identification of phosphopeptide ligands for the src-homology 2 (sh2) domain of grb2 by phage...
TRANSCRIPT

Eur. J. Biochem. 246, 633-637 (1997) 0 FEBS 1997
Identification of phosphopeptide ligands for the Src-homology 2 (SH2) domain of Grb2 by phage display Hermann GRAM, Rita SCHMITZ, Jean Franqois ZUBER and Gotz BAUMANN
Novartis Pharma AG, Arthritis and Bone Metabolism, Basel, Switzerland
(Received 29 January/7 April 1997) - EJB 97 0139/1
We report here on the identification of phosphopeptide ligands which interact with the Src-homology 2 (SH2) domain of the adapter protein Grb2 by screening a random peptide library established on phage. Phage were phosphorylated in vitro at an invariant tyrosine residue by a mixture of phosphotyrosine kinases c-Src, Blk and Syk. Selection of binding motifs was carried out by interaction of the library with the recombinant SH2 domain of Grb2 expressed as a glutathione S-transferase (GST) fusion protein. Several subsequent cycles of selection led to the enrichment of phage which bound to the GST-Grb2 SH2 domain only when previously phosphorylated. Sequence analysis revealed that all of the selected phage displayed peptides with the consensus motif Y*M/ENW (Y* denotes phosphotyrosine). One of these peptides, bearing the Y*ENW motif, bound the Grb2 SH2 domain with a threefold higher affinity than the peptide motif Y*VNV derived from the natural ligand Shc. Thus, phage display can be employed to rapidly identify high affinity ligands to SH2 domains.
Keywords: peptide library; phage display; SH2 domain: Grb2.
Many intracellular proteins implicated in signal transduction, such as tyrosine kinases, phosphatases or adapter proteins are associated with src-homology 2 (SH2) andor src-homology 3 (SH3) protein domains which serve to regulate their activity or recruit them into specific signaling complexes. SH2 domains can fold into functional protein modules independently of neighbor- ing sequences and bind to their interacting proteins by recog- nition of linear phosphotyrosine-containing sequence motifs (Overduin et al., 1992; Eck et al., 1993). The binding pocket of an SH2 domain for most phosphotyrosine-containing peptides is bipartite (Sierke et al., 1993): The phosphotyrosine residue typically makes molecular contacts to an invariant arginine resi- due residing in a groove lined with basic residues, while amino acids C-terminal to the phosphotyrosine bind to the less con- served surface of the SH2 domain. This former interaction pro- vides most of the binding energy (Russell et al., 1992) and the latter governs the selectivity of a given SH2 domain in binding to a specific partner. SH2 domains can bind to phosphotyrosine- containing peptides with high affinity (KD = 10- 100 nM) which enables systematic approaches to select from combinato- rial libraries phosphotyrosine peptides which bind to the eliam- ined SH2 domain with high affinity. The two approaches de- scribed sofar rely on synthetic combinatorial peptide libraries containing an invariant phosphotyrosine residue. Those libraries were subjected to interaction with a given SH2 domain and pools of strongly binding peptides were identified by affinity chromatography or fluorescence-activated sorting of beads (Son- gyang et al., 1994; Miiller et al., 1996). Preferentially selected
Correspondence to H. Gram, c/o Novartis Pharma A.G., Arthritis
Fax: +41 61 324 9457. Abbreviations. SH2, Src-homology 2; SH3, Src-homology 3 ; GST,
glutathione S-transferase; MI3 gpIII, phage MI3 gene product 111; Y*, phosphotyrosine residue.
Enzyme. Recombinant phosphotyrosine kinases (EC 2.7.1.1 12).
and Bone Metabolism, Building 3861927, CH-4002 Basel, Switzerland
amino acid motifs were determined by pool sequencing. Though the resolution of these techniques is sufficiently high to identify amino acids crucial for binding to the SH2 domain, the result may be biased, e.g. due to low cycle yield for particular amino acids such as serine, threonine and tryptophan.
