information to users - mcgill universitydigitool.library.mcgill.ca/thesisfile21519.pdf · ckb...
TRANSCRIPT

INFORMATION TO USERS
This manuscript MS been reproduœcl fn:)m the microfilm master. UMI films
the text diredly from the original or copy submitted. Thus, some thesis and
dissertation copies are in typewriter face, while others may be from any type of
computer printer.
The quality of this reproduction is dependent upon the quality of the
copy submitted. Broken or inetistinct prim, coIored or poor quality illustrations
and photographs, print bleedthrough, substandard margins, anet improper
alignment can adversely affect reproduction.
ln the unlikely event that the author did nct send UMI a complete manuscript
and there are missing pages, these will be noted. Also, if unauthorized
copyright material had to be removec:t, a note will indicate the deletion.
Oversize materials (e.g., maps, drawings, d'larts) are reproduced by
sectioning the original, begiming at the upper Ieft..hand corner and continuing
trom left to right in equal sections with small overlaps.
Photographs includecl in the original manuscript have been reproduœd
xerographically in this copy. Higher quality 6- x 9" black and white
photographie prints are available for any photographs or illustrations appearing
in this copy for an additional charge. Contact UMI directly to order.
BeU & Howell Information and Leaming300 North Zeeb Raad, Ann Arbor, MI 48106-1346 USA
800-521.Q600


•
•
•
Evidence for a receptor bindiDg 24R,2S-dihydroxyvitamin D3 indeveloping bone
by
ALYSONBYRD
Department of Surgery, Division of Experimental Surgery
MeGUI University, Montreal
March 1999
A thesis submitted to the Faeulty of Graduate Studies and Research in partialfulfUment of the requiremenfs of the degree of
MASTER OF SCIENCE
e ALYSON BYRD, 1999

1+1 National Libraryof Canada
Acquisitions andBibliographie Services
395 Wellington StreetOttawa ON K1A 0N4canada
Bibliothèque nationaledu Canada
Acquisitions etservices bibliographiques
395. rue WellingtonOttawa ON K1 A 0N4canada
The author bas granted a nonexclusive licence allowing theNational Library of Canada toreproduce, loan, distribute or seOcopies of this thesis in microform,paper or electronic formats.
The author retains ownership of thecopyright in this thesis. Neither thethesis oor substantial extracts from itmay be printed or otherwisereproduced without the author' spermISSIon.
L'auteur a accordé une licence nonexclusive permettant à laBibliothèque nationale du Canada dereproduire, prêter, distribuer ouvendre des copies de cette thèse sousla forme de microfiche/film, dereproduction sur papier ou sur formatélectronique.
L'auteur conserve la propriété dudroit d' auteur qui protège cette thèse.Ni la thèse ni des extraits substantielsde celle-ci ne doivent être imprimésou autrement reproduits sans sonautorisation.
0-612-50730-0
Canadl

•
•
•
TABLE OF CONTENTS
Sections Page
LIST OF FIGURES 4
~<:~O~IJH(J~S 5
ABBREVIATIONS 7
1. ~STRAcr 10
II. RÉsUMÉ... .. ... . .. . .... . ... .. .. . .. . .. .. .. Il
ID. INTRODUCTION 13
A. The Vitamin D Endocrine System 13
B. Bone Histogenesis 16
C. Nuclear Hormone Receptors. .. .. . . . . . . . . .. . .. . . . . . . . . . . . . . . . ... . . .. .. 18
D. 24R,25-dihydroxyvitamin D3...........................•..•........... 22
IV. HYPOTlŒSIS 28
v. OB~~~~1rl{)~~ ...............................•..•...........3()
VI. MATE~AND METHODS 32
A. Preparation of Nuclear and <:ytosol Extracts 32
B. <:rode Extracts 33
C. <:ompetition Assays 34
D. Saturation Analysis Experiments 35
E. Sucrose Gradient Sedimentation Experiments 36
F. mRNA Extraction 37
G. Two..Hybrid cDNA Library Constnlctïon 38
H. DNA-Binding Domain Vector <:onstIUction 38
2

IX.
X.
XI.
•
•
•
L Yeast Two-Hybrid Screening 39
J. PCR Screening of the Yeast Two-Hybrid Libraries 39
vu. RESULTS 42
A. Ligand Saturation Analysis of Nuclear Extracts 42
B. Competition Analysis of Nuclear and Cytosol Extraets
and of DBP 42
C. Sucrase Gradient Sedimentation of Nuclear Extraets 45
D. Tissue Specificity 47
E. Yeast Two-Hybrid Screening 48
F. PCR Screening of the Yeast Two-Hybrid Libraries 49
Vill. DISCUSSION 50
A. Ligand Saturation Analysis of Nuclear Extracts 50
B. Competition Analysis of Nuclear and Cytosol Extraets
and of DBP 52
C. Sucrose Gradient Sedimentation of Nuclear Extraets 55
D. Tissue Specificity 57
E. Yeast Two-Hybrid Screening 59
F. PCR Screening of the Yeast Two-Hybrid Libraries 62
G. Alternative Strategies to Clone the 24R,2S(OH)2D3 Receptor 64
S~YJ\lS[[) CONCLlJSIONS 69
ORIGINAL CONTRIBUTION TO KNOWLEDGE 71
REFERENCES 72
3

•Figure
LIST OF FIGURES
Following page
•
•
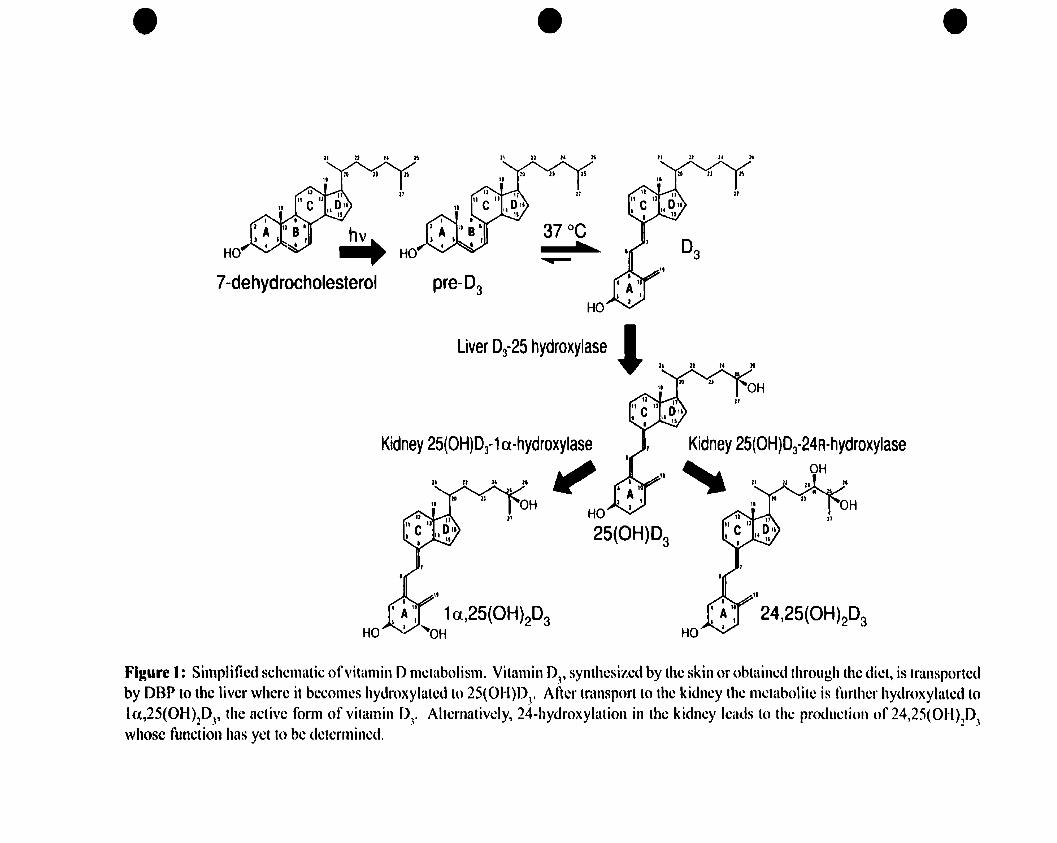
Figure 1: Simplified schematic of vitamin D metabolism 13
Figure 2: Nuclear hormone receptor superfamily structure and classification... 19
Figure 3: VDR-mediated transcription 20
Figure 4: Abnormal bone fonnation in 24-0Hase-deficient mice 25
Figure 5: Saturation analysis of eH]-24R25(OH)2D3 binding bymandiblelcalvaria nuclear extraet. 43
Figure 6a: Specificity of eH]-24R2S(OHhD3 binding bymandiblelcalvaria nuclear extract. 44
Figure 6b: Specificity of eHl-24R25(OH)2D3 binding bymandiblelcalvaria cytosol extract 44
Figure 6c: Specificity of eH]-24R2S(OHhD3 binding by liver nuclear extract 44
Figure 7a: Specificity of eH]-24R25(OHhD3 binding to bovineGlobulin Cohn Fraction IV (Sigma) as a source of DBP 45
Figure Th: Specificity of eH]-24R25(OH)2D3 binding to Gc Globulin(Sigma) as a source of DBP 45
Figure 8a: Complex fonnation between DBP7 actin, and DNase 1. 46
Figure 8b: Sucrose gradient sedimentation of liver nuclearextract after labeling with IoM eHl-24R25(OH)203 46
Figure 9a: Complex fonnation between OBP, actin and anti-actin antibody 48
Figure 9b: Sucrose gradient sedimentation of Iiver nuclear extractincubated with eH]-24R25(OH)2D3 and anti- actin antibody 48
Figure 10: Tissue specificity of the eHl-24R25(OH)2D3 binding- proteinin nuclear extraets 48
4

•
•
•
ACKNOWLEDGEMENTS
To Dr. René St-Arnau~ my thesis supervisor, without whom 1 could never bave
accompüsbed my goals. He bas offered me endIess advice, encouragement, and patience,
both in this project and thesis, as weU as in my pursuit of extra-eurricular interests. 1
would like to thank him for providing me with top of the line scientific and personal
guidance, bath to which 1 am indebted.
To Janet Moir-Brazeau, a coUeague who bas endured, with exceptional patience, my
countless questions and worries. 1 thanle her for taking me througb Many of the
procedures 1 was performing for the fml time, offering sound suggestions and great
companionship.
To Josee Prud'homme, also a cO-worker and always full of encouragement and good
humour. 1 appreciate her belp with setting up the mice matings, the genotyping and the
dissections.
To Serge Messerlian, a feUow graduate student with wbom 1 bave shared a myriad of
frustrations and celebrations and from whom 1have sougbt advice and answers to infinite
queries. 1 thank him for bis companionsbip, understanding, and more formaUy, for bis
contribution of the VDR primers.
5

•
•
•
To Alice Arabian, my co-worker, 1 owe mucb to ber expert instruction and ber sound
reasoning. 1 thanle ber for ber belp with perfonning and interpreting the ligand binding
assays.
To Louise Marineau and Mia Esser, the animal teehnicians, 1 thank them both for aIl their
help with the mice matings and dissections.
To Jane Wishart, the MediCal imager, 1 wisb to thank ber for her help in preparing the
figures in the Introduction.
To Dr. Xiangming Gao, Dr. Olivier Dardenne, Dr. Isabelle Quelo and Dr. Gourgen
Ambartsoumian, my colleagues, 1 very much value aIl of their advice in overcoming my
experimental difficulties, as weIl as their indispensable camaraderie. 1 truly could not
have found a better group of people to work with and 1 hopc the friendships forged
remain strong, despite our diverging paths.
Finally 1 would like to give my appreciation to my Mother and Father, and Tom, for
supporting me in my decisions and allowing me the freedom to choose my own direction.
This research was conducted thanks to funding from the Shriners of North America.
6

• ABBREVIATlONS
25(0H)D3 25-hydroxyvitamin D3
1a,25(OH)2D3 1a,25-dihydroxyvitamin D3
24R925(OH)2D3 24R925-dihydroxyvitamin D3
la-ORase 25-hydroxyvitamin D la-hydroxylase
24-0Hase 25-hydroxyvitamin D 24-hydroxylase
25-0Hase vitamin D 2S-hydroxylase
AF-2 activation function-2 domain
AR androgen receptor
3-AT 3-amino-1 92A-triazole
• ATP adenine triphosphate
bp base pairs
Bmax maximal binding
BSA bovine senun albumin
CAT chloramphenicol acetyltransferase
cDNA complementary deoxyribonucleicacid
CKB creatine kinase brain isoform
CMV cytomegalovirus
C-tenninal carboxy-terminal
DBD DNA-binding domain
DBP vitamin D binding protein
• dCTP deoxycytosine triphosphate
7

• DNA deoxyribonucleicacid
DR3 direct repeat separated by 3 base pairs
dNTP deoxy N triphosphate
DIT dithiothreitol
E17.5 embryonic clay 17.S
EDTA ethylene diamine tetra acetic acid
EGTA ethylene glycol tetra acetic acid
ER estrogen receptor
EtOH ethanol
FXR famesoid X receptor
GR glucocorticoid receptor
• ms histidine
hsp90 heatshock protein of 90kDa
Hyp hypophosphatemic
IP9 inverted palindrome separated by 9 base pairs
Kb kilo base pairs
K<t dissociation constant
kDa kilo dalton
LBD ligand-binding domain
LEU leucine
LUC luciferase
LXR liver X receptor
rnRNA messenger ribonucleicacid• 8

• NCBI National Center for Bioteebnology Information
N-tenninal amino terminal
one omithine decarboxylase
Olïgo dT oligomeric deoxythymidine
PBS phosphate buffered saline
PCR polymerase chain reaction
PMSF phenylmethylsulfonyl fluoride
PPARy Peroxisome proliferator-activated receptor gamma
PR progesterone receptor
PTH parathyroid hormone
Pfu Pyrococcus furiosus
• RAR retinoic acid receptor
RT-PCR reverse transcriptase-polymerase chain reaction
RXR retinoid X receptor
RXRE retinoid X response element
S sedimentation coefficient
Td temperature of dissociation
TR thyroid receptor
VAS upstream activating sequence
UTR untranslated region
VDR vitamin D receptor
VDRE vitamin D response element
• 9

• 1• ABSTRACf
•
•
Although 24~25(OH)2D3 bas been implicated in bone development, its biological
role and mechanism of action remain controversial. In searcb for evidence of a receptor,
nuclear and cytosol exttacts were isolated from mandibles and calvaria of E 17.5 mice.
Competition and saturation analysis identified a saturable, specific and higb affinity
(~=I.lnM) 24R,25(OHhD3 binding-protein. The results of these and sucrose
sedimentation studies indicate tbat this protein is not vitamin D receptor (VDR) or
vitamin D binding protein (DBP). Tissue specificity experiments suggest that this
putative receptor is also present in liver but Dot brain.
pBDGal4-hRXRa bait was used to screen neonate and embryonal
mandiblelcalvaria cDNA libraries using the yeast two-bybrid system. PCR screening was
also perfonned using primees from the zinc-fmger region of the VDR. To date no
positive clones bave been identified. Isolation of this putative receptor will provide
valuable insight ioto the mechanism of this metabolite's role in bone development.
10

•
•
•
o. RÉSUMÉ
Des travaux récents de notre laboratoire suggèrent que le métabolite de la vitamine
D, 24R,25-dihydroxyvitamin 03 (24R,25(OH)2D3), pourrait être impliqué dans le
développement osseux intramembranaire. Nous avons émis l'hyPOthèse qu'un récepteur
spécifique pour la 24R,25(OHhD3 était exprimé dans l'os intramembranaire en
développement. Des extraits nucléaires et cytosoliques de calottes craniennes et de
mandibules ont été préparés à partir d'embryons à 17.5 jours de développement. Des tests
de liaisons utilisant la 24R,25(OH)2D3 tritiée ont mis en évidence un site de liaison
nucléaire de haute affmité et spécificité (Kcs=1.1 nM). L'analyse de ces sites de liaison par
sédimentation en gradient de sucrose a pennis d'éliminer la possibilité que l'activité de
liaison soit dOe au récepteur classique de la vitamine D ou à la globuline sérique de
transport de la vitamine D. L'activité de liaison de la 24R,2S(OHhD3 rot détectée dans
l'os et le foie, mais pas dans le cerveau, suggérant une certaine restriction d'expression
tissulaire.
Une stratégie de criblage par interaction fonctionnelle chez la levure Cyeast two
hybrid screen'), utilisant le partenaire potentiel 'Retinoid X Receptor', a été utilisée dans
l'espoir de cloner le récepteur 24R,25(OH)2D3. La librairie criblée fOt produite à partir
d'os intramembranaire fétal. Une stratégie basée sur l'homologie de séquence probable
entre le récepteur pour la 24R,25(OH)2D3 et le récepteur classique de la vitamine D et
utilisant l'amplification en chaîne ('polymerase chain reaction'; PCR) a aussi été utilisée.
Ces tentatives sont restées infructueuses jusqu'à maintenant.
Il

•
•
•
Le clonage d'un récepteur spécifique à la 24R,2S(OH)ZD3 pennettrait d'approfondir
de façon marquée notre compréhension du mécanisme d'action de la vitamine D et des
mécanismes moléculaires contrôlant le développement osseux.
12

•
•
•
m. INTRODUCTION
A. The Vitamjn D Endocrine System:
Despite its name, vitamin 0 is not a vitamin at a1l, but rather, a hormone. Both
plants and animais are able to syntbesize vitamin 0 during exposure to sunlight. In
humans, what is not formed endogenously in the skin is obtained through the diet from a
variety of plants, dairy products, eggs and 6sh. Vitamin 02 (also referred to as
ergocaiciferol) is the form of the vitamin that is produced in plants, whereas vitamin D3
(cholecaiciferol) is that which is syntbesized by vertebrates. Vitamin~ is unsaturated at
carbon centers Cn and C23 and bas an extra methyl group at C24 (1). Although this form
has been shown to contribute significantly to the vitamin 0 functions of humans and other
vertebrates (2, 3) and undergoes similar metabolic pathways to form a hormonally active
metabolite (4), for the pUl'POse ofthis paper vitamin 03 is the focus.
Figure 1 illustrates a simplified schematic of vitamin D metabolism. Penetration of
sunlight's ultra violet B photons through the skin allows photolysis of a cholesterol-like
precursor called 7-dehydrocholesterol (also termed pro-vitamin D3) present in
mammalian skin, converting it into pre-vitamin 03 (5). UPOn thermoisomerization, this
becomes the more stable secosteroid, vitamin D3 that then enters the circulation where it
binds to the vitamin D binding protein (DBP) (6).
The vitamin D binding protein is the principal carrier of vitamin DJ and its major
metabolites. Il is a serum protein that circulates in high amounts (5 x lO~ M) compared
to 25(OH)D3, its major ligand (5-12 x IO·IM) (7). Although the ligand binding domain of
DBP has highest affinity for 2S(OH)DJ (the most abundant circulating vitamin 0
13
• • •II n Il lt l1 n Il :t II II Il 20
24,25(OH)2D3
ri Il It
HO
Kidney 25(OH)Df 24R-hydroxylase" , OH
HO
25(OH)D3
~
37 oC.....
HO
Liver D3-25 hydroxylase l "pre-D3
Kidney 25(OH)Df 1a-hydroxylase
" " · · JI'
4 Â'THO'-'V, 1a,25(OH)2D
OH 3
~7-dehydrocholesterol
Figure 1: Simplified schcmatic of"itamin Dmetabolism. Vitamin DJ, synthesized by the skin or obtained throllgh the diet, is transportedby DBP to the liver where it becomes hydroxylated to 25(01-l)D
3• After transport to the kidney the Illetabolite is further hydroxylatcd to
1a.,25(OH)2DJ' the active form of vitamin DJ• Alternativcly, 24-hydroxylation in the kidney Icatls to the production of 24,25(OH)2DJwhosc function has yct to be dctcrmined.

