modification of the brain proteome of africanized honeybees (apis mellifera) exposed to a...
TRANSCRIPT

Modification of the brain proteome of Africanized honeybees (Apismellifera) exposed to a sub-lethal doses of the insecticide fipronil
T. C. Roat • J. R. A. dos Santos-Pinto •
L. D. dos Santos • K. S. Santos • O. Malaspina •
M. S. Palma
Accepted: 6 August 2014
� Springer Science+Business Media New York 2014
Abstract Fipronil is a phenylpyrazole insecticide that is
widely used in Brazilian agriculture for pest control.
Although honeybees are not targets of fipronil, studies
indicate that this pesticide can be harmful to honeybees. To
assess the effects of fipronil in the brain of Africanized
Apis mellifera workers, this study focused on the toxico-
proteome profiling of the brain of newly emerged and aged
honeybee workers that were exposed to a sub-lethal dose
(10 pg fipronil per day. i.e. 1/100 of LD50/bee/day during
5 days) of the insecticide. Proteomic analysis identified 25
proteins that were differentially up-regulated or down-
regulated when the fipronil-exposed and non-exposed
groups were compared. These proteins are potentially
related to pathogen susceptibility, neuronal chemical stress,
neuronal protein misfolding, and occurrence of apoptosis,
ischemia, visual impairment, damaged synapse formation,
brain degeneration, memory and learning impairment. The
exposure of honeybees to a very low dose of fipronil, even
for a short period of time (5 days), was sufficient to cause a
series of important neuroproteomic changes in the brains of
honeybees.
Keywords Brain proteome � Phenylpyrazole �Insecticide � Proteomic analysis � 2-DE � Mass
spectrometry
Introduction
Honeybees Apis mellifera L., 1758 (Hymenoptera: Apidae)
are considered excellent pollinating insects in agroecosys-
tems (Kremen et al. 2007) giving to this specie a great
agronomic, environmental and economic importance.
However the reduction in the number of pollinators,
especially bee species, has become a major problem for
beekeepers, and it may threat food security and ecosystem
integrity due to reduced pollination. The causes of this high
mortality are not well understood. Many stress factors,
acting alone or together, can weaken the colony including
the decreased availability of resources due to the degra-
dation and fragmentation of the honeybee habitat, patho-
gens and parasites, beekeeping management, and the
extensive use of chemical pesticides on crops (vanEn-
gelsdorp and Meixner 2009; Bromensshenk et al. 2010;
Wratten et al. 2012).
Honeybees can be exposed to insecticides during their
foraging activities and while carrying out activities within
the colony through contact with contaminated pollen and
nectar. Consequently, different larval stages and adults of
different ages can be exposed to these chemical products
(Pham-Delegue et al. 2002; Malaspina and Silva-Zacarin
2006; Desneux et al. 2007). There are different viewpoints
in the discussion about the exposure of honeybees to dif-
ferent levels of insecticide residues and whether this
T. C. Roat (&) � J. R. A. dos Santos-Pinto � O. Malaspina �M. S. Palma
Institute of Biosciences of Rio Claro, Department of Biology,
Center of the Study of Social Insects, University of Sao Paulo
State (UNESP), Avenida 24-A, N.1515-Bela Vista, Rio Claro,
SP, Brazil
e-mail: [email protected]
L. D. dos Santos
Center for the Study of Venoms and Venomous Animals
(CEVAP), University of Sao Paulo State (UNESP), Botucatu,
SP, Brazil
K. S. Santos
Discipline of Allergy and Immunology (HC/Incor/FMUSP),
Sao Paulo, SP, Brazil
123
Ecotoxicology
DOI 10.1007/s10646-014-1305-8

exposure directly causes honeybee death (Greig-Smith
et al. 1994; Rortais et al. 2005; Rondeau et al. 2014).
The fipronil is the active component of different agricul-
tural products (Tingle et al. 2003). Fipronil belongs to the
phenylpyrazole group and acts on the nervous system of
insects by blocking GABA receptors and inhibiting ionotro-
pic glutamate-gated chloride channels, inhibiting the central
nervous system (El Hassani et al. 2005). GABA receptors are
located in several neuropils of the honeybee brain (Schafer
and Bicker 1986), and they are involved in modulating
learning, memory and sensory perception (El Hassani et al.
2005; Aliouane et al. 2009; Bernadou et al. 2009).
Studies assessing the toxicity of fipronil to honeybees
reported that it is harmful to these insects (Decourtye et al.
2005; Pochi et al. 2012). It can modulate insects’ sensi-
tivity to sucrose (Aliouane et al. 2009), affect learning
(Decourtye et al. 2005), orientation and foraging behavior
(Decourtye et al. 2009), and cause hyperactivity followed
by paralysis and death (Barbara et al. 2005; Gunasekara
et al. 2007). The results from studies of the oral toxicity of
fipronil against the stingless bee Melipona scutellaris
showed that these insects exhibited signs of intoxication,
such as initial tremors followed by paralysis, before their
death (Lourenco et al. 2012). It was reported that low doses
of fipronil impaired the olfactory learning (0.5 ng/bee) and
memory abilities (0.1 ng/bee) in A. mellifera through the
action of multiple targets, including glutamatergic and
GABAergic inhibitory signaling (El Hassani et al. 2005)
Roat et al. (2013) reported the lethal dose (LD50) and
lethal concentration (LC50) values of fipronil for newly
emerged Africanized honeybees showing that this insecti-
cide may be harmful to these bees independently of exposure
route: LD50 = 1.06 ng fipronil/bee and LC50 = 1.27 ng
fipronil/lL of food, respectively. Besides, this study assessed
the side effect of a sublethal dose of fipronil on neuron
metabolic activity from Africanized honeybees and showed
that the insecticide causes metabolic changes by increasing
the respiratory activity of mitochondria.
A series of discussions about the use and restrictions of
different classes of pesticides has been reported in the lit-
erature; however, despite the fact that many studies have
been published, there is still a lack of knowledge on the
relationship between the exposure to insecticides and the
possible effects on the honeybee physiology (Maxim and
van der Sluijs 2007). Studies performed using neonicoti-
noids already showed effects of sublethal doses on hon-
eybee behaviors, e.g. antifeedant effect (Han et al. 2010a),
and reduced visual learning capacities and decreased
olfactory learning performances (Decourtye et al. 2009;
Han et al. 2010b).
The most studies that were performed with phenylpy-
razoles were conducted by evaluating acute toxicity, i.e.,
determining the LD50 values for these pesticides (OEPP/
EPPO 2003). The assessment of risk to honeybee popula-
tions using this approach has been contested because the
pesticides are generally used at sub-lethal doses (Halm
et al. 2006), which would require the evaluation of chronic
toxicity caused by continued use of low doses of pesticide
(Rortais et al. 2005); there is a need of more accurate
methods for risk assessment of pesticides on bees (De-
courtye et al. 2013).
Because fipronil is widely used in Brazilian agriculture,
we used it as a pesticide model in a proteomic approach
aiming to analyze the biochemical effects of sublethal
doses in the brains of Africanized A. mellifera. We
developed a toxicoproteomic approach using an experi-
mental protocol; it combined 2-DE gel electrophoresis with
in-gel protein digestion, followed by mass spectrometry
analysis to assess the effects of fipronil by profiling the
protein expression pattern in the brains of newly emerged
and aged workers.
Materials and methods
Honeybees
Honeybees were obtained from queen-right colonies of
Africanized A. mellifera at the experimental apiary of the
Institute of Biosciences of Rio Claro, Department of
Biology, University of Sao Paulo State (UNESP), Rio
Claro, SP, Brazil. To obtain newly emerged workers, three
sealed brood combs from three different colonies were
placed in a climatic room at a temperature of 34 ± 2 �C
and a relative humidity (RH) of 70 ± 10 %, and the adults
were collected after emergence. Of the 740 newly emerged
honeybees that were collected, 240 were used in bioassays
and 500 were marked with ink on their thorax and were
returned to the colony for capture when they reached
20 days of age (considered here as aged honeybees).
