infecciones respiratorias virales: influenza aspectos ... · la primera descripción de un...
TRANSCRIPT

4 Ciencias clínicas
CIENCIASCLÍNICAS
No. 18 • Volumen 6
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos
• Dr. Adrián Valle de la O.1
• Dr. Carlos Jorge Castillo Gómez2
Desde la antiguedad, la mayor amenaza de calamidad universal asociada a una enfermedad infecciosa han sido las pandemias de influenza. Actualmente, con una población mundial que supera los 6,700 millo-nes de habitantes, hasta una pandemia “leve” podría causar muchos millones de muertes. Varios aconteci-mientos y factores recientes han generado una gran preocupación sobre una inminente pandemia de in-fluenza a corto plazo, la cual podría ser causada por el virus de influenza A H5N1 que ha circulado en los últimos años por Asia, Europa y África. En estos momentos, no es posible saber con precisión en qué momento podría hacer su aparición dicha pandemia. Sin embargo, lo que sí es cierto es que la realidad de una próxima pandemia no puede evitarse, y sólo se pueden tomar medidas para amortiguar sus efectos mediante una oportuna planeación basada en el co-nocimiento de la enfermedad, y que va más allá de cualquiera que se haya realizado hasta la fecha por algún país u organización.
Aspectos históricos
La influenza es uno de los problemas de salud más notorios y antiguos. La primera descripción de un padecimiento identificado como influenza se realizó en el año 412 a. C. por Hipócrates. El primer repor-te de una enfermedad epidémica, cuyos síntomas fueron probablemente de influenza, fue hecho en el año 1173-1174, y no se le considera pandémico. En cambio, el brote del año 1510 fue probablemente pandémico, se extendió desde África hacia Europa.1
En América, la primera aparición de un problema res-piratorio severo de este tipo se documentó en el año 1552 en Texcoco, y se le denominó “pestilencia ca-tarral”. No obstante, el primer brote pandémico reco-nocido por todos los autores ocurrió en 1580, y a éste en Italia se le llamó la “influencia planetaria”, por la asociación que hicieron con ciertos fenómenos astro-nómicos observados durante ese tiempo.2 Esta pan-demia se originó en Asia en el verano de aquel año, y se extendió hacia África, luego a Europa a lo largo de dos corredores, desde Asia menor y África noroc-cidental. Toda Europa fue afectada de sur a norte en seis meses, y la infección posteriormente se extendió a América.
La primera pandemia de influenza del siglo XVIII co-menzó en Rusia en 1729, durante los meses de pri-mavera y se extendió hacia el occidente, abarcó toda Europa en un periodo de seis meses. La siguiente pan-demia ocurrió 40 años después, entre 1781 y 1782; la mayoría de los autores piensan que se originó en China durante los meses de otoño, luego se extendió a Rusia, y de allí se expandió en círculos cada vez más extensos hasta comprender toda Europa en un periodo de ocho meses.1
La pandemia de 1830 a 1833 se equipara, en térmi-nos de severidad, con la pandemia de 1918 a 1920; esta pandemia comenzó en el invierno de 1830 en China y se extendió hacia el sur, por mar, y llegó has-ta Filipinas, India e Indonesia, y a través de Rusia se introdujo a Europa. La tasa de ataque fue de alrede-dor del 25%; sin embargo, su tasa de mortalidad no fue excepcionalmente alta.1
1 Profesor del Departamento de Ciencias Básicas de la División Ciencias de la Salud de la Escuela de Medicina del Tecnológico de Monterrey.2 Médico especialista en otorrinolaringología del Instituto Mexicano del Seguro Social, UMAA No. 65.

Ciencias clínicas 5
No. 18 • Volumen 6
Epidemiología
En la era moderna, el entendimiento de la influenza fue iniciado por Smith y cols. cuando aislaron el virus de la influenza tipo A en hurones.3 Años más tarde, en 1939, el virus de la influenza tipo B fue aislado por Francis y cols.,4 y en 1950, Taylor y cols. aislaron al virus tipo C.5 El descubrimiento en 1936 de que el virus de la influenza podía cultivarse en huevo em-brionado de gallina permitió el estudio extensivo de las propiedades del virus y el desarrollo de vacunas inactivadas. El fenómeno de la hemaglutinación, el cual fue descubierto por Hirst en 1941, condujo al desarrollo de métodos más simples y baratos para la determinación de la presencia del virus y la medición de los anticuerpos específicos contra el mismo.6
La naturaleza “explosiva” de la influenza epidémica, junto con el gran número de pacientes afectados y los hallazgos clínicos específicos de la enfermedad han dado credibilidad a los registros de esta infección realizados desde los inicios del siglo XVIII. El mejora-miento y mayor precisión de los datos registrados, así como el aislamiento y estudio del virus desde 1933, han proporcionado datos más precisos sobre el com-portamiento de esta infección durante los últimos cien años. La incidencia de brotes y el número de casos involucrados no ha descendido durante este periodo. La historia ha registrado epidemias frecuentes, casi anuales, de influenza en muchos países casi todos los años. La aparición súbita y sin advertencias de la in-fluenza, así como la impredecibilidad de la severidad de un brote en particular, son rasgos característicos de la enfermedad. Estos brotes están asociados a ta-sas de ataque del 5% al 30% de la población, lo que
origina un incremento significativo en la tasa de hos-pitalizaciones y de mortalidad, predominantemente entre personas de la tercera edad y muy jóvenes, y a la disrupción de los servicios y de la vida social.1
La historia registra nueve pandemias desde 1700, las cuales empezaron en un punto focal y se diseminaron rápidamente a través de todo el mundo para infectar a cientos de millones de individuos.7
La documentación precisa de una pandemia fue rea-lizada por primera vez en la pandemia que se expan-dió desde Rusia hacia Europa y Estados Unidos en los años 1889 a 1892; la infección se presentó como una enfermedad aguda del tracto respiratorio de inicio súbito y de corta duración. El número total de casos fue alto y las muertes fueron más numerosas entre infantes y personas mayores de 60 años; desafortuna-damente, pese a que del 25% al 30% de la población mundial fue infectada, únicamente existen registros de las muertes ocurridas en un pequeño porcentaje (0.05%) de los casos.
La pandemia de 1918 a 1920, conocida como la influenza española, es uno de los eventos más dra-máticos registrados en la historia de la medicina. El comienzo de la pandemia no se conoce. Algunos autores piensan que se originó en China; sin embar-go, los primeros brotes ocurrieron aproximadamente al mismo tiempo en Norteamérica, específicamente en Detroit, Carolina del Sur, y en la prisión de San Quentin, localizada en el estado de California, Es-tados Unidos. Esta pandemia se expandió por todo el mundo por unos seis a ocho meses, y dio lugar a una infección de una marcada e inusual virulencia
Figura 1. NC = No conocido. Principles and practice of clinical virology, 5th edition. John Wiley & Sons, LTD 2004:271-297.
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos
6 Ciencias clínicas
No. 18 • Volumen 6
que comúnmente causaba una forma nueva y severa de neumonía en los pacientes infectados. Se ha cal-culado que ocurrieron aproximadamente entre 40 y 50 millones de muertes, principalmente entre adultos jóvenes. Los efectos de esta pandemia causaron pá-nico internacional. El número de muertes registrado es probablemente sólo una fracción del número total real de muertes. Muchos países no hicieron registros. Además, la propaganda que circundaba respecto a la Primera Guerra Mundial de 1914 a 1918 socavó la precisión de los datos registrados en Europa, y la revolución Bolchevique en Rusia hizo que fueran ig-norados los efectos de la pandemia en ese país.1,7
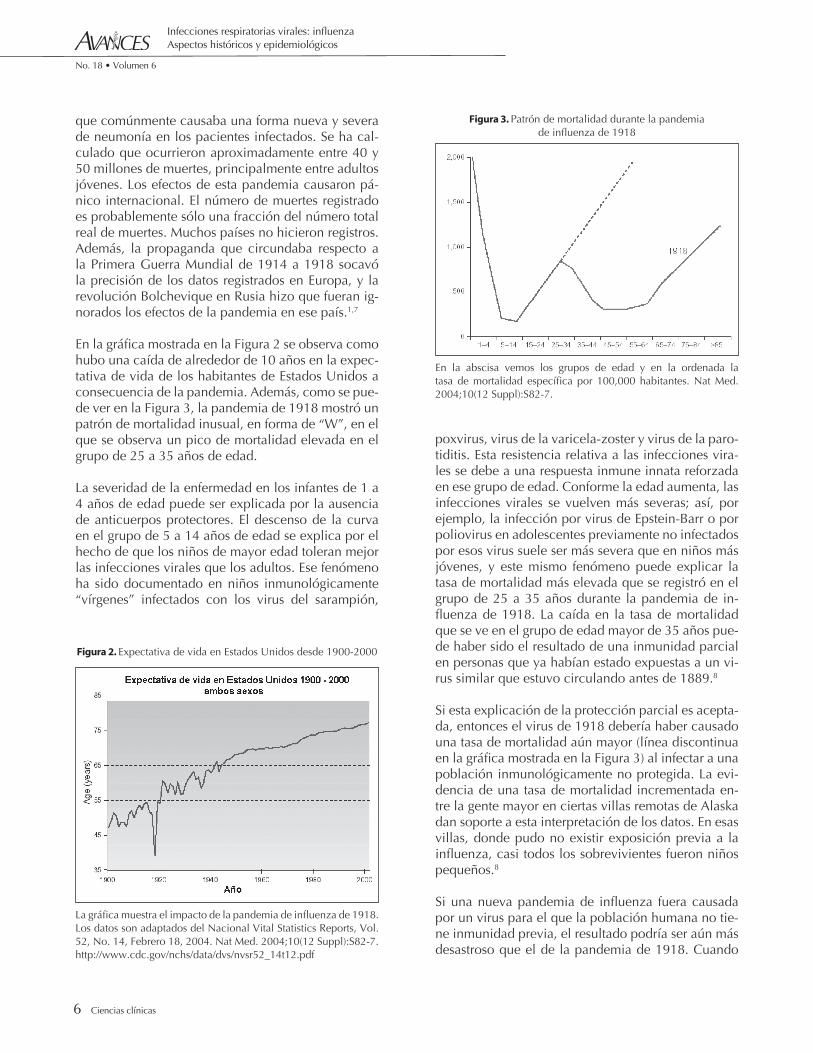
En la gráfica mostrada en la Figura 2 se observa como hubo una caída de alrededor de 10 años en la expec-tativa de vida de los habitantes de Estados Unidos a consecuencia de la pandemia. Además, como se pue-de ver en la Figura 3, la pandemia de 1918 mostró un patrón de mortalidad inusual, en forma de “W”, en el que se observa un pico de mortalidad elevada en el grupo de 25 a 35 años de edad.
La severidad de la enfermedad en los infantes de 1 a 4 años de edad puede ser explicada por la ausencia de anticuerpos protectores. El descenso de la curva en el grupo de 5 a 14 años de edad se explica por el hecho de que los niños de mayor edad toleran mejor las infecciones virales que los adultos. Ese fenómeno ha sido documentado en niños inmunológicamente “vírgenes” infectados con los virus del sarampión,
poxvirus, virus de la varicela-zoster y virus de la paro-tiditis. Esta resistencia relativa a las infecciones vira-les se debe a una respuesta inmune innata reforzada en ese grupo de edad. Conforme la edad aumenta, las infecciones virales se vuelven más severas; así, por ejemplo, la infección por virus de Epstein-Barr o por poliovirus en adolescentes previamente no infectados por esos virus suele ser más severa que en niños más jóvenes, y este mismo fenómeno puede explicar la tasa de mortalidad más elevada que se registró en el grupo de 25 a 35 años durante la pandemia de in-fluenza de 1918. La caída en la tasa de mortalidad que se ve en el grupo de edad mayor de 35 años pue-de haber sido el resultado de una inmunidad parcial en personas que ya habían estado expuestas a un vi-rus similar que estuvo circulando antes de 1889.8
Si esta explicación de la protección parcial es acepta-da, entonces el virus de 1918 debería haber causado una tasa de mortalidad aún mayor (línea discontinua en la gráfica mostrada en la Figura 3) al infectar a una población inmunológicamente no protegida. La evi-dencia de una tasa de mortalidad incrementada en-tre la gente mayor en ciertas villas remotas de Alaska dan soporte a esta interpretación de los datos. En esas villas, donde pudo no existir exposición previa a la influenza, casi todos los sobrevivientes fueron niños pequeños.8
Si una nueva pandemia de influenza fuera causada por un virus para el que la población humana no tie-ne inmunidad previa, el resultado podría ser aún más desastroso que el de la pandemia de 1918. Cuando
Figura 2. Expectativa de vida en Estados Unidos desde 1900-2000
La gráfica muestra el impacto de la pandemia de influenza de 1918. Los datos son adaptados del Nacional Vital Statistics Reports, Vol. 52, No. 14, Febrero 18, 2004. Nat Med. 2004;10(12 Suppl):S82-7. http://www.cdc.gov/nchs/data/dvs/nvsr52_14t12.pdf
En la abscisa vemos los grupos de edad y en la ordenada la tasa de mortalidad específica por 100,000 habitantes. Nat Med. 2004;10(12 Suppl):S82-7.
Figura 3. Patrón de mortalidad durante la pandemiade influenza de 1918
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

