nuclear matrins: identification major nuclearmatrix proteins · proc. nati. acad. sci. usa vol. 88,...
TRANSCRIPT

Proc. NatI. Acad. Sci. USAVol. 88, pp. 10312-10316, November 1991Cell Biology
Nuclear matrins: Identification of the major nuclear matrix proteinsHIROSHI NAKAYASU* AND RONALD BEREZNEYtDepartment of Biological Sciences, State University of New York at Buffalo, Buffalo, NY 14260
Communicated by Keith R. Porter, August 23, 1991 (received for review June 27, 1991)
ABSTRACT A preparative two-dimensional polyacrylam-ide gel system was used to separate and purify the majorCoomassie blue-stained proteins from the isolated rat livernuclear matrix. Approximately 12 major proteins were con-sistently found. Of these, 5 proteins represented identifiedproteins, including nuclear lamins A, B, and C, the nucleolarprotein B-23, and residual components of core heterogeneousnuclear ribonucleoproteins. The remaining eight major pro-teins termed the nuclear matrins consisted of matrin 3 (125kDa, slightly acidic), matrin 4 (105 kDa, basic), matrins D-G(60-75 kDa, basic), and matrins 12 and 13 (42-48 kDa, acidic).Peptide mapping and two-dimensional immunoblot studiesindicate that matrins D-G compose two pairs of related pro-teins (matrins D/E and F/G) and that none of the matrinsresemble the nuclear lamins or any of the other major proteinsdetected on our two-dimensional gels. Subfractionation immu-noblot experiments demonstrated the nearly exclusive localiza-tion of matrins F/G and other matrins to the nuclear matrixfraction of the cell. These results were further supported byindirect immunofluorescence microscopy that showed a strictlyinterior nuclear localization of the matrins in intact cells incontrast to the peripherally located nuclear lamins. We con-clude that the nuclear matrins are a major class of proteins ofthe nuclear matrix interior and are distinct from the nuclearlamins.
There is a growing awareness that many of the importantanswers to questions concerning the expression and regula-tion of the eukaryotic genome will require an understandingof the higher-order arrangement and function of the geneticcomponents as they interact within the complex three-dimensional architecture of the cell nucleus (1-13). Thenuclear matrix, a residual nuclear structure that has beenisolated from a wide variety of eukaryotic cells throughoutthe phylogenetic scale (1-3, 6, 7, 13-16), offers a potentiallyvaluable in vitro approach for studying nuclear processes inrelation to nuclear structure. Indeed, numerous studies haveimplicated the matrix as a site of organization for virtually allknown nuclear processes (1-11, 13, 16), such as, DNA loopattachment, DNA replication, transcription, RNA splicingand transport, hormone receptor function, carcinogen bind-ing, oncogene proteins, viral proteins, and protein phosphor-ylation.
Despite this progress, our knowledge of the proteins thatcompose this proteinaceous nucleoskeletal structure is still inits infancy. In this study we have used high-resolutionpreparative PAGE to identify and purify many of the majorCoomassie blue-stained nuclear matrix proteins. A class ofnuclear matrix proteins termed the nuclear matrins is iden-tified and characterized by peptide maps, polyclonal anti-bodies generated against the individual purified matrins, andindirect immunofluorescence microscopy.
MATERIALS AND METHODSIsolation of Nuclei and Nuclear Matrix. Nuclei and nuclear
matrix were isolated from rat liver by a modification of thehigh-salt extraction procedures of Berezney and Coffey (6,14, 15) as described (17). Phenylmethylsulfonyl fluoride (1mM) and sodium tetrathionate (0.1 mM) were added to all theisolation solutions through the high salt extraction (2 M NaCl)step (17).Two-Dimensional (2D) Electrophoresis, Purification of In-
dividual Nuclear Matrix Proteins, and Preparation of ChickenPolyclonal Antibodies. The 2D nonequilibrium pH gradientand SDS/PAGE system of O'Farrell et al. (18) was modifiedto optimize separation of the major nuclear matrix proteinson a large scale. Total rat liver nuclear matrix protein (2-3mg) was separated on 13-cm tube gels for the first-dimensional pH gradient (pH 3-10 ampholytes, 400 V, 14-16h, 22°C). The second-dimensional gels measured 15 cm x 30cm x 1.5 mm and consisted of a 2-cm 3.5% polyacrylamidestacking gel on top of a 28-cm 5-12% gradient gel. Electro-phoresis was performed at 15 V for 4 h followed by 250 V for20-24 h at 4°C. The gels were stained with Coomassie blueand the major stained spots were excised and stored at-20°C. The individual proteins were further purified afterextraction from the gel pieces (in 60 mM TrisHCl, pH6.8/0.1% SDS/2% 2-mercaptoethanol) on one-dimensional(1D) SDS/PAGE gels (10% polyacrylamide) according toLaemmli (19) and stored as gel slices at -20°C.For polyclonal antibody production, multiple gel slices for
each protein were washed four times with 10 mM sodiumphosphate, pH 7.2/0.15 M NaCl and homogenized in thesame buffer with 2 vol of Freund's complete or incompleteadjuvant. Intramuscular and subcutaneous injections intomature laying hens were given with complete adjuvant at day1 (100-200 gg of protein) and every 2 weeks with incompleteadjuvant (50-100 ,ug of protein). Sera were prepared at day28 and subsequently every 2 weeks.2D Western Blot Analysis. Total nuclear matrix protein (100
,ug) was separated on the 2D gel system described above ona miniscale (5.5-cm tube gels and 5 cm x 7.5 cm x 0.45 mmslab gels) and electrophoretically transferred to a nitrocellu-lose sheet (20). After blocking with 0.2% Tween 20/10 mMTris-HCI, pH 7.4/0.15 M NaCl for 2-4 h, the blots werestained with india ink (21) or incubated for 12 h with thevarious polyclonal antibodies (1: 100 dilution of sera) followedby 2 h with alkaline phosphatase-conjugated goat anti-chicken IgG secondary antibodies (1:1000 dilution) and de-veloped as described (22).