Honeybee toxicity assays
The honeybee toxicity assay was performed using a sub-
lethal dose of fipronil that was based on the LD50 value for
newly emerged honeybees, which was determined by Roat
et al. (2013) to be 1.06 ng fipronil per bee. Fipronil
(95.0 %, analytical grade) was supplied by Dalian Raiser
Pesticides (China). To intoxicate the honeybees, a stock
solution of fipronil (1,000 ng/lL) was prepared in acetone,
and from this solution, several dilutions were prepared and
applied to a syrup preparation (50 % m/v sucrose) to obtain
a solution containing 1 pg fipronil per lL diet and
0.0001 % of acetone. Groups of 60 newly emerged and 60
aged honeybees (from three different colonies-20 of each
colony) were equally divided into three disposable cages
T. C. Roat et al.
123

(20 workers per cage). The honeybees were collectively fed
with the sucrose syrup containing fipronil, and the total
volume was adjusted to ensure that each honeybee could
consume approximately 10 lL sucrose solution daily.
Therefore, each bee ingested 10 pg fipronil per day (1/100 of
LD50), during 5 days. After ingestion of the contaminated
syrup, the bees were fed with syrup (the same that was
furnnihed for the control group). The experimental control
used in these experiments consists of honeybees that were
fed with syrup. The food was provided in a small plastic
cup that was placed on the bottom of the experimental
cages and covered with a plastic mesh. Water was supplied
ad libitum through impregnated cotton. The cages were
kept in a climatic room at 32 ± 2 �C and 60 ± 10 % RH.
Samples of newly emerged and aged honeybees were
collected to assess the alterations that were induced by
fipronil 5 days after the beginning of exposure to fipronil.
Thus, newly emerged bees were 5 days old at the time of
collection and aged bees were 25 days old.
Protein assay
For the analysis of the protein fraction, the brains of 30
bees derived from each treatment were homogenized in
1 mM PMSF (phenylmethylsulphonyl fluoride) in dis-
solved bi-distilled water (containing 1 % v/v isopropanol).
Protein quantification was determined by the Bradford
method (Bradford 1976) using bovine serum albumin
(BSA) as a standard.
Two-dimensional gel electrophoresis
Samples (250 lg proteins) were applied by rehydration to
7 cm IPG strips, pH 3–10. Isoelectric focusing (IEF) was
carried out on an IPGphor system (GE Healthcare) at
3,500 V for 7,100 Vh. Immobilized pH Gradient (IPG)
strips were incubated in equilibration buffer [50 mM Tris–
HCl, pH 8.8, 6 M urea, 30 % (v/v) glycerol, 2 % (w/v) SDS]
containing 0.5 % (w/v) dithiothreitol (DTT) for 15 min,
followed by equilibration buffer containing 4 % (w/v)
iodoacetamide for 15 min. The second dimension was run on
self-cast SDS-PAGE gels [12.5 % (w/v) polyacrylamide and
0.8 % (w/v) bis-(N,N’-methylenebisacrylamide)] at 15 mA/
gel for 15 min and 30 mA/gel for 1 h at 10 �C in a Mini-VE
system (GE Healthcare). The gels were stained overnight
with Coomassie Brilliant Blue R-250 (CBB) and stored at
21 �C in preserving solution [7 % (v/v) acetic acid].
Image acquisition, protein quantitation and statistical
analysis
For each treatment or control group were carried out three
gels, representing a triplicate. 2-DE gels stained with
Coomassie Brilliant Blue (CBB) were scanned and digi-
tized (BioImage, GE Healthcare) in the transparency mode
with 24-bit red-green-blue colors and 400 dpi resolution for
documentation. The gel staining was performed as indi-
cated by the manufacturer of the staining system (GE
Healthcare) as follows: gels were fixed for 30 min in a
solution of 40 % (v/v) methanol containing 7 % (v/v)
acetic acid; the gel was then stained overnight in Coo-
massie (0.02 % PhastGel Blue R, 60 % methanol, 10 % (v/
v) acetic acid). The destaining was conducted by incubat-
ing the gel for 30 min in 40 % methanol containing 7 % (v/
v) acetic acid, and a solution containing 7 % (v/v) acetic
acid and 5 % (v/v) methanol, until the background was
clear. All images were analyzed by ImageMaster 2-DE
Platinum 7.0 software from GE Healthcare (Uppsala,
Sweden) using a white light tray; the f/stop was set 22
(minimal exposure). The integrated optical densities were
determined for the full width of each protein spot and the
parameters for image capture of the CBB stained gel were:
time = 0.28 s, gain = 1, b.lev = 7, and gamma = 0.55;
the duration of exporting the image to a computer was
0.45 s, gain = 0.1, b.lev = 6 and gamma = 1.0. The gel
with the most spots was chosen as a reference. The refer-
ence gel was then used to match the corresponding protein
spots between gels. After an average mode of background
correction, manual subtraction of the ‘‘joined valleys’’ was
performed. The rate values of individual spot volume
intensity and the total volume intensity (sum of the inten-
sity volumes obtained from all spots in the same 2-DE gel)
were calculated. Then, a comparison was made between
the control (honeybees not exposed to fipronil) and the
fipronil-exposed group; thus, the rate values of individual
spot volume intensity were transformed to inter-experi-
mental normalized spot volume intensity (INSVI), and the
values of this rate were used to determine the differential
protein expression between the control and fipronil-
exposed groups, which were considered as up- or down-
regulated when the INSVI values were greater than 1.50
and less than 0.50, respectively. One-way ANOVA was
used for the analysis of the INSVI value differences of the
spots between both groups; a probability of p \ 0.05 was
considered to be statistically significant.
In-gel digestion
Pieces (*1 mm3) were excised from the protein spots of
the 2-DE gel, destained twice for 30 min at 25 �C with
50 mM ammonium bicarbonate/50 % acetonitrile (MeCN),
dehydrated in the presence of 100 lL pure MeCN at 25 �C,
dried in a speed-vac system for 30 min at 38 �C, and
treated with 25 lL trypsin solution (20 lg/mL, Promega,
Madison, USA) in 50 mM ammonium bicarbonate, pH 7.9
at 37 �C for 18 h. Digests were extracted from each gel
Brain proteome of Africanized honeybees
123

piece with 30 lL 50 % (v/v) acetonitrile containing 5 %
(v/v) formic acid, desalted and cleaned with PerfectPure
C18 pipet tips (Eppendorf, Hamburg, Germany) according
to the manufacturer’s instructions, and the desalted digests
were vacuum dried. The concentrated digests were mixed
with 0.5 lL matrix [10 mg/mL a-cyano-4-hydroxycin-
namic acid in methanol/acetonitrile (1:1, v/v) mixed with
an equal volume of 0.2 % (v/v) aqueous trifluoroacetic acid
(TFA)] and spotted onto a MALDI plate target.
Mass spectrometry analysis
Mass spectrometric analysis was performed with a MALDI
ToF/ToF–MS (matrix-assisted laser desorption ionization
time of flight/time of flight-mass spectrometry) instrument
(Shimadzu, Axima Performance). The MS and MS/MS
spectra were acquired in the positive ion reflectron mode
using a N2 laser. Typically, 250–500 laser shots were
acquired for the MS and MS/MS mode, and spectra were
obtained at a laser power that was maximized for the
highest possible resolution and peak intensity in each
analytical condition. CID spectra were acquired using a
dual-timed ion gate at a laser power that was approximately
20 % higher than for MS acquisition. MS data were
acquired in the m/z range of 700–3,500, with an acceler-
ating voltage of 20 kV, delayed extraction, a peak density
of a maximum 50 peaks per 200 Da, a minimal S/N ratio of
10 and a maximum peak at 60. MS/MS data were acquired
in the mass range from 60 Da to each precursor mass with
a minimum S/N ratio of 10; the maximum number of peaks
was set to 65 and a peak density of a maximum of 50 peaks
per 200 Da.
