onion-like glycodendrimersomes from sequence-defined janus
TRANSCRIPT

Onion-like glycodendrimersomes from sequence-defined Janus glycodendrimers and influence ofarchitecture on reactivity to a lectinQi Xiaoa, Shaodong Zhanga, Zhichun Wangb, Samuel E. Shermana, Ralph-Olivier Moussodiaa, Mihai Petercaa,Adam Muncana, Dewight R. Williamsc, Daniel A. Hammerb,d, Sabine Vértesye, Sabine Andrée, Hans-Joachim Gabiuse,Michael L. Kleinf,1, and Virgil Perceca,1
aRoy & Diana Vagelos Laboratories, Department of Chemistry, University of Pennsylvania, Philadelphia, PA 19104-6323; bDepartment of Bioengineering,University of Pennsylvania, Philadelphia, PA 19104-6321; cElectron Microscopy Resource Laboratory, Department of Biochemistry and Biophysics, PerelmanSchool of Medicine, University of Pennsylvania, Philadelphia, PA 19104-6082; dDepartment of Chemical and Biomolecular Engineering, University ofPennsylvania, Philadelphia, PA 19104-6391; eInstitute of Physiological Chemistry, Faculty of Veterinary Medicine, Ludwig-Maximilians-University, 80539Munich, Germany; and fInstitute of Computational Molecular Science, Temple University, Philadelphia, PA 19122
Contributed by Michael L. Klein, December 22, 2015 (sent for review November 14, 2015; reviewed by Timothy J. Deming and Eduardo Fernandez-Megia)
A library of eight amphiphilic Janus glycodendrimers (GDs) withD-mannose (Man) headgroups, a known routing signal for lectin-mediated transport processes, was constructed via an iterativemodular methodology. Sequence-defined variations of the JanusGD modulate the surface density and sequence of Man after self-assembly into multilamellar glycodendrimersomes (GDSs). Thespatial mode of Man presentation is decisive for formation of ei-ther unilamellar or onion-like GDS vesicles. Man presentation andJanus GD concentration determine GDS size and number of bila-yers. Beyond vesicle architecture, Man topological display affectskinetics and plateau level of GDS aggregation by a tetravalentmodel lectin: the leguminous agglutinin Con A, which is structur-ally related to endogenous cargo transporters. The agglutinationprocess was rapid, efficient, and readily reversible for onion-likeGDSs, demonstrating their value as versatile tools to explore thenature of physiologically relevant glycan/lectin pairing.
self-assembly | synthetic multilamellar vesicles | glycolipid mimics
Supramolecular chemistry has enormous potential to helpresolve fundamental questions in the realm of cell biology.
One of the key challenges is the design of programmable modelsfor vesicles and cells and their surfaces as a means of establishinga chemical platform that mimics natural features in size andshape, and also allows customized implementation of bioactiveepitopes, in structural and topological terms. Natural complexitycan conveniently be reduced to simple systems, whose degree ofdiversity can then be rationally reconstituted in a stepwise pro-cess. Focusing on surface properties, the recently gained accessto uniform populations of stable glycodendrimersomes (GDSs)by a simple injection method using a solution of amphiphilicJanus glycodendrimers (GDs) as building blocks for self-assem-bly in a water-soluble aprotic solvent (1–3), has afforded a prom-ising opportunity to realize this concept. Interestingly, the resultingGDSs, which have tunable surface features, can cover the sizerange of naturally occurring vesicles such as endo- and exosomes.When considering the choice of surface determinants, a nat-
ural class of molecules to study comes from the third alphabet oflife, carbohydrates. In fact, glycan structures, unsurpassed incoding capacity relative to proteins and nucleic acids, serve assignals in many bioprocesses, from cell adhesion and growthregulation to diverse transport processes (4–9). Consequently, awide array of synthetic products for the many roles of glycansmust be prepared by diversity-oriented approaches (10–15). Theself-assembly of Janus GDs into GDSs enables analysis of trans(bridging) contacts. Initial experiments with unilamellar GDSshave proven their reactivity with sugar receptors (lectins) (16).Thus, model studies of the impact of glycans on vesicle propertiesand on glycan–lectin recognition are now possible. The studies will