•
•
•
metabolite), it is also able to bind other metabolites, including 24R,2S(OHhD3 (but not
24S,25(OH)2l>J, its synthetic cpimer) and la,25(OH)2D3 (8). Several conflicting reports
exist on the relative order ofbinding affinity for sterols by DBP. A couple of reports state
the order is thought to he: 25(OH)D3 = 24R,25(OH)2D3 > la,25(0H)203 > vitamin 03 (7,
9), while others suggest it is: 25(0H)03 > 24R,25(OH)2D3 > la,25(OHhl>J (10, Il), and
still a third shows it could he 24R,25(0H)03 > 25(08)D3> la,25(0H)2D3 (12). Sorne of
these variations may he due to the source of DBP used and/or the conditions in which the
experiments were performed.
Vitamin 03, associated with OBP, gets transported to the liver, at which point the
vitamin D 25-hydroxylase enzyme (25-0Hase) adds a hydroxyl group to C25 (figure 1) (5,
6). This is the initial step in vitamin D activation and leads to the production of
25(0H)D3, the most abundant, but still inactive, form of vitamin D3. The liver production
of 25(0H)D3 does not seem to he stringently regulated and mainly depends on substrate
concentration (1). This inactive metabolite is then carried to the kidney, again by DBP,
where it can follow one of two pathways (figure 1). It can become hydroxylated at either
the la-position by the renal enzyme, 25-hydroxyvitamin D la-hydroxylase (la-OHase),
or al the 24-position by the 25-hydroxyvitamin D 24-hydroxylase (24-0Hase) (5, 6). In
the former situation, the resulting metabolite is la,25(OH)2D3 (calcitriol), the biologically
active form of vitamin D3. The circulating concentration of 1a,25(OH)203 is about 1000
fold less than 25(OH)03 al 5-15 x 10-H M (13). The latter pathway leads to the formation
of 24R,25-dihydroxyvitamin D3 (24R,25(OHhD3), the most abundant dihydroxylated
metabolite in blood (-6 x lO-~ (14). Countless other side chain oxidation pathways
14

•
•
•
lead to the formation of over 40 other vitamin I>J Metabolites, Most of which, however,
are considered non-functional and eatabolic in nature.
The primary role of la,2S(OHhD3 is to regulate calcium and phosphoms
homeostasis. It does this in three ways: 1) by increasing intestinal calcium and
phosphorus absorption, 2) by inducing mobilization of calcium from bone whenreq~
in synergy with parathyroid hormone (PfH) and 3) by increasing renaI reabsorption of
calcium and phosphoros, aIso in conjonction with PTH (15).
1a,25(OH}2D3 and 24R,25(OH)2D3 production are reciprocally regulated. When
calcium is needed, 1a,25(OH)2D3 synthesis is enhanced, while the 24-0Hase
hydroxylation pathway is suppressed. Conversely when calcium levels are adequate,
synthesis shifts to 24R,2S(OH)2D3. This increase in 1a,2S(OH)2D3 synthesis upon
hypocalcemia is secondary to increased PTH and is due to an induction of the 1a-OHase
enzyme. Thus there appears to he a calcium-mediated feedback system regulating the
la-OHase enzyme whicb prevents sustained levels of 1a,25(OH)2D3 that could lead to
hypercalcemia (16). 1a,2S(OH)2D3 production is aIso reguiated by plasma phosphate
levels. As plasma phosphate level decreases, production shifts from 24R,25(OHhD3 to
increased la,25(OH)2D3 synthesis. In addition, high levels of la,2S(OHhD3 act to
decrease 1a-OHase activity and stimulate 24-0Hase activity (1), further regulating
Metabolite levels.
The major site of 24-bydroxylation is in the kidney (1). The cytochrome P450
enzyme, 25-hydroxyvitamin D-24-hydroxylase, is able to hydroxylate both 2S(OH)D3
and la,25(OH)2D3. Contrary to la-OHase, it is positively regulated by la,2S(OH)2D3
15

•
•
•
and negatively regulated by PI'H (17). It seems tbat the 24-hydroxylation of 25(08)1»
and 1(1,25(0H)203 is the first step in a metabolic pathway to inactivate these Metabolites
and halt their fonctions. However, although the major site of expression of 24-0Hase is
in the kidney, its expression in osteoblasts and other tissues indicates it May serve to
sYDthesize active Metabolites (such as 24R,25(OH)203) that could play an important role
in bone mineralization (18). The possible fonctions of 24R,25(OH)203 are discussed in a
later section.
B. Bone Bistogenesis:
It is weil known that 1a,25(08)203 is vital for normal bone formation, as it is an
important regulator of osteoblastic differentiation and function. Abnormalities along the
pathway of the vitamin D endocrine system leads to osteomalacia and rickets. In
addition, a significant amount of evidence exists which suggests that 24R,25(OH)2D3 also
plays an important biological role in bone development. A brief overview of bone
histogenesis will allow for a better understanding of these raIes.
There are three main cell types in the skeleton: (a) chondrocytes, the cartilage
fonning cells which differentiate from eartier multipotential mesenchymal progenitor
(stem) cells; (h) osteoblasts, the bone forming cells which aIso differentiate from
mesenchymal progenitor ceUs and deposit a specialized matrix that undergoes
mineraiization (see below); and (c) osteoclasts, the bone resorbing ceUs which are large,
multinucleated cells derived from baematoPOietic stem cells. Two distinct tyPes of bone
histogenesis exist: intramembranous and endocbondral ossification, the major difference
being the presence or absence of a cartilagjnous phase (19, 20).
16

•
•
•
Endochondral ossification, which occurs in long bones and the growth plate,
involves the formation of a cartilaginous matrix befoIe conversion ioto bone. Essentially,
mesenchymal ceUs differentiate into prechondroblasts and !hen chondroblasts. These
cells secrete a cartilaginous matrix into which they embed tbemselves and become
chondrocytes. During the development of this avascular, embryonic cartilage a ring of
woven bone is formed through the process of intramembraneous ossification (see below)
in the future midshaft area. Angiogenesis then allows blood vessels to invade this
calcified woven bone, bringing along osteoclast precursors. The osteoclasts begin to
destroy the calcified cartilage matrix as weil as excavate the hematopoietic bone marrow
cavity, while osteoblasts are recroited to replace the cartilage witb bone matrve At the
ends of long bones, the cartilage serves as a growtb plate to aIIow longitudinal growth.
The growth plate chondrocytes form columns beginnjng with resting cartilage cells,
followed by proliferating chondrocytes that become progressively larger, hypertrophie,
and eventually die. Their cartilaginous matrix becomes mineralized just below the
growth plate hypertrophie zone. Eventually the calcified cartilage is partially resorbed by
osteoclasts, and osteoblasts (that differentiated from the mesenchymal cens brought in by
the invading blood vessels) begin to deposit woven bone on top of the cartilaginous
remnants. This immature woven bone and calcified eartilage is further remodeled lower
in the growth plate, eventually becoming replaced by mature lameUar bone (19,20).
Intramembranous bone formation, which occurs in the fiat bones (such as the
calvari~ mandible, and clavicle) as well as the outer cortex of the long bones
(periosteum), also begins as a cluster of mesenchymal cells. However, in this case they
do not differentiate into ehondrocytes, but rather undergo proliferation and differentiation
17

•
•
•
directIy into osteoblasts. The proliferating precursors secrete an organic mabix, called
osteoi<L that contains coUagen fibers embedded within it. Once differentiated, osteoblasts
begin to form an immature woven bone by dePOsiting crystals of calcium phosphate on
and around the coUagen fibers in random orientation. EventuaUy the woven bone
becomes invaded by blood vessels, remodeled and replaced by mature, lameUar bane (19,
20).
c. Nuclear Hormone Receptors:
As early as the 1970s steroid actions were shown to he mediated by SPeCifie high
affinity receptor proteins. It was believed that the binding of hormone to its receptor
induced an allosteric change that enabled the hormone-receptor complex to bind to high
affmity sites in chromatin and modulate transcription. However, it wasn't until 1985 that
the nrst nuclear hormone receptor was isolated (glucocorticoid receptor, GR) (21). Sïnce
then receptors have been identified for alI the known fat-soluble, nuclear hormones.
Furthermore, an endless number of so-called orphan receptors have been identified for
which no known ligands have been found, bringing the total number of members of this
nuclear receptor superfamily to over ISO (22). Of these, only about 50 are mammalian
receptors, the rest coming from C. elegans and other species. The majority of these
orphan receptors were discovered by using low stringency hybridization screening, while
still others were found by using more modem molecular cloning techniques, sucb as the
yeast two-hybrid.
Nuclear hormone receptors are Iigand-modulated transcription factors that mediate
responses to steroids, retinoids, and thYroid hormones. They are modular proteins
18

•
•
•
consisting of five distinct domains" each with separate fonctions (figure 2) (22" 23). The
N-tenninal (NB) and the C-terminal (F) domains are highly variable, as is the hinge (D)
region, which allows Cree rotation of the ligand-binding and DNA-binding domains. This
hinge region has also been implieated in heatshock protein 90 (hsp90) interaction as well
as nuclear localization (24). The N-terminal domain is involved in transcriptional
activation (AF-l domain) (in sorne receptors) and promoter selection (24). AlI nuclear
hormone receptors have a DNA-binding (C) domain (DBD) consisting of two highly
conserved zinc-fmgers. This region is responsible for localizing the receptor to specific
DNA sequences (called hormone response elements) on target genes (22). In addition to
protein-DNA interactions this domain also alIows protein-protein interactions (25).
Finally, the ligand-binding (E) domain (LBD) of these receptors, aIso highly conserved, is
responsible for ligand recognition which ensures specificity and selectivity of the
physiologic response. This region aIso encodes the dimerization properties of the
receptors (possessing homo- and/or hetero- dimerization interfaces). Furthermore,
hormone-dependent transcriptional activation (AF-2 domain) and/or repressor functions
are also encoded in this domain (23). Essentially, the LBD is a molecular switch that, in
most cases, tums the receptor on to a transeriptionally active state upon ligand binding
(22). The mechanism for this involves complicated interactions with various co
activators and co-repressors. In simplest terms, co-repressors interact with the unliganded
receptors, resulting in inhibition of basal transcriptionaI activity. Binding of hormone
causes the AF-2 domain to change confonnation, which subsequently leads to the
dissociation of the co-repressors, recJUitment of co-activator proteins, and transcriptional
activation of the receptor (23).
19

•
"....-__....A_I...8 ........;;;,C__~-D...... E FN < i'iIIIft..'_. c
Staroid Receplora
Dimeric 0rphM Aeceptora
tIryroId hotmoI..."."... RA
'.2S-lOHJ7~
~ttoItâ
~
NGf1.8 (CE8-1') '1ELP 1 SF-1 (Rz-F1) '1,
ri i
lIonomeric Orphan Aeceptors
glu(C1 COificoitI
""'.,&01"',,, 'fi
PR7"...........a'.n....e,."
AXA (wp) ~RACOUP/.... '1HNF.... .,
•
•Figure 2: Nuclear hormone receptor superfamily structure and classification. Ail nuclearhonnone receptors have an N-tenninal region (AIB), a DNA-binding domain (C), a hingeregion (0), a ligand-binding domain (E), and aC-terminal region (F). The superfamily isdivided into four classes depending on their dimerization and DNA-binding properties asdescribed in the text. From: Mangelsdorf, DJ., et al. 1995. Cell. 83: 835-839.

•
•
•
The nuclear bormone receptor superfamily is divided into four classes depending on
their dimerization and DNA-binding propertics (figure 2) (22). Class 1 receptors are the
steroid bormone receptors. These fonction as ligand induced bomodimers and bind to
DNA-response elements consisting of inverted repeats. Class fi receptors fonction as
obligate RXR heterodimers and usuaUy bind to directly repeated DNA balf-sites. This
group contains the rest of the known ligand-dependent receptors. Most orpban receptors
belong to either class m or class IV. Class mconsists of the dimeric orphan receptors,
which bind as homodimers to direct repeats. Finally, class IV monomeric orphan
receptors bind to DNA response elements as monomers (22).
The actions of la,25(OHhD3 are mediated by a bigh affinity, highly specifie
receptor, called the vitamin D receptor (VDR). VDR is a elass fi member of the nuelear
hormone receptor superfamily, and thus is able to beterodimerize with RXR. Figure 3
depicts the mechanisms by whicb la,25(OH)2D3, VDR and RXR act to modulate
transcription. When la,25(OH)2D3 enters its target celI, it binds to the unliganded VDR
in the nucleus. The reœptornigand complex heterodimerizes with RXR, conferring upon
the receptor an increased affmity for specific DNA sequences located upstream of the
target genes (Le. in the target gene promoters). Tbese are refened to as vitamin D
response elements (VDRE). This interaction leads to the modulation of transcriptional
activity of target genes responsible for carrying out the pbysiologjcal functions of
A typical VDRE consists of two directly repeated hexanucleotide sequences
separated by three base pairs (DR3). However, DR4 and DR6 type VDREs also exis~ as
do inverted palindromic arrangements of two core binding motifs spaced by nine20

•9-cis retinoic acid (.6)
+1i •
t..U-'ra"s ftlinol0' -\ ~ .... ,("11,011
t1
DietarySources
Sunlight
~coon
.U·trllns retin.ld~hyde
~Cll()
• ll-trllns retinoic uid (8)~C()()fl
t
~4
if ...--110
RXRE
-- .~VDRE t.::::/
ŒI"---c1-.I~~~
1,2S(OH)2D3 (e)
~OH +-4_
.& kidneyHO OH
/t!
l~1: Target1\ Cell\ Nucleus
•
•Figure 3: VDR-mediated transcription. In the presence of la,25(OH)..,D., RXR- )
heterodimerizes with the bound VDR, fonning a transcriptionally active complex on theVDRE. 9-cis-retinoic acid may promote RXR homodimer fonnation, shifting occupiedRXR to RXREs. From: Whitfield, GK., et al. 1995. J. Nutr. 125: 16905-16945.

•
•
•
Ducleotides (IP9) (25). 1be general consensus sequence of the core binding motifs of
Most members of the supcrfamily is RGGTCA (R= A or 0), although there is a high level
of divergence from this consensus in VDREs of various promoters.
Figure 3 also shows the effect of 9-cis retinoic acid (the RXR ligand) on VDRIRXR
heterodimerization. For some RXR heterodimers (termed I&pennissive"), the presence of
RXR ligand acts to potentiate the activation action of the partner ligand. However, this
seems Dot to he the case with the VDR, where, in the presence of 9-cis retinoic acid, RXR
May preferentiaUy homodimerize and bind to RXR. response elements (RXREs), thereby
activating different target genes. This may indirectly destabilize the VDRIRXR
heterodimer formation.
The VDR is present in ail tissues that exhibit a respoDse to la,25(OH)2D3
(including intestine, kidney, parathyroid glands, and bane). Its sedimentation coefficient
is 3.2 S in human and rat, and 3.7 S in chicken (26). Ils abundance (as measured by
la,25(OH)2D3 binding capacity of extraets) in Most la,25(OHhD3 target tissues is fairly
low, between 10 - 100 fmol/mg proteÎD, but can he as high as 1pmollmg protein in certain
tissue extracts (26). This could indicate that those ceUs with a higher VDR content are
more responsive to la,2S(OHhD3, than those with lower VDR expression. However, it
is also likely to involve differences in the ability to intemalize and metabolize the ligand
and/or the availability of partner proteins needed for gene activation (26). Finally,
although highly specifie for la,2S(OH)2D3 (Kct =1-2 x lO-I~, VDR does bind to other
vitamin D metabolites (including 24R,25(OHhD3) with a 500-1000 fold lower affinity
(27,28).
21

•
•
•
It is important to note here that in addition to its genomïc actions, la,25(OHhI>] is
also thought to carry out a nomber of non-genomic activities in various tissues (reviewed
in 28a). These include transcaltaehia (movement of calcium from the intestinal lumen to
the bloodstream), as weil as rapid changes in membrane Iipid turnover and prostaglandin
production. These actions are proposed to he mediated by the interaction of
la,25(08)2D3 with a œil surface (membrane) receptor (VDRmcm) that is distinct from the
nuclear receptor, VDRnuc (28b). Though it bas Dot yet been studied, it is wonhwbile to
keep in mind the possibility that some of the actions of 24~25(0H)203May a1so he
mediated by non-genomic mecbanisIDS. Interestingly, a putative candidate for a
24R,25(OH)203 membrane receptor has already been identified (Il).
The role of 1a,25(OH)2D3 in intestinal calcium and phosphorus absorption,
mobilization of calcium from bone, and renal reabsorption of calcium and phosphoros is
weil established and widely accepted. On the other band, the physiological mIe of
24R,25(OH12D3 remains a controversial topie. 24R,25(OH)2DJ is the Most abundant
dihydroxylated Metabolite of vitamin 03, yet for years scientists bave believed that it is
simply a catabolite of 25(0H)03 whose production is meant only to regulate
la,25(OH)2D3 levels. This line of thought stemmed, in part, from experiments using
analogs of vitamin D that had been tluorinated on C24 (thereby preventing hydroxylation
at this position). When this analog (24,24-ditluoro-25-hydroxyvitamin 03) was used as
the sole source of vitamin D, no effects different from those obtained wben using
25(OH)D3 alone were observed. This was taken to indicate that metabolites of vitamin D
22

•
•
•
that are hydroxylated al position 24 serve no biological function, other than as a
mechanism to inactivate 2S(OH)D3 (29, 30). However, the first evidence contradicting
this hypothesis and suggesting that other vitamin 0 metabolites migbt have biologjcal
importance came from experiments sbowing that administration of la,2S(OH)2D3 alone
was unable to produce the same biologjcal effects as those obtained wben vitamin 03 was
given (31). In these eXPeriments, bens raised solely on la,25(OH)2D3 produced fertile
eggs that were unable to batcb. In contrast, bens raised OD a combination of
la,25(OHhD3 and 24R,2S(OH)2DJ produced eggs able to batcb equally effectively as
those of bens given vitamin D3. These results suggested a bioiogical role for
24R,25(OH)20J tbat bad never been sunnised.
Since then growing evidence bas been obtained for physiological funetions of
24R,25(OH)20J. One of the fust in vivo experiments that suggested a role of
24R,25(OH)2D3 in embryonic development came from Sunde et al. in 1978 (32). These
investigators observed that the impaired development of vitamin D-deficient cbick
embryos could he rescued by treatment of the eggs with vitamin D3 itself, but not
completely by treatment with la,25(OH)2D3. This suggested that other vitamin D
metabolites were necessary for normal chick embryogenesis.
A significant number of reports iodicate that growth plate cartilage is a target organ
for 24R,25(OH)2D3 (reviewed in 33). It was sbown that injected radiolabeled
24R,25(OH)2DJ was preferentially taken up by growtb plate cartilage. In the same set of
in vivo experiments, when radiolabeled 25(08)D3 was injected ioto vitamin D-replete
rats an accumulation of labeled 24R,25(OHhD3 in growtb plate cartilage but not in
articular cartilage was discovered. Moreover, Corvol et al. (34) demonstrated that very
23