Ciencias clínicas 7
No. 18 • Volumen 6
hablamos de una pandemia de influenza nos referi-mos a un brote de la infección que se originó en una región geográfica específica, luego se disemina por todo el mundo, infectando a un alto porcentaje de la población, y es causado por una cepa del virus de la influenza que no pudo haberse originado por mutación de cepas previamente circulando. Toman-do en cuenta esta definición, los brotes de influenza de 1932 a 1933 y de 1947 a 1948 fueron causados por virus relacionados entre sí y con el virus que cau-só la pandemia de influenza de 1918 a 1920; por lo tanto, por definición, los brotes de influenza de 1932 y 1947 no fueron pandemias.7
Sin embargo, efectivamente ocurrió una pandemia de influenza en 1957, a la cual se la ha conocido como “influenza asiática”. Esta pandemia se originó en la provincia de Yunan, China, en febrero de ese año; alcanzó Hong Kong en abril, y luego se expandió rá-pidamente a Singapur, Taiwán y Japón. En este mo-mento, la Organización Mundial de la Salud (OMS) se percató de la existencia de este brote por un nuevo subtipo viral. Posteriormente, este brote se diseminó al hemisferio sur, durante el invierno de esa región geográfica, y desde allí se extendió al hemisferio nor-te, donde ocurrieron epidemias durante el invierno. La infección alcanzó a India, Australia e Indonesia en mayo de 1957; a Pakistán, Europa, Norteamérica y
Medio Oriente, en junio de ese año; al sur de África, Sudamérica, Nueva Zelanda y a las islas del pacífico, en julio; y a África central, oriental y occidental, Eu-ropa del este y al Caribe, en agosto del mismo año. Una segunda oleada se observó en 1958 con múlti-ples brotes en Europa, Norteamérica, Unión Soviética y Japón. Esta pandemia se extendió por todo el mun-do e infectó del 40% al 50% de la población, y causó más de un millón de muertes, principalmente en la población mayor de 60 años de edad; sin embargo, las fatalidades entre víctimas más jóvenes fueron más sobresalientes en los primeros meses de la pandemia, y esto es algo que se ha observado en otras pande-mias.1,7
En el virus de la pandemia de 1957, dos nuevas glu-coproteínas hicieron su aparición en la superficie del mismo. La hemaglutinina (HA) era del subtipo H2, y mostraba sólo una homología del 66% en su se-cuencia de aminoácidos en relación a la del virus pandémico previo, el cual había sido del subtipo H1. La neuraminidasa (NA) de este nuevo virus era del subtipo N2, y sólo compartía un 37% de la secuencia de aminoácidos con la NA, subtipo N1, del virus de la influenza española. Así, después de 39 años de vi-rus H1N1, apareció un H2N2 contra el cual no había una protección inmunológica preexistente.8
Figura 4. Pandemia de influenza de 1957-1958
El punto de origen se ha señalado con un cuadrado negro y ocurrió en febrero de 1957. Líneas de expansión de la pandemia ( ); los números en la figura se refieren al número de meses, a partir de febrero (mes cero) que tomó a la pandemia extenderse; los números acompañan a las flechas correspondientes. J Appl Microbiol 2001;91(4):572–579.
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

8 Ciencias clínicas
No. 18 • Volumen 6
Once años más tarde, en 1968, otra vez se presen-tó un cambio en las glucoproteínas de superficie del virus y éste se hizo pandémico. En este virus, sólo los genes que codificaban para la HA y la PB1 ha-bían cambiado. Este virus era del subtipo H3N2; las glucoproteínas H3 del nuevo virus y la H2 del virus previo diferían en más del 60% en su secuencia de aminoácidos. La conservación de la neuraminidasa N2 pudo haber proporcionado cierta protección a la población que había estado previamente expuesta al virus H2N2.8
Cabe mencionar que hubo un cuarto brote pandé-mico en 1976-1977 y fue debido a un virus muy re-lacionado con la cepa del virus que desapareció en el brote de 1947. Esta pandemia estuvo, por lo tanto, limitada a los jóvenes, puesto que muchas personas nacidas antes de 1947 ya habían tenido contacto pre-viamente con este virus y habían conservado inmuni-dad a la infección. Antes de 1976-1977 la aparición de un nuevo virus pandémico se había acompañado de la desaparición del subtipo viral previo circulante entre los humanos; sin embargo, desde 1976-1977, los subtipos viejo y nuevo han circulado simultá-neamente, y esta conducta continúa hasta nuestros días.7
Aunque no hay evidencia suficientemente sólida, se piensa que la introducción de este virus H1N1 fue el resultado de los ensayos de vacunación realizados en el lejano Oriente, en el cual varios miles de reclutas militares fueron inmunizados con virus vivos H1N1. Desafortunadamente, como ya se ha mencionado, este virus y sus descendientes continúan circulando junto con el subtipo H3N2, desde entonces.8
No se ha identificado ningún patrón discernible entre las pandemias que permita predecir cuándo ocurrirá una futura pandemia. Sin embargo, los registros his-tóricos indican que el periodo de tiempo más largo entre dos pandemias, a lo largo de los últimos 150 años, es de 40 años, lo cual sugiere que la próxima pandemia ocurrirá antes del año 2017, y no hay nada que permita sugerir que esta situación cambiará en el futuro.1
Variación antigénica del virus
Los virus de la influenza evolucionan de forma cons-tante por acumulación de mutaciones puntuales en cada uno de sus ocho segmentos de RNA, lo que conduce a cambios en las proteínas virales y subse-cuentemente a nuevas variantes antigénicas del virus,
esto permite la evasión de la respuesta inmune y la aparición de brotes regulares de la enfermedad que en ocasiones pueden causar epidemias de impacto global. Las modificaciones antigénicas importantes son llamadas “cambio antigénico”, mientras que los cambios antigénicos menores son llamados “desvia-ción antigénica”.
Cambio antigénico mayor (Antigenic shift)Desde 1933, los virus que han causado tanto brotes pandémicos como epidémicos han sido aislados y comparados en el laboratorio. Los virus aislados de pacientes entre los años 1933 a 1946 y entre 1947 a 1956 muestran una amplia variación, y las pruebas de inhibición de la hemaglutinación (HI) utilizando antisueros obtenidos de hurones infectados con el vi-rus no muestran reactividad serológica cruzada. Sin embargo, los datos indican que los dos grupos de vi-rus, representados por las cepas o subtipos A/PR/8/34 y A/FM/1/47, están relacionados y se incluyeron en el subtipo H1. Además, las secuencias de RNA re-cuperadas y amplificadas mediante PCR (polymerase chain reaction) de tejido pulmonar post mortem de dos soldados norteamericanos muertos en septiem-bre de 1918 y de una tercera muestra recuperada de una mujer Inuit de Alaska –que había sido ente-rrada en permacongelamiento desde su muerte por influenza en noviembre de 1918– indicaron que la pandemia de 1918 a 1920 fue también causada por un virus de la influenza A(H1). En cambio, los sub-tipos que emergieron y causaron las pandemias de 1957 y 1968 no están relacionados entre ellos ni con el subtipo H1N1, y no pueden haberse originado por mutación de los subtipos precedentes.
Virus con tres subtipos de hemaglutininas (H1, H2 y H3) y dos sub-tipos de neuraminidasa (N1 y N2) han sido identificados en huma-nos. Los cuadros indican la introducción de las pandemias H1NI, H2N2 y H3N2 en los años 1918, 1957 y 1968, respectivamente. En 1976-1977 un virus H1N1 similar al que circulara por 1950 fue reintroducido. Las líneas discontinuas indican la ausencia de aisla-mientos virales y la existencia sólo de evidencia indirecta basada en datos sexológicos de las cepas circulantes en ese tiempo. Nat Med. 2004;10(12 Suppl):S82-7.
Figura 5. Virus de influenza A circulandoen la población humana.
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos
Ciencias clínicas 9
No. 18 • Volumen 6
Esta especificidad absoluta de la HA de diferentes subtipos indica que en los años en que se registraron pandemias habían emergido nuevos subtipos virales en una población que no tenía inmunidad para ese nuevo subtipo. Esto es lo que se conoce como “cam-bio antigénico” (antigenic shift). Ahora bien, puesto que es la presencia de anticuerpos contra la HA viral la que indica inmunidad, la infección por virus de la influenza en los años previos no indujo anticuerpos HI contra las nuevas cepas virales emergentes. Como resultado de la infección, se adquiere inmunidad contra el nuevo subtipo por un periodo de años, y los subsecuentes brotes son, por lo tanto, más limitados. Sin embargo, después de 10 a 40 años, un nuevo sub-tipo emerge con una HA que no presenta reacción cruzada con los anticuerpos contra la HA del virus previo, y al que la población es, por lo tanto, alta-mente susceptible, iniciándose así un nuevo ciclo de pandemia y epidemias de influenza.
Aunque las HA de diferentes subtipos de virus de la influenza no muestran reactividad cruzada en las pruebas de HI, existe cierta relación entre ellas, lo cual puede ser observado a través del fenómeno lla-mado “pecado antigénico original” (original antigenic sin), descrito desde 1953. Este fenómeno se refiere a la observación de que la infección por el virus de la influenza induce anticuerpos contra el virus actual-mente infectante y, de alguna manera, también hace que se produzcan nuevamente anticuerpos contra otros subtipos del virus con los cuales el individuo ha tenido experiencias previas. De hecho, los títulos de anticuerpos contra los subtipos previos pueden ser varias veces más grandes que los dirigidos contra el virus actualmente infectante. Esto probablemente se debe a la estimulación de las células B de memoria que persisten después de la infección, que ahora son éstas estimuladas por el nuevo virus, lo cual sugiere que existe cierta relación en los antígenos de la HA de diferentes serotipos que no son detectados en las pruebas de HI ni por otras pruebas serológicas.
La importancia de los cambios antigénicos es eviden-te, pero a pesar de cien años de intensa investigación, el origen de estos nuevos virus es incierto. El proble-ma ha sido de gran interés para investigadores; los ex-perimentos realizados, los estudios epidemiológicos, y la imaginación han generado diversas teorías. Por ejemplo, se ha sugerido que los nuevos serotipos vi-rales podrían surgir por mutación espontánea, y hasta se ha llegado a sugerir que proceden de fuentes “cós-micas”; sólo una minoría de científicos consideran estas teorías como explicaciones válidas, y no existe evidencia convincente para apoyar a alguna de ellas. Así, se esperaría que las mutaciones generadas al azar produjeran cepas intermediarias, y tales cepas no han sido identificadas. Análisis detallados han mostrado que los nuevos subtipos virales tienen múltiples dife-rencias en sus secuencias genéticas, y es difícil imagi-nar que todas éstas ocurran simultáneamente.
La teoría más ampliamente aceptada para el origen de nuevos subtipos del virus de la influenza es que son virus “reordenados” que resultan de la doble in-fección de una misma célula. La aparición de un nue-vo subtipo de virus es seguida por la desaparición del subtipo previo, con la única excepción de 1976, y por lo tanto parece improbable que la infección dual sea por dos subtipos virales humanos. Sin embargo, el virus de la influenza tipo A infecta también otras especies, incluyendo caballos y, particularmente, aves. Una infección dual con un virus humano y otro de origen animal o aviario en una especie que pue-da infectarse y sostener la replicación viral de ambos subtipos virales puede resultar en un reordenamiento genético que conduzca a la producción de un virus con determinantes antigénicos de superficie del sub-tipo no humano y con la infectividad del subtipo hu-mano. Esta teoría es científicamente sostenible; este reordenamiento genético puede ser producido en el laboratorio con subtipos humanos y animales, y puede ser identificado cuando se le da seguimiento a infecciones mixtas de animales y aves. Asimismo, los
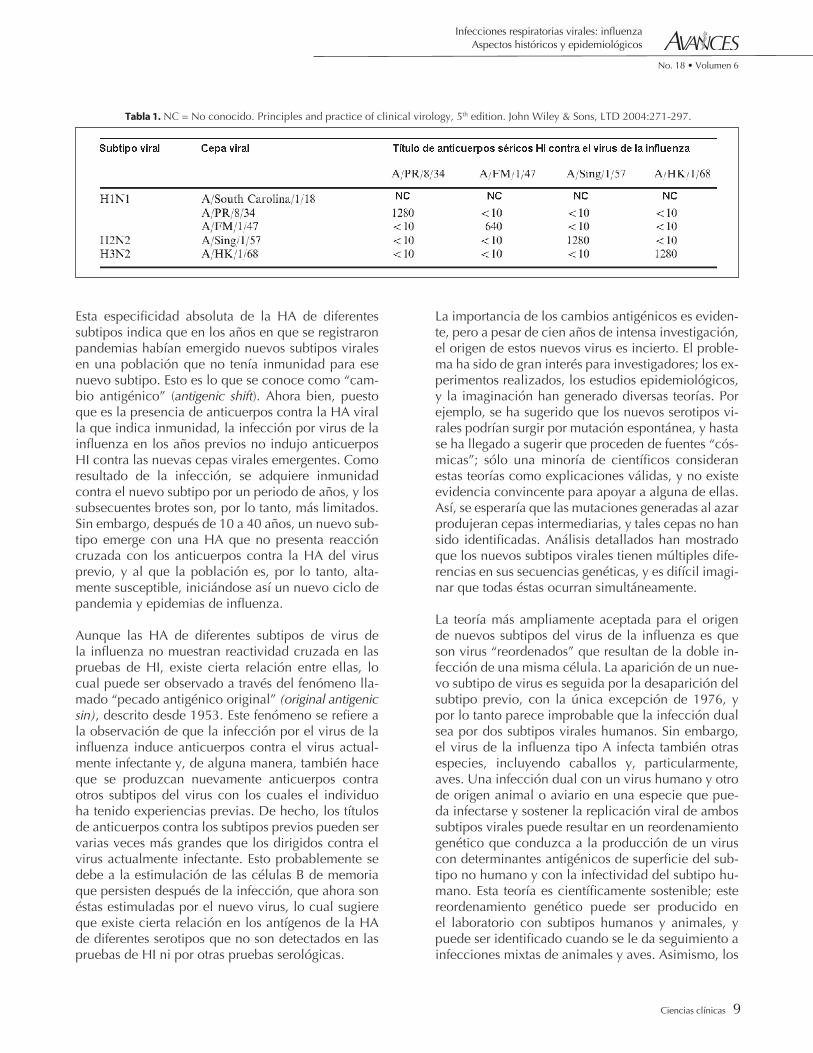
Tabla 1. NC = No conocido. Principles and practice of clinical virology, 5th edition. John Wiley & Sons, LTD 2004:271-297.
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