Peptide Analysis of Purified Nuclear Matrix Proteins. Pro-teins were extracted from the gel slices as described above forthe 2D gel pieces, precipitated with 3 vol of absolute ethanol(-20°C, 16 h), and dissolved in 0.1% SDS/60 mM Tris-HCI,pH 6.8. For 1D peptide analysis, 3 1,u of each purified protein
Abbreviations: hnRNP, heterogeneous nuclear ribonucleoprotein;1D and 2D, one and two dimensional, respectively.*Present address: Department of Pharmacology, Kyoto PrefecturalUniversity of Medicine, Kawaramachi-Hirokoji, Kamikyo, Kyoto602, Japan.tTo whom reprint requests should be addressed.
10312
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 88 (1991) 10313
solution (3 jig of protein) was incubated with 2 ,1l of eitherL-1-tosylamido-2-phenylethyl chloromethyl ketone-treatedtrypsin (80 ng; Sigma) or protease V8 (300 ng; Sigma) for 1 hat 370C and subjected to SDS/PAGE using either a 15%(protease V8 digested) or 12% (trypsin digested) minigel (19).The gels were stained with Coomassie blue. For 2D analysis,14 pug of each nuclear matrix protein was incubated with 160ng of trypsin (370C, 1 h). The reactions were stopped with 0.1mM phenylmethylsulfonyl fluoride, and the peptides wereprecipitated with ethanol and subjected to 2D SDS/PAGEusing the 10%o polyacrylamide minigel system describedabove.
Affinity Purification of Chicken Polyclonal Antibodies andIndirect Immunofluorescence Microscopy. For immunofluo-rescence microscopy, the sera containing the polyclonalantibodies were first affinity purified on nitrocellulose. Nu-clear matrix proteins from 2D gels were transferred tonitrocellulose (20). Individual spots corresponding to themajor proteins were identified by brief staining with india ink(21), excised, and incubated with the corresponding chickenserum (16 h, room temperature). The filter spots were washedfour times with 0.2% Tween 20/10mM TrisHCl, pH 7.4/0.15M NaCI and distilled water. Affinity-purified antibodies wereeluted from the nitrocellulose with 0.1 M glycine hydrochlo-ride (pH 2.2) on ice for 5 min and immediately neutralizedwith 3 M Trizma base. Immunofluorescence microscopy wasperformed on rat kangaroo kidney (PtK1) cells grown oncoverslips as described (23).
RESULTS2D Polyacrylamide Gel Electrophoresis and Identification of
Nuclear Matrix Proteins. Nuclear matrix proteins from ratliver were separated on a preparative 2D SDS/PAGE system(18). Whereas >50 spots were detected on the 2D gels (Fig.1), only =12 major Coomassie blue-stained proteins werepresent. Most of these major nuclear matrix proteins mi-grated between 60 and 200 kDa with an approximately equaldistribution between the acidic and basic sides of the gel. Wehave numbered these major proteins from 1 to 15 including amore minor protein at position 2, which served as a control
Acidic
kDa - -
97- 1
68-II 1t*B)_*!(i
57-12 1t(C)
45-13
'4(B-23)
Basic
2
- 4
6(D) 5(F)_W -7(G)
6(E) .
for certain experiments, and protein 1, which often appearsas a more minor protein by Coomassie blue staining. Sinceour preparations include the nuclear lamins A, B, and C asmajor components, we have also labeled a series of fourproteins, proteins D-G, that migrated at similar apparentmolecular masses (60-75 kDa) but are much more basic incharge.The 2D pattern of the major Coomassie blue-stained poly-
peptides was extremely reproducible in rat liver and was alsocharacteristic of nuclear matrix isolated from a variety ofother mammalian cells including HeLa S-3 (24), Chinesehamster lung cells (Don line), mouse 3T3 fibroblasts, andPtK1 rat kangaroo kidney cells (data not shown).
