in vivo and in vitro metabolism of 5...
TRANSCRIPT

[CANCER RESEARCH 50. 3207-3211. June I. 1990]
In Vivo and in Vitro Metabolism of 5-Deazaacyclotetrahydrofolate, an Acyclic
Tetrahydrofolate AnalogueMary H. Hanlon,1 Robert Perone, Robert J. Mullin, and Barry R. KeithDepartments of Molecular Genetics and Microbiology and of Medicinal Biochemistry, The H'ellcome Research Laboratories, Research Triangle Park,
North Carolina 27709
ABSTRACT
5-Dca/aacyclotctrahydrofolate is a cytotoxic tetrahydrofolate analogue
»hich inhibits glycinamide ribonucleotidc transformylase (Kclley et al.,J. Med. Chem., 33: 561-567, 1990). Cultured mouse L-cells and humanMCF-7 and MOLT-4 cells concentrated the drug several hundred-foldafter 24 h of continuous exposure to a cytotoxic level (100-200 n.M) of
radiolabelcd drug. High performance liquid Chromatograph) analysisrevealed that each cell type metaboli/ed -Sir , of the internalized drugto polyglutamatcd forms, which are more potent glycinamide ribonucle-otide transformylase inhibitors. In L-cells, 45% of the polyglutamatedmetabolites »ere also /V-formylated. The pharmacokinetics and distribution of [14C|-deazaacyclotetrahydrofolate were studied in C57BL/6 male
mice. Its plasma half-life was 2.15 h. Radiolabel was concentrated to
well above plasma level in the kidney, pancreas, and liver. Metabolismwas examined in tumor-bearing and in normal mice. Twenty-four h after
a single i.p. injection (50 mg/kg), drug equivalents were 0.6 nmol/g (83%polyglutamated) in colon-38 adenocarcinoma carried s.c., 2.4 nmol/g
(100% polyglutamated) in ascitic P388 cells, and 3.7 nmol/g (76%polyglutamated and -20% formylated) in mouse liver. Elimination wasmostly in the urine as unmetabolized drug. Feces contained 5-deazaacy-
clotetrahydropteroate (parent compound less glutamate). In conclusion,5-deazaacyclotetrahydrofolate was shown to be concentrated to well
above the extracellular level and metabolized to more active polyglutamated forms by transformed cells grown in culture and in mice.
INTRODUCTION
Various reports in the literature (1-3) have demonstratedhigher levels of de novo purine biosynthetic enzymes and greaterincorporation of simple precursors into purines in cancer cells,compared to normal cells. Thus, the enzymes of purine de novosynthesis may be selective targets for cancer chemotherapy.Additional selectivity might be obtained by inhibition of a targetenzyme with a prodrug which could be selectively activated intumor cells. A folie acid analogue which inhibits purine de novosynthesis and which could be activated by polyglutamationmight fit these criteria. Several such inhibitors, homofolate (4),reduced homofolate analogues (5, 6), 5,10-dideazatetrahydro-folate(7), and 5-DACTHF- (8), have been described.
5-DACTHF is a cytotoxic tetrahydrofolic acid analogue (Fig.1) which inhibits the two folate-requiring enzymes of purine denovo synthesis, GAR and 5-amino-4-imidazole carboxamideribonucleotide transformylases (8). Previous work (8) demonstrated that 5-DACTHF is a good substrate for mammalianfolylpolyglutamate synthetase, that polyglutamation makes it amore potent inhibitor of both transformylases, and that itscytotoxicity can be reversed by hypoxanthine alone and byleucovorin alone, which suggests that it interrupts purine bio-
Reccivcd 12/5/89: revised 2/13/90.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1To »hornrequests for reprints should be addressed.2The abbreviations used are: 5-DACTHF, A'-|4-|3-(2.4-diamino-1.6-dihydro-
6-oxo-5-pyrimidinyl)propylamino|-benzo>l|-i.-glutamic acid; GAR. glycinamideribonucleotide: N-CHO-5-DACTHF. A'-|4-[3-(2.4-diamino-1.6-dihydro-6-oxo-5-pyrimidinyl)-A'-forni\lpropylamino]-benzoyll-L-glutaniic aeid: FPGS. folylpolyglutamate symhetase: TFA. trifluoroacetic acid: PBS. phosphate-buffered saline:HPLC. high performance liquid chromatography; SAX. strong aniónexchange.
synthesis in whole cells at a site which requires a reduced folate.In human MOLT-4 T-cell acute lymphatic leukemia cells, 5-DACTHF inhibits the incorporation of [l''C]formate into for-
myl glycinamide ribonucleotide (8); it also shows antitumoractivity against mouse P388 leukemia cells grown in mice (8).To further define its mode of action, we have examined themetabolism of 5-DACTHF in tumor cells and in mice and itsbioavailability and distribution in mice, using radiolabeled drug.Of particular interest is whether the drug is activated in vivo bymetabolism to polyglutamated forms.