be valuable in clarifying how glycan structure and topology of pre-sentation on cell surfaces team up to achieve the physiological levelof selectivity and specificity of lectin binding to distinct counter re-ceptors (17).In this article, generation of onion-like GDSs is demonstrated
for some D-mannose (Man)-presenting Janus GDs. Of note, theyare structurally related to physiological multilayered vesicles (18–20). The carbohydrate headgroup Man was selected because thissugar is relevant for cellular transport. A group of intracellularcargo transporters are mannose-specific lectins, i.e., ERGIC-53,ERGL, VIP36, and VIPL, and they share the β-sandwich foldand Ca2+ dependence (21–24). However, their nature as type-1membrane proteins hampers studies on GDS aggregation (ag-glutination). Close similarities between the tissue and legumelectins, resulting in their classification as leguminous-like (L type)(25, 26), provide the motivation to run the assays with the tetra-meric leguminous lectin Con A (ConA), a popular model oftenused in glycocluster research (7, 9, 10, 12). The distance betweentwo lectin sites for ConA binding to the surface of the same JanusGDS is about 64 Å or 70 Å, and the cations are bound to ConA tohold amino acid side chains in place for ligand binding (25). Theseaggregation studies unravel the impact of sequence-defined sur-face display of Man on lectin-dependent bridging between vesicles
Significance
The known role of the sugar D-mannose (Man) as a postal codein intracellular cargo routing has herein inspired the design ofMan-presenting synthetic glycolipid-like mimics termed Janusglycodendrimers (GDs). Simple injection of a solution of JanusGDs prepared into a water-miscible solvent into buffer pro-duces, via self-assembly, monodisperse multilamellar onion-like glycodendrimersomes (GDSs). Janus GD structural designimpacts the resulting GDS architecture including surface dis-play of Man. The latter is shown to tune reactivity to a lectin.Thus GDSs provide a model system to enable systematic stud-ies of physiologically relevant glycan/lectin pairing.
Author contributions: Q.X., S.Z., H.-J.G., M.L.K., and V.P. designed research; Q.X., S.Z., Z.W.,and S.E.S. performed research; R.-O.M., M.P., A.M., D.R.W., and D.A.H. contributednew reagents/analytic tools; Q.X., S.Z., S.V., S.A., H.-J.G., M.L.K., and V.P. analyzed data;and Q.X., S.E.S., S.V., S.A., H.-J.G., M.L.K., and V.P. wrote the paper.
Reviewers: T.J.D., University of California, Los Angeles; and E.F.-M., Universidad de San-tiago de Compostela.
The authors declare no conflict of interest.1To whom correspondence may be addressed. Email: [email protected] or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1524976113/-/DCSupplemental.
1162–1167 | PNAS | February 2, 2016 | vol. 113 | no. 5 www.pnas.org/cgi/doi/10.1073/pnas.1524976113
Dow
nloa
ded
by g
uest
on
Dec
embe
r 13
, 202
1

(rate and extent) and on stability of the aggregates under condi-tions of impaired protein–carbohydrate interactions.
Results and DiscussionAn accelerated modular synthetic strategy was used to generate alibrary of eight Man-presenting Janus GDs. The library was ex-tended from the mono- and bivalent and mixed-type headgroups ofthe amphiphilic Janus GDs, previously shown to result in ConA-reactive unilamellar GDSs (27, 28), to five further sequence-defined Janus GDs. Details on their synthesis are shown graphi-cally in SI Appendix, Figs. S1–S3. Methoxytriethoxy fragmentswere used as spacers or space-filling chains to minimize chemicalsurface heterogeneity, except for the lectin-reactive group. Indeed,for various types of lectins, this surface design has been found tobe inert (16, 27–29). Also, the presence of methoxytriethoxy haspreviously been shown to be conducive to the formation of onion-like dendrimersomes (3). This result prompted the use of JanusGDs as building blocks for obtaining onion-like GDSs. As shownin Fig. 1 and SI Appendix, Fig. S5, the surface density of thesugar and its relative positioning on the scaffold result in differentsurface topologies after the Janus GDs self-assembled into bilayers.