Protein identification
Launchpad 2.8 (Shimadzu) was used to submit the com-
bined MS and MS/MS data to the MASCOT protein search
engine v. 2.2 (http://www.matrixscience.com) for search-
ing against a publicly available A. mellifera protein
sequence database deposited in the National Center for
Biotechnology Information non-redundant protein database
(NCBInr) (http://blast.ncbi.nlm.nih.gov/Blast.cgi). The
following search parameters were used: no restrictions on
protein molecular weight, one tryptic missed cleavage was
allowed, and peptide mass tolerances for the searches were
0.5 Da for MS spectra and 0.3 Da for MS/MS spectra.
Iodoacetamide derivatized cysteine and oxidation of
methionine were specified in MASCOT as fixed and vari-
able modifications, respectively. Scaffold (v. 2.04.00,
Proteome Software Inc., Portland, OR) was used to validate
MS/MS-based peptide and protein identifications. The
False Discovery Rate (FDR) assessment was estimated
using the original decoy FDR approach from Mascot; a
separate decoy database was generated from the protein
sequence database with the decoy.pl Perl script provided by
Matrix Science. This script randomizes each entry, while
retaining the average amino acid composition and length of
the entries. The maximum protein and peptide FDR rates
were set to 0.01 and the maximum peptide FDR to 0.1;
FPR was calculated based on the Mascot score. Proteins
were considered identified when at least two peptides were
assigned to the respective sequence. All peptides and
proteins were also confirmed by manual examination of the
spectra.
Results
To investigate the effects of a sub-lethal doses of fipronil
on the protein expression profile of the honeybee brain,
proteomic profiles of this organ from newly emerged and
aged workers were compared and analyzed without (con-
trol) and with exposure to a dose of 10 pg fipronil/day for
5 days; under these conditions no death of bees was
observed. The electrophoretic patterns of samples from
three 2-DE gels were analyzed for each experimental
condition. The patterns were highly similar, as reflected by
the high scatter plot correlation coefficient ([98 %)
between the three gels. Image analysis revealed 96 protein
spots in newly emerged brains from the control group and
the fipronil-treated group (Fig. 1a, b respectively), while
116 protein spots were observed in the aged worker brains
of both groups in the 14.74–67.90 kDa MW range and the
3.79–9.93 pI range (Fig. 1c, d respectively).
Considering that no apparent qualitative differences
were observed in the triplicates of 2-DE gels, and in order
to accurately compare the quantities of each protein in each
spot of the triplicate from each group of newly emerged
bees, or in the group of aged bees, the volume of each
protein in the spot was normalized and automatically cal-
culated by the software as a single spot volume. Differ-
ential expression analysis was performed using the
algorithm ImageMaster 2-DE Platinum 7.0 for the gels that
were representative of each experimental condition. This
analysis revealed a total of 25 proteins spots that were
differentially expressed in all the experiments. From these,
20 proteins spots characterized the proteomic differences
between the brains of the control and fipronil-treated
groups of newly emerged honeybees (Fig. 1a vs. b), and 15
differentially expressed proteins characterized the differ-
ences in the brains of aged honeybees in the presence and
absence of fipronil (Fig. 1c vs. d). Thus, these 25 protein
spots were assigned as numbers from 1 to 25 (Fig. 1), and
the corresponding proteins were identified with the protein
engine search MASCOT, with scores from 61 to 449, with
sequence coverages from 5 to 32 %.
T. C. Roat et al.
123

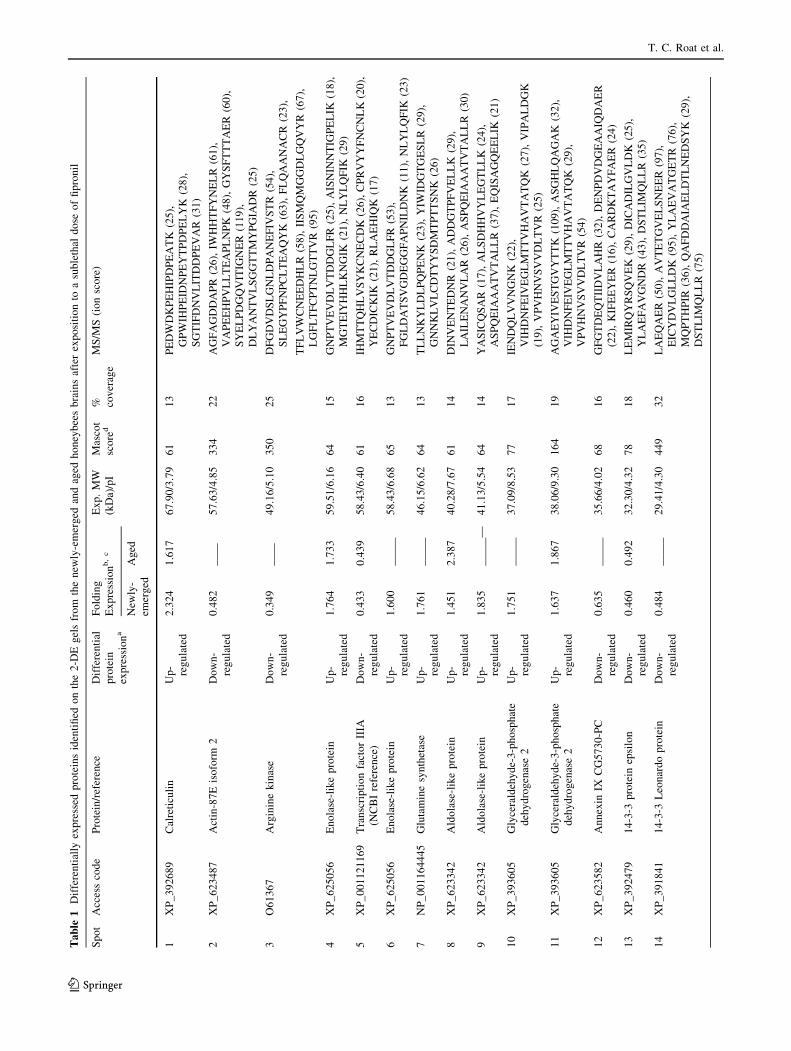
From the 25 selected proteins spots (Table 1), ten were
common to the newly emerged and aged honeybee work-
ers: calreticulin (spot 1), enolase-like protein (spot 4),
transcription factor IIIA (spot 5), aldolase-like protein (spot
8), glyceraldehyde-3-phosphate dehydrogenase (spot 11),
14-3-3 protein epsilon (spot 13), putative dual specificity
tyrosine-phosphorylation-regulated kinase 3 (spot 15),
protein lethal essential for life (spot 16), pyruvate dehy-
drogenase E (spot 18), and fatty acid binding protein (spot
20). Among these proteins, six were up-regulated
(1.63–2.92 fold) in the group that received a sub-lethal
dose of fipronil (calreticulin, enolase-like protein, aldolase-
like protein, glyceraldehyde-3-phosphate dehydrogenase,
protein lethal essential for life, and pyruvate dehydroge-
nase E), while four proteins were down-regulated (0.41 to
0.46-fold) by fipronil (transcription factor IIIA, 14-3-3
protein epsilon, tyrosine kinase 3, and fatty acid binding
protein).
Ten out of the 25 selected proteins were characteristic of
the brains from newly emerged honeybee workers: actin 87
E (spot 2), arginine kinase (spot 3), enolase-like protein
(spot 6), glutamine synthetase (GS) (spot 7), aldolase-like
protein (spot 9), glyceraldehyde-3-phosphate dehydroge-
nase (spot 10), annexin IX (spot 12), 14-3-3 Leonardo
protein (spot 14), thioredoxin peroxidase (spot 17), and
vascular endothelial growth factor receptor 3-like (VEGF-
3) (spot 19). Among these proteins, six were up-regulated
(1.60 to 2.16-fold) in the presence of fipronil (enolase-like
protein, GS, aldolase-like, glyceraldehyde-3-phosphate
dehydrogenase, thioredoxin peroxidase, and VEGF-3-like),
whereas four were down-regulated (0.35 to 0.48-fold) by
effect of fipronil (actin-87E, arginine kinase, annexin IX,
and 14-3-3 Leonardo protein). Five of the selected proteins
were characteristic aged honeybee worker brain proteins
that were up-regulated (1.53 to 1.99-fold) in the presence
of fipronil: defensin precursor (spot 21), neuroligin 2 (spot
22), protein lethal essential for life (spot 23), protein kinase
(spot 24), and amyloid precursor protein (spot 25).