MATERIALS AND METHODS
Preparation of 5-DACTHF and Analogues
5-DACTHF and N-CHO-5-DACTHF were made as described (8).[14C]-5-DACTHF was synthesized by John Hill of these laboratories bycondensation of dimethyl [benzoyl-carbonyl-uC]-Ar-(4-aminobenzoyl)-L-glutamate (purchased from International Chemical Corporation, UK)with 3-(2-acetylamino-4-diacetylamino-6-hydroxy-5-pyrimidinyl)pro-pionaldehyde, as described (9). Before each experiment, radioactivecontaminants were removed from the drug solution by extraction on a500-mg Cls cartridge, as described below. The eluent (in 5% ammoniumacetate) was lyophilized and redissolved in PBS, pH 7, or cell culturemedium. Both radiochemical and UV HPLC analysis showed that all5-DACTHF solutions used were >98% pure. Escherichia coli folylpolyglutamate synthetase (10) was used to synthesize the analogues of 5-DACTHF with one and two additional glutamates. These productswere extracted on a 100-mg C,8 cartridge, as described below, andseparated by C,«HPLC. Fractions containing the reaction productswere collected, brought to 100 mM 2-mercaptoethanol, lyophilized todryness, and redissolved in water. These and all solutions containinganalogues and metabolites of |'4C]-5-DACTHF were protected from airand light by saturation with oxygen-free argon and storage in ambergas-tight vials. 5-Deazaacyclotetrahydropteroate (5-DACTHF less theglutamate) and the hcxaglutamated analogue were chemically synthesized.' Partial enzymatic hydrolysis of the latter compound with hogkidney 7-glutamyl hydrolase resulted in a mixture of analogues of 5-DACTHF with three to six glutamates. One g of dried hog kidney(Difco) was extracted (II), precipitated with 55% ammonium sulfate,and centrifuged. The pellet was dissolved in 4 ml of 0.04 M EDTA, IHIML-cysteine, and dialyzed against the same solution. Dialyzed hogkidney extract (10 /jl) was added to 120 ^1 of 100 /IM hexaglutamatedsubstrate in 40 ITIMsodium acetate, pH 4.7, 2 HIML-cysteine. After 2h at 37"C, the hydrolysis was terminated by acidification and extraction
on a 100-mg d»cartridge, as described below.
Cell Cultures
MOLT-4 cells, human MCF-7 breast adenocarcinoma cells, andmouse L-cells (a connective tissue carcinoma) were obtained from theAmerican Type Culture Collection. All cells were maintained at 37°C
in 95% air/5% CO2, in folate-free RPMI 1640 medium (GIBCO)supplemented with 10 nM (6-R,S)-Ca leucovorin and 10% fetal calfserum (which had been dialyzed against 0.3% charcoal in 0.9% NaCl).For metabolism studies, MOLT-4 cells were cultured in 25-cnr flasks
3V. L. Styles and J. L. Kelley. Synthesis of the penta-glutamyl derivative ofyV-U4-t/V-[3-(2,4-diamino-l .6-dihydro-6-oxo-5-p>TÃŒmidnyl)propyl]amino|-benzyol||-i.-glutamic acid (5-DACTHF). An acyclic analogue of tetrahydrofolicacid. J. Heterocyclic Chem.. in press. 1990.
3207
on July 2, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

METABOLISM OF 5-DACTHF
H2N
Tetrahydrofolate
If H ?°°H
C—N—CH
CH2
CH2
COOH
HC-N-CH
COOH
Fig.
CH2
CH2
COOH
5-DACTHF (5-deaza-acyclo-tetrahydrofolate); R=H
N-CHO-5-DACTHF; R=CHOStructures of 5-DACTHF and analogues. *. radiolabcled carbon.
at a seed density of 1 x IO6cells/ml, in the presence of 100 nM |'4C]-5-DACTHF ( 110,000 dpm/nmol). L-cells and MCF-7 cells were grownto 50% confluency in 75-cm2 flasks before the addition of drug to the
medium. After continuous drug exposure, the attached cells werewashed twice on the culture flask with ice-cold PBS and harvested byscraping and centrifugation; MOLT-4 cells were harvested and washedby centrifugation and were washed twice by suspension in ice-cold PBS
followed by centrifugation.