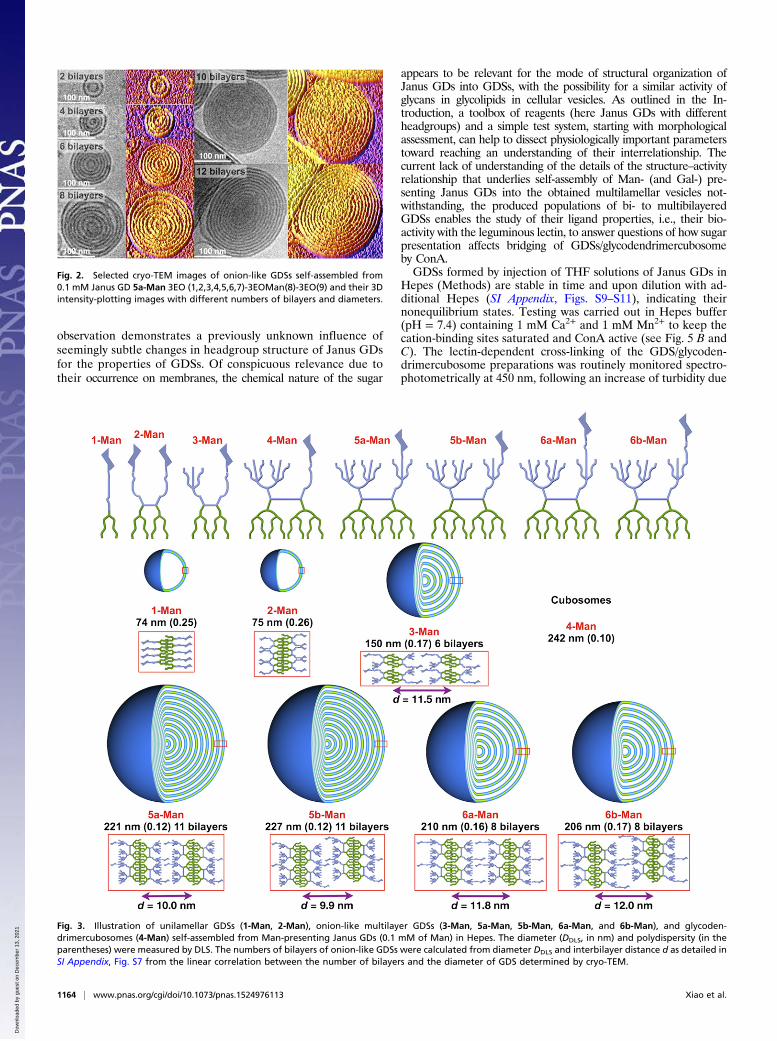
This change on an otherwise constant chemical background wassignificant for yielding new products. The self-assembly process ofseveral Janus GDs of this panel resulted in nonunilamellar vesicles.From 5a-Man onward and 3-Man, onion-like GDSs, resemblingthe structural organization of multilamellar vesicles, were obtained(Fig. 2 and SI Appendix, Fig. S6), and 4-Man formed according tostructural analysis results to be published elsewhere, bicontinuousglycodendrimercubosomes (SI Appendix, Fig. S6).The diameter of the onion-like GDSs is linearly related to the
number of bilayers (Fig. 3 and SI Appendix, Fig. S7) and directlyconnected to the Janus GD concentration (SI Appendix, Table S1and Fig. S8). Overall, the new findings are that the nature andconcentration of a Janus GD’s headgroup determine the size andarchitecture of the GDSs. The various arrangements of the bi-layers and the GDSs resulting from using a fixed concentration of0.1 M Man in Janus GDs are presented in Fig. 3. At the sameconcentration, Janus GDs with D-lactose (Lac) established GDSsthat were invariably unilamellar (16, 29). Intriguingly, when themonosaccharide D-galactose (Gal) was incorporated as part of theamphiphilic Janus GD 3-Gal (SI Appendix, Fig. S4), the occur-rence of several bilayers was noted (SI Appendix, Fig. S12). This
Fig. 1. Summary of Man-presenting amphiphilic Janus GDs. Their diameter (DDLS, in nm) and polydispersity (in the parentheses) were measured by dynamiclight scattering (DLS) with 0.1 mM of Man in 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (Hepes) buffer (10 mM, pH = 7.4).