Discussion
The present investigation was carried out to evaluate the
effects of honeybee exposure to a sub-lethal dose (1/100 of
LD50/day/bee) of the insecticide fipronil for 5 days and to
determine the proteomic profile of the honeybee brain. Up
to now, no previous studies have been performed to
investigate some potential biochemical damage to the
Fig. 1 Representative 2-DE
gels from the newly emerged
(a–b) and aged (c–d) honeybee
brain. a Control sample, 5 days
and b fipronil sample, 5 days.
c Control sample, 25 days and
d fipronil sample, 25 days
Brain proteome of Africanized honeybees
123
Ta
ble
1D
iffe
ren
tial
lyex
pre
ssed
pro
tein
sid
enti
fied
on
the
2-D
Eg
els
fro
mth
en
ewly
-em
erg
edan
dag
edh
on
eyb
ees
bra
ins
afte
rex
po
siti
on
toa
sub
leth
ald
ose
of
fip
ron
il
Sp
ot
Acc
ess
cod
eP
rote
in/r
efer
ence
Dif
fere
nti
al
pro
tein
exp
ress
ion
a
Fo
ldin
g
Ex
pre
ssio
nb,
cE
xp
.M
W
(kD
a)/p
I
Mas
cot
sco
red
% cov
erag
e
MS
/MS
(io
nsc
ore
)
New
ly-
emer
ged
Ag
ed
1X
P_
39
26
89
Cal
reti
culi
nU
p-
reg
ula
ted
2.3
24
1.6
17
67
.90
/3.7
96
11
3P
ED
WD
KP
EH
IPD
PE
AT
K(2
5),
GP
WIH
PE
IDN
PE
YT
PD
PE
LY
K(2
8),
SG
TIF
DN
VL
ITD
DP
EV
AR
(31
)
2X
P_
62
34
87
Act
in-8
7E
iso
form
2D
ow
n-
reg
ula
ted
0.4
82
__
__
57
.63
/4.8
53
34
22
AG
FA
GD
DA
PR
(26
),IW
HH
TF
YN
EL
R(6
1),
VA
PE
EH
PV
LL
TE
AP
LN
PK
(48
),G
YS
FT
TT
AE
R(6
0),
SY
EL
PD
GQ
VIT
IGN
ER
(11
9),
DL
YA
NT
VL
SG
GT
TM
YP
GIA
DR
(25
)
3O
61
36
7A
rgin
ine
kin
ase
Do
wn
-
reg
ula
ted
0.3
49
__
__
49
.16
/5.1
03
50
25
DF
GD
VD
SL
GN
LD
PA
NE
FIV
ST
R(5
4),
SL
EG
YP
FN
PC
LT
EA
QY
K(6
3),
FL
QA
AN
AC
R(2
3),
TF
LV
WC
NE
ED
HL
R(5
8),
IIS
MQ
MG
GD
LG
QV
YR
(67
),
LG
FL
TF
CP
TN
LG
TT
VR
(95
)
4X
P_
62
50
56
En
ola
se-l
ike
pro
tein
Up
-
reg
ula
ted
1.7
64
1.7
33
59
.51
/6.1
66
41
5G
NP
TV
EV
DL
VT
DD
GL
FR
(25
),A
ISN
INN
TIG
PE
LIK
(18
),
MG
TE
IYH
HL
KN
GIK
(21
),N
LY
LQ
FIK
(29
)
5X
P_
00
11
21
16
9T
ran
scri
pti
on
fact
or
IIIA
(NC
BI
refe
ren
ce)
Do
wn
-
reg
ula
ted
0.4
33
0.4
39
58
.43
/6.4
06
11
6IH
MT
TQ
HL
VS
YK
CN
EC
DK
(26
),C
PR
VY
YF
NC
NL
K(2
0),
YE
CD
ICK
IK(2
1),
RL
AE
HIQ
K(1
7)
6X
P_
62
50
56
En
ola
se-l
ike
pro
tein
Up
-
reg
ula
ted
1.6
00
__
__
_5
8.4
3/6
.68
65
13
GN
PT
VE
VD
LV
TD
DG
LF
R(5
3),
FG
LD
AT
SV
GD
EG
GF
AP
NIL
DN
K(1
1),
NL
YL
QF
IK(2
3)
7N
P_
00
11
64
44
5G
luta
min
esy
nth
etas
eU
p-
reg
ula
ted
1.7
61
__
__
_4
6.1
5/6
.62
64
13
TL
LN
KY
LD
LP
QP
EN
K(2
3),
YIW
IDG
TG
ES
LR
(29
),
GN
NK
LV
LC
DT
YY
SD
MT
PT
TS
NK
(26
)
8X
P_
62
33
42
Ald
ola
se-l
ike
pro
tein
Up
-
reg
ula
ted
1.4
51
2.3
87
40
.28
/7.6
76
11
4D
INV
EN
TE
DN
R(2
1),
AD
DG
TP
FV
EL
LK
(29
),
LA
ILE
NA
NV
LA
R(2
6),
AS
PQ
EIA
AA
TV
TA
LL
R(3
0)
9X
P_
62
33
42
Ald
ola
se-l
ike
pro
tein
Up
-
reg
ula
ted
1.8
35
__
__
_—
41
.13
/5.5
46
41
4Y
AS
ICQ
SA
R(1
7),
AL
SD
HH
VY
LE
GT
LL
K(2
4),
AS
PQ
EIA
AA
TV
TA
LL
R(3
7),
EQ
ISA
GQ
EE
LIK
(21
)
10
XP
_3
93
60
5G
lyce
rald
ehy
de-
3-p
ho
sph
ate
deh
yd
rog
enas
e2
Up
-
reg
ula
ted
1.7
51
__
__
_3
7.0
9/8
.53
77
17
IEN
DQ
LV
VN
GN
K(2
2),
VIH
DN
FE
IVE
GL
MT
TV
HA
VT
AT
QK
(27
),V
IPA
LD
GK
(19
),V
PV
HN
VS
VV
DL
TV
R(2
5)
11
XP
_3
93
60
5G
lyce
rald
ehy
de-
3-p
ho
sph
ate
deh
yd
rog
enas
e2
Up
-
reg
ula
ted
1.6
37
1.8
67
38
.06
/9.3
01
64
19
AG
AE
YIV
ES
TG
VY
TT
K(1
09
),A
SG
HL
QA
GA
K(3
2),
VIH
DN
FE
IVE
GL
MT
TV
HA
VT
AT
QK
(29
),
VP
VH
NV
SV
VD
LT
VR
(54
)
12
XP
_6
23
58
2A
nn
exin
IXC
G5
73
0-P
CD
ow
n-
reg
ula
ted
0.6
35
__
__
_3
5.6
6/4
.02
68
16
GF
GT
DE
QT
IID
VL
AH
R(3
2),
DE
NP
DV
DG
EA
AIQ
DA
ER
(22
),K
IFE
EY
ER
(16
),C
AR
DK
TA
YF
AE
R(2
4)
13
XP
_3
92
47
91
4-3
-3p
rote
inep
silo
nD
ow
n-
reg
ula
ted
0.4
60
0.4
92
32
.30
/4.3
27
81
8L
EM
IRQ
YR
SQ
VE
K(2
9),
DIC
AD
ILG
VL
DK
(25
),
YL
AE
FA
VG
ND
R(4
3),
DS
TL
IMQ
LL
R(3
5)
14
XP
_3
91
84
11
4-3
-3L
eon
ard
op
rote
inD
ow
n-
reg
ula
ted
0.4
84
__
__
_2
9.4
1/4
.30
44
93
2L
AE
QA
ER
(50
),A
VT
ET
GV
EL
SN
EE
R(9
7),
EIC
YD
VL
GL
LD
K(9
5),
YL
AE
VA
TG
ET
R(7
6),
MQ
PT
HP
IR(3
6),
QA
FD
DA
IAE
LD
TL
NE
DS
YK
(29
),
DS
TL
IMQ
LL
R(7
5)