•
•
•
low concentrations (10-13 M - 10-10 M) of 24R,25(OHhOJ stimulated proteoglycan
synthesis in cultured rabbit growth plate chondrocytes.
Other lines of evidence suggest a mIe of 24R,2S(OHhD3 in regulation of bone
growth, development and repaire As far back as 1978, Omoy et al. (35) discovered that
24R,25(OH12D] was necessary for the bone healing process of racbitic cbicks. Birds
treated with 24R,2S(OH)2D3 alone grew as weIl as those treated with vitamin D3 but were
unable to maintain calcium homeostasis. Conversely, those treated with la,25(OHhD3
alone could maintain nonnal plasma calcium and phosphorus levels but still were unable
to prevent the racbitic changes in bones. Only birds treated with both dihydroxylated
metabolites were normal in both bone and mineral bomeostasis, indicating that both
metabolites are required but perform distinct functions. Another group of investigators
examined the effects of 24R,25(OH)2D3 on bone fracture healing (36, 37). Firstly, it was
detennined that circulating levels of 24R,25(OH)2D3 are elevated in chicks during tibial
fracture bealing. This is due to an increase in the renal 24-0Hase activity and suggests
that 24R,25(OH)2D3 is involved in the early process of fracture repair (37). Secondly,
Seo et al. (36) aIso demonstrated that a normal physiological concentration (10-9 M) of
24R,25(OH)2D3 is necessary and sufficient for normal bone growth and integrity, and is
essentiaI, in combination with la,2S(OHhD3, for the fracture healing process in chicks.
This confmned in vitro studies by Schwartz et al. (38) based on organ culture of mice
fetallong bones, suggesting a raie for 24R,25(OH)2D3 (at physiological concentrations)
in the growth of fetaI mice bones as shown by increased diaphyseal length, periosteal
bone area, and hydroxyproline content.
24

•
•
•
Using a different approach to examine the putative physiological role of
24R,25(OHhD3 our own laboratory took advantage of the modem teehnology of
homologous recombination to ultimately create mice lacking the 24-0Hase gene (20, 39).
A null mutation in the 24-0Hase gene was created, thereby eliminating the source of
vitamin D3 Metabolites hydroxylated al the 24-position in animaIs homozygous for this
mutation. Nonnal, fertile animaIs heterozygous for the mutation were obtained and
crossed to produce homozygous mutants. These are barn with the expected Mendelian
frequency, though about one-half die before weaning. The reason for this perinatal
lethality has not yet been determined but a possible explanation could he an inability of
these pups to maintain minerai homeostasis. The homozygous pups are unable to
eliminate la,2S(OHhD3, leading to its accumulation and thus MaY die from
hyPercalcemia. The surviving fllSt-generation homozygous mutants are fertile and appear
normal in minerai homeostasis, macroscopic anatomy of liver, spleen, kidney, and 8Ot, as
weIl as in bone histology. This is surmised to he due to the fact that the mother May
provide adequate 24R,25(OH)2~ to the pup during development through the placenta.
However, when homozygous mutants are bom of homozygous mothers, bone
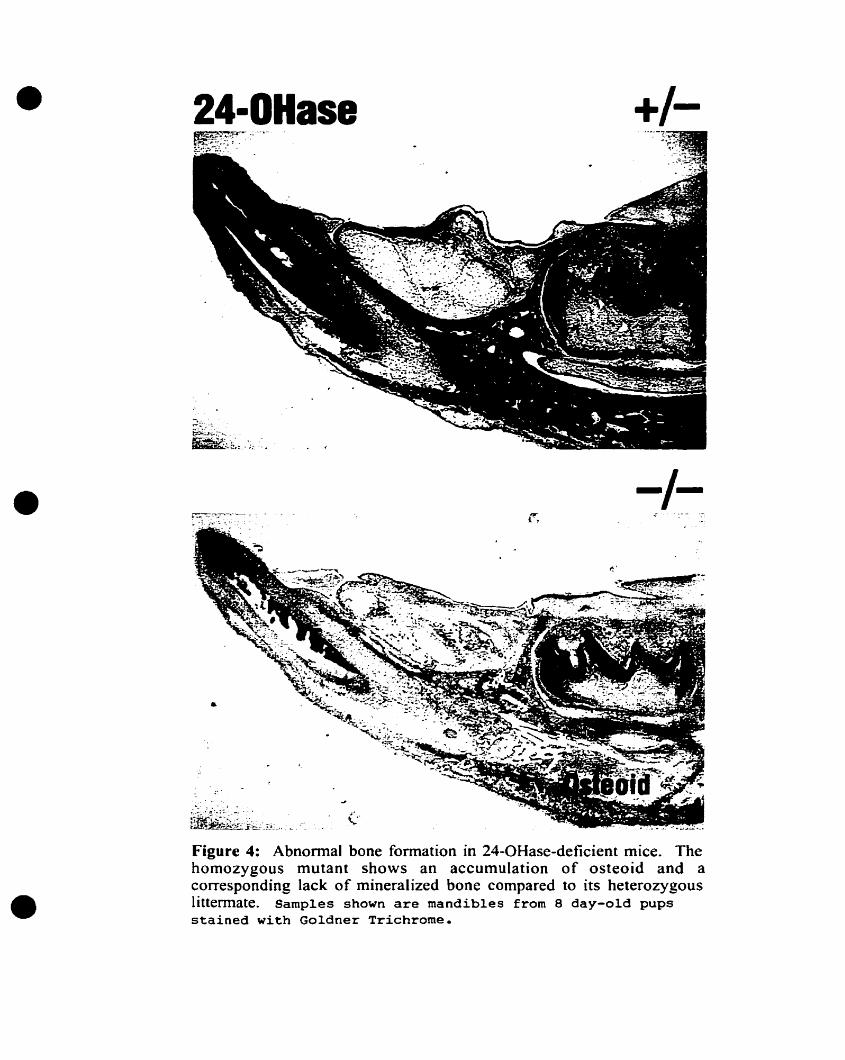
development becomes impaired in the fonn of a mineralization defect. Accumulation of
unmineralized osteoid matrix at sites of intramembranous ossification, such as the
calvaria, mandible, clavicle, and exocortical (periosteal) surface of the long bones, is
evident (figure 4). Heterozygous littermates are nonnal. Though further detailed
examination needs to he done, initial observation of the growth plate seems normal,
possibly indicating that 24R,2S(OHhI>] is not important in chondrocyte maturation in
vivo.
25
•
•
•
24-0Hase
-/-
Figure 4: Abnormal bone formation in 24-0Hase-deficient mice. Thehomozygous mutant shows an accumulation of osteoid and acorresponding lack of mineralized bone compared to its heterozygouslittermate. Samples shawn are mandibles from 8 day-old pupsstained with Goldner Trichrome.

•
•
•
In addition to suggesting a plausible role for 24R,2S(OH)203 during
intramembranous bone development, these results support the existence of specific
putative receptors to carry out the actions of this Metabolite and indicate sites of
intramembranous ossification as a possible location for these receptors. Interestingly, it
has been observed that chick embryos from hens maintained ooly on la,25(08)203 as
their source of vitamin D3 also presented with abnormal mandibles (40). Admittedly, it
could be argued that these effects may be due to the lack of la,25(08)203 catabolism
instead of a requirement for 24R,2S(OH)203. This possibility is currently being
addressed by crossing the 24-0Hase mutant mice with vitamin D receptor (VDR)
mutants. If the observed effects are due ta an excess of la,2S(OH)2D3 acting on the
VDR, then the 24-0HaselVDR double mutant mice should no longer demonstrate the
abnormal intramembranous bone phenotype (39).
Phannacological activity of 24R,2S(OH)2D3 has also been established by various
groups. High doses of 24R,2S(OH)203 have been shown to increase bone volume and
decrease bone resorption in vivo in rats (41), rabbits (42), and ovariectomized dogs (43,
44). Furthermore, pharmacological doses of 24R,25(OH)2OJ produced dose-dependent
effects in promoting bone formation without causing excessive bone resorption in
hyPOphosphatemic (Hyp) mice (a model for familial X-linked hypophosphatemic rickets
in humans). This was in contrast to the effects of equivalent doses of la,25(OH)2D3 (45,
46).
A large number of these slodies have aIso suggested that 24R,25(OH)2D3 and
la,2S(OH)2D3 exert independent effects on various tissues. For example, it bas been
shown that radiolabeled 24R,2S(0H)203 localized to epiphyseaI chondroblasts while26

•
•
•
1~25(OH)2D3 concentrated in diaphyseal booe cells, suggesting that each of tbese
metabolites bas its specifie site of action in developing bone (47). FurthermoIe,
experiments indicate that rat costochoodral cartilage chondrocytes isolated from the
resting zone respond to 24R,25{OHhD3, whereas those from the growth zone respond to
1~25(OH)2D3 (48-50). These responses included protein synthesis, cell proliferation,
plasma membrane and matrix vesicle synthesis, and phospholipid metabolism. Aloog
these same lines, Schwartz et al. discovered that treatmeot of resting zone chondrocytes
with 24R,25(OHhD3 induces their differeotiation and maturation iota growth zone
chondrocytes tbat are lcx,2S(OH)2D3 responsive (51). This suggests that 24R,25(OH)2D3
plays a role in cartilage development. 24R,25(OH)2D3 has also been sbown to stimulate
the activity of the brain isozyme of creatine kinase (CKB) in cultured chick limb-bud
cartilage ceUs, whereas 1~25(OHhD3 does not (52). Moreover, in vivo experiments
demonstrated that CKB aetivity is stimulated by 24R,25(OH)2D3 in the epiphyses of rat
long banes, but by lcx,25(OH)2D3 in the diaphyses of these bones, again suggesting
separate sites of action for the two dihydroxylated metabolites (53). A [mal indication of
the distinct effects of 24R,25(OHhD3 and la,25(OH)2D3 are found in a report by Yamato
et al. (54). These investigators sbowed that 24R,25(OH)2D3 antagonizes the stimulating
effect of la,25(OH)2D3 on the fonnation and fonction of osteoclastic cells.
Although strongly suggestive, the above observations are not unequivocal evidence,
and thus final proof of the different effects of 24R,25(OH)2D3 and1a,25(OH)2D3 will
depend upon the demonstration of two receptors.
27

•
•
•
The presence of hypotbetical 24~25(OH)21>J receptors in growth plate
chondrocytes (55) and epiphysis of rat bone (47, 56, 57) has been suggested by severa!
groups, mostly using autoradiographic and binding saturation slodies. Other slodies have
provided support for the existence of 24R,25(OHhI>J receptors in chick embryo limb-bud
cultures (58) and in the parathyroid gland of chicks (59). These reports are somewhat
outdated and not much follow up has occurred. However, a recent slody by Kato et al.
e11) indicates the quest for the receptor bas not been given up by all, and provides
evidence for a membrane receptor specifie for 24R,2S(OHhD3 in a fracture-healing eallus
of chick tibiae. A1though all these studies are persuasive, they are also merely
insinuative, and thus, the isolation of the receptor has remained elusive to date. We
provide here, evidence of a putative 24R,25(OH)2D3 receptor in intramembranous bone of
the developing mouse embryo and describe attempted efforts for its isolation.
28

•
•
•
IV. HYPOTBESIS
Previous experiments have indicated that a receptor for 24R,25-dibydroxyvitamin
0 3 does exist (56, 58, 59). However, these studies are outdated and have not been
followed up, and so the topic remains controversial. Moreover, tbese studies were
perfonned in long bones (56), parathyroid gland (59) and chondrocytes (58), tissues that
appear normal in the 24-0Hase knockout mice (20). Tbus it is possible that the receptor
bas never been cloned because the wrong tissue was heing examined. The abnonnal
ossification of the mandible and calvaria (and other sites of intramembranous
ossification) in the 24-0Hase knockout mice leads us to propose that a likely site of
expression for the putative receptor would be in these tissues. Furthermore, former
attempts to clone the receptor were often performed on postnatal tissue (56, 59). Since
ossification starts at E14.5, and since it is unknown whether expression of this putative
receptor would be turned off during or after development, it seems more rational to look
at embryonic tissue. Finally, by analogy with the vitamin 0 receptor that binds
1~25(OH)2D3, and by the fact that the structures of other RXR heterodimer ligands are
similar to that of 24R,25(OHhD3, we hypotbesize that the putative receptor will be a class
II member of the nuclear hormone receptor superfamily. It is worthwhile to note that the
retinoic acid system bas more than one Metabolite (all-trans and 9.-cis retinoic acid), eacb
of whicb binds its own nuclear hormone receptor (i.e. RAR, RXR). In contrast, the
estrogen system bas two distinct receptors (ERa and ERP) binding the SarDe Metabolite.
29

•
•
•
Therefore it is not unreasonable to hypothesize that the vitamin D system could similarly
have more than one receptor for its metabolites.
Thus, the hypothesis propose<! is that a receptor for 24R,2S(OH)203 exists in
embryonic mandible and calvaria tissue, and that this receptor is a class n member of the
nuclear hormone receptOf superfamily, heterodimerizing with RXRa.
30

•
•
•
v. OBJEcrIVES AND RATiONALE
The goals of tbis project were tirst ta provide evidence for the existence of a unique
receptor for 24R,2S-dihydroxyvitamin D3 and second, ta attempt ta clone il. This would
further our understanding of the molecular mecbanisms of 24R,2S(0H)21» action and its
function in bone development. The fICSt objective was to look for 24R,2S(OHh03
binding in mouse mandible and calvaria tissue. The rationale behind using these tissues
cornes from the observation that bone development is impaired at sites of
intramembranous ossification (including mandible and calvaria) in mice lacking the 24
OHase enzyme (and hence deficient in 24R,25(OH)203). This was done by isolating
nuclear and cytosol extraets from 17.5 day-old mouse embryo mandibles and calvaria and
performing saturation analysis on them using eH]-24R,2S(OHhD3.
The next objective was to characterize this binding with respect ta its specificity for
24R,2S(OHhD3 versus other vitamin 03 metabolites. Competition assays were carried
out using 10-200 foid excess of various cold metabolites, including 24R,2S(0H)2D3,
245,25(0H)2D3 (the unnatural, inactive epimer), la,2S(OHhD3, and 25(OH)03 as
competitors for binding.
Thirdly, it was necessary to rule out the possibility of the vitamin D receptor (VDR)
or the vitamin D binding protein (DBP) accounting for the observed binding. The VDR
was ruled out on the basis tbat la,25(OH)2D3 did not compete for binding with the eH]
24R,25(0H)2D3 (opposite to wbat would have occurred if the binding protein was the
VDR). Sucrose gradient sedimentation experiments were used to evaluate the possibility
of DBP as the binding protein by comparing proftles elicited by DBP and the nuclear
31

•
•
•
extraet upon addition of actin and DNase I. The basis for this was the observation that
DBP fonns a complex with actin and DNase 1, which cao be visualized as a shift in
sedimentation peak.
Lastly, with enough evidence supporting the existence of a receptor in the tissue,
the next objective was to try to clone it. This was attempted by using the yeast two
hybrid system since il" bad already been successfully employed in our lab and had also
been used by others to identify novel nuclear hormone receptors (60-62). The rationale
for using retinoid X receptor (RXR) as bait was based on functional homology of the
putative receptor with the VDR, which was known to function as a RXR heterodimer. In
addition, the ligands for other class II receptors are structurally similar to
24R,25(OHhD3• Mandible and calvaria tissue was selected for the cDNA libraries to he
screened for the reasons discussed above. The PCR-based screening was employed as an
alternate strategy to isolate the receptor since the highly conserved zinc-fmger region of
the DNA-binding domain of the VDR provided a good region for a 3'primer to allow
amplification of inserts containing a similar motif. Another advantage to this technique is
that it did not require the receptor to he an RXR heterodimer, thereby increasing our
chances of finding the clone.
32

•
•
•
VI. MATERIALS AND METRODS
A. Preparation of Nuelear and Cytosoi Extracts:
Bone (mandible and calvaria), brain, and liver nuclear and cytosol extracts were
isolated using a protocol adapted from Roy et al. 1991 (63). Briefly, bone (O.55g), liver
(2.0g) and brain (l.Og) tissues from 17.5 day-old C57B16 mouse embryos were dissected
and frozen in liquid nitrogen. The tissue was homogenized in a 40ml Dounee manuaI
tissue grinder (Wheaton "200", Minville, NJ) containing 2 ml/gram tissue of NEI buffer
(250mM sucrose, 15mM Tris-HCI [pH 7.9], l40mM NaCI, 2mM EDTA, O.5mM EGTA,
O.15mM spermine, O.5mM spermidine, ImM dithiothreitol [DTI], 0.4mM
phenylmethylsulfonyl fluoride [pMSF), 25mM KCl and 2mM MgCh) (63) using pestle
B. Nonidet P-40 (Sigma Chemical, St Louis, MO) was added to a fmal concentration of
0.5% and the homogenate dounced 5 more strokes with pestle B. The homogenate was
centrifuged in a Sorvall RC-3 centrifuge using the HL-S rotor (Sorvall, Wilmington, DE)
at 2.2K (-lOOOg), 4°C for S minutes. The supematant containing the crude cytosol
extract was flash frozen in 200J.1l a1iquots and stored at -SO°C. The nuclei (pellets) were
washed in 5 ml of NEl buffer containing 0.5% Nonidet P-40 and recentrifuged as before.
This wash supematant was later assayed for protein content and then discarded. The
nuclei were then Iysed in 2 packed cell volumes (PCV) of NE2 buffer (NE1 buffer
containing 3SOmM Kel) (63) containing 0.5% Nonidet P-40, 1110 total volume 4M KCI,
and 11500 total volume O.SM sodium bisulfite (Sigma Chemicals, St. Louis, MO) for 5
minutes at 4°C. After douncing for 20 strokes in a 7ml dounce using pestle A, the
homogenate was transferred to a Sm! Ultra-elear ultracentrifuge tube (Beckman
33

•
•
•
Instruments, Palo Alto, CA) and centrifuged al 24K for 1 hour (4°C) using a Beckman
LB-M ultracentrifuge with the SWS5Ti rotor (Beckman Instruments, Palo Alto, CA).
For brain and liver tissues, the supematant was too viscous to pellet, thus it was
subsequentIy sonicated twice for 5 seconds using an MSE 500 Watt ultrasonic
disintegrator (Measuring and Scientific Equipment Ltd., Sussex, Eng), then recentrifuged
at 35K for 1 hour. The supematant was dialyzed using a O.S-3.0ml Slide-A-Lyzer
cassette (pierce, Rockford lllinois) for 1 houc al 4°C in 500m1s of Dingam's Buffer D
(20mM Hepes [pH 7.9], l00mM KCl, 0.2mM EDTA, O.SmM DIT, O.5mM PMSF, 20%
glycerol). The dialyzed nuclear extract was recovered and spun in a I.Sml
microcentrifuge tube at 13000rpm (4°C) for 10 minutes in an IEC Centra MP4R
microcentrifuge (International Equipment Company, Needham Hts, MA). The
supematant was flash frozen in SOJ1l aliquots and stored at -SO°C. A small amount of
nuclear and cytosol extract was used to determine protein concentration using the Bio-
Rad Protein Assay kit (Bio-Rad Laboratories (Canada) Ltd., Mississauga, Ont.).
B. Crude Extracts:
Crude extraets were isolated from mandible and calvaria of 24-0Hase homozygous
mutant and heterozygous mutant mouse embryos. Tissues were collected at embryonic
day 17.5 (E17.5) as follows: 0.3g of tissue was isolated, placed directIy in liquid nitrogen
and stored at -sooe until ready for extraction. The tissue was crushed in an autoclaved
mortar and pestIe on dry ice until it was a powder. Liquid nitrogen was added to the
mortar, the powder transferred to a 7ml dounce on ice and the liquid nitrogen allowed to
evaporate off. ImL of NE2 buffer containing O.SfI, Nonidet P-40, 1110 total volume of34