Xiao et al. PNAS | February 2, 2016 | vol. 113 | no. 5 | 1163
CHEM
ISTR
Y
Dow
nloa
ded
by g
uest
on
Dec
embe
r 13
, 202
1
observation demonstrates a previously unknown influence ofseemingly subtle changes in headgroup structure of Janus GDsfor the properties of GDSs. Of conspicuous relevance due totheir occurrence on membranes, the chemical nature of the sugar
appears to be relevant for the mode of structural organization ofJanus GDs into GDSs, with the possibility for a similar activity ofglycans in glycolipids in cellular vesicles. As outlined in the In-troduction, a toolbox of reagents (here Janus GDs with differentheadgroups) and a simple test system, starting with morphologicalassessment, can help to dissect physiologically important parameterstoward reaching an understanding of their interrelationship. Thecurrent lack of understanding of the details of the structure–activityrelationship that underlies self-assembly of Man- (and Gal-) pre-senting Janus GDs into the obtained multilamellar vesicles not-withstanding, the produced populations of bi- to multibilayeredGDSs enables the study of their ligand properties, i.e., their bio-activity with the leguminous lectin, to answer questions of how sugarpresentation affects bridging of GDSs/glycodendrimercubosomeby ConA.GDSs formed by injection of THF solutions of Janus GDs in
Hepes (Methods) are stable in time and upon dilution with ad-ditional Hepes (SI Appendix, Figs. S9–S11), indicating theirnonequilibrium states. Testing was carried out in Hepes buffer(pH = 7.4) containing 1 mM Ca2+ and 1 mM Mn2+ to keep thecation-binding sites saturated and ConA active (see Fig. 5 B andC). The lectin-dependent cross-linking of the GDS/glycoden-drimercubosome preparations was routinely monitored spectro-photometrically at 450 nm, following an increase of turbidity due
Fig. 2. Selected cryo-TEM images of onion-like GDSs self-assembled from0.1 mM Janus GD 5a-Man 3EO (1,2,3,4,5,6,7)-3EOMan(8)-3EO(9) and their 3Dintensity-plotting images with different numbers of bilayers and diameters.
Fig. 3. Illustration of unilamellar GDSs (1-Man, 2-Man), onion-like multilayer GDSs (3-Man, 5a-Man, 5b-Man, 6a-Man, and 6b-Man), and glycoden-drimercubosomes (4-Man) self-assembled from Man-presenting Janus GDs (0.1 mM of Man) in Hepes. The diameter (DDLS, in nm) and polydispersity (in theparentheses) were measured by DLS. The numbers of bilayers of onion-like GDSs were calculated from diameter DDLS and interbilayer distance d as detailed inSI Appendix, Fig. S7 from the linear correlation between the number of bilayers and the diameter of GDS determined by cryo-TEM.
1164 | www.pnas.org/cgi/doi/10.1073/pnas.1524976113 Xiao et al.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 13
, 202
1

to aggregation with time (up to 400 s). No turbidity increase wasmeasured in the absence of ConA, excluding self-aggregation ofGDSs. A carbohydrate-independent GDS/glycodendrimercubosomereactivity to the lectin was excluded by a lack of change in absor-bance after mixing GDSs/glycodendrimercubosome bearing thenoncognate sugar Lac with ConA (Fig. 4A). This control aspect,i.e., to check for carbohydrate-independent aggregation, wasfurther backed by addition of a β-sandwich protein with reactivitytoward Lac, i.e., human galectin-3 (Gal-3) to Man-presentingGDSs. A measurable turbidity in the solution was not inducedover a period of 1,000 s, although this lectin is an agglutinin (SIAppendix, Fig. S14). Thus, the response was strictly dependent onthe presence of the cognate pair of Man-bearing GDSs withConA, and the presence of inhibitors Man (100 mM)/EDTA(20 mM) completely abolished the activity (please see below).Structural properties other than the binding of Man are thereforenot relevant for ConA-dependent bridging between GDSs. Theformation of such aggregates was determined in terms of theplateau level and the initial rate.