T. C. Roat et al.
123

Ta
ble
1co
nti
nu
ed
Sp
ot
Acc
ess
cod
eP
rote
in/r
efer
ence
Dif
fere
nti
al
pro
tein
exp
ress
ion
a
Fo
ldin
g
Ex
pre
ssio
nb,
cE
xp
.M
W
(kD
a)/p
I
Mas
cot
sco
red
% cov
erag
e
MS
/MS
(io
nsc
ore
)
New
ly-
emer
ged
Ag
ed
15
XP
_3
95
33
8P
uta
tiv
ed
ual
spec
ifici
ty
tyro
sin
e-p
ho
sph
ory
lati
on
-
reg
ula
ted
kin
ase
3(N
CB
I
refe
ren
ce)
Do
wn
-
reg
ula
ted
0.4
18
0.3
72
35
.16
/6.4
36
21
3T
IIA
TP
DV
VL
K(2
5),
HIY
FIG
AN
AK
(23
),
NE
KR
FH
RQ
AQ
EE
VK
(22
),N
RII
HC
DM
KP
EN
VL
LK
(27
)
16
XP
_0
01
11
98
84
Pro
tein
leth
ales
sen
tial
for
life
(Pro
tein
Efl
21
)
Up
-
reg
ula
ted
2.0
04
1.9
85
21
.56
/5.9
41
31
27
LL
DQ
NF
GL
GL
YP
EQ
LL
SP
SR
(36
),R
DE
HG
WIS
R(5
8),
LS
SD
GV
LT
ITA
PR
(36
),IE
QT
GK
PA
IQT
K(4
1)
17
XP
_3
93
44
5T
hio
red
ox
inp
ero
xid
ase
1U
p-
reg
ula
ted
2.1
58
__
__
_2
0.8
3/5
.90
64
28
DY
GV
LD
EE
SG
VP
FR
(40
),Q
ITIN
DL
PV
GR
(27
),
LV
QA
FQ
YT
DK
HG
EV
CP
AG
WK
(31
)
18
XP
_3
97
34
6P
yru
vat
ed
ehy
dro
gen
ase
E1
Up
-
reg
ula
ted
2.9
21
1.6
72
18
.79
/6.5
36
35
NN
SQ
LH
DN
EN
LE
DL
ILP
SN
ED
WR
(23
),
IFIR
AL
ED
AG
FW
QR
NR
(18
),A
AG
LT
DID
NA
LS
GM
K
(20
),S
IEL
EE
NE
FA
FD
PIC
PK
(26
),E
ML
AV
SE
LV
CN
HR
(16
)
19
XP
_0
01
12
14
92
Vas
cula
ren
do
thel
ial
gro
wth
fact
or
rece
pto
r3
-lik
e(N
CB
I
refe
ren
ce)
Up
-
reg
ula
ted
1.6
72
__
__
_1
7.4
4/5
.46
67
11
YIT
NIP
EV
LY
TV
PK
(22
),M
SQ
QM
DL
SID
SE
TR
(25
),
GG
DL
LT
YL
HN
QR
(27
),G
ITH
RD
LA
AR
NIL
LT
ED
LT
VK
(20
)
20
Q7
6L
A4
Fat
tyac
idb
ind
ing
pro
tein
Do
wn
-
reg
ula
ted
0.4
30
0.7
67
17
.68
/7.4
86
92
1L
GE
EF
EE
ET
VD
GR
(45
),S
VC
TL
DG
NK
LIQ
VQ
K(2
8)
21
C7
AH
S8
Def
ensi
np
recu
rso
r-li
ke
pro
tein
Up
-
reg
ula
ted
__
__
_1
.72
12
5.3
3/8
.45
76
24
VT
CD
LL
SF
K(3
5),
TT
FK
NL
WD
KR
(26
)
22
B9
VM
Q8
Neu
roli
gin
1U
p-
reg
ula
ted
__
__
_1
.99
82
6.5
9/6
.27
68
11
LIV
VS
INF
RL
GV
LG
FL
K(2
3),
LT
LF
GY
GT
GA
AL
AN
FL
AV
SP
MV
K(2
7),
VV
LL
GG
SA
LS
PW
AIQ
R(2
5),
VA
TG
CP
GN
VE
AD
DIA
PC
LR
(20
),Y
HL
HE
IYS
TL
R
(18
)
23
XP
_0
01
12
00
70
Pro
tein
leth
ales
sen
tial
for
life
(pro
tein
Efl
21
)
Up
-
reg
ula
ted
__
__
_1
.53
02
2.2
6/5
.19
63
22
LH
MD
YY
RP
WG
EL
LR
(21
),V
VD
RF
VIV
EA
K(2
2),
VIN
IEH
TG
KP
CD
AQ
NE
ER
(25
)
24
B7
S8
R5
cAM
P-d
epen
den
tp
rote
in
kin
ase
cata
lyti
csu
bu
nit
Up
-
reg
ula
ted
__
__
_1
.62
82
0.8
0/9
.70
67
12
TV
SN
TS
QD
LV
TS
LK
(24
),D
DF
TK
R(2
7),
TL
GM
GA
FG
RV
KL
IKH
K(2
2),
VL
QS
INY
PF
VV
TL
K(1
8)
25
XP
_6
24
12
4A
my
loid
pro
tein
pre
curs
or-
lik
eC
G7
72
7-P
A
Up
-
reg
ula
ted
__
__
_1
.85
11
8.7
1/9
.93
61
12
GM
LV
PA
LL
GV
LM
VC
R(3
1),
VA
TL
CE
AG
EV
YL
AQ
HM
GE
QG
R(2
4),
DIT
NIV
ES
SH
YV
R(2
2),
CW
ES
HR
WN
AT
AA
DT
CR
(26
),
SF
AM
LL
PC
GIS
LF
SG
VE
FV
CC
PK
(18
)
aP
rote
ins
dif
fere
nti
ally
exp
ress
edw
ere
clas
sifi
edas
up
-o
rd
ow
n-r
egu
late
din
fip
ron
il-t
reat
edg
rou
pin
com
par
iso
nto
con
tro
lg
rou
pb
Val
ues
det
erm
ined
bas
edo
nin
ter-
exp
erim
enta
ln
orm
aliz
edsp
ot
vo
lum
ein
ten
sity
(IN
SV
I)c
Dif
fere
nce
ssi
gn
ifica
nt
atle
vel
of
P\
0.0
5d
Mas
cot
sco
re:
pro
tein
sco
res
hig
her
than
60
are
sig
nifi
can
t(P
\0
.05
)
Brain proteome of Africanized honeybees
123

honeybee’s nervous system that can be caused by fipronil.
Two groups of workers were used, i.e., 5 day old (newly
emerged workers) and 25 day old (aged workers), and their
exposure to the insecticide was biochemically character-
ized by the differential expression of 25 proteins in their
brains. These proteins can be organized into three groups:
i) ten proteins that are common to the newly emerged and
aged honeybee workers; ii) ten proteins that are charac-
teristic of the newly emerged honeybee workers; and iii)
five proteins that are typical of the aged honeybee workers.
The structural and functional features of these groups
are described as follows
i) The group of ten proteins that are common to the newly
emerged and aged honeybee workers was composed of six
proteins that are up-regulated by the effect of fipronil on
the brain (calreticulin, enolase-like protein, aldolase-like
protein, glyceraldehyde-3-phosphate dehydrogenase, pro-
tein lethal essential for life, and pyruvate dehydrogenase
E), and four proteins that are down-regulated in the hon-
eybee brain due to the action of fipronil (transcription
factor IIIA, 14-3-3 protein epsilon, tyrosine kinase 3, and
fatty acid binding protein).
Among the up-regulated proteins, the enzymes enolase,
aldolase, and glyceraldehyde-3-phosphate dehydrogenase,
are directly involved in glycolysis, while pyruvate dehy-
drogenase links glycolysis to the acid citric cycle (within
the mitochondria) releasing energy via NADH; thus, the
over-expression of these proteins is likely related to the
requirement for metabolic energy in the neurons of hon-
eybees that are effected by fipronil. According to Vidau
et al. (2009), in addition to being neurotoxic, fipronil also
exerts cytotoxic action on other tissues, acting on common
cellular targets that are components of mitochondrial
energy metabolism. The ability of fipronil to induce cyto-
toxicity is related to its ability to prevent the production of
ATP by mitochondrial disruption (Vidau et al. 2011).