•
•
•
4M KCl, and 1/500 total volume of NaBisulfite was added and the mixture was dounced
severa! tilDes with pestle B until viscous. It was then transferred to an ultraelear
centrifuge tube on ice and sonicated twice for 5 seconds. The homogenate was then spun
at 24K for 1 hour at 4°C. The cmde extraet supematant was dialyzed, spun, aliquoted and
quantified as above.
C. Competition Assays:
Mandiblelcalvaria or liver, nuclear (25~g protein) or cytosol (65~g protein) extraets
(in lOO~ of NEl buffer) were incubated with InM (nuclear) or O.25nM (cytosol) eH]24R,25(OH)2D3 (Amersham Pharmacia Biotech, Baie d'Urfe, QC) (in S~ 95% ethanol).
This was done both in the presence and absence of increasing amounts (10-200 molar fold
excess) of cold Metabolites. Non-radioactive 24R,25(08)2D3, 24S,25(OH)2D3 (the
unnatural, biologjcally inactive epimer (64, 65», 1«,25(08)2D3. or 25(0H)D3 (in SJ.1l of
95% ethanol) were used as the competitors for the eHl-24R,2S(OH)2D3 putative receptor
binding interaction. In control assays, either O.5Jlg of Gc-Globulin (Sigma Chemicals, St.
Louis, MO) with O.2Smglml BSA (66) in the same reaction volume, or 50flg of Bovine
Globulin Cohn Fraction IV (Sigma Chemicals, St. Louis, MO) in 320J.Ü fmal volume, was
used as a source of DBP. Each binding reaction was performed in duplicate.
In the ligand binding assays for determining tissue specificity and for all cmde
extraet experiments, 25~g of relevant extract was incubated with 1nM eH]24R,2S(OH)2D3 in the presence and absence of 2S-fold molar excess of unlabeled
24R,2S(OH)2D3.
35

•
•
•
The incubation tubes (I.Sml microcentrifuge tubes), containing extraet or DBP,
labeled ligand and cold competitor, were vortexed to start the binding reaction and
incubated at room temperature for 4S minutes (67). The reaction was terminated by
transferring the tubes to ice and immediately adding 400IJl of hydroxyapatite slurry (67).
The tubes were vortexed and left on ice for IS minutes with vortexing every S minutes.
The samples were then centrifuged at 13000rpm for 3 to 4 seconds in the microfuge. The
hydroxyapatite pellets were washed tbree times with SOOJ.ü of 10mM Tris/HCI-o.S%
Triton X-lOO [pH 7.5] by vortexing and then centrifuging as above. The final washed
hydroxyapatite pellets were extraeted twicc with 9OOJJ.l 2: 1 methanol:chloroform to
ensure complete removal of radioactive ligand (67). The extraction solvent was then
transferred to scintillation vials and dried under a stream of nitrogen. Econotluor-2
scintillation fluid (packard, Meriden, CI') was added to the vials and the samples counted
for 5 minutes at 57% counting efficiency using a TRI-CARB liquid scintillation analyzer
(Canberra Packard Canada, Mississauga, ON). Competition curves were then obtained
using Prism's Non-linear Regression One Site Competition equation (GraphPad Prism
Version 2.0, San Diego, CA). From the ligand binding experiments to determine tissue
specificity, maximum binding in each tissue was calculated (in fmol) and graphed as a bar
graph using Prism.
D. Saturation Analysis Experiments:
The saturation analyses were carried out using a similar method as described for the
competition assays. SJ.lg of mandiblelcalvaria (or liver) nuclear extract were incubated
with eH]-24R,2S(OHhD3 over the range of O.OS - 6.4nM, in the presence and absence of
36

•
•
•
2S-fold excess cold 24R.25(OHhD3 in a final reaction volume of 110J,Ll. AlI reactioos
were perfonned in duplicate. Separation of bound and unbound ligand was earried out
with BioGel HTPGeI hydroxyapatite (BiO-Rad Laboratories, Hercules, CA) as in the
competition assays (67). Bound ligand was counted as above, where total binding
represents bound eH]-24R,2S(OH)2D3 in the absence of non-Iabeled hormone and oon
specifie binding represents bound eH]-24R,2S(OH)2D3 in the presence of excess non-
labeled hormone. Specifie binding was ealeulated by subtracting the non-specifie from
the total binding. The saturation curve was obtained using Prism's Non-linear Regression
One Site Binding (Hyperbola) equation.
E. Sucrose Gradient Sedimentation Experiments:
2SJ1g of liver nuelear extrael (in lOSJ1l of NEt buffer) or IJlg DBP (in lOSIJ,1 of
PBS) was incubated with toM of eHl-24R,2S(OH)2D3 (in SJ.Ll 95% EtOH) for 4S minutes
at room lemperalUre. As per modifications in the protocol of Van Baelen, et al. (68), in
tubes where actin was ta he added, 2J.lg (DBP experiments) or 501J.g (nuelear extraet
experiments) of actin from bovine muscle (Sigma, St. Louis, MO), dissolved in Hepes
buffer (SmM HePes [pH 1.5], O.lmM CaCh, O.lmM NaN3, O.2mM ATP) was used. An
equal volume of bePes buffer alone was added to tubes wbere actin was Dot ineluded. The
actin (or Hepes buffer) was added to the tubes following the 45-mînute incubation and
incubated for another 15 minutes al room temperature.
In tubes where DNase 1 was to he added, the DNase 1 from bovine pancreas
(Boehringer Mannheim, Laval, Canada) was dissolved in O.IM KeI. Once the 15
minutes incubation of actin was completed, 16J.lg (DBP experiments) or 400J,Lg (nuelear37

•
•
•
extract experiments) of DNase 1 was added to the desired tubes containing actin and
incubated for a final 15 minutes at room temperature. To the tubes not containing DNase
1. O.IM KCI was added instead. In aU tubes water made up the final reaction volume to
220J.Ù.
In the anti-actin antibody experlments, 1Of.ù of a 2001J.glml stock of anti-actin
antibody in PBS (Santa Cruz Biotechnology, Santa Cruz, CA) was added to the
incubation mixture containing nuclear extraet or DBP and actin. Finally, 5.5J,11 of 4M KCl
and water was added to the tubes to obtain a fmal volume of 22otJ.l.
Linear 5-25% sucrose density gradients (4.2ml) in 0.3M KCl-TED (O.3M KCl,
10mM Tris, 1.5mM EDTA, ImM dithiothreitol, pH 7.4) (11) were prepared using a
peristaltic pump, in 5m! ultraelear centrifuge tubes that bad been coated for 1 bour with
5% BSA and dried for 1 bour. FoUowing the incubations, aU 220fJ.l of sample was loaded
onto the sucrose gradient with a pipet and tben centrifuged at 240000g (45000rpm) for 20
hours at 4°C (68). Fractions of 4 drops were tben collected into scintillation vials from
the bottom of the tube using a peristaltic pump and counted as in the competition assays.
F. mRNA Extraction:
Mandibles and calvaria (0.59g) from twenty-four 17.5 day old mouse embryos were
extracted and immediately placed into a sterile 50ml screw cap conical tube containing
4ml GTC Extraction Buffer (4M guanidine tbiocyanate, 25mM sodium citrate [pH 7.1])
and 1641Jl of p-Mercaptoethanol. mRNA was isolated from this fresh tissue by foUowing
38

•
•
•
the large-seale protocol for mRNA isolation from tissue samples of the PolyATtraet
System 1000 kit from Promega (Promega, Madison, WI).
G. Two-Hybrid cDNA Library Construction:
Stratagene's HybriZAP-2.1 Two-Hybrid eDNA synthesis kit (Stratagene, La Jalla,
CA) was used ta eonstnlct a cDNA library for ycast two-bybrid screening. FoUowing
Stratagene's protocol, SlJ.g of the isolated mandiblelcalvaria poly(AtmRNA was used to
synthesize the fust strand cDNA, labeled with [a_32P]dCfP (800 Ci/mmol). eDNA was
size fractionated using a Sepharose CL-2B spin eolumn and the fust two fractions were
used separately to continue the procedure. The Gigapack III Gold Packaging Extraet
(Stratagene, La Jalla, CA) was used to package the two fractions, the primary libraries
were amplified, and the secondary libraries in-vivo mass excised, as per the instnletion
manual for the kit. The exeised libraries were amplified following Stratagene's protocol
and the DNA isolated by a1kaline lysis.
H. DNA·Binding Domain Vector Construction:
Stratagene's pBD-Gal4 pbagemid vector (lOJ1g) was digested with EcoRI and Pstl.
The ligand-binding domainlhinge region of buman RXRa was obtained for the bait insert
by cutting the full-Iength clone with EcoRI and Pst 1. The insert was tben subcloned iota
the digested vector and the new bait plasmid was sequeneed to ensure it was in the correct
reading frame.
39

•
•
•
1. Yeast Two-Hybrkl Screening
The prepared two-hybrid libraries were screened foUowing the Library
Transformation and Screening Protocols in Clonteeh's Matehmaker GaI4 Two-Hybrid
user manual (CLONTECH Laboratories, Palo Alto, CA) with the following
modifications: YRG-2 (Stratagene, La Jalla, CA) was the yeast host strain used for the
simultaneous co-transformation of SOOJ.t.g of library plasmid and Img of bait plasmid.
24~S(OHhD3 (lO-6M) was added to ~e transformed yeastjust before plating onto 10-
20 SD 3- plates containing S-lOmM 3-AT. For sorne screeos, the ms3 Ujump-start"
protocol was followed as per the Clontech user manual. HIS3+ clones were assayed for
Il-Gal activity using the colony-lift ~-galactosidase fliter assay (Clontech) and positive
clones grown for 1-2 days in LEU" media. The yeast plasmid DNA was isolated using
Stratagene's Isolation of Plasmid DNA From Yeast protocol (Stratagene, La Jolla, CA),
transfonned ioto XL-l Blue Mlf cells by electroporation and plated on LBIAmp plates.
Insert DNA was isolated from the colonies by alkaline lysis and sequenced using the
ThermoSequenase radiolabeled tenninator cycle sequencing kit (Amersham Phannacia
Biotech, Baie d'Urfe, QC). The sequence Was resolved on a 6% acrylamide gel (19:1 Bis
to acrylamide), analysed by EditSeq program (DNASTAR IDc, Madison, WI) and
compared to known sequences using a BLAST search of the NCBI (National Center for
Biotecbnology Information) data bank.
J. PCR ScreeniDg of the Yeast Two-Hybrid Libraries:
PCR was used as an aItemate method of screening the yeast two-hybrid libraries.
PCR reactions (SOJÙ) were set up containing 1~ of library DNA (sOOng/JÙ), S~ lOx
40

•
•
•
cloned Pfu DNA polymerase reaction buffer (Stratagene, La JoUa, CA), 4OIJ.l 820, and
IJ.Ù eacb of IOmM dNTPs, S'and 3' primers (SOpmlJ1l) and Pfu polymerase enzyme
(Stratagene, La Jolla, CA). As a negative control no DNA was added to the tube, and as a
positive control IIJl (-IOOng) of a plasmid containing a full-Iength buman vitamin D
receptor (VDR) insert was added instead of library DNA.
Analysis of the full-Iength sequence of VDR showed that it has the highest
sequence similarity with FXR (Famesoid X Receptor) and LXR (Liver X Receptor),
especially in the zinc-finger domaine Based on this, an internaI 3' primer was chosen in
an area of tbis domain where there is only a 1 bp difference between VDR and FXR and a
2 bp difference between VDR and LXR out of the 21 bp length of the primer. Thus this
primer consisted of 21 bases of the 3' end of the zinc-finger region of mVDR (S'> AAT
GTC CAC GCA GCG TIT GAG <3') (Td=70.10C). The 5' primer was either 21 bases
of the 5' end of the zinc-linger region of mVDR (S'> TGT GOA GTG TGT GOA GAC
CGA <3') (Td=68.5°C) or 21 bases of the pADGal4 vector upstream of the c10ned inserts
(5) GAT ACC CCA CCA AAC CCA AAA <3') (Td=67.8).
The peR reactions were placed in a Hybaid OmnïGene (InterSciences, Markham,
Ontario). The screening by PCR was done as follows: After a 2 min denaturation at
95°C, the cycling parameters were: 95°C for 30 s,65°, 62°, or 59°C for 1 min, and 72°C
for 1 min, for a total of 30 cycles. A finaI extension of 10 min at 72°C was added. The
PCR products were directly anaIyzed by gel electrophoresis through a 1.2% agarose gel.
Bands of interest (<300bp when using both VDR primers, and up to 3kb when using the
41

•
•
•
pADGal4 5' primer) were isolated from the gel and the DNA extraeted usmg the
QlAquick Gel Extraction Kit (Qiagen Inc., Mississauga, Ont).
The DNA of severa! bands was subcloned into TOPIO ceUs using the TOPO TA
Cloning System (Invitrogen, Carlsbad, CA). Before perfonning the TOPO Cloning and
the One Shot Transformation Reactions, the addition of 3' A-overbangs post
amplification was perfonned as per the protocol appendix. White colonies (containing
insert) of each PCR reaction product were cultured ovemight in LB medium containing
lOOlJ,glJ.1l ampicillin and the miniprep DNA isolated using alkaline lysis. This was then
digested with Eco RI and run on a 1.2% gel. A couple of IDÏnipreps of each band were
selccted for DNA sequencing using the ThennoSequenase radiolabeled terminator cycle
sequencing kit (Amersham Pharmacia Biotech, Baie d'Urfe, QC). The sequence
reactions were loaded on a 6% acrylamide gel and the DNA sequences obtained were
subjected to a National Center for Biotechnology Information BLAST search to compare
with known sequences.
42

•
•
•
vu. RESULTS
A. Ligand Saturation Analysis of Nuclear Extrads:
The search for a nuclear receptor which binds specifically to eH]-24R,2S(OH)2D3
was based upon, among other studies, the observation that mice lacking the 24-0Hase
enzyme suffer from abnonnal development of bones formed by intramembraneous
ossification (39). Since this phenotype was observed in the mandible and calvari~ these
tissues were isolated from 17.5 day-old CS7B 16 mouse embryos, homogenized, and
purified nuclear and cytosol extracts were obtained. Saturation analysis using tritiated
24R,25(OH)2D3 over the range of O.OS-6.4nM was carried out on the nuclear extraet to
test for the presence of saturable ligand binding to a protein present in the extract. Figure
5 shows the saturation curve obtained from this experiment. As is evident in the figure,
saturable binding of eH]-24R,2S(OH)2D3 was observed in the mandiblelcalvaria nuclear
extraet. This binding is of high affmity (Kci =1.1 ::t 0.14nM) and has a Bmax of 1.79 ::t
0.08 pmoVmg proteine An identical saturation analysis experiment on liver nUclear extract
was later carried out and also demonstrated saturable binding, with a Kct =0.44 ± 0.07 DM
and a Bmax of 2.34 ±0.11 pmollmg protein (data not shown).
B. Competition Analysis of Nuclear and Cytosol Extracts and of OBP:
In arder to elucidate the ligand specificity of the nuclear extract protein binding
24R,2S(OH12D3, competition analysis experiments were conducted on both cytosol and
nuclear fractions using related vitamin D Metabolites as competitors of eH]24R,2S(OH12D3 binding. These Don-radioactive, competitor Metabolites were: the
43

•
2000
76543
Bmax 1.79 ± O.OSpmoVmg protKd 1.1nM ±O.1nM
21O-T=---~---r-----r-----""------'r----"""-----'
o
500
al 1500c=ac_:aE.!:! e- 1000I-f/)
•Figure 5: Saturation analysis of [3H]-24R25(OH)2D3 binding bymandible/calvaria nuclear extract. Aliquots of nuclear extract (5J.1g
protein) were incubated with [3Hl-24R25(OH)2D3 over the range ofO.05-S.4nM, in the presence or absence of 25-fold excess ofnonlabeled hormone for 45min at room temperature. Specifiebinding was calculated as outlined in Materials and Methods.
•

•
•
•
synthetic epimer 24S,2S(OHhI>], la,2S(OHhDJ, and 25(OH)~, in addition to unlabeled
24R,25(OHhD3. Figures 6a and b show the competition curves for eHl-24R,2S(OH)2D3
binding in mandiblelcalvaria nuclear and cytosol extraets, respectively. In figure 6a, it is
evident that non-radioactive 24R,2S(OH)2D3 competes Most effectively with the labeled
ligand for binding to the nuclear protein and thus has the highest affinity for the binding
sites compared to the other vitamin D3 metabolites. At only a 2S-fold molar excess
([competitorl =2S DM) of cold 24R,2S(OHhD3, the specific binding of labeled ligand to
the protein drops to 6%, compared to the 33-38% specific binding seen with competition
by the same excess of either 24S,2S(OHhD3 or 25(0H)03. These latter two Metabolites
were equal in tbeir ability to displace the eHl-24R,2S(OHhD3 from the binding protein.
Finally, la,25(OH)2D3 competes only minimally for binding, even at 200-fold molar
excess ([competitor] = 200nM). This specificity of the 24R,2S(OHhD3 binding protein in
the nuclear extraet suggests that it is not the vitamin D receptor (VDR) since
1a,25(OHh03 would have had the highest affmity for the VDR (27, 28, 69), which is
clearly not the situation.
As seen in figure 6b, similar trends appear in the cytosol extraet, though in aU cases,
there is somewhat less competition tbroughout the curves (e.g. at So-fold molar excess of
unlabeled 24R,2S(OH)203 specific binding is less than 5% in the nuclear extraet whereas
it is 29% in the cytosol extraet). Finally, nuclear and cytosol extraets were a1so isolated
from the livers of the same 17.5 day-old embryos and competition assays perfonned on
these. Ligand binding was observed, and the competition assays elicited a similar profùe
to those of the bane extraets (figure 6c).
44

•100
6
75mc:cc:a • 24R2503u
i 50 0 24825036 1a25D3
(1)25(OH)03
~A
0
25
•00.0 0.5 1.0 1.5 2.0 2.5
log (competitor]
Figure 6a: Specificity of [ 3Hl-24R25(OH)2D3 binding tomandible/calvaria nuclear extract. 251J,g of nuclear extract protein was
incubated for 45min at room temperature with 1nM eHl-24R25(OH)203and various concentrations (1o-200nM) of unlabeled vitamin 03 analogs.
•

•
Cft 75c:acliu;; 50"ü8.fi) •fi. 24R25D325 •
0 2482503• --6-1a25D3• 25(OH)03
a0.0 0.5 1.0 1.5 2.0
log [competitor]
Figure 6b: Specificity of [ 3H]-24R25(OH)2D3 binding tomandible/calvaria cytosol extract. 65J.1Q of cytosol extract proteinwas incubated for 45min at room temperature with O.25nM
[3H]-24R25(OH)2D3 and various concentratoins (2.5-50nM) ofunlabeled vitamin 03 analogs.
•