Relationship of Nuclear Matrix Proteins to Known NuclearProteins. To determine whether any of the major proteinsother than the nuclear lamins were known nuclear proteins,we obtained a variety ofantibodies to nuclear proteins. Theseincluded antibodies to core heterogeneous nuclear ribonu-cleoproteins (hnRNPs) A, B, and C (25-27), other presump-tive hnRNPs (26, 27), proteins of the small nuclear RNPparticles (28, 29), the nucleolar specific proteins B-23 ornumatrin (30, 31), and C-23 or nucleolin (32, 33). Immunoblotanalysis showed that spot 14 is B-23 and spot 15 containshnRNP core proteins (data not shown). All the other anti-bodies to RNP and nucleolar proteins reacted only with moreminor polypeptides on the 2D gels (data not shown). Proteins3, 4, 12, 13, and D-G are, therefore, uncharacterized majorproteins of the nuclear matrix. We have termed these pro-teins the nuclear matrins to distinguish them from nuclearlamins A, B, and C.
Purification and Peptide Mapping of Individual NuclearMatrix Proteins. We next purified large quantities of individ-ual major matrix proteins from 64 preparative 2D gels (Fig.2c). The 1D V8 peptide maps of most of the proteins werevery different (Fig. 2a). As reported (34), the peptide mapsof lamins A and C were very similar to each other but quitedistinct from that of lamin B. Since matrins D-G were veryresistant to V8 protease, we used trypsin digestion. Thedigestion patterns of matrins D and E resembled each other
akDa97-68-45-35-24-18-14-
3 4 F G D E A C B 12 13 14 15
A B C D E F G
15(hn RNP)
* _ -- .....
..-
_ q
- S
A BO * G_
A B C D E F G
68- __45-35-24-
FIG. 1. Preparative 2D PAGE of nuclear matrix proteins. Totalnuclear matrix proteins from rat liver were separated on a prepara-tive (2-3 mg of protein) PAGE system. The major Coomassieblue-stained proteins are numerically labeled including one minorspot (protein 2) and another spot (protein 1) that often stain lessintensely than shown. Nuclear lamins A, B, and C are indicated withletters along with a group of matrin proteins that migrate in the samemolecular weight range (matrins D-G). A broken line indicates theapproximate position of pH 7 on the pH gradient. Molecular massmarkers from top to bottom were thyroglobulin, phosphorylase b,bovine serum albumin, pyruvate kinase, ovalbumin, and lactatedehydrogenase.
18-14-
b c
FIG. 2. 1D peptide maps of nuclear matrix proteins. Nuclearmatrix proteins purified from 2D gels (identified by lane labels) weredigested with V8 protease (a) or trypsin (b) and subjected to 1DPAGE. (c) Purified proteins before protease digestion. Molecularmass markers were, from top to bottom, bovine serum albumin,ovalbumin, glyceraldehyde 3-phosphate, trypsinogen, lactoglobulin,and lysozyme.