The phage-display technique offers an alternative route to generating and screening diverse combinatorial peptide libraries (reviewed by Smith, 1991; Wells and Lowman, 1992), and al- lows for the analysis of single-peptide motifs as opposed to the analysis of peptide pools in the above mentioned techniques.
We have shown previously that peptides displayed on phage can be phosphorylated and selected using an anti-phosphotyro- sine antibody (Schmitz et al., 1996). Here, we demonstrate the feasibility of selecting from a phage library single phosphotyro- sine peptides which bind to the SH2 domain of Grb2, an adapter protein that links phosphotyrosine-mediated signaling to the mi- . -
logen-activated-protein kinase pathway (Lowenstein 1992).
EXPERIMENTAL PROCEDURES
Enzymes and reagents. Human phosphotyrosine
et al.,
kinases Blk, an i Syk expresseld in insect cells by-bac;lovirus were a generous gift from Dr M. Reth and Dr H. Flaswinkel (Max Planck Institute, Freiburg, Germany). The expression of the ca- talytic domain of human c-Src in Escherichia coli and its refold- ing will be reported elsewhere. Expression and purification of the glutathione-S-transferase(GST)-Grb2 SH2 domain was essentially as described (Baumann et al., 1994; Miiller et al., 1996) and phosphotyrosine peptides were synthesized as de- scribed (Miiller et al., 1996). Coupling of recombinant GST and GST fusion proteins to CNBr-activated Sepharose (Pharmacia) was performed according to the instructions. The peptide library used in this study was prepared as reported previously (Schmitz et al., 1996).
634 Gram et al. ( E m J. Biochem. 246)
Phosphorylation and selection. For selection of phage binding to the Grb2 SH2 domain, phosphorylation of the library was carried out using a mixture of the phosphotyrosine kinases c-Src, Blk and Syk (3500 units, 600 units and 160 units, respec- tively). For phosphorylation, 10 p1 phage stock (4.5 X 10" ampi- cillin-resistant transducing unitslml) were incubated for 3 h at ambient temperature with the indicated amounts of phosphotyro- sine kinase in reaction buffer (50 mM Tris/HCl, 5 mM MgCl,, 5 mM MnCl,, 1 mM ATP) in a total volume of 50 p1. Selection of phage on GST-Grb2 SH2 domain fusion protein coupled to CNBr-activated Sepharose beads was done as follows. The reac- tion mixture was transferred to a tube containing 10 pg Sepha- rose-coupled GST-Grb2-SH2 domain, 20 p1 4% non-fat milk- powder in 137mM NaC1, 2.7mM KCI, 4.3mM Na,HPO, . 7 H,O, 1.4 mM KH,P04, pH 7.2 (NaCI/P,) and 0.05% Tween 20. Purified GST was added to a final concentration of 30 pgl ml to the entire mixture in order to prevent non-specific binding to the fusion protein. After incubation for 16 h at ambient tem- perature, the mixture (80 pl) was loaded into a cotton-plugged disposable tip. Elution and amplification of the phage were per- formed exactly as described (Schmitz et al., 1996). For binding studies with single phage clones, 10" ampicillin-resistance transducing units were either treated with a mixture of phospho- tyrosine kinases as described above or left untreated. Affinity selection and determination of ampicillin-resistance transducing units eluted from the columns was performed as described above.