10 Ciencias clínicas
No. 18 • Volumen 6
virus de la influenza pueden cruzar la barrera interes-pecie, y el análisis antigénico de dos subtipos virales humanos ha revelado similitudes entre la HA de estos subtipos y la de conocidos subtipos aviarios.
Finalmente, se ha sugerido que hay un número limi-tado de subtipos de virus de la influenza que son re-ciclados en la población humana. La evidencia para esta teoría proviene de estudios seroepidemiológicos de anticuerpos contra el virus de la influenza en suero tomado en diferentes tiempos de sujetos de diferentes edades. De este modo, pueden detectarse anticuer-pos a lo largo de un periodo de cien años, puesto que la presencia de anticuerpos contra un subtipo viral particular en una persona de edad conocida puede indicar las fechas en que este virus estuvo en circu-lación en la población, incluso antes de que fuera posible el aislamiento del virus. El resultado de estos análisis indican cuándo ocurrieron las pandemias y cuáles serotipos virales estuvieron involucrados. De esta manera, se mostró que las pandemias que co-menzaron en los años 1889, 1900, 1918, 1957, 1968 y 1976 fueron debidas a virus de la influenza tipo A, subtipo respectivamente (ver la Tabla 1). Un ciclo de aproximadamente 70 años involucrando a estos tres subtipos virales podría satisfacer este requerimiento. Sin embargo, la evidencia para apoyar esta teoría es frágil. Aunque los estudios han detectado la presen-cia de anticuerpos para el virus de la influenza tipo A(H3), el cual fue visto por primera vez en 1968 en sueros de pacientes de edad avanzada recolectados antes de la pandemia, no se ha encontrado ningún reservorio animal o humano para el virus antes de la fecha en que fue encontrado.
Sin embargo, una vez más, puede argumentarse que no se necesita un reservorio para apoyar a esta teoría. Los reordenamientos genéticos pueden aparecer todo el tiempo y sólo cuando emerge un reordenamiento que es capaz de diseminarse rápidamente en la po-blación humana, y cuando el estatus inmune de la población ha menguado por la muerte de las genera-ciones mayores, entonces puede ocurrir una pande-mia producida por un virus similar que circuló años atrás. Resulta claro, de todo lo mencionado hasta este momento, que el origen de los nuevos subtipos virales no está probado aún, pero los avances en la vigilancia epidemiológica de la influenza, en el diag-nóstico de influenza y en el análisis antigénico viral podrán ser aplicados al virus que cause la próxima pandemia, y esto puede ayudar a probar cuál es el mecanismo que conduce a la producción de cambios antigénicos.7,9
Desviación antigénica (Antigenic drift)Cuando los virus de varias epidemias entre las pande-mias son comparados mediante pruebas de HI, es po-sible encontrar ciertas diferencias entre las cepas. Así, aunque todos los virus pertenecen al mismo subtipo, no presentan una reacción cruzada completa. Estos cambios son denominados “desviación antigénica” (antigenic drift). El grado de reactividad antigénica cruzada tiende a disminuir conforme se incrementa la diferencia temporal entre dichos virus, y puesto que el grado de inmunidad cruzada está directamen-te relacionado con el grado de reactividad antigénica cruzada, la inmunidad contra las variantes del mismo virus disminuye al pasar el tiempo.
¿Cómo se originan estas desviaciones antigénicas? La teoría aceptada propone que las variantes virales ocurren naturalmente por mutación, y éstas son selec-cionadas por la presión de la inmunidad humoral en una población inmune o parcialmente inmune. Pues-to que estas nuevas variantes no son completamente neutralizadas por los anticuerpos dirigidos contra las cepas virales preexistentes, éstas con capaces de cau-sar nuevas epidemias.7,9
Ecología de los virus de influenza A
Las aves silvestres acuáticas migratorias, tanto de la-gos como costeras, son los reservorios naturales de toda la constelación genética de los virus de influen-za, en particular las especies de los órdenes Anserifor-me (los patos), Passeriforme (los gansos) –ambas aves silvestres migratorias de lagos– y las del orden Chara-driforme (gaviotas y golondrinas marinas, entre otras) –aves acuáticas migratorias de las costas. De hecho, todos los subtipos de HA, incluidos los identificados en los últimos 17 años (HAs 14, 15, y 16), se aislaron de patos y gaviotas, lo que demuestra que ocurrieron mayores cambios genéticos virales en estas aves.2,10-12 En todas estas aves silvestres, los virus han alcanzado un estado evolutivo estable, y por lo general no cau-san trastornos clínicos en ellas.2,13,14
Los virus no patogénicos de influenza aviar son ubi-cuos en la naturaleza, sobre todo, como ya se comen-tó, en las aves acuáticas, y todos los diferentes sub-tipos de influenza A son perpetuados en estas aves. En los patos silvestres, el virus se replica preferente-mente en las células epiteliales del tracto gastrointes-tinal, sin causar signos de enfermedad, y es excretado en grandes cantidades por las heces. Asimismo, se ha aislado el virus del agua de los lagos. Esta infor-mación nos indica que las aves acuáticas tienen una
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

Ciencias clínicas 11
No. 18 • Volumen 6
forma muy eficiente de transmitir virus: a través de la contaminación, con materia fecal, de los suministros de agua y del suelo.
Puesto que un gran número de patos nacen cada año en todo el mundo, muchos de ellos son infectados por el virus que se encuentra en el agua de los lagos. En Norteamérica, un gran número de patos jóvenes susceptibles se reúnen cada año en los lagos cana-dienses, lo que explica la alta incidencia (hasta del 30%) de infección por el virus de la influenza en los patos canadienses. La transmisión por heces también puede ser fuente de infección durante la migración anual otoñal, hacia el sur, que efectúan estas aves. Los virus que se eliminan en las heces frescas conta-minan el suelo y agua de los lagos o canales de otras regiones, en donde las aves domésticas, en especial aquéllas que se crían en espacios abiertos, buscan alimento y consumen agua, lo que constituye una efi-ciente ruta de transmisión para el virus. 2,13,14
La naturaleza avirulenta de la infección por virus de la influenza aviar en patos puede ser el resultado de la adaptación del virus a su hospedero a lo largo de los siglos, creando así un reservorio que asegura la perpetuación del virus. Esto fuertemente sugiere que los patos ocupan una posición única y muy importan-te en la historia natural de los virus de la influenza.
Los virus de influenza de las aves acuáticas silvestres llegan a diferentes especies de mamíferos de manera directa o a través de aves domésticas, y se han des-crito infecciones en mamíferos marinos como focas y ballenas; mamíferos terrestres como cerdos, caballos,
visones, gatos, tigres y leopardos; y en fecha más re-ciente, también en perros.2,13-15 No obstante, aunque los virus de la influenza puedan infectar una gran va-riedad de aves y mamíferos, el hospedero natural del virus son las aves acuáticas. Cuando otras especies de animales –incluyendo pollos, pavos, cerdos, caba-llos– y humanos son infectados, éstos son considera-dos hospederos aberrantes, lo que origina mutaciones en los virus como un mecanismo de adaptación para una óptima replicación y transmisión en la nueva es-pecie.2,14,16
El estudio de la ecología del virus de la influenza ha conducido a la hipótesis de que todos estos virus de-rivan de los reservorios de influenza aviar (ver Figura 6). En apoyo a esta teoría están los análisis filogenéti-cos de las secuencias de ácidos nucleicos de virus de la influenza tipo A en una gran variedad de hospede-ros, regiones geográficas y subtipos virales. El análisis
del gen de la nucleoproteína (NP) muestra que los virus de la influenza aviar han evolucionado en cin-co linajes específicos: el equino antiguo –el cual no se ha aislado en años–, el equino reciente, el de las gaviotas, el de los cerdos y el de los humanos. El virus de la influenza humano y el porcino evolucionaron de un ancestro común que parece haber sido un virus aviar que, como el virus de la influenza porcino que circula en Europa, derivó todos sus genes de fuentes aviares.2,14
Asimismo, el análisis filogenético de un mismo sub-tipo de virus encontrado en aves migratorias muestra linajes diferentes entre aquéllos que circulan en Eu-ropa y Asia y los del continente Americano, lo que pone en evidencia que las migraciones longitudinales tiene un papel clave y son, de hecho, más importan-tes que las migraciones intercontinentales en la difu-sión global de los virus.
Del mismo modo, a través de análisis filogenéticos de los cambios en las secuencias de aminoácidos, se ha podido ver que los virus de influenza aviar, en contraste con las cepas de mamíferos, tiene bajas tasas evolucionarias. De hecho, podría decirse que los virus de la influenza en aves acuáticas se encuen-tran en una especie de estancamiento evolutivo, sin evidencia de cambios evolutivos francos en las últi-mas seis o siete décadas. Los cambios de nucleótidos han continuado con la misma tasa de cambio en las
Figura 6. Las aves acuáticas son el principal reservoriodel virus de influenza tipo A para aves y mamíferos.
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