In Vivo Procedures
Mouse colon 38 adenocarcinoma was obtained from Southern Research Institute (Birmingham, AL) and was maintained as s.c. axillaryimplants in C57BL/6 mice (Charles River), with a passage time of 21-28 days. P388 cells were obtained from Trudeau Institute (SeranacLake, NY). For metabolism studies, 1x10* P388 cells were implantedi.p. into each of nine 20-g CDF, male mice (Charles River). Sixteen hlater, drug was administered (¡.p.).Ascitic cells were harvested with twoconsecutive 5-ml i.p. rinses of 4°CPBS. The cell suspension wasbrought to 40 ml with 4°CPBS and was centrifuged for 7 min at 365x g at 4"C. The pellet was suspended for 20 s in 1 ml of ice-coldhypotonie PBS (0.2%) to lyse RBC. A 20-ml aliquot of 4°CPBS was
immediately added and the suspension was centrifuged as above. Thecells were resuspended in 10 ml 4°CPBS, counted in a hemocytometer,
and repelleted.All mouse experiments, except those with P388 cells, were done with
20-22-g C57BL/6 male mice (Charles River). Unless noted otherwise,"C-labeled drug (4,000-12,000 dpm/nmol), in PBS, was administered
i.p. at 50 mg/kg. After sacrifice by cervical dislocation, blood was drawnfrom the vena cava, nitrogen saturated to minimize oxidation of thedrug and its metabolites, and placed on ice. Plasma was separated bycentrifugation at 4°Cfor 15 min at 5,000 x g. Tissues were excised,
rinsed with ice-cold sterile 0.15 M NaCl, weighed, and frozen in specimen containers which had been precooled on a block of dry ice. Tissueswere stored at -70°Cfor up to 2 months before extraction and metab
olite analysis. Feces and urine were collected in a metabolism cage (fourmice/cage). Feces were collected at 24 and 48 h and frozen. Urine andcage washes were immediately frozen. At 3, 6, 24, and 48 h after drugadministration, urine was thawed, argon saturated, sterilized by filtration (0.22-^m Millex GV), and frozen (-70T).
Pharmacokinetics
The pharmacokinetics of 5-DACTHF in mouse plasma were evaluated from concentration data obtained from 63 mice (3 mice/timepoint) after administration of a single 50 mg/kg dose. The drug was
administered either i.v. (through the tail vein) or i.p. The total 14C
content in the plasma was determined by combustion, as describedbelow. That fraction of the total plasma IJC which was parent drug was
determined by HPLC analysis. Phramacokinetic parameters were estimated with a nonlinear least-squares regression computer program,NONLIN (12). Mean plasma drug concentrations were fit to a two-compartment open model with first-order elimination. The bioavail-ability of 5-DACTHF after i.p. administration was determined fromthe ratio of the areas under the concentration-time curves.
Detection of Radioactivity
Total radioactivity was determined in all tissues described in thisstudy. Whole tissues, aliquots of plasma and crude tissue homogenates,and all cell pellets and precipitates were combusted in a model 306Packard oxidizer. The generated 14Cwas trapped in a column filledwith Packard Carbo-sorb. flushed into a vial using the scintillantPermafluor V (Packard), and counted in a Beckman model LS230liquid scintillation counter. Aliquots of all cell lysates and tissue homogenates were counted in the Beckman liquid scintillation counter in10 ml of Beckman Ready-Safe. Recovery of radioactivity through allstages of sample preparation for HPLC was >85%.
Extraction and Preparation of Samples for HPLC
Urine. Urine was diluted with an equal volume of aqueous 2% (w/v)TFA. Up to 100 fi\ of the acidified urine were applied to a 100-mg C18Bond Elut cartridge (Analytichem International) which had been pre-washed sequentially with 1 ml acetonitrile, 1 ml water, and 1 ml 0.1%TFA. After sample application, the column was washed with 8 ml 0.1 %TFA. The l4C-metabolites were eluted in 1 ml of either 0.1% TFA in
50% acetonitrile (for SAX HPLC) or 5% ammonium acetate in ethanol(for C,8 HPLC). The TFA-acetonitrile eluents were filtered (MilliporeHV) and examined directly on SAX HPLC. The ammonium acetate-ethanol eluents were dried under an argon stream at 45°C,dissolved in
5% ammonium acetate in water, and filtered. Up to 3 ml of this filtratewere loaded onto the Cm HPLC column.
Plasma. An aliquot of plasma (5-300 pi) was diluted with water to900 n\, made pH <2 by the addition of 100 n\ 10% TFA, and extractedon a C|g cartridge, described as above.
Cell Pellets. MCF-7, MOLT-4, and P388 cells were lysed (IO7 cells/
ml) in boiling extraction buffer [50 niM Na,K phosphate, pH 7; 100HIMNa citrate (to inhibit cellular -y-glutamyl hydrolases); 100 mM ß-
mercaptoethanol (to protect the drug and its metabolites from oxidation)] and boiled for 10 min. Lysates were clarified by centrifugation(30 min. 4°C.17,000 g). The supernatants were adjusted to pH <2
with 0.15 volumes of 40% TFA, centrifuged, and extracted on 100-mgC,«cartridges, as described above.
Colon-38, Livers, and Feces. Samples were removed from the —70°C
freezer, held at room temperature for 2 min, shattered with a hammer,added to a tube containing 6 ml/g sample of ice-cold extraction buffer,and homogenized with a Brinkman Polytron (20 sec at setting 11). Thehomogenate was held in boiling water for 10 min, cooled on ice, andcentrifuged (30 min, 4°C,17,000 x g). The pellet was re-extracted as
above. The two supernatants were pooled, adjusted to pH <2 with 0.15volumes of 40% TFA, and centrifuged (15 min, 4°C,3,000 x g). Tumor
supernatants were extracted on 100-mg C18 cartridges, as describedabove. Up to 10 ml of liver supernatant were extracted on a 500-mgCu cartridge, which was developed as above except that the volume ofall washes and elutions was increased 5-fold. A 1-ml aliquot of fecalsupernatant was extracted on a 500-mg C»cartridge, which was developed like the 500-mg C,8 cartridge. This cartridge was used for fecalsamples because much of the radioactivity in these samples could notbe removed from a C,Hcartridge.