Optical biosensor measurements. The Pharmacia BIAcore 2000 was employed for all studies. For affinity measurement, streptavidin was coupled covalently to the sensor chip CM5 sur- face (corresponding to a signal of about 2000 resonance units) by derivatizing the carboxymethylated dextranhydrogel on the chip surface with N-ethyl-N'-(dimethylaminoproY*l) (Y*, phos- photyrosine) carbodiimide and N-hydroxysuccinimide ; unre- acted groups were quenched by exposure to ethanolamine. Bio- tinylated phosphotyrosine peptides were immobilized on the streptavidin-coated surface and different concentrations of GST- Grb2 SH2 domain fusion protein (range 5-200 nM) were in- fused at a rate of 25 pl/min for 4 min. To minimize re-binding effects during determination of kOff, the corresponding non-bioti- nylated peptide was injected at a concentration of 10 pM. All runs were performed at 25 "C in 20 mM Hepes, pH 7.4, 150 mM NaCl, 0.5 mM dithiothreitol, 0.005% (by vol.) Tween 20 (solu- tion A). The kinetic parameters for binding of the GST-Grb2 SH2 domain were determined using the BIAcore kinetic evalua- tion software version 2.1. Assuming pseudo-first order kinetics, binding of the fusion protein is described by the equation dR/ dt = k,,XcX(R,,,-R,)-k,,XR,., whereas dR/dt is the overall binding rate in resonance units, k,,, describes the association rate constant, k,, the dissociation rate constant, c is the concentration of the fusion protein, R,,, is the maximum amount of protein bound to the chip and R, is the amount of fusion protein bound at time t. For competition experiments, the peptide Biot-cap- DPSY*VNVQ (SDZl17-432) was coated to the chip, and the binding of 50 nM Grb2-SH2-GST monitored during 4 min of association (10 pl/min flow rate) and 3 min in free buffer flow. Competing peptides were added to the SH2 domain sample at concentrations ranging between lo-" M and M. Results were expressed as the concentration giving rise to half-maxi- mum signal obtained after 7 s into the dissociation of a sample without competitor (in order to eliminate contributions of solvent effects to the signal)
RESULTS Preparation of a diverse phage library. To provide a combina- torial and diverse substrate for the SH2 domain of Grb2, a ran-
grb2-I ' grb2-5 ' grb2-9 'grb2-15' pgem3 '
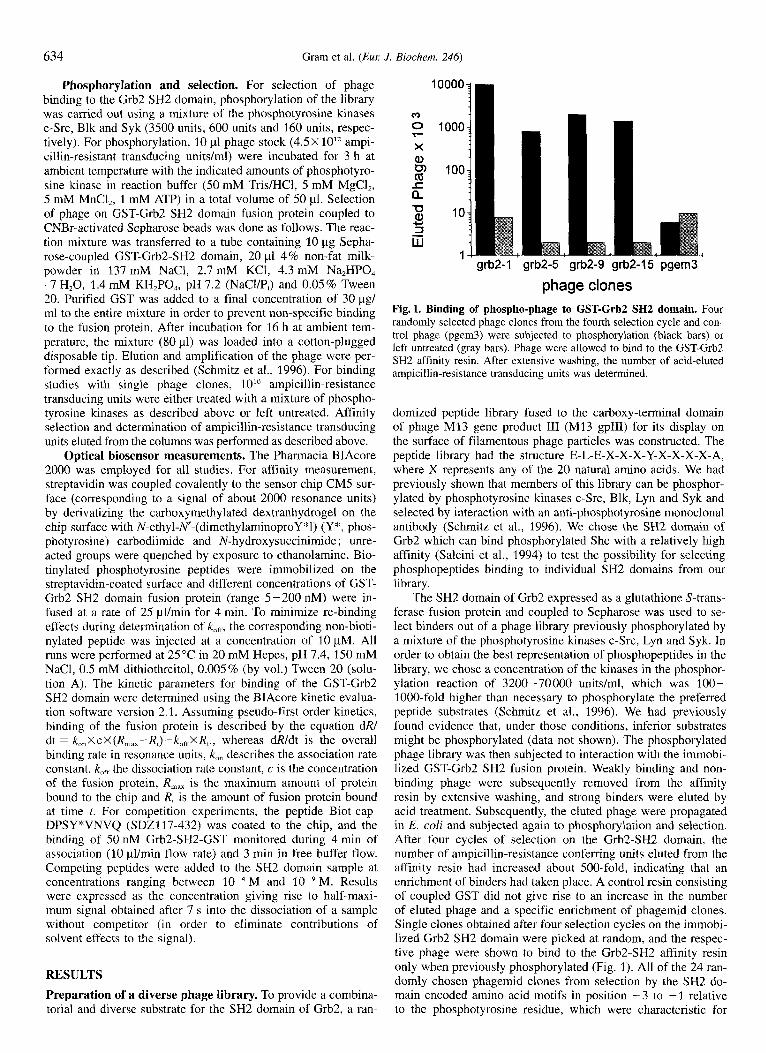
phage clones Fig. 1. Binding of phospho-phage to GST-Grb2 SH2 domain. Four randomly selected phage clones from the fourth selection cycle and con- trol phage (pgem3) were subjected to phosphorylation (black bars) or left untreated (gray bars). Phage were allowed to bind to the GST-Grb2 SH2 affinity resin. After extensive washing, the number of acid-eluted ampicillin-resistance transducing units was determined.