Following confirmation of previously reported data on 1-Man,2-Man, and 3-Man at a constant concentration of Man (27),experiments under identical conditions revealed a strong activityof the glycodendrimercubosome from 4-Man and the onion-likeGDSs consisting of 5a/b-Man and 6a/b-Man, as graded by themolar attenuation coefficient (Fig. 4A). When experiments wererun at a constant size of 145 ± 15 nm and data normalized, asimilar grading was obtained, with 4-Man holding the prominentposition with a coefficient of 40 (Fig. 4B). Among the onion-likeGDSs, relative positioning (sequence) of Man on the scaffold(5a/b-Man) and length of the linker (6a/b-Man) indicate a clearimpact of both sequence and spacer length on agglutination (Fig. 4)and its rate (Fig. 5). This is in in agreement with results on Lac-presenting GDSs (29). In unilamellar GDSs (1-Man, 2-Man) theconcentration of Man on the outside and inside part of assemblycan be statistically estimated to be 50%. However, in multilamellarGDSs the Man concentration on the outside part of the assemblymust be very low. Despite this, multilamellar GDSs with identicaldiameter are much more efficient than the unilamellar assemblies
Fig. 4. Agglutination assays between different Man-presenting GDSs and glycodendrimercubosomes with ConA. (A) Man-presenting GDSs with identicalconcentration (0.1 mM of Man in 900 μL of Hepes, 1.0 mM CaCl2 and 1.0 mM MnCl2), (B) Man-presenting GDSs with identical sizes by using the methoddetailed in SI Appendix, Fig. S8 (145 ± 15 nm, in 900 μL of Hepes, 1.0 mM CaCl2 and 1.0 mMMnCl2) were incubated with ConA (0.5 mg·mL–1 in 100 μL of Hepes,1.0 mM CaCl2, and 1.0 mM MnCl2). The molar attenuation coefficient e = A/(cl), adapted from Beer–Lambert law, where A = plateau OD value, c = molarconcentration of Man, and l = semimicro cuvette path length (0.23 cm). Control experiments were carried out by incubating GDSs generated from 3-Lac3EO(1,2,3)-3EOLac(4), 0.1 mM of Lac in 900 μL of Hepes (1.0 mM CaCl2 and 1.0 mM MnCl2) with ConA (0.5 mg·mL–1 in 100 μL of Hepes, 1.0 mM CaCl2 and1.0 mM MnCl2). Chemical structure of 3-Lac was described in SI Appendix, Fig. S4.
Fig. 5. Rate of change in turbidity, k, of GDSs and glycodendrimercubosomes with ConA calculated from the curves in of GDSs (0.1 mM of Man, 900 μL) withConA (0.5 mg·mL–1, 100 μL) in Hepes (1.0 mM CaCl2 and 1.0 mM MnCl2) as in Fig. 4A at t1/2, where t1/2 is the time at which the observed absorbance is equal tohalf of the plateau absorbance (A). Each rate of GDS was the average value from triplicate measurements. Representative conformation (B) of tetramericstructure of ConA with Ca2+ (green), Mn2+ (purple), and (C) its binding site loaded with methyl α-D-mannopyranoside (Protein Data Bank ID code 5CNA).
Xiao et al. PNAS | February 2, 2016 | vol. 113 | no. 5 | 1165
CHEM
ISTR
Y
Dow
nloa
ded
by g
uest
on
Dec
embe
r 13
, 202
1

(Fig. 4B) although the onions are underestimated. Therefore, thedifference in efficiency is due to Man density, sequence, and spacerlength when compared at identical diameter (Fig. 4B). In contrast,without its implementation into an intimate structural context thatlikely has some spatial flexibility, theMan headgroup of 4-Man is bestsuited to adapt to binding to a lectin, especially when strengtheningan initial contact by a second pairing. In the case of the tetravalentConA, two sites on each side have a fixed distance (Fig. 5B); whenfully loaded with ligand, this yields a contact pattern of two Manmoieties bound by two of the four ConA sites per GDS in an ag-gregate. Such a spatial adaptation process probably takes sometime to occur so that it is reasonable that the rate of change inturbidity was lower for the 4-Man than onion-like GDSs from5a-, 5b-, 6a-, and 6b-Man (Fig. 5A). Conversely, if the lectin alsointroduces spatial adaptability to the system, as natural or engi-neered human tandem-repeat-type galectins with a linker peptidedo, then plateau levels are increased relative to linkerless variants(28) and the difference in the attenuation coefficient is reduced (29).In addition to the concentration- and size-normalized extent
of aggregation and its rate, another parameter of GDS–lectininteraction is the stability of the aggregates in an environmentwith blocking compounds. As mentioned above, no aggregationoccurred when adding a mixture of the cognate sugar for saturatinglectin sites (Man) and the chelating agent (EDTA) for disrupting
