aesculin was at its height.Under such conditions it is not possible to decide whether acriflavinstains the epithelium of the tubules alone or their lumen as well, as isoften observed when acriflavin is injected after fluorescein. Acriflavinunlike fluorescein is found in a lower concentration in the capsules thanin the blood. This might be due to the fact that acriflavin forms com-paratively stable compounds with the blood protein which cannot befiltered in the glomerulus. The elimination of acriflavin in the urine isalso far slower than that of fluorescein, since the former is partly fixedin the tissues.
Fluorescein and acriflavin are dealt with in the rat's kidney just asin the frog. Fluorescein is filtered in the glomeruli and concentrated inthe proximal tubules, where water, some alkali, and a little of the dyeare reabsorbed. The concentration of the dye in the distal tubules is notmuch higher, perhaps because some is absorbed with water in the loopsof Henle which cannot be seen, as well as in the distal tubules themselves,but the urine is more acid there. Acriflavin in low concentration entersthe capsules, stains the nuclei of the capsular epithelium and of the cellsof the blood vessels, and enters the epithelial cells of the proximaltubules from the lymph spaces. This staining of the pericellular lymphspaces indicates that lymph takes some part in the formation of urine[cf. Keller, 1933] especially in the absence of glomerular circulation[cf. also Richards & Barnwell, 1927].
The present experiments give new evidence for an excretory functionof the convoluted tubules. Acid urine was shown to be formed in the
P. EllINGER446

URINE IN AMPHIBIAN AND MAMMALIAN KIDNEY 447
proximal tubules of the normal kidney of frogs and rats when urea orsodium bisulphate was injected. A similar response was obtained in theexperiments with abolished glomerular circulation. The intravitalmicroscopic study completely confirmed Nussbaum's observation andthe objections raised against his interpretation could be refuted. Nourine was formed when the arteries were tied, but on injection of ureaacid urine was formed containing, after these dyes had been injected,fluorescein but not acriflavin. These two dyes therefore correspondin their behaviour to sulphindigotate and carmine, respectively, inNussbaum's experiment. There is no doubt that collateral circulationfrom the renal portal vein or genital, vesical, or ureteric arteries maybe opened up by increased pressure or hypertonic urea injections, andglomerular filtration may so be re-established as in the experiments ofBieter & Hirschfelder [1922, 1924, 1926, 1929], Richards & Walker[1927], Bainbridge, Collins & Menzies [1913, 1914] and Kempton [1937];while the injection of urea into frogs with intact arterial supply tothe kidney quickly activates quiescent glomeruli as shown also byRichards [1922] and Bieter & Hirschfelder [1924]. Nothing of thekind occurred in the present experiments in which the glomerularaction was directly observed. Nor can restored glomerular activityexplain the results of Nussbaum [1878, 1879, 1886], Halsey [1902],Bainbridge & Beddard [1906a, b], Tamura, Miyamura, Fakuda, Hosoya,Kishi & Kihara [1927] or Tamura, Miyamura, Nagasawa, Hosoya,Kishi & Fushita [1927], for in these, as in those now reported and in thoseof Ellinger & Hirt [1930a] on winter frogs, the urine differed from thenormal in acidity and in the lack of certain injected dyes. According toBeddard [1902] the activating effect of urea can only be observed whenligation of the arteries has preceded the injection of urea sufficiently longto allow degeneration of the epithelium to occur. This objection isexcluded in the present experiments in which there was no time availablefor degeneration. The only explanation is that, instead of glomerularfiltration followed by partial reabsorption in the tubules, a differentmechanism of excretion through the proximal tubules is brought intoplay and water and fluorescein can in fact be seen being discharged fromthe proximal tubules. The present results differ from those recentlyobtained by Kempton who observed no formation of urine if all collateralswere tied and urea was injected. It is possible that this discrepancy isagain the result of seasonal or climatic variations, frequently referred to.In fact, Ellinger & Hirt [1930a, 1931] found that after arterial ligationspontaneous excretion of urine rarely occurred in the summer, whereas

P. ELLINGER
it was regularly observed in winter frogs. In experiments of the typerecorded in this paper, the urine containing acid and fluorescein but noacriflavin unquestionably was excreted through the cells of the proximaltubules.