•100r---"2r'"----..A..-_
en 75c:ac 24R2503:s •() 0 24S2503;;: 50 1Œ25D3·ü 6
B- .6 25OHD3en';R.0
25•o-l------'T-----~----=:::;:=~I-------,0.0 0.5 1.0 1.5 2.0
log [competitor]
Figure 6e: Specificity of [3Hl-24R25(OH)203 binding to Iivernuclear extract. 251J,9 of nuclear extract protein was incubated for
45min at room temperature with 1nM [3H]-24R25(OH)203 and variousconcentrations (2.5-50nM) of unlabeled vitamin 03 analogs.
•

•
•
•
ln order to determine if there was a difference in the amount of binding seen in
heterozygous versus homozygous 24-0Hase knockout mice, crode extraets were obtained
from the mandible and calvaria of bo~ as explained in Methods. Traditional ligand
binding assays, using 25J1g of extraet, tnM of eHl-24R,2S(OH)2D3, and the presence or
absence of 25-fold excess cold 24R,2S(OH)2OJ, were Performed on these extracts.
Specific binding was calculated in fmollmg protein and compared between both
genotyPes. The heterozygous mouse embryo crode extract bound 748 ± Il fmol
ligand/mg protein while the homozygous extraet bound less than half of that at 336 ± 9
fmolligand/mg protein (data not shown).
It is weU known that 24R,2S(OH12D3 can bind to other proteins (e.g. VDR and
OBP) (7, 69). DBP (vitamin D binding protein) acts as a carrier protein for 2S(OH)D3 in
the circulation (7). It is possible that DBP could bave been present in the nuclear extract
due to contamination from blood, though this is unlikely since the nuclei were washed
weIl before lysing. Nevenheless, it was necessary to mie out the POssibility that the
protein in the extraet that was binding the ligand was DBP. Thus competition assays
were conducted using DBP as the binding protein, and the same metabolites as
competitors, with the intention of comparing the competition profIles of the two proteins.
lnitially, Bovine Globulin Cohn Fraction IV (Sigma Chemicals, St Louis, MO),
known to bind 24R,2S(0H)2D3 specifically, was used as a source of DBP to carry out the
experiment. Figure 7a shows that the order of competition in this case was different from
that obtained when using the nuclear extraet. Instead of a noticeably more effective
competition by 24R,25(OH)2D3 as seen with the extraet, when using DBP
24R,25(OH)2D3 and 25(0H)D3 competed equally with the labeled ligand, followed by
4S

•
• 24 5 3
o 24S25034 la25D3A 25(OH)D3
25
100r- ~
•O.....l...----r"------~-----__r_-----____,
0.0 0.5 1.0
log [competitor]
1.5
Figure 78: Specificity of [3H]-24R25(OH)2D3 binding to bovineGlobulin Cohn Fraction IV (Sigma) as a source of DBP. 50J,lQ ofDBP was incubated for 45min at room temperature with O.OSnM
eH]-24R25(OH)203 and various concentrations (O.S·17nM) ofvitamin 03 analogs.
•

•
en 75c:gc:s •u
i 50
en • 24R250afi!. 0 245250a
25 6 1a25D3
• 25(OH)Da• 01.0 1.5 2.0
log [competitor]
Figure 7b: Specificity of [ 3H]-24R25(OH)2Da binding toGe Globulin (Sigma) as a source of DBP. 0.51-19 of OBPwas incubated for 45min at room temperature with O.25nM[3H]-24R25(OH)20a and various concentrations (6.251OOnM) of unlabeled vitamin Da analogs.
•

•
•
•
24S,25(01{h03 and finaUy la,25(0H)20J. The OBP competition profile obtained here
agrees with one reported arder of binding affinities for sterols by DBP, which is:
25(OH)DJ =24R,25(0H)203 > 1«,25(08)203 (7, 9). This suggests that OBP and the
protein in the nuclear extraet are not the same protein sincc they elicit a different
competition profile.
Other investigators (7, 10, 68) have used Sigma's Gc-Globulin (Sigma Chemicals,
St. Louis, MO) as a source of DBP. In arder to verify their results we repeated the
competition experiments using this agent. However, as cao he seen in figure Th, very
little competition was obtained even when increasing to 400-fold excess. Although the
findings of the fmt experiment suggested that the protein in the nuclear extract binding
the ligand was not DBP, since reports on the order of sterol affinities for DBP are
conflicting (7, 9, 10, 12), as were our own observations, more convincing evidence was
necessary to determine whether or not OBP was binding the 24R,2S(OHhD3 in the
mandiblelcalvaria nucIear extraets.
c. Sucrose Gradient Sedimentation 01 Nuclear Extracts:
To further ensure that the binding observed in the extracts was not due to
contamination by DBP present in semm, an approach was taken that exploits the
interactive propenies of DBP, actin and DNase 1 (7, 68). Using a modified version of
Van Baelen' s approach (68), a sedimentation profile of DBP in a sucrose gradient system
was obtained. Figure 8a shows that when DBP a10ne was incubated with eH]24R,25(OHhD3 the labeled DBP elution peaked al fraction 18. Upon addition of actin,
this peak shifted to fraction Il. This indicates that the actin bound to the DBP (68),
46

•
•
S-u
1500
1000
500
-M-OBP--0- OBP+Actin-e- OBP+Actin+DNase1
403020
fraction number
10o-l-i~~~~~~~~~..,....-r-"""""""""'I"""""-.--r"~--r-1~I""""T"'""'f"""'T"-'f"""'T"-~
o
Figure 8a: Complex formation between DBP, actin and DNase 1. 11J,g of
DBP (Gc Globulin, Sigma) was incubated with 1nM eHl-24R25(OH)2D3 for45 min at room temperature. The 220).11 samples were layered onto 50/0-250/0sucrase gradients prepared in 0.3M KCI-TED butter and spun @ 240000gfor 20h. Fractions were collected using a peristaltic pump. Fraction 1 =bottom of the tube. DBP (1IJ,g), Actin (2IJ,g), DNase 1(16IJ,g) .
•
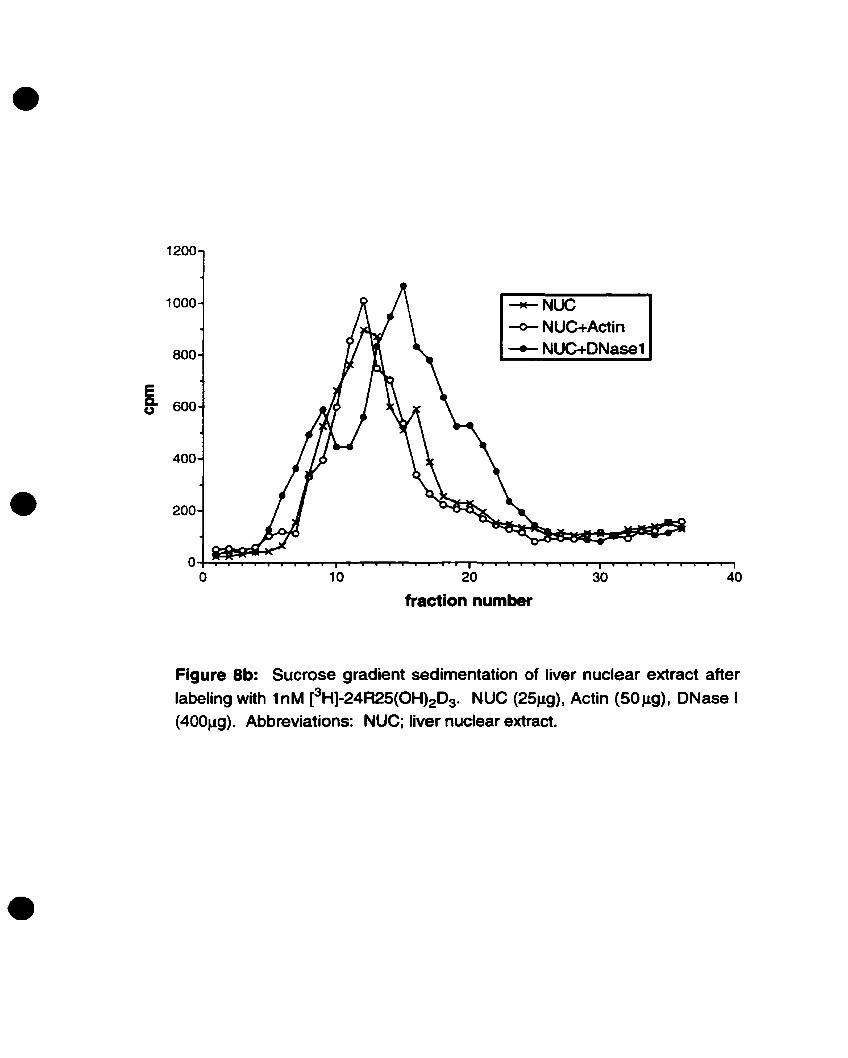
•1200
4030100~;"";";-'T'"""'f"-r-.,....,.........-P"""'T"""'P-"I""--""""'_---"--"'-T"""T"~r""'"r'""T""""I'-T""""I""""T'~t""""T".......-r--T""""""'''''--'''''''''
o
1000 ~NUC
~NUC+Aetin
800 ~ NUC+DNase1
S. 600u
400
• 200
Figure Sb: Sucrose gradient sedimentation of liver nuclear extract after
labeling with 1nM [3H]-24R25(OH)2D3' NUC (25Jlg), Actin (SOJlg), DNase 1(400Jlg). Abbreviations: NUC; liver nuclear extract.
•

•
•
•
making it heavier 50 that il migrated furtber down the tube during the spin and was eluted
in an earlier fraction when drops were collected from the bottom of the tube. Also in
accordance with Van Baelen's observations, the eH]-24R,2S(OHhDJ-DBP peak was
further shifted to an earlier fraction towards the bottom of the tube (fraction 8) with the
addition of both actin and DNase 1, indicating DBP formed a trimer with tbese
compounds (68).
To compare the sedimentation profile of the nuclear extract with that of DBP, the
experiment was repeated onder the same conditions, using 2SJ1g of liver nuclear extraet
instead of DBP. Sïnce previous competition and saturation experiments with liver
nuclear extract showed tbe same propertîes as the mandiblelcalvaria extract, which was in
short supply, liver extraet was used in these experiments. When the nuclear extraet alone
was incubated with eH]-24R,2S(OH)2DJ the bound protein elution peaked at fraction 12
(figure Sb). This eH]-24R,2S(OH)21>]-nuclear protein peak did not shift towards an
earlier fraction upon addition of exogenous actin alone or actin and ONase 1 (figure 8b).
These results show that the protein in the nuclear extraet that is binding eH]
24R,25(OH)20J does not complex with actin or with DNase 1 and suggest that the nuclear
24R,25(OH)2D) binding activity is not due to OBP.
To provide evidence against the possibility that the binding activity was due to DBP
complexed with tissue-derived actin (11), another sedimentation experiment was
perfonned using anti-actin antibodies. This would determine if tissue-derived actin was
present in the sample or not. Anti-actin antibodies would bind to actin present in the
sample and the eH]-24R,2S(OHhD3-nuclear protein-actin peak would shift to an earlier
47

•
•
•
fraction. If no actin was binding the nuclear protein in the sample then no shift would he
seen.
Figure 9a is a conttol experiment using DBP and exogenously added actin to ensure
correct conditions for the anti-actin antibodics. DBP atone elutes neac fraction 18. The
addition of anti-actin antibody to the DBP alone does not cause a shift in the elution~
as expected. When actin and DBP are present in the tube, the elution peak is shifted to
the left (fraction 15). Finally, wben DBP, actin and anti-actin antibody are incubated
together, the elution peak shifts further to the left to fraction Il. This indicates that the
antibody is binding to the DBP/actin dîmer, making the complex heavier so that it elutes
in an earlier fraction.
Next, the experiment was repeated using the nuclear extract. Figure 9b shows that
the elution of the protein in the nuclear extraet alone, when incubated with eH]24R,25(OH)2D3, peaks at fraction 12. When anti-actin antibody is added to the nuclear
extract the elution peak does not shift to an earlier fraction, suggesting there was no actin
hound to the nuclear proteine This was perfonned in duplicate. This provides convincing
evidence that the nuclear protein binding eHl-24R,25(OH)2D3 is not DBP complexed
with endogenous actin.
D. Tissue Speclficlty:
In order to test for tissue specificity of the eH]-24R,25(OH)2D3 binding protein
observed in the mandiblelcalvaria nuclear extraets, ligand binding assays were also
Performed on isolated liver and brain nuclear extraets of the same mouse embryos, using
25J1g protein and loM labeled ligand. The resulls, summarized in figure 10, indicate that
48

•1750
•
1500
1250
i 1000
u750
500
250
~DBP
~DBP+ab
-+- DBP+actin~ DBP+actin+ab
403020
fraction number
10o~_"~i:t:~~~,....-r-"'l~--r-T"""T""""'I"-r-T''''''''''''''''''''''''~::!~~~
o
Figure 9a: Complex formation between DBP, actin and anti-actin
antibody. OBP was incubated with 1nM [3H]-24R25(OH)2D3 for 45min atroom temperature. The final 220J,ll samples were layered onto 5-25%sucrase gradients prepared in O.3M KCI-TED buffer and spun @240000g for 20h. Fractions were collected using a peristaltic pump.Fraction 1 = bottom of the tube. DBP (1IJ.Q). actin (2 IJ.g). ab (2 J.lg).Abbreviations: ab; anti-actin antibody.
•

•
504020 30
fraction number
10O-+-r...........~.,...,.....,...,..."I""""'I"""I""""I"~ .......... "I""""'I".,........~ .......................r--r-r""'T""'r"""I"""............,........~..........r--r-....................'Y""'T"""I
o
200
1000
800 ~nuc
-<>- nuc+ab-4- nuc+ab (2)
600
S-u
400
•
Figure 9b: Sucrose gradient sedimentation profile of 25J.lQ liver nuclearextract incubated with 1nM [3H]-24R25(OH)2D3 and anti-actin antibody toensure the absence of tissue-derived actin in the extract. Duplicates wereperformed with the antibody. NUC (25J.1Q), ab (2f.1g). Abbreviations: NUC;Iiver nuclear extract, ab; anti-actin antibody.
•

•
12.5
10.0
'0C~0,g 7.5
CI):::::'"
Q 0LI) EN:e,a:::~ 5.0~%
CI)
• 2.5
0.0....1.---bone liver brain
•
Figure 10: Tissue specificity of the protein binding eH]-24A25(OH)2D3 in
nuclear extracts. Maximum fmol of [3H]-24A25(OH)2D3 bound by 25f.1Q ofnuclear extract protein from 17.5 day old mouse embryo bone, liver, andbrain.

•
•
•
there is very little binding in brain extraets (60.0 ± 2.0 fmol/mg nuclear protein),
compared with that of bone (304 ± 16 fmoVmg nuclear protein). Furthermore, liver
produced a maximal binding of (432 ± IOfmoVmg nuclear protein), even greater than that
of bone. This data suggests some degree of tissue specificity of the protein expression.
E. Yeast Two-Hybrid Screening:
The yeast two-hybrid strategy for cloning the putative 24~2S(OH)2D3 receptor was
based on the premise that since 24R,2S(OH)203 and la,2S(OH)2D3 are functionally and
structurally related ligands, the putative 24R,25(OH)203 receptor may he of the same
class as the vitamin D receptor (VDR) that binds la,25(OH)2D3. As a member of the
Class fi nuclear hormone receptors, VDR heterodimerizes with the retinoid X receptor
(RXR) (70-73). On this basis we constructed a bait protein consisting of the hRXRa
ligand-binding domain fused in frame with the GAL4 DNA-binding domain in the pBD
GAL4 yeast expression vector. As mentioned previously, this ligand-binding domain
(LBD) of the receptor contains the dimerization interface. This bait was used to screen
two cDNA libraries fused to the GAlA activation domain in the pAD-GAIA vector: a one
week-old mouse calvaria library, and a 17.S-day-old embryonal mouse mandiblelcalvaria
library. After verification of the size of the inserts in the libraries it was determined that
the average insert size was between SOObp - 1.Skh, though inserts ranging to 5kb were
obtained.
The yeast two-hybrid screens of the post-natal library repeatedly pulled out a
-1.7kb insert whicb turned out to be the mouse peroxisome proliferator-activated receptor
49

•
•
•
gamma (pPARy). This is anather member of the class n nuclear bormone receptors and
is known ta interact with RXR (74). This finding demonstrated that the tools and the
methodology used were functional. In addition, non-specific artifacts~ such as mouse
osteopontin~ collagen~ osteonectin and ribosomal proteins were also pulled out of bath the
post-natallibrary and initial screenings of the embryonic library. These false-positives
are one of the major drawbacks of the yeast two-bybrid system. As of yet~ no potential
positive clones bave been identified. Continued screening of the embryonic library is
underway.
F. PCR ScreeniDg of the Yeast Two-Bybrld Libraries:
The rationale for using a PCR-based strategy for attempting to clone the putative
24R,25(OH)2D3 receptor is based on the fact tbat the DNA-binding domains of nuclear
hormone receptors are highly conserved and contain two zinc-fmger motifs (22). Thus by
PCR-amplifying the cDNA libraries using primers from the zinc-fmger region of the
VDR and lower annealing temperatures, il sbould he possible ta amplify fragments in the
library containing similar zinc-finger regions. In addition ta potentially identifying such
transcripts, this strategy may also pull out the VDR since the primers were generated
from its sequence. The PCR screens, performed onder the parameters outlined in
Materials and Methods section~ produced four bands. Deposition of their sequences in
the NCBI <1ata bank ÏDdicated that they were artifacts. Further screens altering the PCR
parameters and/or cbanging the primers will continue to he carried out.
so

•
•
•
VOL DISCUSSION
Many previous studies bave indicated a role for 24R.2S(OHhD3 in bone function
(35-39,45-47, 51, 54, 64, 75, 76). Still others have suggested the existence of receptors
for this vitamin D Metabolite (56, 58, 59). However, the evidence to date bas been
inconclusive and therefore the exact biological role and the mecbanism of action of
24R,25(OH)2DJ have not yet been elucidated. The purpose of this study was to provide
evidence for the presence of, and try to isolate, a putative receptor for 24R,2S
dihydroxyvitamin D3 in developing bone.
Recent evidence in our own laboratory bas demonstrated that mice lacking the 24
OHase enzyme (and consequently, vitamin D metabolites hydroxylated at the 24
position) suffer from abnonnal development of bones formed by intramembraneous
ossification, such as the mandible, calvaria, and clavicle (figure 4) (20, 39). These results
support the notion that 24R,25(OHhD3 does bave an important fonction in bane, that
receptors mediating its function could exist, and that a probable sile of their expression
would he intramembraneous bane.
Based on these observations, we bave isolated nuclear and cytosol exttacts from
mandiblelcalvaria tissue of 17.5 day-old CS7B16 mouse embryos in an attempt to identify
binding of 24R,2S(OHhD] to a protein in this tissue using ligand binding studies.
A. Ligand Saturation ADalysis of Nudear Extracts:
Saturation analysis is used to determine if, with a given amount of protein and the
addition of increasing amounts of ligand, binding of ligand saturates as all the available
51