Cell Biology: Nakayasu and Berezney
10314 Cell Biology: Nakayasu and Berezney
but were very different from those of lamins A and C (Fig.2b). Matrin G showed a digestion pattern distinct from thelamins and matrins D and E.The 2D peptide maps of lamins A and C were very similar
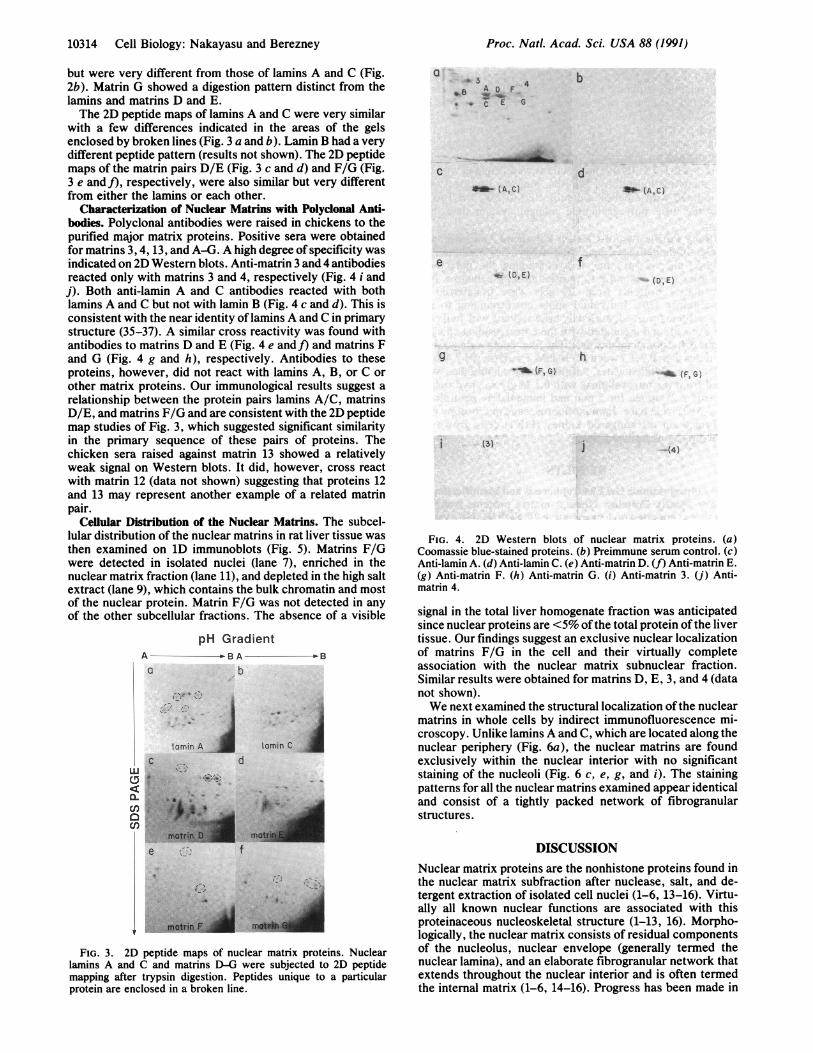
with a few differences indicated in the areas of the gelsenclosed by broken lines (Fig. 3 a and b). Lamin B had a verydifferent peptide pattern (results not shown). The 2D peptidemaps of the matrin pairs D/E (Fig. 3 c and d) and F/G (Fig.3 e and f), respectively, were also similar but very differentfrom either the lamins or each other.
Characterization of Nuclear Matrins with Polyclonal Anti-bodies. Polyclonal antibodies were raised in chickens to thepurified major matrix proteins. Positive sera were obtainedfor matrins 3, 4, 13, and A-G. A high degree of specificity wasindicated on 2D Western blots. Anti-matrin 3 and 4 antibodiesreacted only with matrins 3 and 4, respectively (Fig. 4 i andj). Both anti-lamin A and C antibodies reacted with bothlamins A and C but not with lamin B (Fig. 4 c and d). This isconsistent with the near identity of lamins A and C in primarystructure (35-37). A similar cross reactivity was found withantibodies to matrins D and E (Fig. 4 e and]) and matrins Fand G (Fig. 4 g and h), respectively. Antibodies to theseproteins, however, did not react with lamins A, B, or C orother matrix proteins. Our immunological results suggest arelationship between the protein pairs lamins A/C, matrinsD/E, and matrins F/G and are consistent with the 2D peptidemap studies of Fig. 3, which suggested significant similarityin the primary sequence of these pairs of proteins. Thechicken sera raised against matrin 13 showed a relativelyweak signal on Western blots. It did, however, cross reactwith matrin 12 (data not shown) suggesting that proteins 12and 13 may represent another example of a related matrinpair.