12 Ciencias clínicas
No. 18 • Volumen 6
cepas aviarias y en las de mamíferos; sin embargo, estos cambios ya no dan lugar a cambios en los ami-noácidos en las cepas aviarias, mientras que en los mamíferos, los ocho segmentos genéticos del virus de la influenza continúan acumulando cambios en los aminoácidos.
El alto nivel de conservación genética en los virus aviarios sugiere que éstos se están aproximando a, si no es que ya han alcanzado, un estado óptimo en el cual los cambios en las secuencias de nucleótidos ya no proporcionan ninguna ventaja selectiva. Esto tam-bién significa que la fuente de los genes para los virus de influenza pandémicos existen fenotípicamente sin cambios en los reservorios aviarios. Y la implicación más importante de los estudios filogenéticos es que los virus ancestrales que causaron la pandemia de in-fluenza española en 1918, así como los virus que pro-porcionaron los genes para las pandemias de 1957 y 1968, aún están circulando en las aves silvestres con escasas mutaciones o incluso sin cambio alguno.14
El hombre desempeña un papel importante como transmisor directo o indirecto del virus, ya sea por transporte mecánico a través de la ropa, jaulas, ali-mento, equipos o vehículos contaminados, o bien, por el contrabando de aves silvestres, de ornato, de pelea y comerciales. Los virus altamente patogéni-cos de influenza aviar (HPAIs) pueden sobrevivir por largos periodos de tiempo en el ambiente, especial-mente cuando la temperatura es baja. Por ejemplo, el HPAI H5N1 puede sobrevivir en las heces por, al menos, 35 días a baja temperatura (4ºC); a 37ºC se ha demostrado que este virus sobrevive por 6 días.17-19
Es importante reconocer el papel del cerdo como hospedero intermediario, adaptador y amplificador de ciertos subtipos virales para infecciones en huma-nos. Con frecuencia, los cerdos y las aves de corral son criados en las mismas granjas comerciales, e in-cluso en ocasiones se alimenta a los cerdos con ca-dáveres de aves de corral. Lo anterior es indeseable porque facilita la transmisión entre especies del virus de la influenza.
Hasta este punto, la ruta de transmisión ha sido de tipo fecal-oral. Después de la transmisión a cerdos, caballos o humanos, el método de diseminación de la influenza es principalmente por vía respiratoria. Los gatos pueden infectarse tanto por vía respiratoria como por vía fecal-oral.2,14,20
Influenza aviar: emergencia del virus H5N2 en Norteamérica
En 1983 se aisló un virus de influenza aviar H5N2 que infectó pollos y pavos en Pennsylvania, y que se volvió altamente patogénico para aves de corral. Los estudios virológicos y epidemiológicos no mos-traron evidencia de transmisión a humanos. El virus fue eventualmente erradicado mediante cuarentenas y el exterminio de más de 17 millones de aves con un costo directo de 60 millones de dólares y un costo indirecto para la industria de más de 250 millones de dólares.
Posteriormente, emergió un virus altamente patogéni-co de influenza aviar (HPAI) H5N2 en pollos domés-ticos en México.21 En octubre de 1993, la producción de huevos disminuyó y se incrementaron las muertes entre los pollos mexicanos, se obtuvo evidencia se-rológica de infección por el virus H5N2 en dichos pollos. El virus fue aislado en mayo de 1994, des-pués de haberse extendido ampliamente en el país, y sólo causaba un síndrome respiratorio leve en po-llos. Dado que la erradicación del virus a través de la destrucción de las aves infectadas representaba un gran obstáculo para la industria de las aves de corral en el país, fue posible llevar a cabo un “experimento de campo” para determinar el destino de las cepas virales avirulentas después de repetidos ciclos de replicación en millones de pollos. Hacia finales de 1994, se encontró que el virus había mutado y aho-ra contenía una HA fácilmente escindible, pero aún continuaba siendo sólo levemente patogénico para
Figura 7. El cerdo puede actuar como hospedero intermediarioen la generación de virus con reordenamiento genético
y potencial pandémico. Nat Med 1998;4:1122-3.
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

ProGin - - ArgGluThrArg/Gly ProGin ArgLys Arg Lys ThrArg/Gly
HA1 HA2 HA1 HA2
1994 1995
Ciencias clínicas 13
No. 18 • Volumen 6
pollos. Sin embargo, en cuestión de meses el virus se volvió letal para las aves de corral. Las principales zonas productoras de aves del país fueron infectadas, incluyendo Aguascalientes, Distrito Federal, Gua-najuato, Guerrero, Hidalgo, Jalisco, México, More-los, Puebla, Querétaro, Sonora y Veracruz. El análisis filogenético indicó que la epidemia había sido inicia-da por la introducción de un único virus H5 del linaje norteamericano en los pollos mexicanos. El virus fue erradicado mediante cuarentena y el uso de vacunas inactivadas. No obstante, la variedad de baja patoge-nicidad siguió circulando y se extendió a Guatemala y El Salvador.2,14,21
Pero, ¿cómo se diseminó este virus? El brote de in-fluenza aviar H5N2 en Pennsylvania de 1983 a 1984 demostró que el mercado de aves vivas juega un pa-pel muy importante en la diseminación del virus de la influenza en especies aviares. El virus H5N2 estuvo siendo aislado hasta 1986 y causó infección subclí-nica en pollos en los mercados, de la misma manera en que lo hizo en 1997 el virus H5N1 en Hong Kong. Mas aun, los patos en los mercados de Estados Unidos estaban infectados con muchos subtipos diferentes de virus de la influenza tipo A, incluyendo el subtipo H2N2 relacionado antigénicamente con el virus de la influenza asiática de 1957 y el cual ha desaparecido de los seres humanos.
Figura 8. Cambios moleculares asociados con la emergencia del HPAI H5N2 en pollos en México.
En 1994 una cepa no patogénica del virus H5N2 fue aislada en pollos mexicanos y se encontró que estaba relacionada con un virus H5N2 aislado en Delaware, Estados Unidos, en 1991. Los aislamientos del virus en 1994 a partir de pollos se replicaban principalmente en el tracto respiratorio, se diseminaba rápidamente entre los pollos, y no era altamente pa-togénico. Durante el siguiente año, el virus se volvió altamente patogénico, y la HA había adquirido una inserción de dos aminoácidos básicos (Arg-Gly) y una mutación de Glu a Lys en el sitio de escisión HA1/HA2.
Se ha postulado que un virus de la influenza H5N1 no patogénico se diseminó de aves costeras migratorias hacia los patos por conta-minación fecal del agua. El virus fue transmitido de los patos a los pollos y se estableció en el mercado de aves vivas en Hong Kong. Durante la transmisión entre las especies, el virus se volvió alta-mente patogénico para pollos, y ocasionalmente fue transmitido de los pollos a los humanos en los mercados. A pesar de su alta patogenicidad para pollos (y humanos), el virus no era patogénico ni para patos ni para gansos.
Figura 9. Emergencia del virus H5N1 en Hong Kong.
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos
Transmitido por heces y aguaTransmitido por aire, heces y agua
Desarrollo de una cepaaltamente patogénicay transmisión fecal

14 Ciencias clínicas
No. 18 • Volumen 6
Influenza aviar en humanos
La incidencia de influenza aviar en humanos se ha incrementado durante los últimos años. Inicialmen-te, los casos de influenza aviar H7N7 en humanos habían ocurrido en asociación con brotes en las aves de corral y se manifestaba como una conjuntivitis au-tolimitada. En 1997, un brote a gran escala de HPAI H5N1 ocurrió entre las aves de corral de Hong Kong, se documentaron 18 casos humanos, principalmente adultos jóvenes, de los cuales seis murieron.
El caso índice de influenza aviar H5N1 en humanos fue el de un niño de 3 años de edad de Hong Kong, sin antecedentes de enfermedades subyacentes, y que murió el 21 de mayo de 1997, cinco días des-pués de su hospitalización en una unidad de cuida-dos intensivos, con diagnóstico final de síndrome de Reye, neumonía viral aguda por virus de la influenza e insuficiencia respiratoria. Del aspirado traqueal se aisló el virus; ninguna bacteria patogénica creció a partir de las muestras respiratorias. Mediante pruebas de HI se vio que este virus no presentaba reacción cruzada con antisueros de hurón contra los virus re-cientemente aislados de humanos y cerdos. Posterior-mente, se identificó al virus como subtipo H5N1, de origen aviar, y se vio que era altamente patogénico para pollos.22
Afortunadamente, el virus no tenía la capacidad para transmitirse eficientemente de persona a persona, y el brote pudo ser detenido mediante el sacrificio masivo de aves de corral en esa región. Después de ese brote de HPAI H5N1 en 1997, se aisló en Hong Kong el virus de influenza tipo A subtipo H9N2 en
marzo de 1999, que causó una enfermedad autoli-mitada en dos niños de 1 y 4 años de edad, respec-tivamente. Con respecto a este virus, se ha visto que éste se encuentra ampliamente distribuido en las aves de corral en Asia, y se ha ido documentado que has-ta el 4% de los donadores de sangre en Hong Kong presentan anticuerpos contra este virus.23 Dos brotes subsecuentes de influenza aviar ocurrieron en Hong Kong, uno en 1999 y otro en 2003, ambos causados por HPAI H5N1. A principios de 2003, tres miem-bros de una familia se infectaron por HPAI H5N1 en China; dos, murieron.24 En diciembre del 2003 apa-recieron dos brotes de HPAI H5N1 en Corea y China, y desde el 2003 el brote se ha extendido por todo el mundo, en lo que sería el brote más grande que se haya registrado en la historia de las aves de corral.17 Los casos humanos de HPAI H5N1 han seguido a los brotes entre las aves de corral, con un total de 412 casos y 256 fatalidades (62.13%) hasta marzo 23 del 2009.25 Los países más afectados han sido Indonesia, Vietnam, Egipto y China. Otro virus de influenza A, subtipo H7N7, causó un brote en los países bajos, se documentaron un total de 89 casos con conjuntivitis y sólo una fatalidad (ver Tablas 2 y 3).
Influenza aviar y la pandemia de 1918
La única forma de entender cómo es que el virus de la pandemia de 1918 fue tan altamente patogénico en adultos sanos es reconstruir sus genes y estudiar sus propiedades. Los virus de la influenza tienen varias proteínas que están implicadas en su virulencia: las proteínas de superficie HA y NA, el complejo de la polimerasa (incluyendo las proteínas PB1, PB2 y PA) y las proteínas no estructurales (NS). Taubenberger y
Figura 10. Virus de la influenza aviar (*) circulantes desde 1900.
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos
H1H2
H3H1
H7*
H5*
H9*
1915 1925 1935 1945 1955 1965 1975
1977
1980 1996
1997
2003
1998 1999
2003-2006
200220042003
1985 1995 2005
Ciencias clínicas 15
No. 18 • Volumen 6
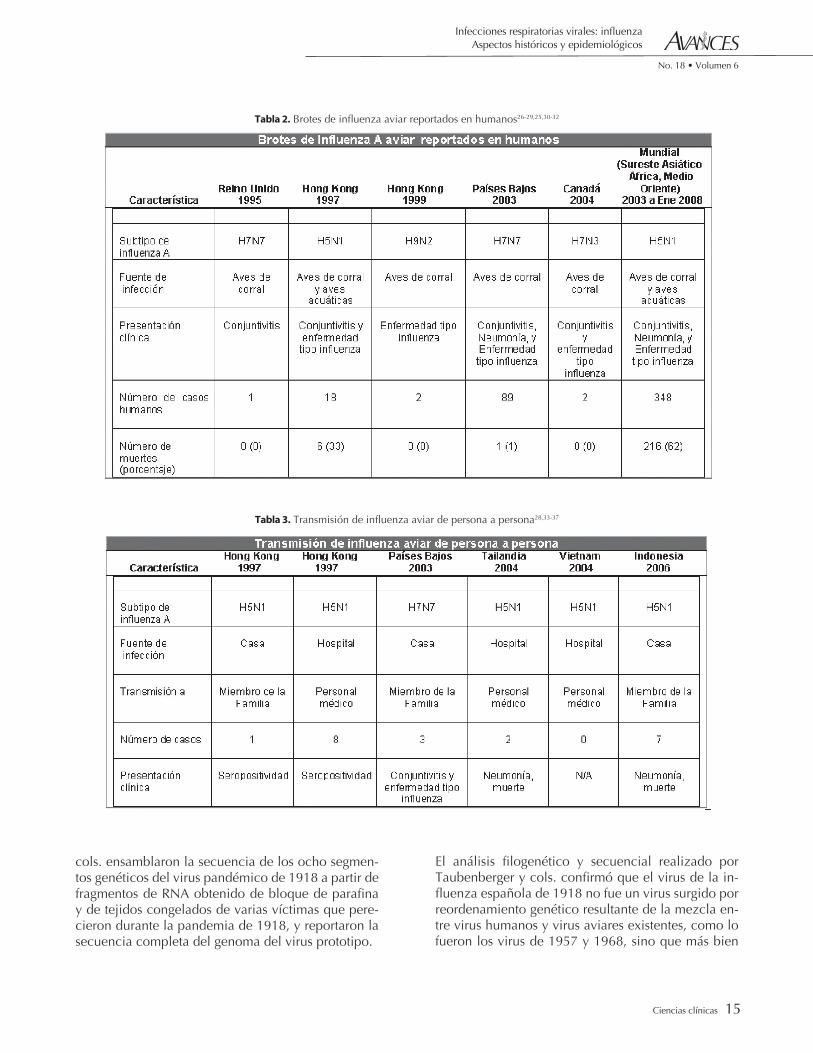
Tabla 2. Brotes de influenza aviar reportados en humanos26-29,25,30-32
Tabla 3. Transmisión de influenza aviar de persona a persona28,33-37
cols. ensamblaron la secuencia de los ocho segmen-tos genéticos del virus pandémico de 1918 a partir de fragmentos de RNA obtenido de bloque de parafina y de tejidos congelados de varias víctimas que pere-cieron durante la pandemia de 1918, y reportaron la secuencia completa del genoma del virus prototipo.
El análisis filogenético y secuencial realizado por Taubenberger y cols. confirmó que el virus de la in-fluenza española de 1918 no fue un virus surgido por reordenamiento genético resultante de la mezcla en-tre virus humanos y virus aviares existentes, como lo fueron los virus de 1957 y 1968, sino que más bien
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