The two other up-regulated proteins, calreticulin and
lethal protein (Efl21) are most likely related to neuronal
protection against the action of fipronil. Calreticulin binds
to misfolded proteins, preventing them from being expor-
ted from the endoplasmic reticulum to the Golgi apparatus
(Michalak et al. 2012), whereas the lethal protein Efl21 is
related to the heat shock protein family, playing the role of
a chaperonin (Kurzik-Dumke and Lohmann 1995).
According to Ki et al. (2012), oxidative stress plays a
central role in fipronil-induced cytotoxicity. Thus, both
proteins are likely related to neuronal protection against
protein misfolding that is caused by fipronil-induced
chemical stress in the neuronal soma.
This group also contains four proteins that are down-
regulated in the honeybee brain due to the action of
fipronil: transcription factor IIIA, 14-3-3 protein epsilon,
tyrosine kinase 3, and fatty acid binding protein. The
transcriptional repressor factor IIIA plays a role in regu-
lating the G2/M transition of the cell cycle (access code:
XP_001121169) 0.14-3-3 protein epsilon is involved in
neural signaling, neuronal development and neuroprotec-
tion and are known to promote cell survival by inhibiting
apoptotic processes via multiple mechanisms (Foote and
Zho 2012). Tyrosine kinase-3 plays a significant role in a
signaling pathway that regulates cell proliferation and is
involved with brain development (Galceran et al. 2004).
Fatty acid-binding protein (FABP) belongs to a family of
small proteins that act as cytoplasmic fatty acid trans-
porters, transferring fatty acids to membranes, and medi-
ating the effects of fatty acids on gene expression in
different tissues, including the brain. Thus, the group of
four down-regulated proteins is likely to be related to
neuronal development and neuroprotection; the fipronil-
induced decreased level of expression of these proteins in
the honeybee brain could be associated with neuronal
damage in these bees.
ii) The group of ten proteins that are characteristic of the
newly emerged honeybee workers was composed of six
proteins that are up-regulated by the action of fipronil
(enolase, GS, aldolase, glyceraldehyde-3-phosphate dehy-
drogenase, thioredoxin peroxidase, and VEGF-3-like pro-
tein) and four down-regulated proteins (actin-87E, arginine
kinase, annexin IX, and 14-3-3 Leonardo protein).
The overexpression of enolase, aldolase, and glycer-
aldehyde-3-phosphate dehydrogenase clearly indicates an
activation of glycolysis that is caused by the action of
fipronil in the honeybee brain, which was also observed in
the group of proteins that are common to the newly
emerged and aged honeybee workers, as discussed in the
above paragraphs. Because the proteomic identification of
these enzymes was based on the same accession numbers,
although the proteins had different MW and pI values
(Table 1), it is likely that they are different forms/isoforms
of the same proteins.
Glutamine synthetase (GS) acts in the brain by regu-
lating the levels of glutamate and detoxifying the neurons
by removing excess ammonia (Suarez et al. 2012); the up-
regulation of GS indicates that the honeybee’s brain may
be over-producing glutamate and/or ammonia as a result of
the action of fipronil. The enzyme thioredoxin peroxidase
is also related to protection against oxidative stress (Santos
et al. 2010); thus, the up-regulation of this enzyme is also
likely to be related to the protection of the honeybee’s
brain against the metabolic toxicity caused by the action of
fipronil. The VEGF-3-like protein stimulates vasculogen-
esis and angiogenesis to restore oxygen supply to the tis-
sues with inadequate blood circulation (Shin et al. 2010);
thus, the over-expression of this protein suggests that the
T. C. Roat et al.
123

fipronil-affected honeybee brain could have suffered from
some damage that restricts hemolymph circulation,
requiring the over-production of VEGF-3 to protect the
brain against possible ischemia that is caused by fipronil.
Additionally, four other proteins were down-regulated in
the newly emerged honeybee brains that were affected by
fipronil: actin-87E, 14-3-3 Leonardo protein, arginine
kinase, and annexin IX. It was recently reported in hon-
eybees that the depolymerization of actin in the brain is
related to the enhancement of the associative olfactory
memory and the increase of long-term memory retention in
insects (Ganeshina et al. 2012). The 14-3-3 Leonardo
protein is involved in forming olfactory learning and
memory structures that are necessary for scent recognition
and learning abilities (Skoulakis and Davis 1996). The
down-regulation of these two proteins in the brains of
newly emerged bees that are exposed to fipronil suggests
that the low level of expression of these proteins could be
related to the impairment of learning and/or decreased
memory. The arginine kinases (AK) from the honeybee
brain belongs to the family of ATP:guanidino phospho-
transferases. They are important components of the energy
shuttle that delivers ATP generated by the mitochondria to
biochemical processes of the visual system that require
high energy, such as pigment regeneration in the retina
(Kucharski and Maleszka 1998). The down-regulation of
this protein could contribute to the occurrence of visual
problems in worker honeybees that are affected by fipronil.
The annexins are a large family of calcium- and
phospholipid-binding proteins that regulate cellular growth
and a series of different signal transduction pathways
(Gerke and Moss 2002). However, currently, there are no
known processes that involve annexin IX in animal brains.
iii) The group of five proteins that are up-regulated in
the brains of aged honeybee workers is composed of a
defensin precursor, neuroligin 2, protein lethal essential for
the life (Efl21), a protein kinase, and amyloid precursor
protein. These proteins could potentially be related to the
age differences that characterize the changes of the work-
ers’ function in the nest, according to the insect’s age.
Defensins are non-specific proteins that mediate the innate
immune defense of insects and protect tissue surfaces from
invading pathogens (Bulet and Stocklin 2005). Thus, the
up-regulation of a defensin could indicate that fipronil
affects the honeybee immune system and thus the honey-
bee became more susceptible to microorganism infection in
the presence of fipronil. Neuroligins are a family of pro-
teins that mediate the synapse formation between neurons,
which affects the signaling of neural networks; in mam-
mals, structural alterations of these proteins are involved in
the occurrence of cognitive disorders (Sudhof 2008). The
overexpression of neuroligin 2 in the brains of aged worker
bees that are exposed to fipronil may indicate that the
insecticide could have affected synapse formation in the
bee’s neurons. The lethal protein essential for life (Efl21) is
related to a family of small heat shock proteins that play
the role of chaperonins (Kurzik-Dumke and Lohmann
1995). The up-regulation of this protein suggests that some
important neuronal proteins in these insects are misfolded,
which requires the action of the chaperonin to protect the
neural proteins against denaturation that can result in loss
of biological activity. The cAMP-dependent protein kinase
catalytic subunit mediates cAMP-dependent signaling that
is triggered by receptor binding to GPCRs. Its activation
regulates a series of cellular processes such as cell prolif-
eration, cell cycle, differentiation and regulation of
microtubule dynamics, and it is important for brain plas-
ticity (Qi et al. 1996). The overexpression of this protein
could be related to neuronal proliferation and/or differen-
tiation that are induced by fipronil in the brains of aged
honeybee workers. The amyloid protein is an important
component of senile plaques in animal brain; the overex-
pression of this protein is strongly associated with neuronal
degeneration (Murakami et al. 1998). Therefore, the up-
regulation of the proteins discussed above indicates that
fipronil rendered the aged bees more susceptible to path-
ogenic infection, and likely damaged the formation of
synapses, protein misfolding and neuronal degeneration.
The proteins identified in the present investigation were
already reported in previous studies of honeybee brain
proteomics; as exemple Li et al. (2010) and Hernandez et al.
(2012) reported the presence of calreticulin, actin-87E,
arginine kinase, enolase, GS, aldolase, glyceraldehyde-3-
phosphate, annexin, 13-3-3 protein epsilon, Leonardo pro-
tein, protein lethal, thioredoxin peroxidase, pyruvate
dehydrogenase, fatty acid binding protein, cAMP-depen-
dent protein kinase and amyloid protein precursor-like in
the brains of honeybees submitted to differential protein
expression in the caste differentiation studies and compar-
ative proteomic analysis between nurse and forager subc-
astes, respectively. The cAMP-dependent protein kinase,
for example, contributes to the induction of a long-term
memory formation in honeybees (Fiala et al. 1999). Garcia
et al. (2009) also reported the proteins such as calreticulin,
GS and protein lethal in the honeybee brains upon onto-
genetic and behavioral development. Neuroligins have been
previously reported in studies of the honeybee brain syn-
aptic development, since this family of proteins is involved
in synapse formation in A. mellifera, and thus it is used as a
model for comparison with synaptic development in ver-
tebrates (Biswas et al. 2008). However, in none of these
studies honeybee brain proteomics was used as strategy to
investigate the toxicological effects of an insecticide in the
nervous system, as done in the present study.