High Pressure Liquid Chromatograph) and Mass Spectral Analyses
Two HPLC columns were used to separate metabolites of |'4C]-5-DACTHF. A Whatman SAX column (25 cm), developed with a 1222-s linear gradient from water to 650 m\i KH:PO4. pH 6. at 1.5 ml/min.gave superior baseline separation of the drug and its polyglutamatcdanalogues. UV absorbance was monitored with Waters 480, Perkin
3208
on July 2, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
METABOLISM OF 5-DACTHF
Elmer LC95, and LKB 2140 detectors. The latter provided spectra ofmetabolites. The column eluent was mixed with Fisher Scintiverse LCat 4.5 ml/min and 14Cwas detected by a radioactivity flow detector(Flo-one; Radiomatic, Inc.). A Waters C18 Novapak Radial Compression Column, developed with a 1222-s linear gradient from 0 to 13%acetonitrile in 140 mM ammonium acetate, pH 5.5, at 1.5 ml/min, gaveoptimum separation of several /V-substituted drug analogues and mon-oglutamated metabolites. This eluent was mixed with Flo-Scint III(Radiomatic) at 4.5 ml/min for detection of radioactivity. The outputfrom each of the detectors was captured by a Digital Specialitiesminicomputer and analyzed as described previously (13).
Liquid chromatography-mass spectral analysis was done with aWaters Ci8 n-Bondapak column, developed with a 600-s linear gradient
from 6 to 24% acetonitrile in 140 mM ammonium acetate, pH 5.5,coupled to a Finnigan MAT 4500 mass spectrometer with a VestecCorporation thermospray interface.
Rat Plasma 7-Glutamyl Hydrolase and GAR Transformylase Reactions
Metabolism of [14C]-5-DACTHF other than polyglutamation was
examined following enzymatic hydrolysis of added glutamates. Samplealiquots (200 ^1 in 5% ammonium acetate, 30 mM 2-mercaptoethanol)were incubated for 2 h at 37°Cwith 50 n\ dialyzed rat plasma, whichcontains -y-glutamyl hydrolase activity (14). Following this hydrolysis,the samples were HV filtered and fractionated by C,8 HPLC. Thosefractions which contained I4C were collected and further characterized.Those which co-eluted with synthetic /V-CHO-5-DACTHF were lyoph-ilized, subjected to an enzymatic treatment (see below) which hadpreviously been shown to deformylate ¿V-CHO-5-DACTHF, and re-examined on HPLC.
/V-CHO-5-DACTHF and samples suspected of containing A'-CHO-5-DACTHF were dissolved in 400 n\ 100 mM 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid, pH 8.5. Twenty-five ¿tl5 mM GAR and6 units hog liver GAR transformylase in 10 M'(8) were added, and themixture was incubated at 37°C.The enzymatic deformylation was
followed by examination by HPLC of aliquots taken at intervals up to20 h. Identifications of substrate and product peaks were based onretention times and UV spectra.
RESULTS
In Vitro Metabolism. Each cell line was exposed to a cyto-toxic level (100 HM for MCF-7 and 200 DM for MOLT-4 andL-cells) of [14C]-5-DACTHF for up to one cell doubling time.
At the times indicated in Table 1, the cells were extracted andthe 14C-metabolites were identified. After one cell doubling,intracellular concentrations of radioactivity were 21-26 pmol/IO6cells in each cell line. This represents a concentration 108-228-fold greater than extracellular levels (if the intracellularvolume is estimated to be 1 /il/106 cells). This intracellular
accumulation of drug was accompanied by rapid metabolism.As early as l h after drug exposure, polyglutamated metabolites
Table 1 In vitro polyglutamation of 5-DACTHF
Intracellular concentration(pmol/106 cells)
SourceL-cells
MCF-7Extra
cellularconcentration(nM)200
100Expo
sure(h)3
1914
245-DACTHF0
02.8
1.62.05-
DACTHF+
1 or 2glutamates0
00.4
0.805-
DACTHF+
3-7glutamates6.4"
21.1*0.8
3.620.8Total,4C6.4
21.14.06.422.8
MOLT-4 200 24 0.4 3.2 21.6 25.6"81% were formylated as well as polyglutamated.6 45% were formylated as well as polyglutamated.