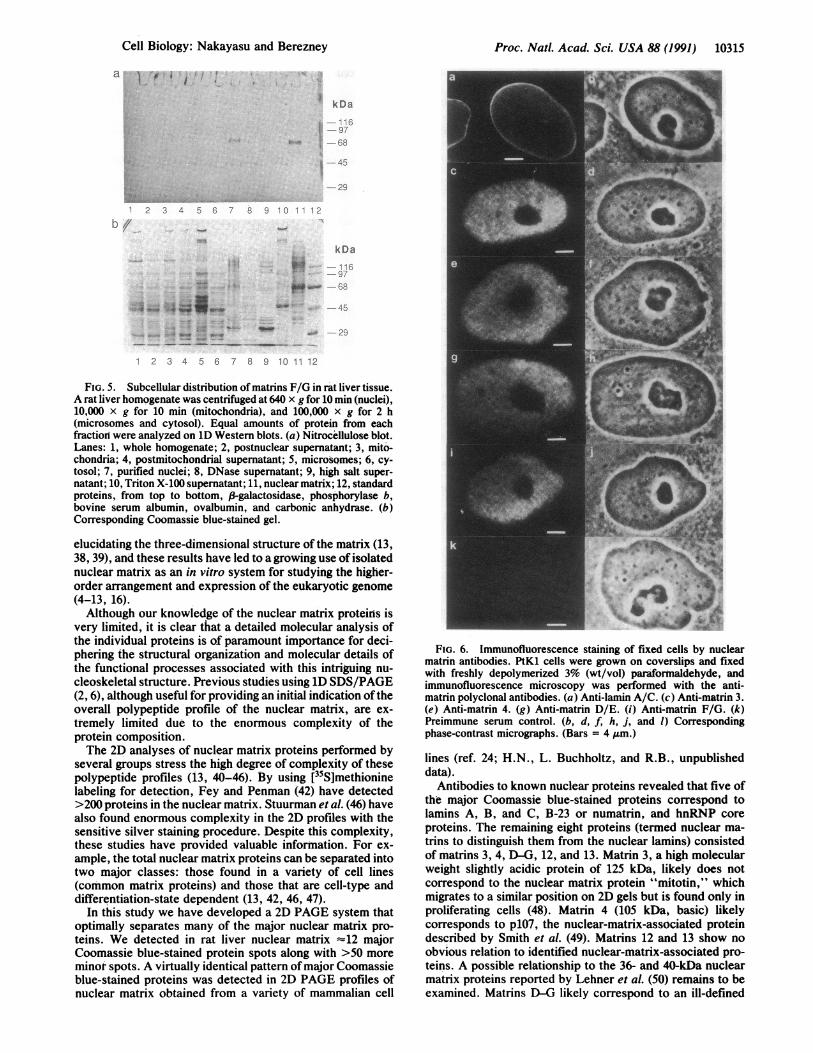
Cellular Distribution of the Nuclear Matrins. The subcel-lular distribution of the nuclear matrins in rat liver tissue wasthen examined on 1D immunoblots (Fig. 5). Matrins F/Gwere detected in isolated nuclei (lane 7), enriched in thenuclear matrix fraction (lane 11), and depleted in the high saltextract (lane 9), which contains the bulk chromatin and mostof the nuclear protein. Matrin F/G was not detected in anyof the other subcellular fractions. The absence of a visible
pH GradientA - - BA 1
a b
iC~laminA Lamin C
e d
FIG. 3. 2D peptide maps of nuclear matrix proteins. Nuclearlamins A and C and matrins D-G were subjected to 2D peptidemapping after trypsin digestion. Peptides unique to a particularprotein are enclosed in a broken line.
a 4B A D F
* a C E G
C
b
d*- (A,C)
e
9
*r (CE)
AP- (AC)
f(0, E)
h-I (F, G) ' (FG)
i (3) '(4~)
FIG. 4. 2D Western blots of nuclear matrix proteins. (a)Coomassie blue-stained proteins. (b) Preimmune serum control. (c)Anti-lamin A. (d) Anti-lamin C. (e) Anti-matrin D. (f) Anti-matrin E.(g) Anti-matrin F. (h) Anti-matrin G. (i) Anti-matrin 3. (j) Anti-matrin 4.
signal in the total liver homogenate fraction was anticipatedsince nuclear proteins are <5% ofthe total protein of the livertissue. Our findings suggest an exclusive nuclear localizationof matrins F/G in the cell and their virtually completeassociation with the nuclear matrix subnuclear fraction.Similar results were obtained for matrins D, E, 3, and 4 (datanot shown).We next examined the structural localization of the nuclear
matrins in whole cells by indirect immunofluorescence mi-croscopy. Unlike lamins A and C, which are located along thenuclear periphery (Fig. 6a), the nuclear matrins are foundexclusively within the nuclear interior with no significantstaining of the nucleoli (Fig. 6 c, e, g, and i). The stainingpatterns for all the nuclear matrins examined appear identicaland consist of a tightly packed network of fibrogranularstructures.
DISCUSSIONNuclear matrix proteins are the nonhistone proteins found inthe nuclear matrix subfraction after nuclease, salt, and de-tergent extraction of isolated cell nuclei (1-6, 13-16). Virtu-ally all known nuclear functions are associated with thisproteinaceous nucleoskeletal structure (1-13, 16). Morpho-logically, the nuclear matrix consists of residual componentsof the nucleolus, nuclear envelope (generally termed thenuclear lamina), and an elaborate fibrogranular network thatextends throughout the nuclear interior and is often termedthe internal matrix (1-6, 14-16). Progress has been made in
Proc. Natl. Acad Sci. USA 88 (1991)
i
Proc. NatL. Acad. Sci. USA 88 (1991) 10315
a I .o i I ,
kDa
I -97
8 -112
8 9 1C 1 1?1
kDa
#0 6Q.