Figure 2 summarizes the interpretation of the effects of
honeybee exposure to a sub-lethal dose of fipronil in the
Brain proteome of Africanized honeybees
123

brains of newly emerged and aged honeybees. The effects
were detected as differential protein expression changes by
comparing the control versus fipronil-treated groups of 5
and 25 day old honeybees. Considering the results that
were obtained for both ages, fipronil initially causes
structural damage that requires the activation of the
anaerobic phase of glycolysis to supply energy to repair the
damage. The presence of fipronil causes chemical stress in
the brain, which leads to the increased expression of pro-
teins that are related to neuronal protection; fipronil also
renders the aged bees more susceptible to pathogenic
infection. The chemical stress in neurons causes protein
misfolding, which results in apoptosis, brain ischemia, and
possibly visual problems that result from impaired pigment
regeneration after oxidation/reduction. These processes are
followed by damage to synapse formation and neurode-
generation, which could result in memory and learning
impairment in aged honeybee workers.
Conclusions
Honeybees have fewer xenobiotic detoxifying enzymes
than other non-social insects, which render honeybees
highly sensitive to a series of insecticides, when compared
to Drosophila melanogaster and Anopheles gambiae
Fig. 2 Flow chart indicating
the interpretation of the possible
effects of a sub-lethal dose of
fipronil on the brains of newly
emerged and aged honeybees.
The actions resulting from the
up-regulation ( ) or down-
regulation ( ) of proteins are
assigned outside of the dashed
square blocks, while the
identified proteins that are
differentially expressed are
indicated within the
corresponding action block
T. C. Roat et al.
123

(Claudianos et al. 2006). This aspect makes the honeybee
an interesting living model for testing novel pesticides and/
or xenobiotics. Recently, a review that was conducted by
the European Food Safety Authority stated that phenylpy-
razoles pesticides present a high risk to the survival of
honeybees (European Food Safety Authority 2013). How-
ever, the degree of risk that is presented by this class of
compounds requires further investigation. To obtain more
details regarding the effect of fipronil on the honeybee
brain, the present investigation focused on a toxicoprote-
omic analysis of newly emerged and aged honeybee
workers that were subjected to a sub-lethal dose of fipronil
for 5 days.
This study demonstrated the differential brain protein
expression profile of the Africanized A. mellifera, an insect
that is not targeted by fipronil. Our results suggest that low
doses of fipronil (sub-lethal doses) may cause changes in
the expression pattern of certain proteins that are involved
in pathogen susceptibility, neuronal chemical stress, neu-
ronal protein misfolding, a higher occurrence of apoptosis,
ischemia, visual problems, damage to synapse formation,
and brain degeneration, that could result in memory and
learning impairment. Taken together, these actions can
induce several types of injuries to honeybee physiology.
Besides, this study demonstrates that honeybees chroni-
cally exposed to sublethal doses of fipronil could have a
reduction of their life span, showing the importance of
establishing more accurate methods for risk assessment of
pesticides on bees as discussed by Decourtye et al. (2013).
Acknowledgments This work was supported by Grants from FA-
PESP (Proc. 2008/05018-7 and 2012/13370-8), BIOprospecTA/FA-
PESP program (Proc. 11/51684-1), CNPq and CAPES. M.S.P. and
O.M. are researchers from the National Research Council of Brazil-
CNPq. T.C.R., L.D.S. and K.S.S. were Postdoctoral fellows from
FAPESP, and J.R.A.S.P is Ph.D. student fellow from FAPESP.
Conflict of interest The authors declare that they have no conflict
of interest.
References
Aliouane Y, El Hassani AK, Gary V, Armengaud C, Lambin M,
Gauthier M (2009) Subchronic exposure of honeybees to
sublethal doses of pesticides: effects on behavior. Environ
Toxicol Chem 28(1):113–122
Barbara GS, Zoube C, Rybak J, Gauthier M, Grunewald B (2005)
Acethylcoline, GABA and glutamate induce ionic currents in
cultured antennal lobe neurons of the honeybee, Apis mellifera.
J Comp Physiol 191:823–836
Bernadou A, Demares F, Couret-Fauvel T, Sandoz JC, Gauthier M
(2009) Effect of fipronil on side specific antennal tactile learning
in the honeybee. J Insect Physiol 55:1099–1106
Biswas S, Russell RJ, Jackson CJ, Vidovic M, Ganeshina O,
Oakeshott JG, Claudianos C (2008) Bridging the synaptic gap:
neuroligins and neurexin I in Apis mellifera. PLoS One
3(10):e3542
Bradford MM (1976) A rapid and sensitive method for the
quantitatioin of microgram quantities of protein utilizing the
principle of protein-dye binding. Anal Biochem 2:248–254
Bromensshenk JJ, Henderson CB, Wick CH, Stanford MF, Zulich
AW, Jabbour RE et al (2010) Iridovirus and microsporidian
linked to honey bee colony decline. PLoS One 5:1–11
Bulet P, Stocklin R (2005) Insect antimicrobial peptides: structures,
properties and gene regulation. Protein Pept Lett 12:3–11
Claudianos C, Ranson H, Johnson RM, Biswas S, Schuler A,
Berembaum MR, Feyereisen R, Oakeshott JG (2006) A deficit of
detoxification enzymes: pesticide sensitivity and environmental
response in the honeybee. Insect Mol Biol 15:615–636
Decourtye A, Devillers J, Genecque E, Le Menach K, Budzinski H,
Cluzeau S, Pham-Delegue MH (2005) Comparative sub-lethal
toxicity of nine pesticides on olfactory learning performances of
the honeybee Apis mellifera. Arch Environ Contam Toxicol
48:242–250
Decourtye A, Lefort S, Devillers J, Gauthier M, Aupinel P, Tisseur M
(2009) Sublethal effects of fipronil on the ability of honeybees
(Apis mellifera L.) to orientate in a complex maze. In: Oomen
PA, Thompson HM (eds) Hazards of pesticides to bees. Arno
Brynda GmbH, Berlin, pp 75–83
Decourtye A, Henry M, Desneux N (2013) Environment: overhaul
pesticide testing on bees. Nature 497(7448):188
Desneux N, Decourtye A, Delpuech JM (2007) The sublethal effects
of pesticides on beneficial arthropods. Ann Rev Entomol
52:81–106
El Hassani AK, Dacher M, Gauthier M, Armengaud C (2005) Effects
of sublethal doses of fipronil on the behavior of the honeybee
(Apis mellifera). Pharmacol Biochem Behav 82:30–39
European Food Safety Authority (2013) Conclusion on the peer
review of the pesticide risk assessment for bees for the active
substance clothianidin. EFSA J 11(1):3066
Fiala A, Muller U, Menzel R (1999) Reversible downregulation of
protein kinase A during olfactory learning using antisense
technique impairs long-term memory formation in the honeybee,
Apis mellifera. J Neurosci 19(22):10125–10134
Foote M, Zho Y (2012) 14-3-3 proteins in neurological disorders. Int J
Biochem Mol Biol 3(2):152–164
Galceran J, de Graaf K, Tejedor FJ, Becker W (2004) The MNB/
DYRK1A protein kinase: genetic and biochemical properties.
J Neural Transm Suppl 67:139–148
Ganeshina O, Erdmann J, Tiberi S, Vorobyev M (2012) Depolymer-
ization of actin facilitates memory formation in an insect. Biol
Lett 8(6):1023–1027
Garcia L, Garcia CHS, Calabria LK, Cruz GCN, Puentes AS, Bao SN,
Fontes W, Ricart CAO, Espindola FS, de Sousa MV (2009)
Proteomic analysis of honey bee brain upon ontogenetic and
behavioral development. J Proteome Res 8:1464–1473
Gerke V, Moss S (2002) Annexins: form structure to function. Physiol
Rev 82(2):331
Greig-Smith PW, Thompson HM, Hardy AR, Bew MH, Findlay E,
Stevenson JH (1994) Incidents of poisoning of honeybees (Apis
mellifera) by agricultural pesticides in Great Britain 1981–1991.