domized peptide library fused to the carboxy-terminal domain of phage MI3 gene product 111 (M13 gpIII) for its display on the surface of filamentous phage particles was constructed. The peptide library had the structure E-L-E-X-X-X-Y-X-X-X-X-A, where X represents any of the 20 natural amino acids. We had previously shown that members of this library can be phosphor- ylated by phosphotyrosine kinases c-Src, Blk, Lyn and Syk and selected by interaction with an anti-phosphotyrosine monoclonal antibody (Schmitz et al., 1996). We chose the SH2 domain of Grb2 which can bind phosphorylated Shc with a relatively high affinity (Salcini et al., 1994) to test the possibility for selecting phosphopeptides binding to individual SH2 domains from our library.
The SH2 domain of Grb2 expressed as a glutathione S-trans- ferase fusion protein and coupled to Sepharose was used to se- lect binders out of a phage library previously phosphorylated by a mixture of the phosphotyrosine kinases c-Src, Lyn and Syk. In order to obtain the best representation of phosphopeptides in the library, we chose a concentration of the kinases in the phosphor- ylation reaction of 3200-70000 units/ml, which was 100- 1000-fold higher than necessary to phosphorylate the preferred peptide substrates (Schmitz et al., 1996). We had previously found evidence that, under those conditions, inferior substrates might be phosphorylated (data not shown). The phosphorylated phage library was then subjected to interaction with the immobi- lized GST-Grb2 SH2 fusion protein. Weakly binding and non- binding phage were subsequently removed from the affinity resin by extensive washing, and strong binders were eluted by acid treatment. Subsequently, the eluted phage were propagated in E. coli and subjected again to phosphorylation and selection. After four cycles of selection on the Grb2-SH2 domain, the number of ampicillin-resistance conferring units eluted from the affinity resin had increased about 500-fold, indicating that an enrichment of binders had taken place. A control resin consisting of coupled GST did not give rise to an increase in the number of eluted phage and a specific enrichment of phagemid clones. Single clones obtained after four selection cycles on the immobi- lized Grb2 SH2 domain were picked at random, and the respec- tive phage were shown to bind to the Grb2-SH2 affinity resin only when previously phosphorylated (Fig. 1). All of the 24 ran- domly chosen phagemid clones from selection by the SH2 do- main encoded amino acid motifs in position -3 to -1 relative to the phosphotyrosine residue, which were characteristic for

Gram et al. (EM J. Biochem. 246) 635
Table 1. Phosphopeptides binding to Grb2 SH2 domain. The ana- lyzed phage clones were randomly chosen after four selection cycles and the encoded peptide sequences were deduced from the respective nucleotide sequences. The amino acids given in positions -4 and +5 were kept invariant in the library. The anticipated phosphotyrosine resi- due is represented by Y*. Amino acids corresponding to the deduced motif Y*E/MNW are represented in bold.