Ghiron's experiment was also confirmed by our method. The lowblood pressure abolished glomerular circulation and formation of urine,as well as elimination of fluorescein or acriflavin, although both were tobe seen in the tubular epithelium. After injection of urea, the eliminationof urine started again, but acriflavin still was not excreted. Fluorescein,however, was excreted through the epithelium of the proximal tubulesand not through the glomeruli, which remained inactive.
The results give some indication of the role of the excretory func-tion of the tubules. The epithelium of the proximal convoluted tubules, ifits excretory function is repeatedly called into play, loses its capacity forreabsorption, for excretion, and for being stained by the dyes used. Iffrogs are repeatedly treated with urea or bisulphate and then injectedwith fluorescein and urea after ligation of the arteries, the urine formedin the tubules contains fluorescein in a similar concentration and of thesame pH as in the plasma. Hence it is evident that the power of selectiveexcretion has also been lost. Further evidence of the damage sufferedis given by the increased tendency of the epithelium to take up calcium.Excretion through these cells cannot, therefore, be a constant function.It may come into action in emergency or in pathological conditions whenthe glomeruli fail or the body is loaded with substances which must beremoved quickly. This does not exclude the possibility that under normalconditions some urine constituents may be partly eliminated through theproximal tubules in a concentration more or less similar to that presentin blood. Such an elimination would not involve great osmotic workwhich apparently damages the cells concerned, and could be responsiblefor the tubular excretion of phenol red postulated by Marshall & Vickers[1923] and Marshall [1931] by an indirect method. The studies by Walker& Hudson [1937a, b] of a possible tubular excretion of urea and phos-phate, which would also represent such a type of elimination, did notlead to conclusive evidence.
The epithelium of the proximal tubules may act in two ways: innormal conditions by absorbing water or certain solutes from theglomerular filtrate and perhaps by elimination of certain constituents incomparatively low concentration, and in abnormal conditions by ex-creting water, solids, and acid from the blood. The way in which it actswill depend on the concentration gradient of substances to be absorbed
448

URINE IN AMPHIBIAN AND MAMMALIAN KIDNEY 449
or excreted, as between the epithelium and lumen on the one hand andthe blood and the lymph on the other. What substances are absorbed orexcreted apparently depends on a selective one-way permeability of thecell walls bordering the lumen.
This conception combines some features of the view of Ludwig andCushny with others of the theory of Bowman and Heidenhain, the normalinterplay of filtration and reabsorption being supplemented in specialcircumstances by an excretory function of the proximal tubules. Thisextension of Ludwig and Cushny's theory may possibly explain somepathological conditions, e.g. the secondary nephrosis following acute orchronic glomerulitis, and the constitution of the urine in acute glomerulo-nephritis.
SUMMARY1. The intravital microscopic study of the response of the normal
frog's kidney to injected fluorescein and acriflavin confirms generally theexperiments by Ellinger-& Hirt and reveals
(a) that seasonal variations of the behaviour of the frog's kidneyobserved in Heidelberg did not occur in this country;
(b) that acriflavin stained the epithelium of the proximal tubulesindependently of the elimination of the dye, entering the cells from theintracellular lymph spaces;
(c) that acriflavin, contrary to fluorescein, is eliminated in theglomerular capsules in a lower concentration than it is present in theblood.
2. In the normal rat's kidney both dyes were eliminated in a mannersimilar to that in the frog.
3. In normal frogs and rats a single injection of a large dose of eitherurea or sodium bisulphate stimulates the excretion of acid urine throughthe epithelium of the proximal tubules.
4. The study here made, by intravital microscopy, of Nussbaum'sand Ghiron's experiment confirms the original observations and in-terpretations of these workers: ligation of the arteries of the frog'skidney or reducing the blood pressure by severing the spinal cord in rats,.arrests the glomerular function and the formation of urine in both cases;while after injection of urea, urine different from the normal urine isexcreted in the proximal tubules.