1 2 3 4 5 6 7 8 9 10 1 12
FIG. 5. Subcellular distribution of matrins F/G in rat liver tissue.A rat liver homogenate was centrifuged at 640 x g for 10 min (nuclei),10,000 x g for 10 min (mitochondria), and 100,000 x g for 2 h(microsomes and cytosol). Equal amounts of protein from eachfractiorl were analyzed on iD Western blots. (a) Nitrocellulose blot.Lanes: 1, whole homogenate; 2, postnuclear supernatant; 3, mito-chondria; 4, postmitochondrial supernatant; 5, microsomes; 6, cy-tosol; 7, purified nuclei; 8, DNase supernatant; 9, high salt super-
natant; 10, Triton X-100 supernatant; 11, nuclear matrix; 12, standardproteins, from top to bottom, 83-galactosidase, phosphorylase b,bovine serum albumin, ovalbumin, and carbonic anhydrase. (b)Corresponding Coomassie blue-stained gel.
elucidating the three-dimensional structure of the matrix (13,38, 39), and these results have led to a growing use of isolatednuclear matrix as an in vitro system for studying the higher-order arrangement and expression of the eukaryotic genome(4-13, 16).Although our knowledge of the nuclear matrix proteins is
very limited, it is clear that a detailed molecular analysis ofthe individual proteins is of paramount importance for deci-phering the structural organization and molecular details ofthe functional processes associated with this intriguing nu-cleoskeletal structure. Previous studies using 1D SDS/PAGE(2, 6), although useful for providing an initial indication oftheoverall polypeptide profile of the nuclear matrix, are ex-tremely limited due to the enormous complexity of theprotein composition.The 2D analyses of nuclear matrix proteins performed by
several groups stress the high degree of complexity of thesepolypeptide profiles (13, 40-46). By using [35S]methioninelabeling for detection, Fey and Penman (42) have detected>200 proteins in the nuclear matrix. Stuurman et al. (46) havealso found enormous complexity in the 2D profiles with thesensitive silver staining procedure. Despite this complexity,these studies have provided valuable information. For ex-ample, the total nuclear matrix proteins can be separated intotwo major classes: those found in a variety of cell lines(common matrix proteins) and those that are cell-type anddifferentiation-state dependent (13, 42, 46, 47).
In this study we have developed a 2D PAGE system thatoptimally separates many of the major nuclear matrix pro-teins. We detected in rat liver nuclear matrix =12 majorCoomassie blue-stained protein spots along with >50 moreminor spots. A virtually identical pattern ofmajor Coomassieblue-stained proteins was detected in 2D PAGE profiles ofnuclear matrix obtained from a variety of mammalian cell
FIG. 6. Immunofluorescence staining of fixed cells by nuclearmatrin antibodies. PtK1 cells were grown on coverslips and fixedwith freshly depolymerized 3% (wt/vol) parafortnaldehyde, andimmunofluorescence microscopy was performed with the anti-matrin polyclonal antibodies. (a) Anti-lamin A/C. (c) Anti-matrin 3.(e) Anti-matrin 4. (g) Anti-matrin D/E. (i) Anti-matrin F/G. (k)Preimmune serum control. (b, d, f, h, j, and 1) Correspondingphase-contrast micrographs. (Bars = 4 jum.)
lines (ref. 24; H.N., L. Buchholtz, and R.B., unpublisheddata).
Antibodies to known nuclear proteins revealed that five ofthe major Coomassie blue-stained proteins correspond tolamins A, B, and C, B-23 or numatrin, and hnRNP coreproteins. The remaining eight proteins (termed nuclear ma-trins to distinguish them from the nuclear lamins) consistedof matrins 3, 4, D-G, 12, and 13. Matrin 3, a high molecularweight slightly acidic protein of 125 kDa, likely does notcorrespond to the nuclear matrix protein "mitotin," whichmigrates to a similar position on 2D gels but is found only inproliferating cells (48). Matrin 4 (105 kDa, basic) likelycorresponds to p107, the nuclear-matrix-associated proteindescribed by Smith et al. (49). Matrins 12 and 13 show noobvious relation to identified nuclear-matrix-associated pro-teins. A possible relationship to the 36- and 40-kDa nuclearmatrix proteins reported by Lehner et al. (50) remains to beexamined. Matrins D-G likely correspond to an ill-defined
23 4 5 6
Cell Biology: Nakayasu and Berezney

10316 Cell Biology: Nakayasu and Berezney
cluster of 60- to 75-kDa basic nuclear matrix proteins ob-served in several studies (41, 43, 45, 46). Halikowski andLiew (51) have also identified a presumptive chromatinprotein(s) termed B2 that migrated in the same generalposition as this basic cluster. B2, which is also a highlyphosphorylated protein, was shown to be also associatedwith the nuclear matrix (52).