16 Ciencias clínicas
No. 18 • Volumen 6
se trató de un virus aviar enteramente diferente que se adaptó a los humanos directamente a través de mutación. Esto significa que los HPAI actualmente circulando podrían evolucionar hacia virus humanos pandémicos, ya sea a través de reordenamiento ge-nético o a través de mutación directa de los genes virales (ver Figura 11).38,39
Un hallazgo inesperado fue el que la secuencia de las proteínas constituyentes del complejo de la polimera-sa (PA, PB1 y PB2) del virus de 1918 y los subsecuen-tes virus humanos difieren únicamente en 10 aminoá-cidos de la secuencia consenso del virus de influenza aviar. Muchos de estos residuos deben dar cuenta de la contribución del complejo de la polimerasa a la adquisición de la transmisibilidad humana por parte de un virus de influenza aviar. Las formas humanas de siete de estos 10 residuos de la polimerasa ya han sido observadas, individualmente o en combinación, en los virus de la influenza aviar H5N1 recuperados de aves y/o humanos. Bajo la presión selectiva de una tasa de crecimiento subóptimo en humanos, los ge-nes de la polimerasa de un virus aviar H5N1 actual-
mente circulando podrían potencialmente mutar, de tal forma que estos 10 residuos sean convertidos a sus “formas humanas”. Como resultado, el virus estaría mejor preparado para transmitirse eficientemente de humano a humano.40
El surgimiento de un virus de influenza pandémico a partir de un progenitor aviar parece también invo-lucrar un cambio hacia una unión preferencial de la HA, cambiando ésta de los ácidos siálicos α-2,3, que es la forma predominante en el tracto gastrointestinal de las aves, a los ácidos siálicos α-2,6, que es la for-ma predominante en el tracto respiratorio humano. La HA de los virus de la influenza aviar contienen en su secuencia de aminoácidos los residuos Gln226 y Gly228, los cuales forman un espacio estrecho que favorece la unión a los ácidos siálicos α-2,3. Por su parte, las especies humanas usualmente contienen en esas posiciones a los residuos Leu226 y Ser228, que forman un espacio más amplio para la unión de los ácidos siálicos α-2,6. El análisis de la estructura de la HA del virus de 1918 muestra que los residuos aviares Gln226 y Gly228 forman un espacio angosto
El virus aviar H5N1 podría adquirir el potencial para causar una pandemia de influenza en humanos mediante dos mecanismos. La coinfección de un hospedero humano con un virus aviar H5N1 y un virus de influenza humano lograría conducir a un reordenamiento genético que generaría un virus aviar H5 que podría transmitirse de humano a humano. Alternativamente, el virus aviar H5N1 podría mutar directamente y convertirse así en un virus capaz de transmitirse de humano a humano. Los virus H5N1 actualmente circulando son antigénicamente nuevos, altamente patogénicos en humanos, y se diseminan sistémicamente en modelos animales. Estos virus tienen un sitio de escisión de la HA independiente de tripsina y una deleción en el gen NS, hechos que están asociados a una elevada patogenicidad. Con una o dos mutaciones en la HA y alrededor de 10 mutaciones en el complejo de la polimerasa podrían convertirse en un virus de influenza aviar capaz de causar una pandemia en humanos. Cell 2005;123(3):368-71.
Figura 11. Origen de un virus pandémico H5N1.
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos
Fuente Aviar H5N1
Humanoinfectado
Virushumano
Viruspandémico
Reordenamiento
Mutación

Ciencias clínicas 17
No. 18 • Volumen 6
para la unión a ácidos siálicos, pero que aún permite la unión de alta afinidad por los ácidos siálicos α-2,6. Al parecer, el residuo 190 por sí solo determina la preferencia del virus de la pandemia de 1918 por los ácidos siálicos α-2,6, en contraposición a lo que sucede con los virus H5N1 que prefieren los ácidos siálicos α-2,3. Así, sólo una única mutación podría cambiar la preferencia de unión de la HA del virus aviar H5N1.40,41 Por su parte, el gen para la proteína NS está relacio-nado con la patogenicidad del virus porque interfiere con la respuesta inmunológica, dependiente de inter-ferón, por parte del hospedero contra el virus. Otro rasgo característico de alta patogenicidad es la dise-minación de la infección por el virus de la influenza a tejidos que carecen de la enzima tripsina. Los ciclos repetidos de replicación de cepas menos patogénicas del virus de la influenza está restringida al tracto res-piratorio superior, que es donde se expresa la enzima proteolítica tripsina. Las cepas altamente patogénicas del virus de la influenza tienen una HA con un sitio de escisión que contiene machos residuos de ami-noácidos básicos. Ese sitio “multibásico” es recono-cido por proteasas de las células del hospedero que se expresan más ampliamente, tales como la furina. Se piensa que la susceptibilidad de la HA para ser es-cindida es el rasgo que controla la diseminación del virus dentro de un organismo que ha sido infectado, incrementando así su patogenicidad. Como resultado de poseer una HA con un sitio multibásico, estos vi-rus pueden diseminarse a través de los pulmones y, en algunos casos, por todo el organismo.43,40
La transmisibilidad y la patogencidad del virus de la influenza son rasgos poligénicos. Y, como previa-mente se ha mencionado, la única forma de enten-der cómo es que éste fue tan altamente patogénico es reconstruir sus genes y estudiar sus propiedades. Las primeras pistas en la patogénesis del virus de 1918 se obtuvieron cuando Kobasa y cols. fueron capaces de insertar la HA y la NA del virus de 1918 en otro virus menos patogénico. Los investigadores encontraron que los virus híbridos se diseminaban más amplia-mente y causaban una mayor respuesta inflamatoria, así como hemorragia en el pulmón de los ratones al causar la liberación de grandes cantidades de cito-quinas y quimoquinas, en lo que ha sido llamado una “tormenta de citoquinas”.41,42
Sin embargo, aún no estaba claro si otros genes, como los de las proteínas no estructurales (NS) o los de la polimerasa, contribuyeron a la letalidad del virus de
1918. La única forma de contestar esta pregunta era reconstruir el virus completo. Esto es lo que Tumpey y cols. hicieron en su estudio.43 Por primera vez, un virus extinto ha sido resucitado. Ellos demostraron que el virus reconstruido mata a los ratones más rápi-damente que cualquier otro virus de la influenza pre-viamente caracterizado. Al igual que otros virus de la influenza altamente patogénicos, como el H5N1 por ejemplo, el virus de 1918 tiene una HA que es escindida en ausencia de tripsina. Sin embargo, a di-ferencia de cualquier otra HA de cualquier otro virus altamente patogénico, la HA del virus de 1918 no tiene un sitio de escisión multibásico que pueda ser escindido por la furina. En lugar de eso, su propia NA está involucrada en la escisión de la HA mediante un nuevo mecanismo aún no completamente entendido. Esto representa otro mecanismo mediante el cual un virus de la influenza de baja patogenicidad puede po-tencialmente incrementar su virulencia.43
La aparente similitud entre la génesis del virus de 1918 y el de la cepa aviar H5N1 ha causado gran preocupación a nivel mundial. Tanto el virus aviar H5N1, como el virus de la pandemia de 1918 activan su HA en forma independiente de tripsina, aunque por diferentes mecanismos. Ambos virus son alta-mente patogénicos en ratones. En el modelo de ra-tón de Tumkey, el virus de la pandemia de 1918 no se disemina más allá de los pulmones, mientras que muchos virus H5N1 se diseminan por todo el orga-nismo, incluso a través del cerebro. El único rasgo que aún le falta al virus H5N1 para convertirse en un virus pandémico es la transmisibilidad eficiente entre humanos.43
Casi tan pronto como el virus aviar H5N1 comenzó su letal diseminación por Asia, mucha gente inmedia-tamente señaló a las aves silvestres migratorias como los probables culpables de dicha diseminación. Sin embargo, estas aves comúnmente portan virus menos virulentos, de baja patogenicidad (LPAI, Low Patho-genic Avian Influenza), aunque con, virtualmente, to-das las combinaciones de HA y NA posibles.
No obstante, los científicos expertos en aves han esta-do relativamente escépticos en relación al papel que estas aves han desempeñado en la diseminación del virus. Esto es debido, en parte, a que el muestreo de decenas de miles de aves no ha logrado recolectar una sola ave viva portadora del HPAI H5N1, el cual ha causado la muerte de más de cien millones de aves domésticas y, como se dijo antes, de 256 vidas humanas hasta el 23 de marzo del presente año.
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