Table 2. Competition experiments. Phosphotyrosine-containing pep- tides were tested for competition of the interaction between Grb2 SH2 domain and the immobilized target peptide DPSY*VNVQ. The experi- ments were performed on a BIAcore instrument. The results represent mean value of two experiments and are expressed as the concentration at which half-maximal competition was observed (ICso). Peptides repre- sented in bold are based on the sequence motif identified in this study.
Peptide Amino acid positions Phosphopep tide IC;,
-4 -3 -2 -1 0 +1 +2 +3 + 4 +5
1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
E E H L Y * M E E P P Y * M E E G V Y * E E E H V Y * L E E P L Y * F E E P M Y * Q E E A T Y * M E Q P L Y * M E A P L Y * W E Q H L Y * M
N F N W N I N W N W N F N W N W N W N W
P A P A P A S A F A S A A A M A Y A M A
E T P V Y * M N F P A E N P V Y * Q N W I A E N P I Y * Q N W I A E V P I Y * E N F P A E S H I Y * E N I V A E S N V Y * E N W T A E D E L Y * Y N W P A E F R V Y * E N F L A E P S V Y * E N Y T A E P I I Y * E N Y V A E P G I Y * W N W F A E S P I Y * E N F P A - E C L Y * Y N L P Y A - E S L Y * Y N W P F A
substrates of c-Src-like kinases (Table 1). In particular, isoleu- cine, valine or leucine in position -1 are strongly preferred by this kinase family (Songyang et al., 1995; Schmitz et al., 1996). Further, the exclusive selection of asparagine in position +2 by the SH2 domain of Grb2 was noted. Asparagine, in contrast, was rarely seen when the library was selected with an anti-phos- photyrosine serum (Schmitz et al., 1996). The frequent occur- rence of glutamic acid and methionine in position + l was also noted. Position +3 is governed by the aromatic amino acids tryptophan, phenylalanine and tyrosine. By comparison of se- quence motifs from the 24 randomly chosen clones emerging after selection of the phage library, the consensus sequence Y* EMNW was derived for binders to the SH2 domain of Grb2. Apart from the invariant asparagine residue in position $2, the methionine/glutamic acid residues and tryptophan frequently found in positions + 1 and +3, respectively, were regarded as potentially important determinants for binding to this SH2 do- main since those amino acids were less frequently observed in these positions when phosphorylated phage were selected by an anti-phosphotyrosine serum.
Competitive binding experiments. Phosphopeptides incorpo- rating the derived motif Y*E/MNW were synthesized and com- pared in their binding activity to previously identified high affin- ity ligands for the SH2 domain of Grb2. The BIAcore biosensor was used to determine the degree of competition of the resynthe- sized peptides for the binding of Grb2-SH2-GST to a reference peptide coated on a sensor chip. The reference peptide was DPSY*VNVQ, the target sequence for the SH2 domain of Grb2 in the natural ligand Shc (Rozakis-Adcock et al., 1992). The concentration of the competing peptides at which half-maximal
PM 1 DPSY*VNVQ 1.9 f 0.6 2 EPFY*ENDPF 1.6 I 0.05 3 EPFY*VNDPD 1.0 t 0.05 4 EPFY*ENVPD 0.8 t 0.05 5 EPFY*QNVPD 0.6 t- 0.05 6 EPFY*VNVDD 0.5 f 0.1 7 EPFY*MNWPD 0.3 t- 0.04 8 EPFY*ENWPD 0.2 -C 0.05 9 EPFY*VtWPD 0.2 t- 0.02
binding of the Grb2 SH2 domain to the immobilized target se- quence occurred was determined by titration (Table 2). While the target sequence itself (peptide 1) had a half-maximal binding constant of 1.9 pM in this assay, peptides 2-6, previously iden- tified from a synthetic library (Muller et al., 1996), competed this interaction more effectively with a half-maximal binding constant of 1.6-0.4 pM. Half-maximal binding constants of 0.2-0.3 pM were determined for peptides 7-9 carrying a tryp- tophan residue in position +3. Peptides 7 and 8 carried methio- nine and glutamic acid in position + 1, respectively, which were identified as predominant amino acids in this position in our phage screen. Peptide 9 with a valine in position + 1 was synthe- sized for comparison, as valine in this position is frequently found in naturally occuring target sequences of the SH2 domain of Grb2 (Muller et al., 1996). Despite the chemical difference between methionine, glutamic acid or valine we did not observe an influence in the competition assay when these amino acids were incorporated in position + 1 of the examined peptides.