the carbohydrate-binding site by cation removal (Fig. 5C). Inter-estingly, a markedly reduced but not lost activity was traced forGDSs from 1-Man/2-Man and 3-Man in the presence of Man butnot the Man/EDTA mixture (Fig. 6 and SI Appendix, Fig. S13).Evidently, a high local density of surface Man can compete with thefree sugar for lectin binding. Fittingly, addition of Man only par-tially led to aggregate dissociation of 1-Man/2-Man (Fig. 6A and SIAppendix, Fig. S13A). Under these conditions, the locked confor-mation of active ConA (26) was apparently sufficiently stable, andEDTA addition only led to a minor, if any, enhancement (Fig. 6Band SI Appendix, Fig. S13B). Slow formation with low plateau levelwas coupled with resistance to aggregate dissociation by inhibitors.An inverted relationship was found in the other aggregation pro-cesses of GDSs from 4-Man onward (Fig. 6 and SI Appendix, Fig.S13). Although secondary interactions cannot be precluded (30), akinetically favored mechanism involving a decrease in the macro-scopic off-rate resembling mucin precipitation by the soybean ag-glutinin (31) can be operative. Lowering cognate sugar density willthen increase the off-rate, a boon for transient recognition phe-nomena, when present in context with physiological inhibitors, e.g.,glycan branches on a cell surface. Alternatively, smaller GDSs(1-Man and 2-Man) may be more resistant to the addition of Man/EDTA due to their larger number of inter-GDSs contacts viaConA linkers. By comparison, larger GDSs may be involved in
Fig. 6. Agglutination assays between the Man-presenting GDSs from 2-Man, 3-Man, 5a-Man, and 6b-Man with ConA. Man-presenting GDSs (0.1 mM of Manin 900 μL of Hepes, 1.0 mM CaCl2 and 1.0 mM MnCl2) were incubated with ConA (0.5 mg·mL–1 in 100 μL of Hepes, 1.0 mM CaCl2 and 1.0 mM MnCl2) with100 mM of Man (blue line) or Lac (red line). A high concentration (100 mM) of Man solution in Hepes (100 μL) was added at t = 50 s into Man-presenting GDSs(0.1 mM of Man in 900 μL of Hepes, 1.0 mM CaCl2 and 1.0 mM MnCl2) with ConA (black line) (A). Man-presenting GDSs (0.1 mM of Man in 900 μL of Hepes,without Ca2+ and Mn2+) were incubated with ConA (0.5 mg·mL–1 in 100 μL of Hepes, without Ca2+ and Mn2+) (blue line) or with Man (100 mM) and EDTA(20 mM) in Hepes (100 μL, without Ca2+ and Mn2+) as inhibitors (red line). Man (100 mM) with EDTA (20 mM) solution in Hepes (100 μL, without Ca2+ and Mn2+)was added at t = 50 s into Man-presenting GDSs with ConA (black line) (B).
1166 | www.pnas.org/cgi/doi/10.1073/pnas.1524976113 Xiao et al.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 13
, 202
1

fewer surface-to-surface interactions and therefore would be ex-pected to agglutinate and deaggregate faster. Clarification of thismechanism will be reported elsewhere. Physiologically, denseclusters can be attained in microdomains, with association of gly-colipids to glycoproteins with their branched N- and O-glycans.Thus, the panel of sequence-defined Janus GDs was instrumentalto trace density as a molecular switch between comparatively slowaggregate formation resistant to dissociation and rapid generationwith high susceptibility to inhibitor presence.
ConclusionsIn the quest to simulate the natural multivalent surface display ofcellular glycans, the self-assembly of specific Janus GDs intoonion-like GDSs is intriguing considering the presence of natu-rally occurring multilamellar vesicles. Importantly, the type ofsugar and the chemical context of its presentation govern thegeneration of the onion-like structures. Results in this articleestablish a basis for systematically studying the influence of thephysiological sugar alphabet on functionality. On GDS surfaces,sugar determinants are presented to facilitate rapid lectin-dependent aggregation, especially when the headgroup is spatiallyadaptable to lectin sites in a fixed constellation. These experi-ments with a leguminous lectin ConA demonstrate the bio-activity of the GDS-surface-presented ligands. They thereforeinspire further experiments with cellular cargo transporters, bi-valent galectin-4 being a prominent candidate (32). The sensi-tivity of the test system, i.e., the aggregation assay, has alreadybeen documented to be high. It can even detect a difference inthe capacity for aggregate formation upon slight changes ofsurface density (by sequence-defined display or by using JanusGD mixtures for GDS assembly) or of protein properties (by
testing natural single-site variants due to polymorphism at thegene level) (28, 29, 33). With regard to cargo transport by GDSs,the proven resistance of proteins such as the β-sandwich galectinsto exposure to an aprotic solvent (34) makes the consideration ofcorresponding GDS loading possible. The versatility of chemicalprogramming of the bilayer composition will enable the discov-ery of novel structure–activity relationships of glycans as struc-tural organizers and as docking sites for tissue lectins on GDSs.