18 Ciencias clínicas
No. 18 • Volumen 6
No hay reservorios naturales para virus HPAI; éstos emergen sólo después de que cepas virales LPAI han pasado de las aves acuáticas hacia pollos y pavos. La evidencia sugiere que los virus HPAI H5N1 matan patos y gansos silvestres de forma tan eficiente como lo hace con pollos. “Las aves muertas no vuelan” ha sido el refrán, al tiempo que los expertos en aves se-ñalan que, además, las aves enfermas o moribundas no pueden diseminar el virus muy lejos. Asimismo, los brotes no se ajustan a ningún patrón migratorio conocido. Mas bien, los epidemiólogos investigado-res han encontrado evidencia que sugiere que el mer-cado de aves, así como otras actividades humanas son responsables de su rápida diseminación.18,19
Sin embargo, los brotes de influenza aviar que se de-tectaron en los lagos Qinghai y Erkhel, localizados en el suroeste de China y en Mongolia, respectivamente, han puesto a pensar a los científicos en el papel que las aves silvestres pueden tener en la diseminación del virus. Esos brotes no pueden ser explicados por actividades humanas. En esas regiones hay escasas aves de corral y es difícil explicar cómo llego el virus hasta esas regiones remotas así como definir si fueron las aves de corral o las aves silvestres las que llevaron el virus a esas áreas. Un grupo de veterinarios de la Wildlife Conservation Society tomaron 774 muestras tanto de aves muertas como vivas en el lago Erkhel, y se confirmó la presencia de HPAI H5N1 en las aves muertas; sin embargo, no se encontró evidencia de ese virus en ninguna de las muestras tomadas de ga-viotas, patos y gansos vivos de esa región. Aún no se sabe qué especie llevó el virus a esas zonas, pero quizá haya especies que todavía no son identificadas que podrían estar diseminando el virus a otras partes del mundo. Las implicaciones de esto son enormes, puesto que si son las aves silvestres las que portan la enfermedad, será muy difícil, si no imposible, evitar la diseminación del virus de país en país, y detener la diseminación del virus a las aves domésticas y, de ellas, hacia los humanos.18
El virus H5N1 ha afectado, por lo menos, al 84% de las especies aviarias conocidas. En su puja por anti-cipar las infecciones humanas, los gobiernos han or-denado sacrificar a millones de aves, tanto enfermas como sanas. La preocupación que ha surgido debido a la influenza aviar ha generado una amenaza para la supervivencia de varias especies raras de aves (ver Figura 12). Cuando se detectan cepas virulentas de influenza aviar en alguna región, se elimina a cada ave viva en un radio determinado.
Sin duda, esta práctica es destructiva tanto ecológi-camente como económicamente, pero ésta ha sido, hasta el momento, la mejor manera de contener la expansión del virus. La vacunación de las aves pue-de detener la expansión de la enfermedad siempre y cuando se cuenten con técnicas de seguridad y mo-nitoreo apropiadas, pero habitualmente es imprácti-co, principalmente en países en vías de desarrollo. ¿Cómo vacunar a 30 millones de aves?44
Ahora que el virus H5N1 es endémico en las aves migratorias, el potencial de una diseminación global del mismo parece inevitable. Si a esto agregamos que las cepas humanas del virus de la influenza H3N2 son endémicas en la población porcina del sureste de China, que la cepa porcina H1N1 también anda circulando en esa región, que el virus H9N2 es endé-mico en aves de corral, y que esa misma región es el origen probable del virus H5N1 actualmente circu-lante, podemos entender el por qué existe una eleva-da probabilidad de que ocurran fenómenos genéticos que pueden eventualmente conducir a una pandemia que se diseminaría alrededor del mundo en cuestión de meses, sino es que en semanas.17,18
El impacto de una nueva pandemia sería enorme. En Estados Unidos, por ejemplo, suponiendo que ocu-rriera una pandemia de la magnitud de la de 1957, se ha estimado que resultaría en 18 a 42 millones de visitas médicas, de 314,000 a 734,000 hospitali-zaciones, y de 89,000 a 207,000 muertes. Extrapo-lando estas proyecciones a la población mundial, un cálculo grueso del impacto de la pandemia resultaría en alrededor de 1 a 2 billones de casos de influenza, de 5 a 12.5 millones de casos con enfermedad seve-ra, y de 1.5 a 3.5 millones de muertes alrededor del mundo.45
Figura 12. Cygnus atratus
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

Ciencias clínicas 19
No. 18 • Volumen 6
Patrón de una epidemia
Una epidemia de influenza es un brote de la infec-ción confinado a una localidad, sea una ciudad, un pueblo o un país. Los virus de la influenza se pre-sentan a nivel mundial y causan brotes anuales de intensidad variable. Se estima que en Estados Unidos la epidemia anual causa de 25 a 50 millones de casos por año, lo que origina 150,000 hospitalizaciones y entre 30,000 y 40,000 muertes. Si estos cálculos son extrapolados al resto del mundo, la carga global pro-medio de la influenza interpandémica puede estar en el orden de un billón de casos de influenza, 3 a 5 mi-llones de casos de enfermedad severa, y de 300,000 a 500,000 muertes cada año.46
En una comunidad dada, las epidemias causadas por el virus de la influenza tipo A tienen un patrón carac-terístico. En la Figura 13 se muestra la pauta típica de una epidemia de influenza tipo A en una comunidad urbana. Se trata de la epidemia ocurrida en Houston, Texas, Estados Unidos, en 1976, causada por el virus A/Victoria/75 (H3N2).47
Las epidemias de influenza suelen comenzar abrup-tamente, alcanzan su pico en dos a tres semanas, y duran de cinco a seis semanas. En las fases iniciales de la epidemia, la infección y la enfermedad apare-cen en la población escolar, lo que se refleja por el absentismo a clases, el aumento de las visitas médi-cas y el incremento de las admisiones en hospitales pediátricos. Los niños afectados llevan el virus al ho-gar, donde adquieren la infección sus hermanos pe-queños y los adultos. La infección y la enfermedad en las personas adultas se traduce en mayor absentismo laboral, más admisiones en los hospitales generales por diagnósticos tales como neumonía, exacerbación de enfermedad pulmonar crónica, laringotraquei-tis, insuficiencia cardiaca congestiva, y aumento de la mortalidad por influenza o neumonía. En lo que concierne al virus tipo B, éste presenta una pauta de aparición semejante, aunque no se advierte un incre-mento de la mortalidad.
Si bien la duración de la epidemia suele ser de cinco a seis semanas, el virus está presente en la comuni-dad durante un número variable de semanas antes y después del periodo epidémico. Durante la fase epi-démica, la duración de un brote en cada subgrupo de la población puede ser limitada, esto es, no mayor de dos semanas en un grupo cerrado.
En un brote típico, como el mostrado en la Figura 13, el número de aislamientos se incrementa rápida-mente y alcanza un pico a las dos o tres semanas, lo que se correlaciona con el aumento en el número de casos tipo influenza. Luego, el número de aisla-mientos disminuye rápidamente en las dos a tres se-manas siguientes, y sólo escasos aislamientos pueden obtenerse en las semanas siguientes. Después, miste-riosamente, los virus de la influenza casi nunca son nuevamente aislados durante el resto del año.
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos
Figura 13. Correlación de los índices no virológicos de la epidemia y el número de aislamientos del virus a/Victoria
cada semana, Houston, Texas, Estados Unidos, 1976.
1 2 3 4 5 6 7 8 9 10
Enero Febrero Marzo
Númerode semanas
100
200
300
10
10
20
30
40
175
400
425
450
475
500
10
15
20
25
30
5
15
25
35
45
4
6
8
10
12
35
20
Virus de la gripeA/Victoria
Mortalidadgripe-neumonía
Núm
ero
de a
isla
mie
ntos
Núm
ero
Núm
ero
Med
ia d
iaria
Porc
enta
jede
aus
entis
mo
Admisiones de neumonía
Pacientesadultos
Pacientespediátricos
Frecuenciade las alteracionesrespiratorias
Exámenes
Vacaciones
Absentismo escolar
Absentismo industrial
Media
Admisionesen los serviciosde urgencias
Frec
uenc
ia d
e la
sal
tera
cion
es re
spira
t.Fr
ecue
ncia
de
las
alte
raci
ones
resp
irat.
20 Ciencias clínicas
No. 18 • Volumen 6
Los brotes de influenza ocurren en diferentes patrones estacionales, dependiendo de la región del mundo de la que estemos hablando. En los países tropicales, los virus de la influenza pueden ser aislados durante todo el año, pero las epidemias suelen estar en rela-ción con los cambios en los patrones climáticos. Sin embargo, en los climas templados, sólo escasos ais-lamientos de influenza pueden ser obtenidos en las semanas previas al brote epidémico.
Las epidemias de influenza ocurren casi exclusiva-mente en los meses de invierno (diciembre a abril) en el hemisferio norte (ver Tabla 4), y sólo en raras ocasiones ocurren infecciones en otras épocas el año. Si una nueva cepa viral tiene un mapa antigénico significativamente diferente al del virus que circuló previamente, entonces puede encontrarse brotes tan tempranos como, por ejemplo, durante el mes de oc-tubre. Así mismo, hay ocasiones en que el pico del brote de influenza ocurre tardíamente, hasta abril o mayo.45 En el hemisferio sur, los brotes ocurren en los meses de mayo a septiembre, y pueden predecir el tipo de virus que se encontrará en el hemisferio norte durante el próximo invierno.
Dónde o cómo persiste el virus en la comunidad no se conoce. Debe existir una cadena de transmisión continua persona a persona para que el virus sobre-viva. Puede detectarse cierta actividad viral en gran-des centros de población durante todo el año, lo cual indica que el virus permanece endémico en la po-blación y produce algunas infecciones subclínicas o menores.48
Las tasas de ataque son altamente variables de brote a brote. Durante las epidemias, la tasa de ataque pro-medio es de alrededor del 20%, pero no es inusual que en algunos grupos seleccionados pueda ser muy alta, alcanzando valores que van del 40% al 50%.48
La mortalidad producida por la influenza es un índi-ce fidedigno del alcance de la epidemia, pero tien-de a minimizar la magnitud de las epidemias por el virus tipo B, que generalmente son menos extensas, producen una enfermedad de menor severidad, y en ellas no suele haber un incremento de la mortalidad. Otra característica importante de la actividad de la influenza estacional es que conduce a un “exceso” de muertes, o “muertes ocultas”. Éstas son muertes que no son usualmente atribuidas a la influenza, aun-que muchas de ellas son causadas por neumonía viral o bacteriana, secundaria a la infección por el virus de la influenza. Tomemos, por ejemplo, los caos de In-glaterra y los países bajos. En Inglaterra se ha estima-do que anualmente ocurrieron entre 5,000 y 29,000 muertes en exceso, causadas por influenza, entre los años 1975 y 1990, lo que representa un número cin-co veces mayor que el número de muertes atribuido a influenza en los certificados de defunción.49 En los países bajos, unas 2,000 muertes anuales son atribui-das a la influenza (1.3 muertes por cada 10,000 ha-bitantes); sin embargo, ocurren otras 2.6 muertes por 10,000 habitantes que, aunque no son atribuidas a la influenza, están directamente relacionadas con la infección inicial de influenza.50
La experiencia en cualquier año determinado refleja el interjuego entre la amplitud de la desviación anti-génica del virus predominante y la disminución de la inmunidad de la población. Los tres tipos de virus de la influenza muestran desviaciones antigénicas. Sin em-bargo, sólo el tipo A muestra cambios antigénicos, lo cual, presuntamente, se debe a que los virus tipo B y C restringen más su variedad de hospederos. EI cambio antigénico aparece en una región geográfica limitada y, con el tiempo el nuevo virus se extiende para todo el mundo por las rutas de transporte. Los virus de la in-fluenza no parecen ser objeto de cambios antigénicos en las aves, quizá debido a su corta vida.
Tabla 4. Mes de actividad pico de influenza en Estados Unidos durante 31 temporadas
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