Peptide mapping and antibody cross-reaction studies indi-cated that the nuclear matrins are distinct from the nuclearlamins. Immunofluorescence microscopy, furthermore, re-vealed a strictly interior nuclear location for the matrins asopposed to the peripherally located lamins. These results areconsistent with Kaufmann and Shaper (45) who reported thatcomponents in the cluster of 60- to 75-kDa nuclear matrixproteins had 1D peptide maps different from the lamins and didnot recognize anti-lamin antibodies on immunoblots. Ourresults further indicate that the four proteins identified in thiscluster form two pairs of related proteins (matrins D/E andF/G). Matrins 12 and 13 may form a third pair and matrins 3and 4 showed no relationship to any of the other matrixproteins.We propose that the nuclear matrins compose a broad
family of structural proteins in the nucleus with potentialsubfamilies indicated by the various protein pair homologues.The actual relationship of each putative protein pair, how-ever, will require more detailed molecular studies. In thisregard, this laboratory has demonstrated that matrins D/Eand F/G specifically bind DNA on 2D Southwestern blots,whereas matins 3, 12, and 13 do not (53). Moreover, thecDNA coding regions for matrins F/G and 3 have beensequenced and are-as predicted from this study-completely unrelated (23, 54). Consistent with their DNAbinding properties, the matrin F/G primary structure con-tains putative zinc-finger DNA binding motifs (54), but noknown DNA binding motif was identified in matrin 3 (P.Belgrader and R.B., unpublished data).We thank the following investigators for their generous gifts of
antibodies: Terrence Martin for antibodies to hnRNP core proteins(iD2), small nuclear RNP proteins (SmY12), and the 70-kDa U1 smallnuclear RNP protein (2.73); Gideon Dreyfuss for antibodies tohnRNP C1 and C2 core proteins (2B12) and a 120-kDa hnRNP protein(3G6); Mark Olson for antibodies to B-23 (numatrin) and C-23(nucleolin). We are extremely grateful to Linda A. Buchholtz andSteven Rosenbloom for their expert technical assistance. Jim Stamosprovided the illustrations. This work was supported by NationalInstitutes of Health Grant GM-23922 (to R.B.).1. Berezney, R. & Coffey, D. S. (1976) Adv. Enzyme Regul. 14,
63-100.2. Berezney, R. (1979) in The Cell Nucleus, ed. Busch, H.
(Academic, New York), Vol. 7, pp. 413-456.3. Shaper, J. H., Pardoll, D. M., Kaufmann, S. H., Barrack,
E. R., Vogelstein, B. & Coffey, D. S. (1979) Adv. EnzymeRegul. 17, 213-248.
4. Berezney, R. (1985) UCLA Symp. Mol. Cell. Biol. 26, 99-117.5. Berezney, R. (1991) J. Cell. Biochem. 47, 109-124.6. Berezney, R. (1984) in Chromosomal Nonhistone Proteins, ed.
Hnilica, L. S. (CRC, Boca Raton, FL), Vol. 4, pp. 119-180.7. Bouteille, M., Bouvier, D. & Seve, A. P. (1983) Int. Rev. Cytol.
83, 135-182.8. Jackson, D. A. (1986) Trends Biochem. Sci. 11, 249-252.9. Gasser, S. M. & Laemmli, U. K. (1987) Trends Genetics 3,
16-22.10. Bodnar, J. (1988) J. Theor. Biol. 132, 479-507.11. Nelson, W. G., Pienta, K. J., Barrack, E. R. & Coffey, D. S.
(1986) Annu. Rev. Biophys. Biophys. Chem. 15, 457-475.12. Nigg, E. (1988) Int. Rev. Cytol. 110, 27-92.13. Nickerson, J. A., He, D., Fey, E. G. & Penman, S. (1990) in
The Eukaryotic Nucleus: Molecular Biochemistry and Macro-molecular Assemblies, eds. Strauss, P. R. & Wilson, S. H.(Telford, Caldwell, NJ), Vol. 2, pp. 763-782.
14. Berezney, R. & Coffey, D. S. (1974) Biochem. Biophys. Res.Commun. 60, 1410-1417.
15. Berezney, R. & Coffey, D. S. (1977) J. Cell Biol. 73, 616-637.16. Agutter, P. S. & Richardson, J. C. W. (1980) J. Cell Sci. 44,
395-435.17. Basler, J. B., Hastie, N. D., Pietris, D., Matsui, S., Sandberg,
A. & Berezney, R. (1981) Biochemistry 20, 6921-6929.18. O'Farrell, P. Z., Goodman, H. M. & O'Farrell, P. H. (1977)
Cell 12, 1133-1142.19. Laemmli, U. K. (1970) Nature (London) 227, 680-685.20. Burnett, W. N. (1981) Anal. Biochem. 112, 195-203.21. Hancock, K. & Tsang, V. C. (1983) Anal. Biochem. 133,
157-162.22. Blake, M. S., Johnston, K. H., Russell-Johns, G. J. & Got-
schlich, E. C. (1984) Anal. Biochem. 136, 175-179.23. Belgrader, P., Dey, R. & Berezney, R. (1991) J. Biol. Chem.