500 1000SECONDS
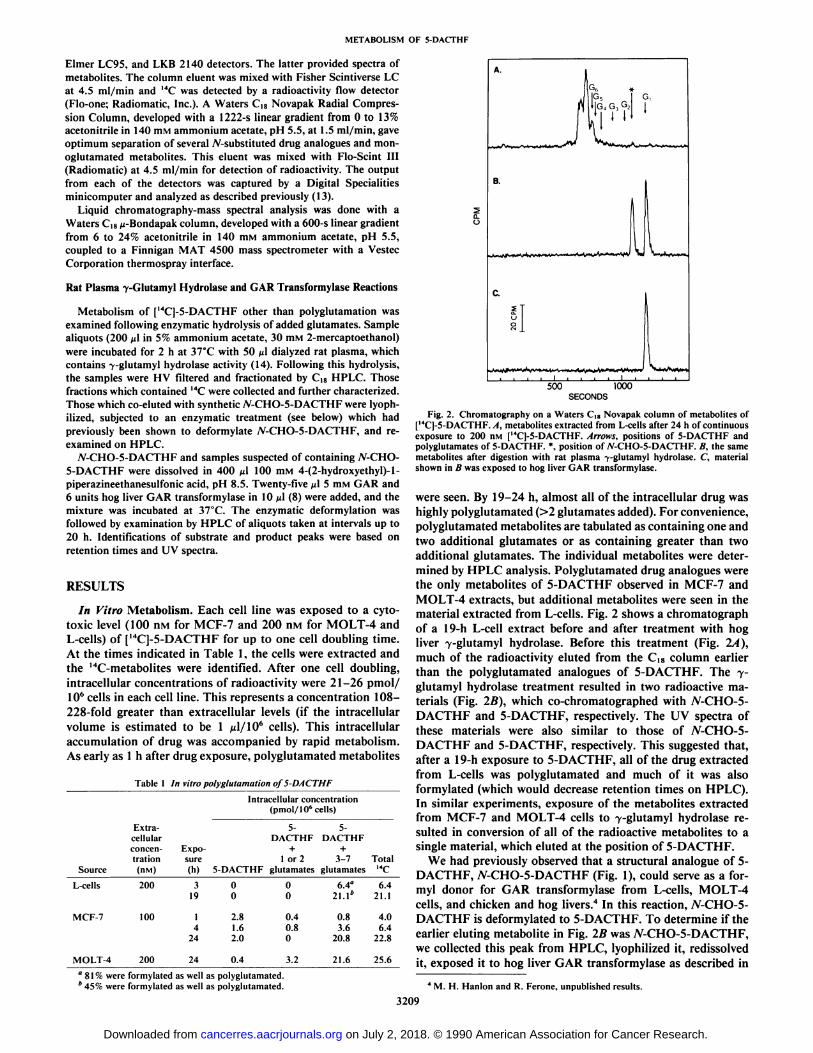
Fig. 2. Chromatography on a Waters CÃaNovapak column of metabolites of[14C]-5-DACTHF. A, metabolites extracted from L-cells after 24 h of continuousexposure to 200 nM |MC]-5-DACTHF. Arrows, positions of 5-DACTHF andpolyglutamates of 5-DACTHF. *. position of /V-CHO-5-DACTHF. B, the samemetabolites after digestion with rat plasma -y-glutamyl hydrolase. C, materialshown in B was exposed to hog liver GAR transformylase.
were seen. By 19-24 h, almost all of the intracellular drug washighly polyglutamated (>2 glutamates added). For convenience,polyglutamated metabolites are tabulated as containing one andtwo additional glutamates or as containing greater than twoadditional glutamates. The individual metabolites were determined by HPLC analysis. Polyglutamated drug analogues werethe only metabolites of 5-DACTHF observed in MCF-7 andMOLT-4 extracts, but additional metabolites were seen in thematerial extracted from L-cells. Fig. 2 shows a Chromatographof a 19-h L-cell extract before and after treatment with hogliver 7-glutamyl hydrolase. Before this treatment (Fig. 2/1),much of the radioactivity eluted from the C|8 column earlierthan the polyglutamated analogues of 5-DACTHF. The 7-glutamyl hydrolase treatment resulted in two radioactive materials (Fig. 2B), which co-chromatographed with 7V-CHO-5-DACTHF and 5-DACTHF, respectively. The UV spectra ofthese materials were also similar to those of W-CHO-5-DACTHF and 5-DACTHF, respectively. This suggested that,after a 19-h exposure to 5-DACTHF, all of the drug extractedfrom L-cells was polyglutamated and much of it was alsoformylated (which would decrease retention times on HPLC).In similar experiments, exposure of the metabolites extractedfrom MCF-7 and MOLT-4 cells to 7-glutamyl hydrolase resulted in conversion of all of the radioactive metabolites to asingle material, which eluted at the position of 5-DACTHF.
We had previously observed that a structural analogue of 5-DACTHF, N-CHO-5-DACTHF (Fig. 1), could serve as a for-myl donor for GAR transformylase from L-cells, MOLT-4cells, and chicken and hog livers.4 In this reaction, yV-CHO-5-DACTHF is deformylated to 5-DACTHF. To determine if theearlier eluting metabolite in Fig. IB was 7V-CHO-5-DACTHF,we collected this peak from HPLC, lyophilized it, redissolvedit, exposed it to hog liver GAR transformylase as described in
4 M. H. Hanlon and R. Perone, unpublished results.
3209
on July 2, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

METABOLISM OF 5-DACTHF
"Materials and Methods," and rechromatographed it. Fig. 2C
shows that after this treatment all of the radioactivity cochro-matographed with 5-DACTHF. Thus, we concluded that theearlier eluting metabolite observed in Fig. 2B was yV-CHO-5-DACTHF.