MethodsGDSs and glycodendrimercubosomes were generated by injection of 100 μLof the stock solution of amphiphilic Janus GDs in freshly distilled THF into2.0 mL Hepes buffer, followed by about 5 s vortexing.
Three-dimensional intensity plotting images of multilayer GDSs weregenerated from their cryogenic transmission electron microscopy (cryo-TEM)images by ImageJ (v1.50a) with interactive 3D plotting plugin in invert fireLUT mode.
Agglutination assays of GDSs or glycodendrimercubosomes with ConAwere monitored in semimicro disposable cuvettes at 23 °C at single wave-length λ = 450 nm by using a Shimadzu UV-vis spectrophotometer UV-1601with Shimadzu/UV Probe software with kinetic mode. One hundred micro-liters of Hepes solution of ConA was injected into 900 μL of Hepes solution ofGDSs or glycodendrimercubosomes. The cuvette was shaken for about 2 sbefore starting recording the absorbance change in time, with the sameGDSs or glycodendrimercubosomes solution in reference cuvette. Hepessolutions of ConA were kept at 0 °C ice bath before the agglutination assays.
ACKNOWLEDGMENTS. The authors thank a reviewer for recommending anew agglutination mechanism. Financial support from the National ScienceFoundation (Grants DMR-1066116 and DMR-1120901), the P. Roy VagelosChair at the University of Pennsylvania, and the Humboldt Foundation (all toV.P.), National Science Foundation (Grant DMR-1120901 to M.L.K.), and the ECSeventh Framework Programme (GLYCOPHARM) is gratefully acknowledged.
1. Percec V, et al. (2010) Self-assembly of Janus dendrimers into uniform dendrimersomesand other complex architectures. Science 328(5981):1009–1014.
2. Peterca M, Percec V, Leowanawat P, Bertin A (2011) Predicting the size and propertiesof dendrimersomes from the lamellar structure of their amphiphilic Janus dendrimers.J Am Chem Soc 133(50):20507–20520.
3. Zhang S, et al. (2014) Self-assembly of amphiphilic Janus dendrimers into uniformonion-like dendrimersomes with predictable size and number of bilayers. Proc NatlAcad Sci USA 111(25):9058–9063.
4. Winterburn PJ, Phelps CF (1972) The significance of glycosylated proteins. Nature236(5343):147–151.
5. Gabius H-J (1988) Tumor lectinology: At the intersection of carbohydrate chemistry,biochemistry, cell biology and oncology. Angew Chem Int Ed Engl 27:1267–1276.
6. Bertozzi CR, Kiessling LL (2001) Chemical glycobiology. Science 291(5512):2357–2364.7. Kiessling LL, Splain RA (2010) Chemical approaches to glycobiology. Annu Rev
Biochem 79:619–653.8. Gabius H-J (2015) The magic of the sugar code. Trends Biochem Sci 40(7):341.9. Seeberger PH (2015) The logic of automated glycan assembly. Acc Chem Res 48(5):
1450–1463.10. Gestwicki JE, Cairo CW, Strong LE, Oetjen KA, Kiessling LL (2002) Influencing receptor-
ligand binding mechanisms with multivalent ligand architecture. J Am Chem Soc124(50):14922–14933.
11. Woller EK, Walter ED, Morgan JR, Singel DJ, Cloninger MJ (2003) Altering the strengthof lectin binding interactions and controlling the amount of lectin clustering usingmannose/hydroxyl-functionalized dendrimers. J Am Chem Soc 125(29):8820–8826.
12. Kramer JR, Deming TJ (2010) Glycopolypeptides via living polymerization of glyco-sylated-L-lysine N-carboxyanhydrides. J Am Chem Soc 132(42):15068–15071.
13. Zhang Q, Haddleton DM (2013) Synthetic glycopolymers: Some recent developments.Adv Polym Sci 262:39–60.
14. Munoz EM, Correa J, Riguera R, Fernandez-Megia E (2013) Real-time evaluation ofbinding mechanisms in multivalent interactions: A surface plasmon resonance kineticapproach. J Am Chem Soc 135(16):5966–5969.