Association (Icon) and dissociation (/cot*) rate constants of the Grb2 SH2 domain towards biotinylated peptides 1 and 8 were determined by surface plasmon resonance measurement. The as- sociation rate constants were found to be similar for both pep- tides; 6.0X105 M-' s-' and 4.3X105 M-' s-' for peptides 1 and 8, respectively. In contrast, the dissociation rate constants dif- fered by almost a factor of four; values of 6.0X10-z s-' and 1.6X10-* s-' were determined in this case for peptides 1 and 8, respectively. The resulting binding affinity (KD) for peptide 1 was 100 nM and 37 nM for peptide 8.
DISCUSSION
We employed phage display of a library of random peptides to delineate the substrate specificity of the SH2 domain of Grb2. The presented approach by phage display offers high resolution and sensitivity because it relies on analysis of individual and clonal sequences rather than on the analysis of pools imminent to synthetic library approaches described sofar (Songyang et al., 1994; Muller et al., 1996). A requisite for our method, however, is the availability of a panel of phosphotyrosine kinases which phosphorylate the peptide library ideally in a quantitative way. As became apparent from our recent study on the substrate spe- cificities of various phosphotyrosine kinases, these enzymes re- quire canonical substrate motifs for efficient phosphorylation of a tyrosine residue (Schmitz et al., 1996). As a consequence, the

636 Gram et al. (Eul: J. Biochem. 246)
distinct substrate requirement of individual phosphotyrosine ki- nases only allows for phosphorylation of a fraction of the library and, thus, the phosphorylation step may introduce a considerable bias for later selection. A possible solution may be the use of larger quantities of enzyme, a prolonged incubation time and the employment of a carefully chosen set of phosphotyrosine lc- nases which widely differ in their substrate specificity. Further, knowledge on the substrate specificity of the phosphotyrosine kinases employed is desirable for defining the canonical binding sequence from the phage clones obtained after selection on a given SH2 domain. Though it was difficult to quantitatively as- sess the degree of phosphorylation of our phage library, it was estimated that the amount of enzyme used was sufficient to phosphorylate about 2-3% of the library. This estimate is con- servative and based on indirect evidence obtained from binding of the phosphorylated library to an anti-phosphotyrosine col- umn. We determined in previous experiments that the amount of c-Src used here (3500 units) alone was sufficient to phosphory- late the library to such an extent that 2.5% of the accessible phage could be captured by an anti-phosphotyrosine column (Schmitz et al., 1996). Though only partial phosphorylation of the library has been achieved, probably because two out of the three phosphotyrosine kinases used are from the c-Src family and exhibit a similar substrate requirement, we were able to iso- late phospho-phage binding to the SH2 domain of Grb2 from this library. Previous studies suggested that residues carboxy ter- minal to the phosphotyrosine are implicated in the binding to the Grb2 SH2 domain, and in particular, an asparagine residue in position +2 was found to be critical in this respect (Songyang et al., 1994; Miiller et al., 1996). Similarly, all of the phage analyzed after the fourth selection cycle encoded an asparagine residue in position +2. While phage display and synthetic li- brary approaches predicted a pivotal role for asparagine at posi- tion +2, differences between the two methods were observed for positions +I and +3. For position +1, glutamine, glutamic acid, valine or tyrosine were predicted from synthetic libraries (Songyang et al., 1994; Muller et al., 1996), while we found a preference for glutamic acid and methionine residues. The most apparent