•
•
•
receptor molecules become occupied. This experiment demonstrated the presence of
saturable binding of eHl-24R925(OHhD3 to a protein in the nuclear extraet. This high
affinity binding <Kd = 1.1 ± O.14nM) strongly supports the notion that there is a
24R,25(OH)2D3 receptorlbinding protein in the nuclear extraet and that the number of
these receptors available for ligand binding is finite and abundant (Bmu = 1.79pmoVmg
protein). These values compare with the~ (1.79nM) and Bmax (2.79pmoVmg protein) of
a putative cytoplasmic 24~25(OH)2D3 receptor described by Somjen et al. (56), but not
with those (1Cd = 15.3nM, Bmax = 43.9 fInol/mg) of a putative membrane receptor
described by Kato et al. (11). This could suggest that different fractions (i.e. nuclear,
cytosol, membrane) or possibly even different tissues, may contain unique
24R,25(OH)2D3 receptors for separate functions. Altematively, variations observed in the
abundance of the receptor could suggest that certain tissues (with higher Bmu) are more
highly responsive to 24R92S(OH)2D3 than those with lower levels ofexpression (26).
The circulating concentration of 24R,2S(OH)2D3 is reported to he hetween loM
(chick) (37) and 6nM (human) (14). This is in the same order of magnitude as the Kct of
the putative receptor (1.1nM) and MaY not cause a problem. However, it could be argued
that, with a circulating 24R,25(OHhOJ concentration of as mucb as 6nM and a receptor
with a dissociation constant of ooly 1.1nM, all available receptors could he occupied all
of the lime and there would he no on/off mecbanism. One possible explanation is that
sorne of the circulating 24R,2S(OH)2D3 is bound to DBP and is thus not available to bind
the putative 24R,25(OH)2OJ receptor. Another possibility could he that 24R925(OH)2D3
is acting via an intraerine mecbanism, wberebya specific ceU produces 24R,2S(OH)2D3
which binds the receptor within the same celle In sucb a case, the receptor would not he
52

•
•
•
exposed to the higher circulating levels of 24R,2S(OHh~. There is evidence of such an
intraerine action of la,25(OHh~with the VDR in a chick myelomonocytic œil line
(77). As weU, there are reports of similar intraerine and autocrine functions for other
nuclear honnone receptors including the androgen receptor, AR (78-80) and the
progesterone receptor, PR (81, 82). Ooly once it is possible to determine if the putative
receptor localizes to specific ceU types known to produce 24R,2S(0H)2DJ will it he
feasible to test this hypothesis.
B. Competition Analysis of Nuclear and Cytosol Extracts and of DBP:
It is known that the vitamin D receptor (VDR) binds lo.,2S(0H)2D3 with highest
affinity and exquisite specificity. However, with 500-1000 fold lower binding affinity, it
can also bind other vitamin D Metabolites, including 24R,2S(OH)2DJ (27, 28). Moreover,
it is also well documented that plasma vitamin D binding protein (DBP), which circulates
in the blood at very high titers (5 x 10-6 M), is also capable of binding 24R,25(OH)2DJ (7,
9, 10, 12). Therefore it remains crucial to determine the ligand specificity of this
24R,25(OH)2D3 receptorlbinding protein and to eliminate the possibility that the binding
seen in the saturation experiments was due to either the VDR or DBP.
In order to do this, ligand competition assays using structurally related vitamin D
Metabolites as competitors were carried out. Assays performed on nuclear and cytosoI
extracts exhibited similar competition profiles, though competition was comparatively
less in cytosol than in nuclear extraets. This does not necessarily signify that there is
another unique receptor for 24R,2S(OHhDJ in cytosol, but could conftrm the notion that
nuclear hormone receptors are found in the cytosol as weU as in the nucleus (83, 84).
S3

•
•
•
Altematively, the lower level of competition seen in the cytosol could be due to the
presence of DBP since the cytosol fraction did not have washes, incœasing the possibility
of contamination with blood. The contaminating DBP could he binding some of the cold
metabolites, theœby leaving less available to compete for binding to the protein.
Another possible explanation for the differing levels of competition between
nuclear and cytosol fractions involves complex formation of nuclear honnone receptors
with accessory factors in the cytoplasm. It is reported that Many nuclear hormone
receptors (e.g. GR, PR, ER, but not TR, RXR or VDR) are found in the cytoplasm
complexed to accessory proteins (83-87). Tbese bave been characterized as 9 S
complexes consisting of the receptor bound with a 90kDa beat shock protein, hsp90
(and/or other accessory factors) and are mostly observed in cytosol fractions (83, 88).
This is in contrast to the 4 S receptor in the nucleus. However, there are significant
differences between receptors in this family with respect to: 1) their ability to bind hsp90
and/or other proteins, 2) where this complex is found (for some receptors (e.g. AR and
ER) the 9 S receptor complex is thought to he mainly in the nucleus in intact cells,
leaking out into the cytoplasm only upon celllysis), and 3) its effect on the affmity of the
receptor for its ligand (84, 88). Tberefore, it remains possible that the lower affinity of
the putative 24R,25(OH)2D3 binding protein for the ligand seen in the cytosol extract
(figure 6b) could he due to the receptor heing complexed to hsp90 or other accessory
factors. To determine if the cytosol protein is complexed to other factors sucrose
sedimentation experiments cao he carried out on both the mandiblelca1varia cytosol and
nuclear extraets. The profl1es elicited from these can men he compared. A sbift of the
peak in the cytosol extract to an earlier fraction (i.e. towards the bottom of the tube)
54

•
•
•
compared with that of the nuclear extraet would indieate that the cytosol binding protein
was complexed to other factors, making it beavier.
In both the cytosol and nuclear fractioDS, the putative receptor clearly demonstrated
highest binding affmity for 24~25(OH)2D3 (figures 6a and b). An important result
obtained from these experiments was tbat the putative receptor showed little competition
from 1a,25(OH)2D3. This indicates the receptor had a low aftmity for la,25(OHhD3. If
the protein in the extraet was the vitamin D receptor, one would expect la,25(OHhD3 to
compete most effectively for the binding sites, compared to the other metabolites. As this
is clearly not the case, these results therefore provide unequivocal evidence that the
protein binding 24R,25(OHhD3 in the extracts is not VDR. Identical competition assays
and saturation analyses were carried out using the liver nuclear extraet as weil (figure 6c).
Since these elicited similar proftles as the bone extract we concluded that the two tissues
contained the same receptor and therefore could he used interchangeably.
Vitamin D binding protein (DBP), which is known to bind circulating 25(OH)DJ, is
present in high amounts in serum (7). Though contamination of the nuclear extract is
unlikely because of the washes performed in its isolation, it remained important to rule
out the possibility of DBP being responsible for the 24R,2S(OH)2D3 binding seen in this
extract. An attempt was made to do this by carrying out similar competition assays using
DBP instead of extract with the intention of comparing the two binding profiles. In the
frnt of these experiments, using bovine globulin Cohn fraction IV as the source of DBP,
the DBP did not appear to distinguish between 24R,2S(OH)2D3 and 2S(OH)D3, whereas
the nuclear extraet protein had a distinctly higher affinity for 24R,2S(OH)2D3. This was
in agreement with some published reports on DBP (7, 9) and suggests that DBP and the
55

•
•
•
nuclear protein are not one and the same. However, as mentioned in Resolts, a suitable
competition assay could not he obtained when using Gc-Globulin as the source of DBP
(figure Th), even at 400-fold excess of cold metabolite. This could bave been because too
little of the protein wu used or too much labeled ligand prevented competition by the
cold metabolites. Since this conttoversy prevented elimination of the plssibility of DBP
involvemen4 more convincing evidence from alternative experiments was necessary.
c. Sucrose Gradient Sedimentation of Nudear Extracts:
The strategy used to determine the presence or absence of DBP in the nuclear
extract was based upon observations tirst described by Van Baelen, et al. (68). The
investigators used sucrose gradient sedimentation to detennine that DBP, actin and
DNase 1 can fonn a trimeric complex that cao he viewed as a shift in the elution peak in
sedimentation experiments. Based on this we compared sedimentation proftles elicited
from DBP with those obtained from the nuclear extraets that we prepared.
The sedimentation constants for DBP, DBP + actin, and DBP + actin + DNase 1 are
4.1 S, 5.8 S, and 7.0 S respectively (68, 89). Since the nuclear protein elutes al a peak
between those of DBP a10ne and DBP + aclin, and assuming a linear relationship, an
estimated sedimentation value of s.s S can he given for the nuclear binding proteine A
more accurate value could he assigned by using known standards in the sante tube as the
sample. This would eliminate error due to slight variations in the gradients as well as in
the number of fractions collected. Again it remains possible that this value is actually that
of the receptor complexed to an accessory factor aod not the value of the receptor alone.
S6

•
•
•
The shifts that occur to the DBP peak upon addition of actin alone or actin + DNase
1 (figure Sa) are not seen with the nuclear extraet peak upon tbese same additions (figure
Sb), suggesting tbat the protein responsible for 24R,2S(OHh1>J binding in the nuclear
extract is not DBP. Only one doubt remained. In a similar experiment by Kato et al. (11)
it was shown that the reason for the lack of shift upon addition of external actin was the
presence of tissue-derived actin already hound to the protein in the sample (which
therefore was confirmed to he DBP). In addition, as is evident in figure Sb, the peak
obtained upon the addition of DNase 1 and actin to the nuclear extraet was not as clean as
the others (probably due to a disturbance of the sucrose gradient during the experiment).
For these reasons, a final experiment was performed in order to be absolutely sure that
DBP was not responsible for the ligand binding seen in the nuclear extraets.
Exploiting the same principles of complex formation, anti-actin antibody was added
to the nuclear extraet tube and layered on a sucrose gradient to try to detect the presence
of tissue-derived actin. In this experiment, if a shift of the eHl-24R,2S(OH)2D3-nuclear
extraet peak towards an earlier fraction was seeo, compared to that of ouclear extract
alone, it would be an indication that tissue-derived actin was hound to the protein in the
ouclear extraet. This would then strongly suggest that the protein binding the ligand was
DBP. However, as shown in figure 9b, wc did not observe a shift of the peak. This
therefore indicates tbat the lack of shift seen upon addition of actin to the nuclear extraet
iD the fust set of sedimentation experiments was not due to the presence of tissue-derived
actin, which further supports the evidence that the 24R,2S(OH)21>J-binding protein in the
nuclear extraet is not DBP.
57

•
•
Absolute, unequivocal praof of this cao only he obtained by using nuclear extraets
Crom DBP knockout mice ta do the ligand binding experiments (90). If the nuclear
extract of these mice no longer binds the eHl-24R,2S(OHhD3 this would he evidence
that, in the normal mice, the binding was due to DBP and not an independent receptor for
24R,25(OHhD3. On the other hand, the presence of binding in the DBP knockout mice
would provide unequivocal evidence that DBP is not a factor. These experiments are
currently being arranged, using the DBP knockout mice from Dr. Nancy Cooke (90).
Taken together, these ligand binding assays strongiy support the existence of a
specific receptorlbinding protein for 24R,25(OH)2D3 in developing intramembraneous
bone. Furthennore, we have proven that this putative receptor is different from the
vitamin D receptor (VDR) and seems not to he the vitamin D binding protein (CBP).
D. Tissue Specificity:
Since the phenotype observed in the 24-0Hase knockout mice occurred in bone
(specifically in those formed by intramembraneous ossification) this suggested a possible
site of receptor expression. However, an interesting question to pose was that of tissue
specificity. In arder to detennine ü this putative receptor was specifie to bone or ü it was
also present in other tissues, nuelear and cytosol extracts were obtained from the livers
and brains of the same 17.5 day-old mouse embryos. Ligand binding assays were tben
carried out to detennine if 24R,25(0H)2D3 binding could he detected in either of these
tissues. Using identieal amounts of nuclear extraet protein (2SJ,lg), identical
•concentrations of labeled ligand (toM) and 2S-fold molar excess of unlabeled
24R,25(0H)2D3 (2SnM), ligand hound (in finol) was calculated and compared between
S8

•
•
•
tissues. It is impottant to note tbat these maximum ligand binding values do not represent
the number of binding sites present (as the SIDa values from the saturation analyses do)
since these experiments are carried out in sub-saturating conditions.
As is evident in figure 10. brain tissue at this stage of development did not show
much binding in comparison to bone, indicating it is unlikely to he a site of expression of
the putative receptor. A literature searcb failed to find reports wbere fonction of
24R,25(OHhD3 bas been described in the brain. However, the level of binding seen in
liver was even greater than in bone. suggesting that there is high expression of the
putative receptor in tbis tissue. This observation is of particular interest since the
presence of putative 24R,25(OHhD3 receptors in the liver could suggest another possible
site of action for 24R,25(OHhD3. The livers of the 24-0Hase knockout mice bave never
been histologically examin~ nor bas the cause of death been detennined since it is
unlikely that these mice are dying from their bone abnonnalities. It is possible that a lack
of 24R,25(OH)2D3 during development could lead to abnormalities in the liver. possibly
severe enough to cause death. Livers from tbese mice are currently heing barvested and
examined by histological techniques for any possible abnormalities to detennine if this is
the case.
Anotber area of interest that would warrant looking into, would he sites of
endochondral ossification. Past reports have indicated endochondral bone as a target for
24R,25(OH)2D3 fonction and a candidate for receptors (35. 36. 38, 56, 64, 75). However,
this contradicts the evidence obtained in our laboratory, which shows tbat endocbondral
bone formation appears nonnal in the 24-0Hase mockout mice (20, 39). In addition.
many of tbese former experiments used long bones (such as tibiae and femur) which have
59

•
•
•
an exocortical surface (periosteum) that is fonned by intramembraneous ossification (20)
and could affect the results. Ligand binding assays on nuclear extraets isolated from
growth plates at the same stage of development could address this controversy.
E. Yeast Two-Hybrid ScreeniDg:
The above ligand binding studies have provided strong evidence that a
receptorlbinding protein for 24~2S(OH)2D3 is expressed in developing
intramembraneous bone. The task remained to clone it. The yeast two-hybrid system
was chosen for this purpose because there is documented use of this strategy for pulling
out new nuclear hormone receptors (60-62). In addition, the system has been used with
success in our own laboratory and many of the tools (the kits, the reagents, the yeast etc.)
were already available. Furthermore, a one-week-old mouse calvaria osteoblast cDNA
library fused to the Gal4 activation domain in Stratagene's pAD-Gal4 vector was already
constructed. This library was used for initial yeast two-hybrid screening using the
hRXRa(LBD)-pBDGal4 bait.
This strategy repeatedly pulled out a -1.7kb insert encoding for the mouse
peroxisome proliferator-activated receptor gamma (mPPARy). PPARyis a known class n
member of the nuclear hormone receptor superfamily that heterodimerizes with RXRa
(74). Thus, this was an encouraging result as it indicated that the sttategy and tools heing
employed were functional. However, due to the unusually high nombers of this insert
pulled out over several screens, it was suspected that during amplification of the excised
library a misrepresentation of inserts bad occurred leading to its over-representation.
Unfortunately, re-excision and re-amplification of the library faile<! to correct this
60

•
•
•
problem and mPPARy was pulled out severa! more tilDes. This strategy eould also he
expected to identify other known RXR-interacting partners present in bane (sueh as VDR,
RXR or RAR). RXR was not isolated since it ooly homodimerizes in the presence of its
own ligand and these experiments were performed using 24R,25(0H)2D3, not 9-cis
retinoic acid as the ligand. Neither VDR Dor RAR were identified in the screens,
suggesting that their representation was low in the screened libraries. In addition to
ppARy, non-specifie artifacts sueh as mouse collagen, osteopontin, and ribosomal
proteins were pulled out. Sueh '-ralse-positives" are a common problem with this system
and certain proteins seem to he pieked up more often than others (e.g. ribosomal proteins,
collagen-related proteins, heat shock proteins, mitochondrial proteins etc.) (91). This
could he because these proteins may have surfaces (such a large hydrophobie surfaces)
that have low affinity for Many different proteins, alIowing them to form complexes
strong enough to he detected in the yeast two-hybrid system (92). These false positives
remain one of the system's major drawbacks.
The bone phenotyPe of the 24-0Hase knockout mice was evident during embryonic
development (20, 39). In addition, it is unknown if the expression of this putative receptor
would he tumed off during development, preventing its c1oning. On this basis, it was
decided to conslIUct another eDNA library from mRNA of the mandible and ealvaria of
17.5 day-old mouse embryos, since allligand binding assays were perfonned al this age.
Screening of this library has so far ooly isolated other non-specifie proteins that seem to
he sticking to the bait.
A Jack of isolation of a potential clone al this stage does not negate the existence of
a receptor. There are several possible explanatioDs as to why this method May not he
61

•
•
•
picking up the desired clone (or other known clones besides ppARy, such as VDR or
RAR). The fust is that Many nuclear hormone receptors have very long 3' untranslated
regions (3' UTR's). Stratagene's Two-Hybrid cDNA sYDthesis kit uses an oligo (dT)
primer to convert the poly(A)+mRNA ioto the first-strand cDNA. If the receptor has a
long 3'UTR it is possible that the reverse transcriptase enzyme was interropted during the
reverse transcription before it reached the region of the gene encoding the dimerization
domain. If this is the case, either no fusion protein is created or it MaY not include the
necessary residues for an interaction with the bait (93). Thus, even though the mRNA
transcript for the putative receptor may have been present, it would not be detected in the
yeast two-hybrid library. The isolation of ppARy, which contains a long 3' UTR, as weil
as the presence of long inserts in our libraries are good indications that this may not he a
problem in our case. Other possibilities are that the gene for the putative receptor is not
present in the correct reading frame in the GAlA-AD library or tbat the hybrid proteins
are not stable in the yeast. preventing detection of an interaction (93). Finally, it remains
possible that the receptor is not a class II nuclear hormone receptor and doesn't interact
with RXR. in which case the yeast two-hybrid strategy using RXR as a bait would not
pull out the clone.
An interesting question to address was that of possible upregulation of the receptor
in mutant mice homozygous for the 24-0Hase gene knockoUL If, in resPOnse to a lack of
its ligand. the putative receptor becomes upregulated (in a similar fashion to some
enzyme/substrate reactions), then it would he wortbwhile to constnlct a new bybrid
library from these homozygous mutant mice (witb a higher receptor representation) in
order to increase our chances of cloning it. Firstly, in order to detennine if the receptor is
62

•
•
•
upregulated in a situation where there is a Jack of ligan~ cmde extraets were obtained
from mandible and calvaria of both the homozygous and heterozygous mutant mice.
Ligand binding assays were perfonned on bath as a measure of receptor levels. Binding
of the labeled ligand (in fmollmg protein) in the homozygous extract was less than balf of
that obtained from the heterozygous extraet (336 ± 9 finol/mg protein vs. 748 ± Il
finol/mg protein). Since this is the case, it would oot he useful to construct a library from
these knockout mice, however, these results MaY suggest that a Jack of 24R.2S(OH)2D3
could lead to downregulation of ils receptor. Interestiogly, Mahonen et al. (94) have
shown that 1a,2S(OHhD3 induces the expression of VDR mRNA and protein in bone
cells. Thus it seeOlS reasonable to suggest that 24R,2S(OH)203 may have a regulatory
fonction on its own receptor, much in the same way as 1a,2S(0H)2D3 does with its
receptor.
F. PCR Screening of the Yeast Two-Bybrid Libraries:
In addition to the yeast two-hybrid strategy, we have employed another approach to
clone the putative receptor. One such method uses PCR to take advantage of the fact that
nuclear hormone receptors have highly conserved DNA-binding domains containing two
zinc-fmgers. On the basis of functional homoJogy with the VDR, it foUows that the
putative receptor for 24R,2S(OH)2D3 would have highest sequence homology in this
region with the VDR. Thus by using a downstream primer from the zinc-linger region of
the VDR and an upstteam primer from the pAD-Gal4 vector, it should he possible to
amplify inserts containing a similar zinc-fmger domain and anything upstream of il in the
63