Pharmacokinetics and Distribution. Plasma concentrations of['4C]-5-DACTHF exhibited biexponential decay after both i.v.and i.p. administration. The mean concentration-time data werevery similar for both routes of administration and are displayedin Fig. 3, with the model-fit profiles. After i.v. administration,the estimated distribution and terminal elimination half-livesof 5-DACTHF were approximately 0.2 and 1.8 h, respectively.The model-predicted clearance was approximately 1300 ml/min/kg and the volume of distribution of the central compartment was about 600 ml/kg. The estimated steady state volumeof distribution after i.v. administration was approximately 1500ml/kg, indicating appreciable distribution of the drug intotissues. Model-predicted areas under the concentration-timecurves were 65 and 72 h-nmol/ml after i.p. and i.v. administration, respectively, resulting in an estimated i.p. bioavailabilityin the mouse of 90%. In a preliminary experiment (data notshown), bioavailability was estimated to be between 13 and 32%following p.o. drug administration.
Table 2 shows the distribution of I4C in mouse organs at 1.5
and 24 h following i.p. drug administration. Radioactivity accumulated over the plasma level in most of the tissues exam-
u- 1000.0 -q
02468
HOURSFig. 3. Plasma levels of 5-DACTHF. C57BL/6 mice received 50 mg ['"CJ-5-
DACTHF/kg either i.v. (A A) or i.p. (D—G). Values shown are averages ofthree determinations.
Table 2 Concentration of'C in mouse tissuesEach C57BL/6 mouse received 50 mg ['4C]5-DACTHF/kg. i.p., at t = 0
14Cconcentration
(nmol/g)
1.5h° 24 h"
KidneyPancreasLiverSpleenHeartLungTesticlesBrain54.858.41139.096.284.606.545.0919f1.635.920.881.050.910.490.48
0.082.972.191.570.760.600.522.861.521.650.300.100.060.38±0.180.38
+ 0.21"At 1.5 h after drug administration, plasma contained 24.94 ±15.40 nmol 5-
DACTHF/ml.* At 24 h after drug administration, plasma contained 0.20 ±0.01 nmol 5-
DACTHF/ml.c Mean ±SE; n = 3.
¡ned,particularly in the kidney, pancreas, and liver. In the first24 h after dosing, 74% of the administered I4C was excreted in
the urine and 2% was recovered in the feces; recoveries were 1and 6%, respectively, in the second 24 h.
In Vivo Metabolism. HPLC analysis of mouse urine revealedtwo radioactive components. Eighty-eight % of the radioactivitywas parent drug through the first 24 h and 34% was parentdrug in the second 24 h. The remainder was a metabolite whicheluted from the SAX column prior to 5-DACTHF and whichwas not further characterized. Two radioactive species wereextracted from the feces. Less than half of the 14Cin each fecal
sample was parent drug. The remainder of the radioactivity wasa metabolite which was identified as 5-deazaacyclotetrahydro-pteroate. Its chromatography on C18 HPLC and its mass,determined by liquid chromatographic-mass spectral analysis(data not shown), were identical to those of the pteroate analogue.
Table 3 shows the concentration of l4C-metabolites in mouse
liver and in tumors at the indicated times after drug administration. The liver data are from mice implanted with colon-38;livers from non-tumor-bearing mice gave similar results. Poly-glutamated analogues of [l4C]-5-DACTHF accumulated in the
liver over the 24 h studied. They were first observed between 2and 5 h in the livers of non-tumor-bearing mice. At 24 h afterdrug administration, 62% of the intracellular drug was highlypolyglutamated and 10-20% of it was also formylated. Totaldrug equivalents (parent drug plus polyglutamated metabolites)in the liver were 3.7 nmol/g at this time. The concentration ofdrug equivalents at 24 h was 6-fold lower in colon-38 takenfrom the same animals and only 33% was highly polyglutamated. In contrast, P-388 cells grown in mice rapidly concentrated and polyglutamated the drug, such that at 24 h intracellular drug equivalents were 2.4 nmol/g and 100% was highlypolyglutamated.
DISCUSSION
We have observed the metabolism of 5-DACTHF in tissueculture, in mice, and in tumors implanted into mice. Each ofthe in vitro cell lines studied here is highly sensitive to the drug.Growth IC50 values for 72-h continuous exposure are 37, 58,
Table 3 In vivopolyglutamation of 5-DACTHFEach mouse received 50 mg |'"C]-5-DACTHF/kg, i.p. at t = 0.
Intracellular concentration(nmol/g)
Source (h)5-DACTHF+1or25-DACTHF
glutamates5-DACTHF+3-6glutamatesTotal,«c
Mouse liver0.17 36.9"
1.5 83.37.0 16.8 ±7.5*
24.0 0.8 ±0.4
Colon-380.171.5724
4.75.4
2.9 ±1.1O.I ±0.1
00
2.4 ±0.6C0.5±0.1C
00
0.4 ±0.20.3 ±0.1
00
1.2±0.4C2.3 ±0.2f
00o
0.2 ±0.1
36.983.3
21.8 + 8.83.7 ±0.3
4.75.4
3.5 ±1.40.6 ±0.1
P38847271.312.800.1000.62.62.42.015.42.4
* Mean ±SE; n = 4.' 10-20% of the polyglutamated mouse liver metabolites were also formy lated.