5. Repeated stimulation of the excretory function of the proximaltubules leads to pathological changes in the cells concerned. This ex-cretory function, therefore, cannot be regarded as normal; rather must
PH. XCVII. 29

450 P. ELLINGER
it be regarded as an emergency function in the absence of glomerularfiltration or when the blood is overloaded with substances requiringquick elimination.
I wish to express my thanks to the Lister Institute, for hospitality, to the Society forthe Protection of Science and Learning, London, for a Research Fellowship, and to theElla Sachs Plotz Foundation, Boston, U.S.A. for a grant for technical assistance. I am verymuch indebted to Prof. J. B. Leathes, Dr W. Feldberg and Dr H. Maclean for their assist-ance in the preparation of the manuscript.
REFERENCES
Bainbridge, F. A. & Beddard, A. P. [1906a]. J. Phy8iol. 34, ixP.Bainbridge, F. A. & Beddard, A. P. [1906b]. Biochem. J. 1, 255.Bainbridge, F. A., Collins, S. H. & Menzies, J. A. [1913]. Proc. Roy. Soc. B, 86, 3,55.Bainbridge, F. A., Collins, S. H. & Menzies, J. A. [1914]. J. Physiol. 48, 233.Beddard, A. P. [1902]. J. Physiol. 28, 20.Bieter, R. N. & Hirschfelder, A. D. [1922]. Proc. Soc. exp. Biol., N. Y., 19, 351, 415.Bieter, R. N. & Hirschfelder, A. D. [1924]. Amer. J. Phy8iol. 68, 327.Bieter, R. N. & Hirschfelder, A. D. [1926]. Proc. Soc. exp. Biol., N.Y., 23, 798.Bieter, R. N. & Hirschfelder, A. D. [1929]. Amer. J. Phy8iol. 91, 178.Ellinger, P. [1934a]. Acta brev. neerl. Physiol. 4, 30.Ellinger, P. [1934b]. J. Phy8iol. 82, lOP.Ellinger, P. [1934c]. J. Physiol. 82, 12P.Ellinger, P. [1935]. Chem. & Ind. 54, 85.Ellinger, P. [1938]. Biochem. J. 32, 376.Ellinger, P. & Hirt, A. [1929a]. Anat. Anz. 67, 225.Ellinger, P. & Hirt, A. [1929b]. Z. ge8. Anat. 1. Z. Anat. EntwGe8ch. 90, 791.Ellinger, P. & Hirt, A. [1929c]. Arch. exp. Path. Pharmak. 145, 193.Ellinger, P. & Hirt, A. [1930a]. Arch. exp. Path. Pharmak. 150, 285.Ellinger, P. & Hirt, A. [1930b]. Handb. Biol. ArbMeth. Abt. v, Teil 22, 1753.Ellinger, P. & Hirt, A. [1931]. Arch. exp. Path. Pharmak. 159, 111.Ellinger, P. & Lambrechts, A. [1937]. J. Physiol. 89, 30P.Ghiron, M. [1912]. Zbl. Physiol. 26, 613.Ghiron, M. [1913]. Pftfug. Arch. ge8. Physiol. 150, 405.Halsey, J. [1902]. Amer. J. Physiol. 6, xvi.Keller, R. [1933]. Der elektrieche Faktor der Nierenarbeit, p. 39. Mahrisch-Ostrau.Kempton, R. T. [1937]. Amer. J. Phy8iol. 119, 175.Marshall, E. K. [1931]. Amer. J. Physiol. 99, 77.Marshall, E. K. & Vickers, J. L. [1923]. Johns Hopk. Ho8p. Bull. 34, 1.Montgomery, H. & Pierce, J. A. [1937]. Amer. J. Phyeiol. 118, 144.Nussbaum, M. [1878]. Pflug. Arch. ges. Physiol. 16, 139.Nussbaum, M. [1879]. Pflug. Arch. gee. Phy8iol. 17, 580.Nussbaum, M. [1886]. Anat. Anz. 1, 67.Richards, A. N. [1922]. Amer. J. med. Sci. 163, 1.Richards, A. N. [1929]. Methods and Results of Direct Investigations of the Function of the
Kidney. Baltimore: Williams and Wilkins Co.Richards, A. N. [1938]. Proc. Roy. Soc. B, 126, 328.