Crop Prot 13:567–581
Gunasekara AS, Truong T, Goh KS, Spurlock F, Tjeerdema RS
(2007) Environmental fate and toxicology of fipronil. J Pestic Sci
32:189–199
Halm MP, Rortais A, Arnold G, Tasei JN, Rault S (2006) New risk
assessment approach for systemic insecticides: the case of honey
bees and imidacloprid (Gaucho). Environ Sci Technol
40:2448–2454
Han P, Niu CY, Lei CL, Cui JJ, Desneux N (2010a) Quantification of
toxins in a Cry1Ac ? CpTI cotton cultivar and its potential
effects on the honey bee Apis mellifera L. Ecotoxicology
19(8):1452–1459
Brain proteome of Africanized honeybees
123

Han P, Niu CY, Lei CL, Cui JJ, Desneux N (2010b) Use of an
innovative T-tube maze assay and the proboscis extension
response assay to assess sublethal effects of GM products and
pesticides on learning capacity of the honey bee Apis mellifera
L. Ecotoxicology 19(8):1612–1619
Hernandez LG, Lu B, da Cruz GCN, Calabria LK, Martins NF,
Togawa R, Espindola FS, Yates JR, Cunha RB, de Sousa MV
(2012) Worker honeybee brain proteome. Proteome Res
11:1485–1493
Ki YW, Lee JE, Park JH, Shin IC, Koh HC (2012) Reactive oxygen
species and mitogen-activated protein kinase induce apoptotic
death of SH-SY5Y cells in response to fipronil. Toxicol Lett
211(1):18–28
Kremen C, Williams NM, Aizen MA, Gemmill-Herren B, Lebuhn G,
Minckley R et al (2007) Pollination and other ecosystem services
produced by mobile organism: a conceptual framework for the
effects of land-use change. Ecol Lett 10:299–314
Kucharski R, Maleszka R (1998) Arginine kinase is highly expressed
in the compound eye of the honey bee, Apis mellifera. Gene
211(2):343–349
Kurzik-Dumke U, Lohmann E (1995) Sequence of the new
Drosophila melanogaster small heat-shock-related gene,
lethal(2) essential for life [l(2)efl], at locus 59F4,5. Gene
154(2):171–175
Li J, Wu J, Begna-Rundassa D, Song F, Zheng A (2010) Differential
protein expression in honeybee (Apis mellifera L.) larvae:
underlying caste differentiation. PLoS One 5(10):e13455
Lourenco CT, Carvalho SM, Maslapina O, Nocelli RCF (2012) Oral
toxicity of fipronil insecticide against the stingless bee Melipona
scutellaris (Latreille, 1811). Bull Environ Contam Toxicol
89:921–924
Malaspina O, Silva-Zacarin ECM (2006) Cell makers for ecotoxico-
logical studies in target organs of bees. Braz J Morphol Sci 23(3/
4):303–309
Maxim L, van der Sluijs JP (2007) Uncertainty: cause or effect of
stakeholders’ debates? analysis of a case study: the risk for
honeybees of the insecticide Gaucho�. Sci Total Environ
376:1–17
Michalak M, Lynch J, Groenendyk J, Guo L, Robert Parker JM, Opas
M (2012) Calreticulin in cardiac development and pathology.
Biochim Biophys Acta 1600(1–2):32–37
Murakami N, Yamaki T, Iwamoto Y, Sakakibara T, Kobori N,
Fushiki S, Ueda S (1998) Experimental brain injury induces
expression of amyloid precursor protein, which may be related to
neuronal loss in the hippocampus. J Neurotrauma
15(11):993–1003
OEPP/EPPO (2003) Environmental risk assessment scheme for plant
protection products. Bull OEPP/EPPO 33:141–145
Pham-Delegue MH, Decourtye A, Kaiser L, Devillers J (2002)
Behavioural methods to assess the effects of pesticides on honey
bees. Apidologie 33:425–432
Pochi D, Biocca M, Fanigliulo R, Pulcini P, Conte E (2012) Potential
exposure of bees, Apis mellifera L., to particulate matter and
pesticides derived from seed dressing during maize sowing. Bull
Environ Contam Toxicol 89:354–361
Qi M, Zhuo M, Skalhegg BS, Brandon EP, Kandel ER, McKnight GS,
Idzerda RL (1996) Impaired hippocampal plasticity in mice
lacking the Cbeta1 catalytic subunit of cAMP-dependent protein
kinase. Proc Natl Acad Sci USA 93(4):1571–1576
Roat TC, Carvalho SM, Nocelli RCF, Silva-Zacarin ECM, Palma MS,
Malaspina O (2013) Effects of sublethal dose of fipronil on
neuron metabolic activity of africanized honeybees. Arch
Environ Contam Toxicol 64:456–466
Rondeau G, Sanchez-Bayo F, Tennekes HA, Decourtye A, Ramırez-
Romero R, Desneux N (2014) Delayed and time-cumulative
toxicity of imidacloprid in bees, ants and termites. Sci Rep 4.
doi:10.1038/srep05566
Rortais A, Arnold G, Halm MP, Touffet-Briens F (2005) Modes of
honeybees exposure to systemic insecticides: estimated amounts
of contaminated pollen and nectar consumed by different
categories of bees. Apidologie 36:71–83
Santos LD, Santos KS, Santos-Pinto JRA, Dias NB, De Souza BM,
Dos Santos M, Perales J, Domont GB, Castro FM, Kalil JE,
Palma MS (2010) Profiling the proteome of the venom from the
social wasp Polybia paulista: a clue to understand the enven-
oming mechanism. J Prot Res 9(8):3867–3877
Schafer S, Bicker G (1986) Distribution of GABA-like imunoreac-
tivity in the brain of the honeybee. J Comp Neurol 246:287–300
Shin YJ, Choi JS, Choi JY, Hou Y, Cha JH, Chun MH, Lee MY
(2010) Induction of vascular endothelial growth factor receptor-3
mRNA in glial cells following focal cerebral ischemia in rats.
J Neuroimmunol 229(1–2):81–90
Skoulakis EMC, Davis RL (1996) Olfactory learning deficits in
mutants for Leonardo, a Drosophila gene encoding a 14-3-3
protein. Neuron 17(5):931–944
Suarez I, Bodega G, Fernandez B (2012) Glutamine synthetase in
brain: effect of ammonia. Neurochem Int 41(2–3):123–142
Sudhof TC (2008) Neuroglins and neurexins link synaptic function to
cognitive disease. Nature 455(7215):903–911
Tingle CC, Rother JA, Dewhurst CF, Lauer S, King WJ (2003)
Fipronil: environmental fate, ecotoxicology, and human health
concerns. Rev Environ Contam Toxicol 176:1–66
vanEngelsdorp D, Meixner MD (2009) A historical review of
managed honey bee populations in Europe and United States
and the factors that may affect them. J Invert Pathol 10:10–16
Vidau C, Brunet JL, Badiou A, Belzunces LP (2009) Phenylpyrazole
insecticides induce cytotoxicity by altering mechanisms
involved in cellular energy supply in the human epithelial cell
model Caco-2. Toxicol In Vitro 23:589–597
Vidau C, Gonzalez-Polo RA, Niso-Santano M, Gomez-Sanchez R,
Bravo-San PJM, Pizarro-Estrella E, Blasco R, Brunet JL,
Belzunces LP, Fuentes JM (2011) Fipronil is a powerful
uncoupler of oxidative phosphorylation that triggers apoptosis
in human neuronal cell line SHSY5Y. Neurotoxicology
32(6):935–943
Wratten SD, Gillespie M, Decourtye A, Mader E, Desneux N (2012)
Pollinator habitat enhancement: benefits to other ecosystem
services. Agric Ecosyst Environ 159:112–122
T. C. Roat et al.
123