•
•
•
insert. Although three different annealing temperatures were used, the four bands that
were isolated using tbis method tumed out to he artifacts.
These results do not invalidate the procedure, however. Good primers are keyand
optimal conditions must he determined in order to ensure that the primers do not stick to
everything, yet are still able to pick up inserts with sequences that vary sligbdy from the
primers. Setting the annealing temperature depends upon the similarity of the primer to
the insert to he amplified, which in our case is UnknOWD. The greater the similarity
between the primers and the putative receptor, the cIoser the annealing temperature
should he set to the Td of the primers. In order to decrease the likelihood of picking up
artifacts the annealing temperature can be increased to 66- 68°C. However, this must he
done on the assomption that the putative receptor sequence in the primer area will he
identical to or only a few base pairs different from that of the VDR primer being used. It
is also possible to decrease the annealing temperature helow 59°C to try to pick up the
receptor under the assumption that its similarity to the VDR will he further away than that
of FXR or LXR. Another alteration that cao he made is to vary the concentration of salt
in the buffer and then alter the annealing temperatures, since salt concentration is known
to play a factor in primer annealing. Altematively, in order to reduce the number of
artifacts that are produced in this procedure the numher of cycles could he decreased from
30 to 25, since increasing the nomber of cycles augments the likelihood of amplifying
non-specifie targets.
As is the case with the yeast two-hybrid system, fallure to identify the putative
receptor using this strategy does not indieate the receptor is not in the tissue we are
looking at. This is because the strategy requires that the DNA-binding domain containing
64

•
•
•
the zinc-linger regioo of the putative receptor was reverse transeribed in the library, not a
guarantee, especially if the receptor bas a long 3'UTR. Non-representative amplification
of the libraries could a1so bias the results of this type of screeo. A possible solution
would he to do RT-PCR on the mRNA rather tban the cDNA libraries. Thus, continued
pursuit using this and other strategies will he ongoing.
G. Alternative Strategies to Clone the Z4R,2S(OR)JI)3 Receptor:
ln the event that the putative receptor is not cloned using the yeast two-hybrid or
PCR screening strategies, alternative methods could he developed to attempt cloning.
One option is to use the strategy that was employed to isolate the majority of the orphan
receptors before the yeast two-bybrid system evolved. This involves constructing a
degenerate oligonucleotide probe corresponding to the most higbly conserved region of
the nuclear hormone receptor family, the DNA-binding domain (DBD) (95). This could
then he used to screen a mandiblelcalvaria cDNA library onder low stringency conditions
(96). The advantage to using such a procedure is that it does not require the putative
receptor to he a class fi member interacting with RXR. However, since there is no way of
increasing the specificity of the screen towards the desired clone tbis sttategy would most
likely identify other nuclear hormone receptors found in bone such as VDR, R.XR, RAR,
or ER.
Another possible strategy involves a modification of the yeast one-hybrid system.
The classical yeast one-hybrid system was devel0Ped as a method to identify and clone
genes encoding for proteins that reco8Dize a specific DNA sequence (97). The system
bas two basic components: 1) an expression library of hybrid proteins constmcted by
65

•
•
•
fusing protein coding sequences to the Gal4 transeriptional activation domain, and 2) a
UAS-less lacZ reporter gene containing multiple copies of the binding site of interest
within its promoter region. Wben transfonned into yeast, hybrid proteins that recognize
this binding site will induce expression of the reporter gene and tum the cell blue in a p
galactosidase assay. This occurs due to the hybrid protein's dual ability to bind the
promoter region and activate transcription. This type of classical one-hybrid system
would not work in our case due to the fact that we do not know the sequence of the
response element to which the putative receptor would bind.
A modification of this system bas been developed to screen for possible ligands for
orphan receptors. In this strategy, the ligand-binding domain (or portions thereof) of an
orphan receptor is fused to the Gal4 DNA-binding domain and transfonned into a
mammalian cellline containing a lacZ reporter gene with a Gal4 UAS. Putative ligands
are added to the ceUs which are then assayed for activation of the reporter gene. The
Gal4 DBD of the fusion protein would bind the Gal4 UAS and, in the event ofbinding by
a tnle ligand, the AF-2 domain of the receptor would adopt a confonnation that would
allow the teeruitment of coactivators and activation of the reporter gene. Clones are then
identified by their blue color upon addition of X-Gal.
In our case, a sunHar protocol could he used for the purposes of isolating a receptor
for a known ligand. In this procedure, a new mandiblelcalvaria cDNA library would he
constnlcted fused with the Gal4 DNA-binding domain in a mammalian expression vector.
This library would he ttansformed into mammalian ceUs and the cells exposed to
24R,2S(OH)2D3. The Gal4 DDD cao interact with the GAIA upstream activating
sequence (UAS) located upstream of an integrated GALl-lacZ gene. If one of the cDNA
66

•
•
•
inserts fused to the GaI4 DBD encodes the putative receptor, then upon addition of
24R,25(OH)2D3 to the transformed cells, a conformational change would occur within the
AF-2 domain of the receptor which, together with the Gal4 DBD would activate
transcription of the GALl-lacZ gene (98). These colonies could then he recognized by
their blue color upon incubation with X-gal. In the absence of 24R,25(OHhD3 or if the
insert does not encode the putative receptor then transcription would not he activated and
the ceUs would not tum blue. A possible drawback to this system is that if the library
insert does not encode the correct portion of the LBD of the receptor, the method would
not pick it up. In addition, other activators could he cloned if they contain an activation
domain that does not require its ligand to he active. On the other hand, this strategy does
not rely on the hypothesis that the putative receptor interacts with RXR and is aIso fairly
straightforward to cany out.
Previous studies indicate that 24R,25(OHhD3 induces the activity of the brain
isoform of creatine kinase (CKB) in cultured chick-embryo limb-bud cartilage ceUs (52)
and in the epiphyses of rat tibiae (53), whereas la,25(OH)2D3 does not. FUIthermore, it
was shown that this increase in activity was due to an increased rate of synthesis of CKB,
not just an increase in the activity of the enzyme that was already present. In addition,
other reports indicate 24R,2S(OH)2D3 induces omithine decarboxylase (ODe) activity in
the epiphyses of rat long bones (64). These results suggest that the CKB and OOC
promoters may contain a 24R,25(OH)2D3 response element. In light of tbis, another
strategy for isolating the putative 24R,25(OH)2D3 receptor arises.
We have obtained plasmids containing various lengths of the ODe promoter from
Dr. Andrew Butler (99) as weil as plasmids containing various lengths of the CKB
67

•
•
•
promoter from Dr Micbael Ritebie (100, 10l). Tbese promoters would he subcloned
behind a lacZ reporter gene. In addition, an expression library driven by a mammalian
promoter (ex. CMV) would he constructed from the EI7.S mouse mandiblelcalvaria
rnRNA. The expression library and one of the promoter constructs would then he
transfected together into CV-1 cells (or any other mammalian cellline that does not sbow
background) and exposed ta 24R,2S(OH)2D3. If the library contained a cDNA insert tbat
encoded for the putative receptor the ligand would bind to il, the receptorRigand complex
(heterodimerized with RXR, if necessary) would bind to the 24R,2S(OHhD3 response
element in the CKB or OOC promoter and drive the expression of the lacZ gene. These
blue clones would he picked, the plasmid DNA isolated, and the insert amplified by PCR
using primers from the library expression vector. A possible drawback to this strategy is
that the insert would bave to encode for bath the DBD and the LBD of the receptor in
order for transcriptional activation to he possible.
An alternative to using the expression library in the strategy would he ta transfect
the promoter constructs into ceUs that are known to respond to 24R,25(OH)2D3 on the
assumption that if the ceUs respond to the ligand then they May contain a receptor for it.
In this situation, it would he necessary to transfect only the promoter plasmid and add
ligand. Past studies have sbown that kidney ceU cultures of one-week old rats respond to
24R,25(OH)2D3 and not 1a,25(OH)2D3 but lose this responsiveness as they approacb 4
weeks, at wbich time they become only responsive ta la,25(OHhD3 (102, 103). In
addition, it has been reported that this response profùe parallels the profl1e of the binding
activity of the respective ligands, suggesting the presence of receptors. Other studies
have indicated that cbick mesencbymal cells (52), rat epipbyseal chondrocytes (58), and
68

•
•
•
cultured growth plate cbondrocytes (34) are also œsponsive to 24R,2S(OHhI>]. If there
was a receptor for the ligand in any of tbese cells, then upon transfection of the promoter
plasmid into, for example, one-week old rat kidney ceUs and addition of 24R,2S(OHhIl],
a ligandlreceptor complex would form (with RXR if necessary), bind to the response
element in the promoter and drive the reporter gene. Progressive deletion of the promoter
would then lead to the identification of the sequence of the response element, whicb could
then he used to isolate the receptor using the classical yeast one-hybrid strategy as
described above.
The above strategies provide a wide may of possibilities to approach the task of
isolating the receptor from a variety of angles. With the curreot technology and the
increasing evideoce supporting the existence of a 24R,2S(OHhD3 receptor, it should just
he a matter of time hefore it is cloned. Once it has beeo cloned it will he necessary to
completely sequence il, to characterize it for binding specificity using other vitamin D
metabolites, and to identify a putative DNA binding site. 115 pattern of expression will
need to he studied, and i15 ttanseriptional regulatory function elucidated in order to
progress towards a full understanding of the mechanisms of 24R,2S(OH)2D3 action.
69

•
•
•
IX. SUMMARY AND CONCLUSIONS
Traditionally, it has been believed that 24R,25-dihydroxyvitamin D3, the most
abundant dihydroxylated vitamin D metabolite circulating in the blood, does not have a
unique physiological role but rather is simply the initial Metabolite in vitamin D
catabolism. However, previous studies have suggested that a fonction for
24R,25(OHhD2 does exist, and still others indieated the existence of specifie receptors
for this metabolite. Most recently, our own laboratory has demonstrated that mice
Iaeking the 24-0Hase enzyme responsible for converting 25(08)D3 to 24R,25(OH)2D3,
suffer from abnormal development of bones formed by intramembraneous ossification,
such as the mandible and ealvaria. These results support the notion that receptors
mediating the action of 24R,2S(OH)2D3 do exist, and also identify the probable sites of
their expression as being intramembraneous bone.
Nuclear and cytosol extraets bave been isolated from the mandible and calvaria of
17.5 day-old mouse embryos. Saturation analysis on these have identified saturable
binding of eHl-24R,25(OH)2D3 to a protein in the extracts. This binding is of high
affinity <Kcs = 1.1nM) and has a Bmu of 2.79pmoUmg proteine Competition assays
showed that this binding protein was specifie for 24R,25(OH)2D3 and was not the
Vitamin D receptor (VDR). Furthermore, sucrose gradient sedimentation assays were
perfonned to eliminate the possibility tbat this protein was the vitamin D binding protein
(DBP), which is a1so known to bind 24R,25(08)2D3. Finally, tissue specifieity
experiments earried out on Uver, brain and bone nuclear extraets revealed that this
putative receptor was also present in liver but not in brain.
70

•
•
•
Two strategies have been employed to isolate the receptor cDNA. Firstly, a hybrid
protein consisting of the GAIA DNA-binding domain fused to the ligand-binding domain
of buman RXRa was used to screen a one-week-old mouse calvaria cDNA library and a
E17.5 mouse mandible/calvaria cDNA library, both fused to the GAIA activation domain.
This strategy bas so far pulled out mouse peroxisome proliferator-activated receptor
(mPPARy) from the neonate library. Continued screening of the embryonallibrary will
follow. Secondly, a 5' primer from the library vector and a 3' primer located within the
highly conserved zinc-finger region of the VDR have been used to screen the embryonal
library by PCR for inserts containing a similar motif. To date no positive clones have
been isolated.
The substantial evidence presented here for the existence of a putative nuclear
receptor for 24R,2S(OHhDJ indicates that continued efforts to isolate the receptor, both
by these methods and by altemate strategies, should result in its successful cloning. The
isolation of such a receptor would provide valuable insigbt into the mechanism of action
of the vitamin D metabolite and its involvement in bone development.
71

•
•
•
Xe ORIGINAL CONTRIBUTION TO KNOWLEDGE
The answer to the question of whetber 24R,25-dihydroxyvitamin OJ is an active
vitamin D3 metabolite or an inactive eatabolite bas long been sought. The bypotbesis tbat
there may exist a receptor for this metaboüte bas been the basis of many studies in the
past two decades. This is the flI'St report to show specifie binding of 24R,2S
dihydroxyvitamin 03 in intramembranous bene during embryonic development. It is also
the flCSt time anti-actin antibodies have been used in sucrose gradient sedimentation
experiments to show that observed ligand binding was not due to DBP complexed witb
endogenous actin. Furthermore, the mRNA that we have isolated from embryonic mouse
mandible and calvaria as weIl as the cONA libraries that we have generated are the fmt
of their kind and provide strong tools with which furtber eloning attempts cao he carried
out.
72

•
•
•
XI. REFERENCES
1. Horst R, Reinhardt T. 1997. Vitamin D Metabolisme In: Feldman D, Glorieux F,Pilee J. (eds). Vitamin D. Academie Press, San Diego, pp 13-31.
2. HartweU D, Hassager C, Christiansen C. 1987. Effeet of vitamin O2 and vitamin 03on the serom concentrations of 1,25(OHh~ and 1,25(OHhD3 in normal subjects.Acta. Endoc. 115: 378-384.
3. HoUis B, Pinard W. 1984. Relative concentrations of 25-hydroxyvitamin D2103 and1,25-dihydroxyvitamin 02103 in maternal plasma at delivery. Nutr. Res. 4: 27-32.
4. Jones 0, Schnoes H, DeLuca H. 1975. Isolation and idenûfication of 1,25dihydroxyvitamin 02. Biochemistry. 14: 1250-1256.
5. Holick Mt MacLaughlin J, Clark M, Holick S, Potts J. Jr, Anderson R, Blank 1,Parrish Jt Elias P. 1980. Photosynthesis of previtamin 03 in human skin andphysiologie consequences. Science. 210: 203-205.
6. Holick M. 1994. Vitamin D: biosynthesis, metabolism, and mode of action. In:DeGroot L, et al. (OOs). Endocrinology (3rd edn). Gnme & Stratton, New York, pp990-1014.
7. Cooke N, and Haddad J. 1989. Vitamin 0 binding protein (Gc-Globulin).Endocrine Reviews. Vol. 10, No. 3: 294-307.
8. Bouillon R, Van Baelen H, DeMoor P. 1980. Comparative study of the affinity ofthe serum vitamin D-binding proteine J. Steroid Biochem. 13: 1029-1034.
9. HoUis B. 1984. Comparison of equilibrium and disequilibrium assay conditions forergocalciferol, cholecalciferol and their major metabolites. J. Steroid Biochem. Vol.21, No. 1: 81-86.
10. Bishop J, Collins E, Okamura W t Norman A. 1994. Profile of ligand specificity ofthe vitamin D binding protein for la,25-dihydroxyvitamin D3 and its analogs.Journal of Bone and Mineral Research. Vol. 9, No. 8: 1277-1288.
Il. Kato A, Seo E-G, Einhorn T, Bishop J, Norman A. 1998. Studies on 24R,25dihydroxyvitamin 03: evidence for a non-nuclear membrane receptor in the chicktibial fracture-healing callus. Bone. Vol. 23, No. 2: 141-146.
73

•
•
•
12. Hollis B, Burton J, Draper H. 1977. A binding assay for 2S-hydroxycalciferols and24R,25-dihydroxycalciferols using bovine plasma globuline Steroids. Vol. 30, No.2: 285-293.
13. Broadus A, Horst R, Lang R, Littledike E, Rasmussen H. 1980. The importance ofcirculating 1,25-dihydroxyvitamin D in the pathogenesis of hypercalciuria and reoalstone formation in primary hyperparatbyroidism. N. Eogl. J. Med. 302: 421-426.
14. Seino Y, Tanaka H. 1997. Vitamin D Metabolites and bone. In: Feldman D,Glorieux F, Pilee J. (eds). Vitamin D. Academic Press, San Diego, pp 305-311.
15. DeLuca H. 1997. Historical overview. In: Feldman D, Glorieux F, Pite J. (eds).Vitamin O. Academie Press, San Diego, pp 3-11.
16. Heaney R. 1997. Vitamin D: role in the calcium eeonomy.ln: Feldman D, GlorieuxF, Pike J. (eds). Vitamin D. Academic Press, San Diego, pp 485-497.
17. Armbrecht H, Wongsurawat N, Zenser T, Davis B. 1984. Effect of PI'H and 1,25(08h03 on renal2S(OH)D3 metabolislll, adenylate cyclase, and protein kinase. Am.J. Physiol. 246: EI02-EI07.
18. Omdahl J, May B. 1997. The 25-hydroxyvitamin D 24-hydroxylase. In: Feldman D,Glorieux F, Pilee J. (eds). Vitamin D. Academie Press, San Diego, pp 69-85.
19. Baron R. 1990. Anatomy and ultrastnlcture of bone. In: Faws M. (ed). Primer onthe metabolic bone diseases and disorders of minerai metabolism (1st ed). WilliamByrd Press, Richmond, pp 3-7.
20. St.-Arnaud R, Glorieux F. 1997. Vitamin D and bone development. In: Feldman D,Glorieux F, Pilee J. (eds). Vitamin D. Academie Press, San Diego, pp 293-303.
21. Hollenberg S, Weinberger C, Oog E, Cerelli G, Oro A, Lebo R, Thompson E,Rosenfeld M, Evans R. 1985. Primary stnlcture and expression of a functionalhuman glucocorticoid receptor cDNA. Nature. 318: 635-641.
22. Mangelsdorf D, Thummel C, Beato M, Herrlich P, Scbutz 0, Umesono K, BlumbergB, Kastner P, Mark M, Chambon P, Evans R. 1995. The nuclear receptorsuperfamily: the second decade. Cell. 83: 835-839.
23. MangelsdorfD, Evans R. 1995. The RXR beterodimers and orphan receptors. Cell.83: 841-850.
24. Jin C, Kerner S, Hong M, Pilee W. 1996. Transcriptional activation anddimerization functions in the human vitamin D receptor. Molec. Endoc. Vol. 10,No. 8: 945-957.
74