Ciencias clínicas 21
No. 18 • Volumen 6
En países de gran extensión, por ejemplo Estados Uni-dos o Australia, pueden verse diferencias regionales en relación al tiempo en que ocurren los brotes e in-fluenza. No es raro tener grandes brotes de influenza en algunas regiones, mientras que otras no muestran en lo absoluto actividad de influenza. A menudo esas comunidades que inicialmente son respetadas tiene sus brotes más tardíamente, sobre todo si el virus que ha estado circulando muestra variación antigénica significativa en relación a los virus previamente cir-culantes.
Por muchos años, se pensó que durante una epidemia de influenza sólo una cepa particular del virus preva-lecía y que los otros virus respiratorios disminuían o incluso desaparecían de la comunidad. Actualmente, sabemos que dos cepas diferentes del mismo subtipo (por ejemplo, A/Victoria/3/75/H3N2 y A/Texas/1/77/H3N2) o que dos subtipos diferentes de influenza tipo A (H1N1 y H3N2) pueden circular simultáneamente. Mas aun, se han visto brotes simultáneos de influenza A y B, e incluso brotes de influenza A y virus sinci-tial respiratorio (RSV) han sido documentados. Ha-cia finales de la temporada de influenza 1987-1988, tres virus diferentes circularon: A/Sichuan/87/H3N2, A/Taiwan/85/H1N1, y B/Ann Harbor/86. En algunas ciudades, los tres virus pudieron ser recuperados en un mismo mes; por ejemplo, A/Taiwan/85/H1N1 apareció en una universidad, A/Sichuan/87/H3N2 causó brotes en un asilo, y B/Ann Harbor/86 estaba ocurriendo en otro asilo y en los pacientes ambulato-rios vistos en una sala de emergencias, todos simul-táneamente.48
En cuanto al virus de la influenza tipo C, a pesar de haber sido caracterizado desde 1947, ha habido po-cos reportes en relación a sus hallazgos clínicos y epidemiológicos. Esto ha sido debido, no a su leve patogenicidad, sino más bien a las dificultades para su aislamiento.51 Por mucho tiempo se pensó que el virus tipo C infectaba sólo a humanos; sin embargo, en 1983 se reportó el aislamiento de este virus en población porcina, e incluso existe evidencia que la transmisión interespecies entre humanos y cerdos ha ocurrido en la naturaleza.52,53
La infección por el virus de la influenza tipo C ocurre principalmente con un patrón epidemiológico espo-rádico, o bien, como brotes de enfermedad de magni-tud limitada.54 Sin embargo, epidemias de extensión nacional han sido reportadas en Japón. La mayoría de estas infecciones ocurre en los meses de enero a ju-lio, un periodo de tiempo que se extiende más allá de
la temporada de influenza convencional, y coexiste con los brotes epidémicos de influenza A y B.55
El virus de la influenza tipo C afecta sobre todos a ni-ños menores de 6 años de edad y a adultos jóvenes.54 En estudios seroepidemiológicos llevados a cabo en Francia se ha demostrado que del 61% al 70% de la población se ha expuesto previamente al virus, se han encontrado las tasas más altas de seropositividad entre las personas de 16 a 30 años de edad.56 Cifras similares han sido reportadas en estudios realizados en España57 y Brasil.58
La diferenciación clínica entre los diferentes subtipos de influenza es muy difícil, se requieren técnicas de diagnóstico más sofisticadas, tales como cultivo viral o RT-PCR para identificar los casos específicos.59 Ha-bitualmente, la infección por el virus de la influenza tipo C suele ser de severidad leve. Pero, el virus de la influenza tipo C no se limita sólo a causar enfer-medad de las vías respiratorias superiores; éste pue-de también causar enfermedad de vías respiratorias bajas de severidad suficiente como para requerir hospitalización, tales como bronquitis, bronquiolitis y neumonía.60,61 Cerca del 20% de los niños infec-tados por virus de la influenza tipo C han requerido hospitalización, y de éstos, más del 70% presentan afección de vías respiratorias bajas.51
La reinfección por el virus de la influenza tipo C es un fenómeno que puede ocurrir frecuentemente, debido a que la infección no induce una inmunidad suficien-temente protectora o duradera.59 Los estudios gené-ticos realizados sugieren que la epidemiología del virus de la influenza tipo C puede caracterizarse por la presencia de muchas variantes cocirculando en un momento dado. Se han descrito seis linajes diferentes de este virus; de esta manera, la infección mixta por virus de la influenza tipo C pertenecientes a diferen-tes linajes daría lugar a frecuentes reordenamientos genéticos en la naturaleza, lo que resultaría en va-riantes virales que, de hecho, ya han ocasionado el surgimiento de brotes epidémicos.
La importancia de los virus de la influenza tipo C no ha sido bien valorada. La realización de futuros estu-dios clínicos y epidemiológicos ayudarán a darle su apropiado lugar a estos virus.54,60,62,63
Impacto socio-económico de la influenza
En 1947, en respuesta al reconocimiento de la va-riación antigénica del virus de la influenza, la OMS
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

22 Ciencias clínicas
No. 18 • Volumen 6
estableció un programa de vigilancia epidemiológica de influenza que opera a través de una red nacional y regional de centros para monitorear la actividad glo-bal de influenza. Los principales centros coordinado-res están localizados en Londres, Melbourne, Atlanta y Tokyo, junto con otros 110 centros nacionales en 83 países alrededor del mundo.49 La vigilancia para los brotes de influenza es más amplia que para cual-quier otra enfermedad, con objeto de identificar la aparición temprana de nuevas cepas y preparar vacu-nas contra ellas antes de que se presente una epide-mia. Esta vigilancia puede extenderse a la población de animales, en especial de aves, cerdos y equinos.
El impacto económico de los virus de influenza tipo A es significativo debido a la morbilidad que acom-paña a las infecciones. Los costos económicos han sido estimados en 10 a 60 millones de dólares por mi-llón de habitantes en países industrializados, lo que depende de la magnitud de la epidemia.
El análisis económico de la enfermedades puede ser separado en dos fases: una fase descriptiva, en la cual se estima la carga o impacto de la enfermedad; y una fase analítica, en la cual se evalúan las intervenciones y sus costos, así como las consecuencias de “remo-ver” la carga de una enfermedad. El costo de cual-quier enfermedad es la suma de los costos directos, los indirectos y los intangibles. Los costos directos re-sultan del uso de los recursos médicos y no médicos; para la influenza, éstos incluyen los costos asociados con el uso de los recursos para el cuidado de la sa-lud y aquéllos que resultan de los gastos relacionados que efectúan tanto la comunidad como la persona. Los costos indirectos son aquéllos relacionados con pérdidas en la productibilidad y el ausentismo en tra-bajos y escuelas. Los costos intangibles se relacionan a los efectos en la calidad de vida y en el desempeño personal. La suma de estos costos individuales varía dependiendo de la edad y el estado de salud subya-cente de la población de pacientes bajo considera-ción.50
Los costos directos relacionados a hospitalización y tratamiento médico son, obviamente, más grandes para los grupos de pacientes de alto riesgo. En térmi-nos generales, las tasas más altas de hospitalización ocurren entre los infantes menores de 1 año de edad, y entre las personas de la tercera edad, es la neu-monía la causa más frecuente de hospitalización. De hecho, alrededor del 90% de las muertes reconocidas como asociadas a influenza ocurren en personas ma-
yores de 65 años de edad. Los costos directos están relacionados también con la presencia de enfermeda-des crónicas, las cuales incrementan tanto el tiempo de hospitalización como la tasa de muerte asociada a influenza.50 Por ejemplo, las enfermedades cardiovas-culares y pulmonares, la diabetes mellitus o el asma pueden incrementar considerablemente la hospitali-zación. Asimismo, la tasa de mortalidad en cualquier grupo de edad está relacionado con la presencia o ausencia de enfermedades crónicas pre-existentes; se ha reportado que la tasa de mortalidad por influenza es 50 veces mayor en pacientes con enfermedad car-diovascular, más de cien veces mayor en pacientes con enfermedad pulmonar, y aún mayor en pacientes con dos o más condiciones mórbidas, que en adultos sanos.47-50
La infección por VIH (virus de la inmunodeficiencia humana) es otra condición crónica que debemos te-ner en cuenta. Hay poca información disponible en relación a la frecuencia y severidad de la influenza en personas con infección por VIH. Un estudio re-trospectivo realizado entre mujeres jóvenes y de me-diana edad del programa Medicaid en el estado de Tennessee, Estados Unidos, determinó que el riesgo de hospitalizaciones por problemas cardiopulmona-res en mujeres con infección por VIH era más alto durante la temporada de influenza que en cualquier otra época del año. El riesgo de hospitalización fue más grande en mujeres infectadas por VIH que en mujeres con otras condiciones médicas subyacentes. En otro estudio se estimó que el riesgo de muerte re-lacionado con influenza era de 94 a 146 por cien mil personas con SIDA (síndrome de inmunodeficiencia adquirida), lo cual es muy grande si lo comparamos con el riesgo de 0.9 a 1.0 por cada cien mil personas de 25 a 54 años de edad sin SIDA, y con el 64-70 por cada cien mil personas de edad mayor de 65 años. Así pues, en personas infectadas con VIH los sínto-mas de la influenza pueden ser más prolongados y el riesgo de muerte se incrementa.64,65 Los costos indirectos pueden llegar a ser de hasta el 80% al 90% de los costos totales y se originan prin-cipalmente del ausentismo y pérdida de la producti-vidad. Asimismo, hasta el 10% del total de ausencias por enfermedad en el trabajo se deben a influenza. El ausentismo a la escuela es también importante, y cuando los niños están enfermos, los padres a me-nudo faltan a su trabajo para cuidarlos. Todos estos factores contribuyen a los altos costos indirectos de la influenza.50
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

Ciencias clínicas 23
No. 18 • Volumen 6
Los costos intangibles se derivan de la disminución en el estatus funcional de los pacientes y de la dis-minución en la calidad de vida tanto de los pacien-tes como de sus familiares o de las personas que los cuidan. Asimismo, la alteración del estatus funcional puede disminuir la capacidad del paciente para reac-cionar ante un estímulo y esto puede tener efectos ad-versos para la salud y la seguridad en el trabajo; se ha demostrado que incluso un cuadro leve de influenza puede disminuir los tiempos de reacción en un 20% al 40%. Los efectos adversos generales sobre la cali-dad de vida, con pérdida de tiempo para el esparci-miento y la inhabilidad para realizar las actividades diarias normales, son también factores importantes que no deberían ser subestimados.50
En Estados Unidos por causa de la influenza se gas-tan alrededor de 1 a 3 billones de dólares en costos médicos directos cada año; los costos indirectos, in-cluyendo la pérdida de salarios por enfermedad y la pérdida de futuros ingresos económicos por muerte, son mucho más altos, del orden de 10 a 15 billones de dólares por año.44 Números similarmente altos han sido obtenidos de otros estudios realizados en Francia y Alemania.50
SI bien es cierto que, en general, se considera a la influenza como un problema de salud relativamente benigno, ésta es una enfermedad grave que mata a miles y miles de personas anualmente. Por su parte, la epidemia de gripe aviar presente en Asia es una crisis de importancia mundial. Ya hemos revisado el riesgo potencial para la humanidad de otra gran pandemia de influenza relacionada con el virus de influenza A H5N1. No se le debe mirar con descui-do, sólo por el hecho de que un país esté conside-rado “libre” de la influenza aviar, lo que ha estado ocurriendo en países de Asia, Europa y África, con el argumento equívoco de que éste es un problema sanitario propio de esos países. El término de “libres” o “exentos” es posible que se pierda en el momento menos pensado. El virus H5N1 de la influenza aviar debe considerarse como una seria amenaza a nivel mundial que requiere de una permanente y sólida vi-gilancia para controlarlo y evitar su difusión a otros continentes. Por ello, todas las autoridades sanitarias del mundo hacen un constante llamado de alerta a todos los gobiernos del planeta para que presten su colaboración con el fin de evitar que la enfermedad se extienda y se pueda, en esta forma, disminuir el riesgo de una posible pandemia a muy corto plazo y/o amortiguar sus efectos.66-70
Referencias bibliográficas:
1. Potter CW. A history of influenza. J Appl Microbiol 2001;91(4):572-579.2. García-García Juan, Ramos C. La influenza, un problema vi-gente de salud pública. Salud Pública, Méx. 2006;48(3):244-267. Disponible en: http://scielo.unam.mx/scielo.php?script=sci_arttext&pid=S0036-36342006000300009&lng=es&nrm=iso.3. Smith W, Andrewes CH, Laidlaw PP. A virus obtained from in-fluenza patients. Lancet 1933; i: 66-68.4. Francis T. A new type of virus from epidemic influenza. Science 1940;92405-408.5. Taylor RM. A further note on 1233 influenza C virus. Archives of Virology 1951;4(4):485-500. 6. Hirst GK. The agglutination of red cells by allantoic fluid of chicks embryos infected with influenza virus. Science. 1941;94(2427):22-23.7. Potter CW. Influenza. In: Zuckerman AJ, Banatvala JE, Pattison JR, Griffiths PD, Schoub BD. Principles and practice of clinical virology, 5th edition. John Wiley & Sons, LTD 2004:271-297.8. Palese P. Influenza: old and new threats. Nat Med. 2004;10(12 Suppl):S82-7. 9. Glezen WP. Emerging infections: pandemic influenza. Epide-miol. Rev. 1996;18(1):64-76. 10. Kawaoka Y, Yamnikova S, Chambers TM, Lvov DK, Webster RG. Molecular characterization of a new hemaglutinin subtype H14 of influenza virus A. Virology 1990;179:759-767.11. Röhm C, Zhou N, Suss J, Mackenzie J, Webster RG. Characte-rization of a novel influenza hemagglutinin, H15. Criteria for de-termination of influenza subtypes. Virology 1996;217:508-516.12. Fouchier RA, Munster V, Wallensten A, Bestebroer TM, Herfst S, Smith DR et al. Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black-headed gulls. J Virol 2005;79:2814-2822.13. Webster RG, Bean WJ, Gorman OT, Chambers TM, Kawaoka Y. Evolution and ecology of influenza A viruses. Microbiol Rev 1992;56(1):152-79. 14. Webster RG. Influenza: An Emerging Disease. Emerg Infect Dis 1998;4(3):436-41.15. Crawford PC, Dubovi EJ, Castleman WL, Stephenson I, Gibbs EP, Chen L, Smith C, Hill RC, Ferro P, Pompey J, Bright RA, Medi-na MJ, Johnson CM, Olsen CW, Cox NJ, Klimov AI, Katz JM, Do-nis RO. Transmission of equine influenza virus to dogs. Science. 2005;310(5747):482-5. 16. Suarez DL. Evolution of avian influenza viruses. Vet Microbiol 2000;74(1-2):15-27. 17. Reina J, Ortiz de Lejarazu R. Mecanismos de patogenicidad y adaptabilidad humana de las cepas gripales aviarias A (H5N1). Rev Esp Quimioterap 2005;18(4):273-280.18. Normile D. Are Wild Birds to Blame? Science 2005;310(5747):426-8.19. Lovell C. Are migratory wild birds being unfairly blamed? Poultry world 2007;161(1):26-7.20. Peiris M, Yuen KY, Leung CW, et al. Human infection with influenza H9N2. Lancet 1999;354:916-17.21. Horimoto T, Rivera E, Pearson J, Senne D, Krauss S, Kawaoka Y, Webster RG. Origin and molecular changes associated with emergence of a highly pathogenic H5N2 influenza virus in Mexi-co. Virology. 1995;213(1):223-30.22. Subbarao K, Klimov A, Katz J, Regnery H, Lim W, Hall H, Per-due M, Swayne D, Bender C, Huang J, Hemphill M. Rowe T, Shaw M, Xu X, Fukuda K, Cox N. Characterization of an Avian influenza A (H5N1) virus isolated from a child with a fatal respiratory illness. Science 1998;279:393-6.
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