URINE IN AMPHIBIAN AND MAMMALIAN KIDNEY 451
Richards, A. N. & Barnwell, J. B. [1927]. Proc. Roy. Soc. B, 102, 714.Richards, A. N. & Walker, A. M. [1927]. Amer. J. Phy8iol. 79, 419.Singer, E. [1933]. Amer. J. Anat. 53, 469.Smith, H. W. [1937]. Phy8iology of the Kidney. New York: Oxford Univ. Press.Robison, R. & Rosenheim, A. H. [1934]. Biochem. J. 28, 684.Tamura, K., Miyamura, K., Fakuda, F., Hosoya, M., Kishi, K. & Kihara, G. [1927].
Jap. J. med. Sci. Pharmacol. 1, 249.'lamura, K., Miyamura, K., Nagasawa, H., Hosoya, M., Kishi, K. & Fushita, T. [1927].
Jap. J. med. Sci. Pharmacol. 1, 263.Walker, A. M. & Hudson, C. L. [1937a]. Amer. J. Physiol. 118, 153.Walker, A. M. & Hudson, C. L. [1937b]. Amer. J. Phy8iol. 118, 167.
EXPLANATION OF PLATES I AND 11
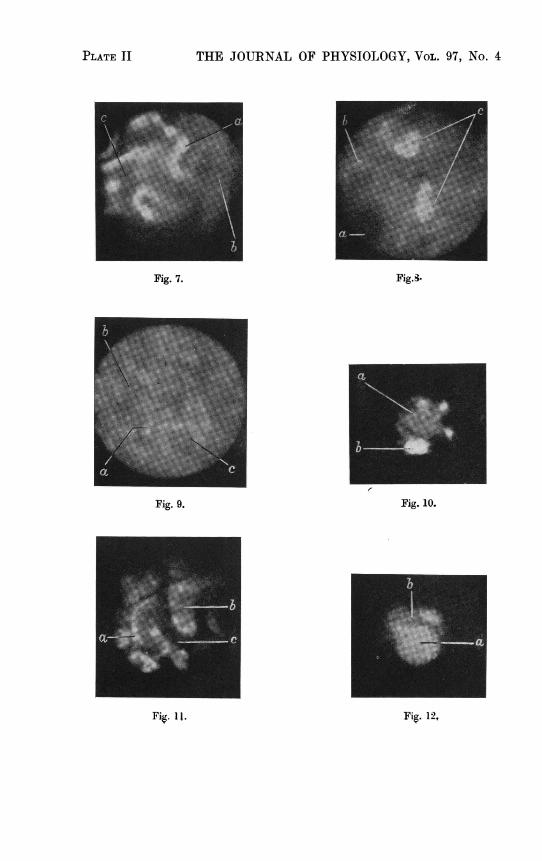
'igs. 1-12. Photographs of living kidneys (magnification: Figs. 1-10 and 12 x 78,Fig. 11 x 150. Zeiss Miflex, reducing the magnification to J. Ilford Panchromatic Plates.Eyepiece: photo x 4 (Zeiss). Objective: Figs. 1-10 and 12, W.I. 39, num. Ap. 1.10;Fig. 11, E.-H. 75, num. Ap. 1.15; both special lenses for intravital microscopy (Zeiss).Filters (Schott): BG3, 2-5 and 3-5 mm., resp. and GG5, 1 mm.). The photographs aretaken directly of the living organs. While these show only one level giving an average stageof the process, in the experiment the micrometer screw is constantly used to observe dif-ferent levels of the object continuously. The different parts of the kidney were identifiedby following up the elimination of fluorescein from the glomerulus downwards and bycomparing the intravital microscopic pictures with those of ordinary microscopic slides ofsections.
PLATE I
Fig. 1. Frog, kidney, ventral side; 2 hr. 51 min. after injection of 0-25 mg. fluorescein:oval active glomerulus (a) with capillaries (al) and bright capsule (a2), some distaltubules (b) with bright narrow lumen (b,) and fairly bright narrow epithelium (b2), onecollecting tubule (c) with bright epithelium (cl) and bright lumen (cq); some of the nucleiof the epithelial cells are stained (cq); the darker intervals between the tubules andsometimes crossing them are capillaries of the renal portal veins (d). Exp. 15 sec.