3210
on July 2, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

METABOLISM OF 5-DACTHF
and 80 nivi, respectively, for MCF-7, MOLT-4, and L-cells (8).These cells rapidly concentrated and polyglutamated the drug.Since polyglutamates of this drug are better inhibitors of GARand 5-amino-4-imidazole carboxamide ribonucleotide transfor-mylases than is the monoglutamated parent drug (8), 5-DACTHF was activated in situ. We have also observed thatmouse L-cells and mouse liver formylate 5-DACTHF; however,the contribution of the formylated metabolites to the drug's
cytotoxicity is unclear, since 5-DACTHF is toxic to other cellswhich do not formylate it.
The in vivo studies showed that, after a single 50-mg/kg doseof 5-DACTHF, plasma levels of the drug which were toxic incell culture (>100 HM) could be maintained in mice for 24 h.The drug was concentrated to well above the plasma level inkidney, pancreas, and liver, as well as in P388 tumor cells.Elimination was principally as unmetabolized drug, in the urine.The feces contained deglutamated drug. The source of thismetabolism was not examined, but it was likely a product ofbacterial metabolism, as has been seen with other folate analogues (15, 16).
Polyglutamates of 5-DACTHF were observed in each of thetissues examined for metabolites. Since intracellular retentionof antifolates against a concentration gradient is thought to bea function of polyglutamation (17), the active polyglutamatedmetabolites are expected to be retained in cells longer than theparent drug. This phenomenon was clearly demonstrated in thein vivo data shown in Table 3. In each of the tissues examined,the amount of total drug equivalents (parent plus all polyglutamated metabolites) present at 24 h after drug administrationwas almost identical to the amount of polyglutamated drugpresent at 7 h. Thus, drug polyglutamation was clearly predictive of intracellular drug retention; ongoing studies are directedat seeing if there is a correlation between in vivo polyglutamation of 5-DACTHF and tumor sensitivity.
The in vivo studies described here did not address the possibility of selective activation (polyglutamation) of 5-DACTHFby tumor cells, but other researchers have shown that malignantcells can be more efficient than host cells at transporting andpolyglutamating an antifolate drug. Ehrlich tumor cells accumulate polyglutamates of methotrexate to well over the level inmouse gut (18), and LI210 cells synthesize more methotrexatepolyglutamates than small intestine, liver, and marrow (19).Galivan et al. (20) showed that the antifolate methotrexate ispolyglutamated to a greater extent in hepatoma cells than inhepatocytes and that the relative enzyme activities of FPGSand folylpolyglutamate hydrolase favor greater polyglutamationin the hepatoma cells. Similarly, other researchers assayed theseenzyme activities in a number of malignant and nonmalignantcells and showed higher levels of FPGS and higher ratios ofFPGS/folylpolyglutamate hydrolase activities in malignantcells (21, 22). In addition, rapidly dividing cells show a greaterability to polyglutamate than do confluent cells (20). Thus, thespectrum of activity of 5-DACTHF, as well as its toxicity tohost cells, will most likely be a function of the ability of a cellor tissue to transport and polyglutamate 5-DACTHF as well asof its dependence on purine de novo biosynthesis.
ACKNOWLEDGMENTS
We wish to thank John Hill for synthesizing radiolabeled 5-DACTHF; Joan Humphreys, Mark Edelstein, and Sara Singer for
growing the cells used in this work; Bob Johnson for mass spectra;Paulo de Miranda, Richard DeAngelis, and Stephen Weller for adviceconcerning analysis of pharmacokinetic data; and Patricia Parker forexcellent technical assistance.
REFERENCES
1. l<USUIMI.Y. M., Takita. H.. and Gomez. G. Metabolic modulation andcellular de novo versus salvage metabolism. Antibiot. Chemother. (Basel), 28:86-93, 1980.
2. Becher, H., Weber, M., and Lohr, G. W. Purine nucleotide synthesis innormal and leukemic blood cells. Klin. Wochenschr., 56: 275-283, 1978.
3. Natsumeda. Y.. Prajda. N., Donohue, J. P.. Glover, J. L.. and Weber, G.Enzymic capacities of purine de novo and salvage pathways for nucleotidesynthesis in normal and neoplastic tissues. Cancer Res., 44: 2475-2479,1984.
4. Divekar, A. Y., and Hakala. M. T. Inhibition of the biosynthesis of 5'-phosphoribosyl-jY-formylglycinamide in sarcoma 180 cells by homofolate.Mol. Pharmacol., 11: 319-325, 1974.
5. Thorndike, J., Gaumont, Y., Kisliuk, R. L., Sirotnak, F. M., Murthy, B. R.,Nair, M. G., and Piper, J. R. Inhibition of glycinamide ribonucleotideformyltransferase and other folate enzymes by homofolate polyglutamates inhuman lymphoma and murine leukemia cell extracts. Cancer Res.. 49: 158-163, 1989.