•
•
•
25. Carlberg C. 1996. The vitamin D reœptor in the context of the nuclear receptorsuperfamily. Endocrine. Vol. 4, No. 2: 91-105.
26. Pike W. 1997. The vitamin D reœptor and its gene. In: Feldman D, Glorieux F,Pike J. (eds). Vitamin D. Academic Press, San Diego, pp 105-125.
27. Holick MF. 1995. Vitamin D: photobiology, metabolism, and clinical :pplications.ln: DeGroot L, Besser H, Burger HG. et al. (eds). Endocrinolgy (3 edn). WBSaunders, Philadelphia. pp 990-1013.
28. Reichel H, Koeffler HP, Norman AW. 1989. The role of the vitamin D endocrinesystem in health and disease. N. Engl. J. Med. 320: 981-991.
28a. Norman A. 1997. Rapid biologjcal responses mediated by la,25-dihydroxyvitaminD3: A case study of transcaltaehia (rapid hormonal stimulation of intestinal calciumtransport. In: Feldman D, Glorieux F, Pike J. (eds). Vitamin D. Academic Press,San Diego, pp 233-256.
28b.Nemere l, Donnanen MC, Hammond MW, Okamura WH, Nonnan AW. 1994.Identification of a specific binding protein for la,25-dihydroxyvitamin D3 in basallateral membranes of chick intestinal epithelium and relationship to transcaltachia J.Biol. Chem. 269: 23750-23756.
29. Miller S, Halloran B, DeLuca H, Yamada S, Takayama H, Jee W. 1981. Studies onthe role of 24-hy::!roxylation of vitamin D in the mineralization of cartilage and boneof vitamin D-deficient rats. Calcif. Tissue Int. 33: 489-497.
30. Parfitt A, Mathews C, Brommage R, Jamagjn K, DeLuca H. 1984. Calcitriol but noother Metabolites of vitamin D is essential for normal bone growth and developmentin the rat. J. Clin. Invest. 73: 576-586.
31. Henry H, Norman A. 1978. Vitamin D: two dihydroxylated Metabolites are requiredfor normal chicken egg hatchability. Science. 201(4358): 835-837.
32. Sunde M, Turk C, DeLuca H. 1978. The essentiality of vitamin D Metabolites forembryonic chick development. Science. 200: 1067-1069.
33. Boyan B, Dean D, Sylvia V, Schwartz Z. 1997. Cartilage and vitamin D: genomicand nongenomic regulation by 1,25(0H)2D3 and 24,25(OH)2D3. In: Feldman D,Glorieux F, Pike J. (eds). Vitamin D. Academic Press, San Diego, pp 395-421.
75

•
•
•
34. Corvol M, Dumontier M, Garabedian M, Rappaport R. 1978. Vitamin D andcartilage. n. Biological activity of 2S-bydroxycbolecalciferol and 24,25- and 1,25dihydroxycholecalciferols on cuItured growth plate chondrocytes. Endocrinology.Vol. 102, No. 4: 1269-1274.
35. Omoy A, Goodwin D, Noff D, Edelstein S. 1978. 24,25-dihydroxyvitamin 0 is ametabolite ofvitamin 0 essential for bone formation. Nature. 276: 517-519.
36. Seo E-G, Einhorn T, Norman A. 1997. 24R,25-dihydroxyvitamin D3: an essentialvitamin 03 metabolite for both nonnal bone integrity and healing of tibial fracture inchicks. Endocrinology. Vol. 138, No. 9: 3864-3872.
37. Seo E-G, Norman A. 1997. Three-fold induction ofrenal25-hydroxyvitamin D3-24hydroxylase activity and increased serum 24,2S-dihydroxyvitamin D3 levels arecorrelated with the healing process after chick tibial fracture. J. Bone Min. Res.Vol. 12, No. 4: 598-606.
38. Schwartz Z, Soskolne W, Atkin 1, Goldstein M, Omoy A. 1989. A direct effect of24,25-(OH)2D3 on the modeling of fetal mice long bones in vitro. J. Bone Min Res.Vol. 4, No. 2: 157-163.
39. St.-Arnaud R, Glorieux F. 1998. Editorial: 24,25-dihydroxyvitamin 0 - activemetabolite or inactive catabolite? Endocrinology. Vol. 139, No. 8: 3371-3374.
40. Miller S, Halloran B, DeLuca H, Lee W. 1982. Role of vitamin D in maternaiskeletal changes during pregnancy and lactation: a histomorphometric study. Calcif.Tiss. Int. 35: 455-460.
41. Nakamura T, Kurokawa T, Orimo H. 1988. Increased bone volume in vitamin Dreplete rat femur by massive administration of 24R,25(OH)2D3. Calcif. Tiss. Int. 43:235-243.
42. Nakamura T, Suzukï K, Hirai T, Kurokawa T, Orimo H. 1992. Increased bonevolume and reduced bone turnover in vitamin D-replete rabbits by the administrationof 24R,25-dihydroxvitamin 03. Bone. 13: 229-236.
43. Yamaura M, Nakamura T, Nagai y, Yoshihara A, Suzuki K. 1993. Reducedmechanical competence of bone by ovariectomy and its preservation with 24R,25dihydroxyvitamin D3 administration in beagles. Calcif. Tiss. Int. 52: 49-56.
44. Nakamura T, Nagai y, Yamato H, Suzuki K, Orimo H. 1992. Regulation of boneturnover and prevention of bone atrophy in ovariectomized beagle dogs by theadministration of 24R,2S(OH)2D3. Calcif. Tiss. lot. SO: 221-227.
76

•
•
•
45. Yamate T, Tanaka H, Nagai Y, Yamato H, Taniguchi N, Nakamura T, Seino Y.1994. Bone-fonning ability of 24R,25-dihydroxyvitamin 1>] in thehypophospbatemic mouse. J. Bone. Min. Res. Vol. 9, No. 12: 1967-1974.
46. Ono T, Tanaka H, Yamate T, Nagai Y, Nakamura T, Seino Y. 1996. 24R25dihydroxyvitamin D3 promotes bone formation witbout causing excessive resorptionin hypophospbatemic mïce. Endocrinology. Vol. 137, No. 6: 2633-2637.
47. Fine N, Bindennan 1, Somjen D, Earon Y, Edelstein S, Weisman Y. 1985.Autoradiographie localization of 24R,25-dibydroxyvitamin D3 in epiphysealcartilage. Bane. 6: 99-104.
48. Schwartz Z, Schlader D, Swain L, Boyan B. 1988. Direct effects of 1,25dihydroxyvitamin D3 and 24,25-dibydroxyvitamin D3 on growtb zone and œstingzone chondrocyte membrane alkaline phosphatase and phospholipase-A2 specificactivities. Endocrinology. Vol. 123, No. 6: 2878-2884.
49. Schwartz Z, Boyan B. 1988. The effects of vitamin D metabolites on phospholipaseA2 activity of growth zone and resting zone cartilage ceUs in vitro. Endocrinology.Vol. 122, No. 5: 2191-2198.
50. Schwartz Z, Schlader D, Ramirez V, Kennedy M, Boyan B. 1989. Effects ofvitamin D Metabolites on collagen production and cell proliferation of growth zoneand resting zone cartilage cells in vitro. J. Bane Min. Res. Vol. 4, No. 2: 199-207.
51. Schwartz Z, Dean D, Walton J, Brooks B, Boyan B. 1995. Treatment of reslingzone chondrocytes witb 24,2S--dihydroxyvitamin D3 [24,25-(OHhil]] inducesdifferentiation inlo a 1,25-(OHhD3-re5PQnsive phenotype characteristic of growthzone chondrocytes. Endocrinology. Vol. 136, No. 2: 402-411.
52. Somjen D, Kaye A, Binderman 1. 1984. 24R,25-dihydroxyvitamin D stimulatescreatine kinase BB activity in chick cartilage ceUs in culture. FEBS. Vol. 167, No.2: 281-284.
53. Somjen D, Weisman Y, Bindennan 1, Kaye A. 1984. Stimulation of creatine kinaseBB activity by la, 25-dihydroxycholecalciferol and 24R,25-dihydroxycholecalciferol in rat tissues. Biochem. J. 219: 1037-1041.
54. Yamato H, Okazaki R, Ishii T, Ogata E, Sato Tt Kumegawa M, Akaogi K, TaniguchiN, Matsumoto T. 1993. Effect of 24R,25-dihydroxyvitamin D3 on the formation andfunction of osteoclastic ceUs. Calcif. Tissue. lot. 52: 255-260.
55. Corvol M, UJmann ~ Garabedian M. 1980. Specifie nuclear uptake of 24,25dihydroxycholecalciferol, a vitamin D3 Metabolite biologically active in cartilage.FEBS Letters. Vol. 116, No. 2: 273-276.
77

•
•
•
56. Somjen D, Somjen G, Weisman Y, Bindennan I. 1982. Evidence for 24,25dihydroxycbolecaIciferol receptors in long bones of newbom rats. Biocbem. J. 204:31-36.
57. Balmain N, Haucbecome M, Pite W, Cuisinier-Gleizes P, Madieu H. 1993.Distribution and subcellular immunolocalization of 24,25-dihydroxyvitamin D3receptors in rat epiphyseal cartilage. Cell. Mol. Biol. 39: 339-350.
58. Somjen D, Somjen 0, HareU A, Mecbanic 0, Bindennan I. 1982. Partialcharacterization of a specific high affinity binding macromolecule for 24R,25dihydroxyvitamin D3 in differentiating skeletal mesencbyme. Biochem. Biophys.Res. Corn. Vol. 126, No. 2: 644-651.
59. Merke J, Norman A. 1981. Studies on the mode of action of calciferol xx:xn.Evidence for a 24(R),2S(OH)2-vitamin 03 receptor in the paratbyroid gland of therachitic chick. Biochem. Biophys. Res. Corn. Vol. 100, No. 2: 551-558.
60. Seol W, Choi H-S, Moore D. 1995. Isolation of proteins that interact specificallywith the retinoid X receptor: two novel orpban receptors. Molec. Endoc. 9: 72-85.
61. Seol W, Choi H-S, Moore D. 1996. An orphan nuclear hormone receptor that lacksa DNA binding domain and heterodimerizes with other receptors. Science. 272:1336-1339.
62. Miyata K, McCaw S, Patel H, Racbubinski R, Capone J. 1996. The orphan nuclearhormone receptor LXRa interacts with the peroxisome proliferator-activatedreceptor and inhibits peroxisome proliferator signaling. J. Biol. Cbem. Vol. 271,No. 16: 9189-9192.
63. Roy R, Gosselin Pt Guerin S. 1991. A short protocol for micro purification ofnuclear proteins from whole animal tissue. Biotechniques Il: 770-777
64. Somjen Dt Binderman It Weisman Y. 1983. The effect of 24R2S-dihydroxycholecalciferol and of la,25-dihydroxycbolecalciferol on ornithinedecarboxylase activity and on DNA sYDthesis in the epiphysis and diaphysis of ratbone and in the duodenum. BiocbemJ. 214: 293-298.
65. Stem P, DeLuca HF, Ikekawa N. 1975. Bone resorbing activities of 24-byroxystereoisomers of 24-hydroxyvitamin D3 and 24,25-dihydroxYvitamin OJ. BiocbemBiophys Res Commun. 67: 965-971.
66. Pierce E, Dame M, Bouillon R, Van Baelen Ht DeLuca H. 1985. Monoclonalantibodies to buman vitamin D-binding protein. Proc. Nad. Acad. Sci. USA. 82:8429-8433.
78

•
•
•
67. Wecksler W and Norman A. 1979. An hydroxylapatite batch assay for thequantitation of la2S-dihydroxyvitamin D3 - receptor complexes. AnalyticalBiochemistry 92: 314-323.
68. Van Baelen H, Bouillon R, De Moor P. 1980. Vitamin D-binding protein (GcGlobulin) binds actin. The Journal of Biological Chemistry. Vol. 255, No. 6: 22702272.
69. Reinhardt T, Horst R, Littledike T, Beitz D. 1982. 1,25-dihydroxyvitamin D3
receptor in bovine thymus gland. Biochemical and Biophysical ResearchCommunications. Vol. 106, No. 3: 1012-1018.
70. Freedman L, Lemon B. 1997. StnJctural and functional determinants of ONAbinding and dimerization by the vitamin D receptor. In: Feldman D, Glorieux F, PileeJ. (eds). Vitamin D. Academic Press, San Diego, pp 127-148.
71. Sone T, Kerner S, Pilee J. 1991. Vitamin D receptor interaction with specifie DNA.Association as a 1,25-dihydroxyvitamin D3-modulated heterodimer. J. Biol. Chem.266: 23296-23305.
72. Yu V, Delsert C, Andersen B, Holloway l, Devary 0, Naar A, Kim S, Boutin J,Glass C, Rosenfeld M. 1991. RXR beta: a eoregulator that enhances binding ofretinoic acid, thyroid hormone, and vitamin 0 receptors to their cognate responseelements. Cell. 67: 1251-1266.
73. Klie~er S, Umesono K, Mangelsdorf D, Evans R. 1992. Retinoid X receptorinteracts with nuclear receptors in retinoie acid, thyroid receptor and vitamin D3signaling. Nature. 355: 446-449.
74. Miyata K, McCaw S, Marcus S, Raebubinski R, Capone J. 1994. The peroxisomeproliferator-activated receptor interacts with the retinoid X receptor in vivo. Gene.148: 327-330.
75. Endo H, Kiyoki M, Kawashima K, Naruchi T, Hashimoto Y. 1980. Vitamin 0 3Metabolites and PTH synergistically stimulate bone formation of chick embryonicfemur in vitro. Nature. 286: 262-264.
76. St.-Arnaud R, Arabian A, Glorieux F. 1996. Abnonnal bone development in micedeficient for the vitamin D 24-hydroxylase gene. J. Bone. Min. Res. Il: S 126(abstract).
77. Adams J, Ren S-Y, Arbelle J, Horiuchi N, Gray R, Clemens T, Shany S. 1994.ReguJated production and intraerine action of 1,25-dihydroxyvitamin D3 in the ehickmyelomonocytic cellline HD-l1. Endocrinology. Vol. 134, No. 6: 2567-2573.
79

•
•
•
78. Demura T, Obashi N, Nonomura~ Mori T, Mamiya M, Seki H, Enami N, KoyanagiT. 1992. [Localization of angrogen receptor in male sex organs, accessory sexorgans and extemal geDitai skin]. [Japanese]. Nippon Hinyokika Gakkai Zasshi Japanese Journal of Urology. Vol. 83, No. 12: 2078-2084.
79. Adashi E, Hsueh A. 1981. Autoregulation of androgen production in a primaryculture of rat te5ticular ceUs. Nature. 293: 737-738.
80. Vomberger W, Prins G, Musto N, Suarez-Quian C. 1994. Androgen receptordistribution in rat testis: new implications for androgen regulation ofspermatogenesis. Endocrinology. Vol. 34, No. 5: 2307-2316.
81. Duffy D, Hess D, Stouffer R. 1994. Aeute administration of a 3p-hydroxysteroiddehydrogenase inhibitor to rhesus monkeys at the midIuteal phase of the menstnJalcycle: evidence for possible autocrine regulation of the primate corpus luteum byprogesterone. J. Clin. Endoc. and Metab. Vol. 79, No. 6: 1587-1594.
82. Hild-Petito S, Stouffer R, Brenner R. 1988. Immunocytocbemical localization ofestradiol and progesterone receptors in the monkey ovary tbroughout the menstnJalcycle. Endocrinology. Vol. 123, No. 6: 2896-2905.
83. Pratt W. 1993. The role of beat shock proteins in regulating the funetion, foldingand traffieking of the glucocortieoid receptor. J. Biol. Chem. Vol. 268, No. 29:21455-21458.
84. Pratt W. 1990. Interaction of hsp90 with steroid receptors: organizing some diverseobservations and presenting the newest concepts. Mol. Cel. Endoc. 74: C69-C76.
85. Smith D, Toft D. 1993. Steroid receptors and their associated proteins. MolecularEndocrinology. Vol. 7, No. 1: 4-11.
86. Dalman F, Sturzenbecker L, Levin A, Lucas D, Perdew 0, Petkovitch M, ChambonP, Grippo J, Pratt W. 1991. Retinoic acid receptor belongs to a subclass of nuclearreceptors that do not fonn "docking" complexes with hsp90. Biochemistry. 30:5605-5608.
87. Dalman F, Koenig R, Perdew 0, Massa E, Pratt W. 1990. In contrast to theglucocorticoid receptor, the thyroid hormone receptor is translated in the DNAbinding state and is not associated with hsp90. 1. Biol. Chem. Vol. 265, No. 7:3615-3618.
88. Bresnick E, Dalman F, Sanchez E, Pratt W. 1989. Evidence that the 9O-kDa heatshock protein is necessary for the steroid binding conformation of the L cellglucocorticoid receptor. J. Biol. Chem. Vol. 264, No. 9: 4992-4997.
80

•
•
•
89. Van Baelen H, Bouillon R, De Moor P. 1977. Binding of 25-hydroxycholecalciferolin tissues. J. Biol. Chem. Vol. 252, No. 8: 2515-2518.
90. Safadi F, Thomton P, Magiera Ht Hollis B, Oentile M, Hadda J, Liebhaber S, CookeN. 1999. Osteopathyand resistance to vitamin D toxicity in mice null for vitamin Dbinding protein. J. Clin. Iov. Vol. 103, No. 2: 239-251.
91. linknown. 1997. Methods and Reagents: False positives from the yeast two-hybridsystem. Computer Corner, TIBS 22: 33.
92. Allen J, Walberg M, Edwards Mt Elledge S. 1995. Finding prospective partners inthe library: the two-hybrid system and phage display fmd a match. TIBS. 20: 511516.
93. Chien Ct Banel P, Stemglanz~ Fields S. 1991. The two-hybrid system: A methodto identify and clone genes for proteins that interact with a protein of interest. Proc.Nat!. Acad. Sci. USA. 88: 9578-9582.
94. Mahonen A, Pirskanen A, Keinanen R, Maenpaa PH. 1990. Effect of 1,25(OH)2D3on its receptor m.RNA levels and osteocalcin synthesis in human osteosarcoma ceUs.Biochim. Biophys. Acta. 1048: 30-37.
95. Forman B, Goode E, Chen J, Oro A, Bradley D, Perlmann T, Noonan D, Burka L,McMorris T, Lamph W, Evans R, Weinberger C. 1995. Identification of a nuclearreceptor that is activated by famesol Metabolites. Cell. 81: 687-693.
96. Issemann l, Green S. 1990. Activation of a member of the steroid hormone receptorsuperfamily by peroxisome proliferators. Nature. 347: 645-650.
97. Li J, Herskowitz I. 1993. Isolation of ORC6, a component of the yeast originrecognition complex by a one-hybrid system. Science. 262: 1870-1874.
98. Bush S, Folla St Lannigan D. 1996. Use of the yeast one-hybrid system to screenfor mutations in the Iigand-binding domain of the estrogen receptor. Steroids. 61:102-109.
99. Mar P, Kumar A, Kang D-e, Zhao B, Martinez L, Montgomery R, Anderson L,Butler A. 1995. Characterization of novel phorbol ester- and serom-responsivesequences of the rat omithine decarboxylase gene promoter. MolecularCarcinogenesis. 14: 240-250.
100. Ritchie M, Trask R, Fontanet H, Billadello J. 1991. Multiple positive and negativeelements regulate human brain creatine kinase gene expression. Nuc. Acids Res.Vol. 19, No. 22: 6231-6240.
81

•
•
•
101. Ritehie M. 1996. Characterization of human B creatine kinase gene regulation inthe heart in vitro and;n v;vo. J. Biol. Chem. Vol. 271, No. 41: 25485-25491.
102. Somjen D, Weisman y, Berger E, Baron y, Kaye A, Bindennan 1. 1986.Developmental changes in the responsiveness of rat kidney to vitamin D Metabolites.Endocrinology. Vol. 118, No. 1: 354-359.
103. Somjen D, Earon y, Harell S, Shimshoni Z, Weisman y, Harell A, Kaye A,Bindennan I. 1987. Developmental changes in responsiveness to vitamin Dmetabolites. J. Steroid Biochem. Vol. 27, No. 4-6: 807-813.
82