difference was observed for position +3, where a strong selection of tryptophan or to a lesser extend, phenylala- nine was described. From approaches using synthetic libraries, no obvious preference for a particular amino acid at this position became apparent. It was reasoned that this discrepancy was due to the fact that tryptophan was either not included in the syn- thetic library (Songyang et al., 1994) or that the cycle yield in sequencing by Edman degradation led to an underestimation of its occurrence. Phenylalanine in this position was the preferred amino acid in the canonical substrate for c-Src, and may, there- fore, reflect a substrate preference by this kinase rather than a true selection by the SH2 domain of Grb2. The canonical sub- strate sequence Y*M/ENW derived in this study was not ob- served when phosphorylated phage libraries were selected by an anti-phosphotyrosine antibody (Schmitz et al., 1996). We con- cluded, therefore, that this sequence motif was specific to selec- tion by the SH2 domain of Grb2 and did not reflect a substrate preference by one of the phosphotyrosine kinases used for phos- phorylation. The more generalized binding sequence Y*-X-N derived for the SH2 domain of the adapter protein Grb2 is in agreement with the sequences D-P-S-Y-V-N-V-Q and D-S-V-Y- A-N-W-M of the Shc protein (Salcini et al., 1994) and the ret protooncogene (Borrello et al., 1994), respectively, both of which were shown to bind Grb2. Interestingly, the latter se- quence motif contains a tryptophan residue in position +3, which meets the predicted consensus sequence in this study.
Most effective competition for binding to the phosphopep- tide ligand DPSY*VNVQ was seen with peptides containing
tryptophan in position +3, suggesting that these peptides would bind with the highest affinity to the SH2 domain. In contrast, choice of either valine, glutamine, glutamic acid or methionine at position fl did not appear to influence the efficiency in the competition experiment (Table 2). Therefore, the frequent occur- rence of glutamic acid and methionine at position + 1 observed in the phagemid clones might not be related to affinity selection of the respective phage but might reflect a yet unexplained bias during phosphorylation or selection. Surface plasmon resonance measurements demonstrated an approximately threefold higher affinity for phosphopeptide 8 containing tryptophan in position + 3 compared to the substrate phosphopeptide derived from Shc. By comparison, peptides 4 and 6 identified from a synthetic li- brary approach were found to bind the SH2 domain of Grb2 with an affinity close to that reported here for peptide 8 (data not shown).
In conclusion, we have demonstrated here that phage display can be used to identify high affinity phosphopeptide SH2 do- main ligands with novel sequence motifs. In more general terms, this work demonstrates that in vitro modification and subsequent selection of a random peptide library displayed on phage is fea- sible.
We thank Drs H. Flaswinkel and M. Reth (Max Planck Institute, Freiburg, Germany) for the generous gift of the phosphotyrosine kinases Blk, Lyn and Syk and Dr F. Gombert for peptide synthesis and character- ization. We further thank Drs U. Manning and J. Eder for critical reading of the manuscript.
REFERENCES Baumann, G., Maier, D., Freuler, F., Tschopp, C., Baudisch, K. & Wie-
nands, J. (1994) In vitro characterization of major ligands for Src homology 2 domains derived from protein tyrosine kinases, from the adaptor protein Shc and from GTPase-activating protein in Ramos B cells, EUK J. lrnmunol. 24, 1799-1807.
Borrello, M. G., Pelicci, G., Arighi, E., De Filippis, L., Greco, A,, Bon- garzone, I., Rizzetti, M., Pelicci, P. G. & Pierotti, M. A. (1994) The oncogenic versions of the Ret and Trk tyrosine kinases bind Shc and Grb2 adaptor proteins, Oncogene 9, 1661 - 1668.