6. Daubner, S. C., and Benkovic, S. J. Characterization of mammalian phos-phoribosylglycineamide formyltransferase from transformed cells. CancerRes., «.-4990-4997, 1985.
7. Beardsley, G. P., Moroson, B. A., Taylor, E. C., and Moran, R. G. A newfolate antimetabolite, 5,10-dideaza-5,6,7,8-tetrahydrofolate, is a potent inhibitor of de novo purine synthesis. J. Biol. Chem., 264: 328-333, 1989.
8. Kelley, J. L.. McLean, E. W., Cohn, N. K., Edelstein, M. P., Duch. D. S..Smith, G. K., Hanion, M. H., and Perone, R. Synthesis and biological activityof an acyclic analogue of 5.6.7,8-tetrahydrofolic acid, /V-[4-[[3-(2,4-diamino-l,6-dihydro-6-dihydro-6-oxo-5-pyTimidinyl)propyl]amino]benzoyl]-i--glu-tamic acid. J. Med. Chem., 33: 561-567. 1990.
9. Kelley, J. L. Novel compounds and use. United States Patent PB0499, 1988.10. Bognar, A. L., Osborne, C., Shane, B., Singer, S., and Perone, R. Folylpoly
glutamate synthetase-dihydrofolate synthetase. Cloning and high expressionof the Escherichia coli fole gene and purification and properties of the geneproduct. J. Biol. Chem., 260: 5625-5630, 1985.
11. Kozloff, L., and Lute, M. Folie acid, a structural component of T4 bacterio-phage. J. Mol. Biol., 12: 780-792, 1965.
12. Metzler, C. M.. Elfring. G. L., and McEwen, A. J. A package of computerprograms for pharmacokinetic modeling. Biometrics, 30: 562-563, 1974.
13. Perone, R., Hanlon, M. H., and Singer, S. C. a-Carboxl-linked glutamatesin the folylpolyglutamates of Escherichia coli. J. Biol. Chem., 267: 16356-16362, 1986.
14. Home. D. W., Krumdieck, C. L., and Wagner. C. Properties of folie acid y-glutamyl hydrolase (conjugase) in rat bile and plasma. J. NutrÃ.,///: 442-449, 1981.
15. Newell, D. R., Alison, D. L., Calvert, A. H., Harrap. K. R., Jarman, M.,Jones, T. R., Manteuffer-Cymborowska, M., and O'Connor, P. Pharmaco-kinetics of the thymidylate synthase inhibitor A"°-propargyl-5,8-dideazafolicacid (CB3717) in the mouse. Cancer Treat. Rep., 70: 971-979, 1986.
16. Valerino, D. M., Johns, D. G., Zaharko, D. A., el al. Studies on themetabolism of methotrexate by intestinal flora. 1. Identification and study ofbiological properties of the metabolite 4-amino-4-deoxy-jV10-methylpteroicacid. Biochem. Pharmacol.. 21: 821-831, 1972.
17. Samuels, L. L., Moccio. D. M., and Sirotnak, F. M. Similar differential fortotal polyglutamylation and cytotoxicity among various folate analogues inhuman and murine tumor cells in vitro. Cancer Res., 45: 1489-1495, 1985.
18. Fry, D. W., Anderson, L. A., Borst, M., and Goldman, 1. D. Analysis of therole of membrane transport and polyglutamation of methotrexate in gut andin the Ehrlich tumor in vivo as factors in drug sensitivity and selectivity.Cancer Res., 43: 1087-1092, 1983.
19. Poser, P. G., Sirotnak, F. M., and Chello, P. L. Differential synthesis ofmethotrexate polyglutamates in normal proliferativi and neoplastic mousetissues in vivo. Cancer Res., 41: 4441-4446, 1981.
20. Galivan, J., Johnson, T., Rhee, M., McGuire, J. J., Priest, D., and Kesevan,V. The role of folylpolyglutamate synthetase and 7-glutamyl hydrolase inaltering cellular folyl- and antifolylpolyglutamates. Adv. Enzyme Regul., 26:147-155. 1987.
21. Samuels, L. L., Goûtas,L. J., Priest, D. G., Piper, J. R., and Sirotnak, F.M. Hydrolytic cleavage of methotrexate polyglutamates by folylpolyglutamate hydrolase from various tumors and normal tissues of the mouse. CancerRes., 46: 2230-2235, 1986.
22. Sirotnak, F. M., and DeGraw, J. I. Selective antitumor action of folateanalogues. In: F. M. Sirotnak, J. J. Burchall, W. B. Ensminger, and J. A.Montgomery (eds.), pp. 43-91. Orlando, FL: Academic Press, 1984.
3211
on July 2, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1990;50:3207-3211. Cancer Res Mary H. Hanlon, Robert Ferone, Robert J. Mullin, et al. Analogue5-Deazaacyclotetrahydrofolate, an Acyclic Tetrahydrofolate
Metabolism ofin Vitro and In Vivo
Updated version
http://cancerres.aacrjournals.org/content/50/11/3207
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/50/11/3207To request permission to re-use all or part of this article, use this link
on July 2, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from