Fig. 2. Frog, kidney, dorsal side; 21 min. after injection of 0-25 mg. fluorescein: proximaltubules, some with bright epithelium (a,) and bright lumen (a2) (active systems),some tubules whose epithelium is stained with yellowish fluorescent lyochrome par-ticles (bL), the lumen being dark (b2) (inactive systems). Epithelium and lumen arebroader than those of the distal tubules in Fig. 1. The comparatively dark intervalsbetween the tubules are capillaries of the renal portal vein (c). Exp. 1 min.
Fig. 3. Frog, kidney, ventral side, immediately after ligating and cutting the renal arteries:oval inactive glomerulus (a) with capillaries (a1) (far less distinct than those in theactive glomerulus, Fig. 1) and dark capsule (a2), some distal tubules (b) with greenishfluorescent lyochromes in low concentration in the lumen (bL) and dark epithelium (b2),dark intertubular capillaries (c). Exp. 3 min.
Fig. 4. Same frog, same part of the kidney, 10 min. after injection of 0.2 mg. fluorescein:no alteration of the picture. Letters mark the same parts of the picture as in Fig. 3.Exp. 3 min.
29-2

452 P. ELLINGER
Fig. 5. Same frog as in Fig. 3, dorsal side, 13 min. after Fig. 4 (23 min. after injection offluorescein): proximal tubules (a), low concentration of lyochromes in the lumen (a,)and in the epithelium (a2). Exp. 5 min.
Fig. 6. Same frog as in Fig. 3, ventral side, 2 hr. after fluorescein injection and 6 min. afterinjection of 5 ml. 5% urea: oval inactive glomerulus (a) with dark capillaries (a,) anddark capsule (a2) and one proximal tubule (b) with very bright yellow broad epithelium(b,) and bright yellow lumen (b2) and some dark distal tubules (c). Exp. 1 min. 15 sec.
PLATE II
Fig. 7. Same frog as in Fig. 3, ventral side, 2 hr. 30 min. after injection of fluorescein and36 min. after urea injection: distal tubules (a) with bright yellow narrow lumen (a,)and bright narrow epithelium (a2), one distal tubule with fairly bright epithelium (b)and dark lumen (b,), fairly bright intratubular capillaries (c). Exp. 45 sec.
Fig. 8. Frog, injected 12 times with 1 ml. 1-65% sodium bisulphate in 17 days: kidney,ventral side, dark glomerulus (a), some distal tubules (b) with greenish fluorescentlyochromes in the lumen and two proximal tubules (c) with very bright fluorescentcalcified epithelial cells (cl) and darker cell edges (c2). Exp. 3 min.
Fig. 9. Rat, kidney, surface; 25 min. after injection of 3 mg. fluorescein: proximal tubules,some with bright lumen (a1) and bright epithelium (a2) (active systems), some with darklumen (bl) and fairly bright epithelium (b2) (inactive systems). Blood capillaries (c)between the tubules, sometimes crossing them. Exp. 1 min.
Fig. 10. Rat, kidney, after the surface had been shaved off; 3 hr. 20 min. after injection of3 mg. fluorescein: active glomerulus (a) with fairly dark capillaries (a1), bright capsule(a2), vas afferens and efferens (a3) and neck leading to a proximal tube (a4), and adistal tube (b) with very bright lumen (b,) and bright epithelium (b2). Exp. 2 min.
Fig. 11. Rat, kidney, surface; 2 hr. after injection of 3 mg. acriflavin: proximal tubules,fairly bright epithelium (al) with very brightly stained nuclei (a2) and fairly darklumen (b). Between the tubules the dark capillaries (c). Exp. 20 sec.
Fig. 12. Rat, kidney, after the surface had been shaved off; 3 hr. 15 min. after injectionof 3 mg. acriflavin. Active glomerulus (a), the nuclei of the glomerular epitheliumbeing stained (a1), and vas afferens (b) with stained nuclei of the vascular muscle cells(bl). Exp. 1 min. 30 sec.