Eck, M. J., Shoelson, S. E. & Harrison, S . C . (1993) Recognition of a high-affinity phosphotyrosyl peptide by the Src homology-2 domain of p561ck, Nature 362, 87-91.
Lowenstein, E. J., Daly, R. J., Batzer, A. G., Li, W., Margolis, B., Lam- mers, R., Ullrich, A,, Skolnik, E. Y., Bar Sagi, D. & Schlessinger, J. (1992) The SH2 and SH3 domain-containing protein Grh2 links re- ceptor tyrosine kinases to ras signaling, Cell 70, 431-442.
Miiller, K., Gombert, F. O., Manning, U., Grossmiiller, F., Graff, P., Zaegel, H., Zuber, J., Freuler, F., Tschopp, C. & Baumann, G. (1996) Rapid identification of phosphopeptide ligands for SH2 domains, J. Biol. Chem. 271, 16500-16505.
Overduin, M., Rios, C. B., Mayer, B. J., Baltimore, D. & Cowhurn, D. (1992) Three-dimensional solution structure of the src homology 2 domain of c-abl, Cell 70, 697-704.
Rozakis-Adcock, M., McGlade, J., Mbamalu, G., Pelicci, G., Daly, R., Li, W., Batzer, A,, Thomas, S . , Brugge, J., Pelicci, P. G., Schles- singer, J. & Pawson, T. (1992) Association of the Shc and Grb2/ Sem5 SH2-containing proteins is implicated in activation of the Ras pathway by tyrosine kinases, Nature 360, 689-692.
Russell, R. B., Breed, J. & Barton, G. J. (1992) Conservation analysis and structure prediction of the SH2 family of phosphotyrosine bind- ing domains, FEBS Lett. 304, 15-20.
Salcini, A. E., McGlade, J., Pelicci, G., Nicoletti, I., Pawson, T. & Pelicci, P. (1994) Formation of She-Grb2 complexes is necessary to induce neoplastic transformation by overexpression of Shc proteins, Oncogene 9, 2827-2836.
Schmitz, R., Baumann, G. & Gram, H. (1996) Catalytic specificity of phosphotyrosine kinases Blk, Lyn, c-Src and Syk as assessed by phage display, J. Mol. Biol. 261, 664-677.

Gram et al. (Eur J. Biochem. 246) 637
Songyang, Z., Carraway, K. L. I., Eck, M. E., Harrison, S. C., Feldman, R. A., Mohammadi, M., Schlessinger, J., Hubbard, S. R., Smith, D. P., Eng, C., Lorenzo, M. J., Ponder, B. A. J., Mayer, B. & Cantley, L, C, (1995) Catalytic specificity ot protein-tyrosine hnases is criti- cal for selective signalling, Nature 373, 536-539.
wells, J, A, & Lowman, H, B. (1992) Rapid evolution of peptide and
Sierke, S. L., Longo, G. M. & Koland, J. G. (1993) Structural basis of interactions between epidermal growth factor receptor and SH2 do- main proteins, Biochem. Biophys. Res. Comnzun. 191, 45-54.
Smith, G. P. (1991) Surface presentation of protein epitopes using bacte- riophage expression systems, Curi: Opin. Biotechnol. 2 , 668-673.
Songyang, Z., Shoelson, s. E., McGlade, J., OliVier, p., PawSon, T., Bus-
Baltimore, D., Ratnofsky, S., Feldman, R. A,, Cantley, L. C. (1994) Specific motifs recognized by the SH2 domains of Csk, 3BP2, fpsl fes, GRB-2, HCP, SHC, Syk, and Vav, Mol. Cell. Biol. 14, 2777- 2785.
telo, X. R., Barbacid, M., Sabe, H., Hanafusa, H., Yi. T., Ren, R., protein binding properties ilz vitro, curx Opin. Biotechnol, 3, 355- 362.