266, 9893-9899.24. Belgrader, P., Siegel, A. J. & Berezney, R. (1991) J. Cell Sci.
98, 281-291.25. Kesser, G. P., Escarla-Wilke, J. & Martin, T. (1984) J. Biol.
Chem. 259, 1827-1833.26. Dreyfuss, G. (1986) Adv. Cell Biol. 2, 459-498.27. Dreyfuss, G., Choi, Y. D. & Adam, S. A. (1986) Mol. Cell.
Biol. 4, 1104-1114.28. Pettersson, I., Hinterburger, M., Mimori, T., Gottlieb, E. &
Steitz, J. (1984) J. Biol. Chem. 259, 5907-5914.29. Billings, P. B., Allen, R. W., Jensen, F. C. & Hoch, S. O.
(1982) J. Immunol. 128, 1176-1180.30. Feurstein, N., Spiegel, S. & Mond, J. J. (1988) J. Cell Biol. 107,
1629-1642.31. Feurstein, N., Chan, P. K. & Mond, J. J. (1988) J. Biol. Chem.
263, 10608-10612.32. Olson, M. 0. J., Wallace, M. O., Herrera, A. M., Marshall-
Carlson, L. & Hunt, R. C. (1986) Biochemistry 25, 484-491.33. Lapeyre, B., Bourbon, H. & Amalric, F. (1987) Proc. Natl.
Acad. Sci. USA 84, 1472-1476.34. Kaufmann, S. H., Gibson, W. & Shaper, J. H. (1983) J. Biol.
Chem. 258, 2710-2719.35. Laiberte, J.-F., Dagenais, A., Filion, M., Bibor-Hardy, V.,
Simard, R. & Royal, A. (1984) J. Cell Biol. 98, 980-985.36. McKeon, F. D., Kirschner, M. N. & Caput, D. (1986) Nature
(London) 319, 463-468.37. Fisher, D. Z., Chandhary, N. & Blobel, G. (1986) Proc. Natl.
Acad. Sci. USA 83, 6450-6454.38. Fey, E. G., Krochmalnic, G. & Penman, S. (1986) J. Cell Biol.
102, 1654-1665.39. He, D., Nickerson, J. A. & Penman, S. (1990) J. Cell Biol. 110,
569-580.40. Peters, K. E., Okada, T. A. & Comings, D. E. (1982) Eur. J.
Biochem. 129, 221-232.41. Verheijen, R., Kuijpers, H., Vooijs, P., van Venrooij, W. &
Ramaekers, F. (1986) J. Cell Sci. 86, 173-190.42. Fey, E. G. & Penman, S. (1988) Proc. Natl. Acad. Sci. USA 85,
121-125.43. Peters, K. E. & Comings, D. E. (1980) J. Cell Biol. 86, 135-155.44. Milavetz, B. I. & Edwards, D. R. (1986) J. Cell. Physiol. 127,
388-395.45. Kaufmann, S. H. & Shaper, J. H. (1984) Exp. Cell Res. 155,
477-495.46. Stuurman, N., Meijne, A. M. L., van der Pol, A. J., de Jong,
L., van Driel, R. & van Renswoude, J. (1990) J. Biol. Chem.265, 5460-5465.
47. Dworetzky, S. I., Fey, E. G., Penman, S., Lian, J. B., Stein,J. L. & Stein, G. (1990) Proc. Natl. Acad. Sci. USA 87,4605-4609.
48. Philipova, R. N., Zhelev, N. Z., Todorov, I. T. & Hadjiolov,A. A. (1987) Biol. Cell 60, 1-8.
49. Smith, H. C., Spector, D. L., Woodcock, L. F., Ochs, R. L. &Bhojee, J. (1985) J. Cell Biol. 101, 560-567.
50. Lehner, C. F., Eppenberger, H. M., Fakan, S. & Nigg, E. A.(1986) Exp. Cell Res. 162, 205-219.
51. Halikowski, M. J. & Liew, C. C. (1985) Biochem. J. 225,357-363.
52. Halikowski, M. J. & Liew, C. C. (1987) Biochem. J. 225,693-697.
53. Hakes, D. J. & Berezney, R. (1991) J. Biol. Chem. 266,11131-11140.
54. Hakes, D. J. & Berezney, R. (1991) Proc. Natl. Acad. Sci. USA88, 6186-6190.
Proc. Natl. Acad Sci. USA 88 (1991)