15. Ponader D, et al. (2014) Carbohydrate-lectin recognition of sequence-defined heter-omultivalent glycooligomers. J Am Chem Soc 136(5):2008–2016.
16. Percec V, et al. (2013) Modular synthesis of amphiphilic Janus glycodendrimers andtheir self-assembly into glycodendrimersomes and other complex architectures withbioactivity to biomedically relevant lectins. J Am Chem Soc 135(24):9055–9077.
17. Gabius H-J, Kaltner H, Kopitz J, André S (2015) The glycobiology of the CD system: Adictionary for translating marker designations into glycan/lectin structure and func-tion. Trends Biochem Sci 40(7):360–376.
18. Hanson PI, Cashikar A (2012) Multivesicular body morphogenesis. Annu Rev Cell DevBiol 28:337–362.
19. Colombo M, Raposo G, Théry C (2014) Biogenesis, secretion, and intercellular interac-tions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol 30:255–289.
20. Cocucci E, Meldolesi J (2015) Ectosomes and exosomes: Shedding the confusion be-tween extracellular vesicles. Trends Cell Biol 25(6):364–372.
21. Velloso LM, Svensson K, Schneider G, Pettersson RF, Lindqvist Y (2002) Crystal struc-ture of the carbohydrate recognition domain of p58/ERGIC-53, a protein involved inglycoprotein export from the endoplasmic reticulum. J Biol Chem 277(18):15979–15984.
22. Satoh T, et al. (2007) Structural basis for recognition of high mannose type glyco-proteins by mammalian transport lectin VIP36. J Biol Chem 282(38):28246–28255.
23. Zheng C, et al. (2013) Structural characterization of carbohydrate binding by LMAN1protein provides new insight into the endoplasmic reticulum export of factors V (FV)and VIII (FVIII). J Biol Chem 288(28):20499–20509.
24. Satoh T, Suzuki K, Yamaguchi T, Kato K (2014) Structural basis for disparate sugar-binding specificities in the homologous cargo receptors ERGIC-53 and VIP36. PLoSOne 9(2):e87963.
25. Loris R, Hamelryck T, Bouckaert J, Wyns L (1998) Legume lectin structure. BiochimBiophys Acta 1383(1):9–36.
26. Bouckaert J, Dewallef Y, Poortmans F, Wyns L, Loris R (2000) The structural features ofconcanavalin A governing non-proline peptide isomerization. J Biol Chem 275(26):19778–19787.
27. Zhang S, et al. (2014) Mimicking biological membranes with programmable glycanligands self-assembled from amphiphilic Janus glycodendrimers. Angew Chem Int EdEngl 53(41):10899–10903.
28. Zhang S, et al. (2015) Dissecting molecular aspects of cell interactions using glyco-dendrimersomes with programmable glycan presentation and engineered humanlectins. Angew Chem Int Ed Engl 54(13):4036–4040.
29. Zhang S, et al. (2015) Glycodendrimersomes from sequence-defined Janus glyco-dendrimers reveal high activity and sensor capacity for the agglutination by naturalvariants of human lectins. J Am Chem Soc 137(41):13334–13344.
30. Horan N, Yan L, Isobe H, Whitesides GM, Kahne D (1999) Nonstatistical binding of aprotein to clustered carbohydrates. Proc Natl Acad Sci USA 96(21):11782–11786.
31. Dam TK, Brewer CF (2010) Multivalent lectin-carbohydrate interactions energeticsand mechanisms of binding. Adv Carbohydr Chem Biochem 63:139–164.
32. Abad-Rodríguez J, Díez-Revuelta N (2015) Axon glycoprotein routing in nerve po-larity, function, and repair. Trends Biochem Sci 40(7):385–396.
33. Zhang S, et al. (2015) Unraveling functional significance of natural variations of ahuman galectin by glycodendrimersomes with programmable glycan surface. ProcNatl Acad Sci USA 112(18):5585–5590.
34. Siebert H-C, et al. (2000) A new combined computational and NMR-spectroscopicalstrategy for the identification of additional conformational constraints of the boundligand in an aprotic solvent. ChemBioChem 1(3):181–195.
Xiao et al. PNAS | February 2, 2016 | vol. 113 | no. 5 | 1167
CHEM
ISTR
Y
Dow
nloa
ded
by g
uest
on
Dec
embe
r 13
, 202
1