24 Ciencias clínicas
No. 18 • Volumen 6
23. World Health Organization. Avian influenza Fac. sheet. Wee-kly epidemiological record 2006;81(14):129-36. 24. Peiris JS, Yu WC, Leung CW, Cheung CY, Ng WF, Nicholls JM, Ng TK, Chan KH, Lai ST, Lim WL, et al.: Re-emergence of fatal human influenza A subtype H5N1 disease. Lancet 2004;363: 617-619.25. World Health Organization: Cumulative Number of Confirmed Human Cases of Avian Influenza A/(H5N1) Reported to WHO. http://www.who.int/csr/disease/avian_influenza/country/cases_ta-ble_2009_01_19/en/index.html26. Kurtz J, Manvell RJ, Banks J: Avian influenza virus isolated from a woman with conjunctivitis. Lancet 1996;348:901-902.27. Peiris M, Yuen KY, Leung CW, Chan KH, Ip PL, Lai RW, Orr WK, Shortridge KF: Human infection with influenza H9N2. Lancet 1999;354:916-917.28. Koopmans M, Wilbrink B, Conyn M, Natrop G, van der Nat H, Vennema H, Meijer A, van Steenbergen J, Fouchier R, Osterhaus A, Bosman A: Transmission of H7N7 avian influenza A virus to human beings during a large outbreak in commercial poultry farms in the Netherlands. Lancet 2004;363:587-593.29. Yuen KY, Chan PK, Peiris M, Tsang DN, Que TL, Shortridge KF, Cheung PT, To WK, Ho ET, Sung R, Cheng AF: Clinical fea-tures and rapid viral diagnosis of human disease associated with avian influenza A H5N1 virus. Lancet 1998;351:467-471.30. Tran TH, Nguyen TL, Nguyen TD, Luong TS, Pham PM, Nguyen VC, Pham TS, Vo CD, Le TQ, Ngo TT, et al.: Avian influenza A (H5N1) in 10 patients in Vietnam. N Engl J Med 2004;350:1179-1188.31. Chotpitayasunondh T, Ungchusak K, Hanshaoworakul W, Chunsuthiwat S, Sawanpanyalert P, Kijphati R, Lochindarat S, Srisan P, Suwan P, Osotthanakorn Y, et al.: Human disease from influenza A (H5N1), Thailand, 2004. Emerg Infect Dis 2005;11: 201-209.32. de Jong MD, Bach VC, Phan TQ, Vo MH, Tran TT, Nguyen BH, Beld M, Le TP, Truong HK, Nguyen VV, et al.: Fatal avian influenza A (H5N1) in a child presenting with diarrhea followed by coma. N Engl J Med 2005;352:686-691.33. Katz JM, Lim W, Bridges CB, Rowe T, Hu-Primmer J, Lu X, Abernathy RA, Clarke M, Conn L, Kwong H, et al.: Antibody response in individuals infected with avian influenza A (H5N1) viruses and detection of anti-H5 antibody among household and social contacts. J Infect Dis 1999, 180:1763-1770.34. Buxton Bridges C, Katz JM, Seto WH, Chan PK, Tsang D, Ho W, Mak KH, Lim W, Tam JS, Clarke M, et al.: Risk of influenza A (H5N1) infection among health care workers exposed to patients with in-fluenza A (H5N1), Hong Kong. J Infect Dis 2000, 181:344-348.35. Ungchusak K, Auewarakul P, Dowell SF, Kitphati R, Auwanit W, Puthavathana P, Uiprasertkul M, Boonnak K, Pittayawonganon C, Cox NJ, et al.: Probable person-to-person transmission of avian influenza A (H5N1). N Engl J Med 2005, 352:333-340.36. Liem NT, Lim W, World Health Organization International Avian Influenza Investigation Team, Vietnam: Lack of H5N1 avian influenza transmission to hospital employees, Hanoi, 2004. Emerg Infect Dis 2005, 11:210-215.37. Avian Influenza, human (09): Indonesia, Archive number 20070111.0133 [www.promedmail.org/pls/promed].38. Taubenberger JK, Reid AH, Krafft AE et al. Initial genetic characterisation of the 1918 Spanish influenza virus. Science 1997;275:1793-1796.39. Taubenberger JK, Reid AH, Lourens RM, Wang R, Jin G, Fa-nning TG. Characterization of the 1918 influenza virus polymera-se genes. Nature 2005;437,889-893.40. Russell CJ, Webster RG. The genesis of a pandemic influenza virus. Cell 2005;123(3):368-71.
41. Lamb RA, Jackson D. Extinct 1918 virus comes alive. Nat Med 2005;11(11):1154-6.42. Kobasa D, Takada A, Shinya K, Hatta M, Halfmann P, The-riault S, Suzuki H, Nishimura H, Mitamura K, Sugaya N, Usui T, Murata T, Maeda Y, Watanabe S, Suresh M, Suzuki T, Suzuki Y, Feldmann H, Kawaoka Y. Enhanced virulence of influenza A viru-ses with the haemagglutinin of the 1918 pandemic virus. Nature 2004;431(7009):703-7.43. Tumpey TM, Basler CF, Aguilar PV, Zeng H, Solorzano A, Swayne DE, Cox NJ, Katz JM, Taubenberger JK, Palese, P, Garcia-Sastre A. (2005). Science 2005; 310:77-80.44. Waltz E. Pandemic prevention schemes threaten diversity, ex-perts warn. Nat Med 2006;12(6):598.45. Prevention and Control of Influenza . Recommendations of the Advisory Committee on Immunization Practices (ACIP), 2007. MMWR 2007;56(RR06) :1.54. http://www.cdc.gov/mmwr/pre-view/mmwrhtml/rr5606a1.htm46. World Health Organization. Acute respiratory infections. In-fluenza. Disponible en: http://www.who.int/vaccine_research/di-seases/ari/en/47. Glezen WP, Couch RB. Interpandemic influenza in the Hous-ton area, 1974-76. N Eng J Med 1978;298(11):587-92.48. Prevention and Control of Influenza. Recommendations of the Advisory Committee on Immunization Practices (ACIP), 2007. MMWR 2007;56(RR06) :1-54. http://www.cdc.gov/mmwr/pre-view/mmwrhtml/rr5606a1.htm49. Betts RF. Influenza Virus. In: Mandell, Douglas and Bennett’s Principles and Practice of Infectious Diseases, 4th edition, Chur-chill Livingstone Inc 1995:1546-67.50. Zambon MC. Epidemiology and pathogenesis of influenza. Journal of Antimicrobial Chemotherapy 1999; 44: 3-9.51. Calvo C, García-García ML, Centeno M, Pérez-Breña P, Casas I. Influenza C virus infection in children, Spain. Emerg Infect Dis 2006;12(10):1621-2.52. Lin JC, Nichol KL. Excess mortality due to pneumonia or in-fluenza during influenza seasons among persons with acquired immunodeficiency syndrome. Arch Intern Med 2001;161:441-6.53. Guo YJ, Jin FG, Wang P, Wang M, Zhu JM. Isolation of in-fluenza C virus from pigs and experimental infection of pigs with influenza C virus. J Gen Virol 1983;64 (Pt 1):177-82.54. Kimuraa H, Abikoa C, Penga G, Murakia Y, Sugawaraa K, Hongoa S, Kitameb F, Mizutac K, Numazakic Y, Suzukic H, Nakamuraa K. Interspecies transmission of influenza C virus be-tween humans and pigs. Virus Research 1997;48(1):71-79.55. Matsuzaki Y, Sugawara K, Mizuta K, Tsuchiya E, Muraki Y, Hongo S, Suzuki H, Nakamura K. Antigenic and genetic cha-racterization of influenza C viruses which caused two outbreaks in Yamagata City, Japan, in 1996 and 1998. J Clin Microbiol 2002;40(2):422-429.56. Matsuzaki Y, Katsushima N, Nagai Y, Shoji M, Itagaki T, Sakamo-to M, Kitaoka S, Mizuta K, Nishimura H. Clinical features of influen-za C virus infection in children. J Infect Dis 2006;193(9):1229-35. 57. Manuguerra JC, Hannoun C, Aymard M. Influenza C virus in-fection in France. J Infect 1992;24:91-9.58. Manuguerra JC, Hannoun C, Saenz MC, Villar E, Cabezas JA. Sero-epidemiological survey of influenza C virus in Spain. Euro-pean Journal of Epidemiology 1994;10:91-4.59. Greenbaum E, Morag A, Zakay-Rones Z. Isolation of Influenza C Virus during an Outbreak of Influenza A and B Viruses. J Clin Microbiol 1998;36(5):1441-1442.60. Matsuzaki Y, Abiko C, Mizuta K, Sugawara K, Takashita E, Mu-raki Y, Suzuki H, Mikawa M, Shimada S, Sato K, Kuzuya M, Takao S, Wakatsuki K, Itagaki T, Hongo S, Nishimura H. A nationwide
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos

Ciencias clínicas 25
No. 18 • Volumen 6
epidemic of influenza C virus infection in Japan in 2004. J Clin Microbiol 2007;45(3):783-8.61. Matsuzaki Y, Mizuta K, Sugawara K, Tsuchiya E, Muraki Y, Hongo S, Suzuki H, Nishimura H. Frequent reassortment among influenza C viruses. J Virol 2003;77(2):871-81.62. Motta FC, Luiz MO, Couceiro JNSS. Serological analysis re-veals circulation of influenza C viruses, Brazil. Rev Saúde Pública 1000;34(2):204-5.63. Peng G, Hongo S, Kimura H, Muraki Y, Sugawara K, Kitame F, Numazaki Y, Suzuki H, Nakamura K. Frequent occurrence of genetic reassortment between influenza C virus strains in nature. J Gen Virol 1996;77 (Pt 7):1489-92.64. Szucs T. The socio-economic burden of influenza. Journal of Antimicrobial Chemotherapy 1999;44:11-45.65. Neuzil KM, Reed GW, Mitchel EF, Griffin MR. Influenza-asso-ciated morbidity and mortality in young and middle-aged Women. JAMA 1999;281(10):901-7.66. World POPClock Projection. http://www.census.gov/ipc/www/popclockworld.html67. Osterholm MT. En previsión de la próxima pandemia. Salud Pública de México 2006;48(3):279-85.68. Ramos C. La influenza, una oportunidad para la prevención y el control. Salud Pública de México 2005;47(6):294-5.69. Mateo P. La influenza aviar podría matar 150 millones de per-sonas. V Congreso de Avicultura que se celebró en La Habana del 11 al 13 de marzo de 2006. http://www.iia.cu/congreso/pilar.htm70. Navarro D. OMS: Una pandemia de gripe aviar podría matar a 150 millones. http://axxon.com.ar/not/155/c-1550006.htm
Correspondencia:Dr. Adrián Valle de la O.Email: [email protected]
Infecciones respiratorias virales: influenzaAspectos históricos y epidemiológicos