paper fisiologi reproduksi udang laut (penaeid): interaksi
TRANSCRIPT

PAPER
Fisiologi Reproduksi Udang Laut (Penaeid): Interaksi antara faktor lingkungan dengan mekanisme dan kontrol aksi hormon
dan pematangan gonad udang windu.
Oleh: B.M. Laimeheriwa
Pendahuluan
Hormon adalah zat kimia organik yang dibentuk dalam sel atau kelenjar sehat dan normal, disekresi langsung kedalam darah dan dibawa ke sel atau organ target, walaupun jumlahnya sangat kecil tetapi pengaruhnya sangat besar, berperan dalam integrasi dan koordinasi fungsi tubuh. Hormon dihasilkan oleh organ endokrin dengan target organ tertentu yang biasanya dibawa oleh darah dan berperan mempengaruhi tubuh.
Keberhasilan reproduksi udang laut (penaeid), termasuk udang windu, merupakan hal mendasar bagi keberlangsungan biota tersebut. Udang laut, memiliki kemampuan adaptasi yang luas terhadap lingkungan dan memiliki strategi reproduksi yang unik sesuai lingkungannya. Strategi adaptasi dan reproduksi ini, telah menjadi hal yang menarik bagi para ilmuwan untuk mempelajarinya karena sangat berkaitan dengan fungsi fisiologi dan tingkah laku yang diekspresikan secara spesifik untuk setiap individu dalam satu spesies maupun antar spesies.
Pengetahuan tentang fisiologi reproduksi udang laut mulai dari mekanisme konrol hormon, determinasi seks, proses pembentukan sel kelamin (gametogenesis) sampai dengan interaksi antara faktor lingkungan, kontrol aksi hormon hingga terjadinya kematangan gonad sangat perlu informasi yang tepat dan lengkap. Bukan pada level organ sampai pada tingkat jaringan bahkan molekul, tetapi juga dalam kaitannya dengan fisiologi reproduksi itu sendiri. Oleh karena itu, penelitian-penelitian yang berkaitan dengan fisiologi reproduksi sangat dibutuhkan pada dewasa ini.
Tujuan paper ini adalah mengkaji (review) interakasi antara faktor lingkungan dengan mekanisme dan kontrol aksi hormon dan perannya dalam pematangan gonad udang laut khususnya udang windu. Penulis berusaha untuk menelaah informasi secara detail dari beberapa artikel ilmiah dalam kaitannya dengan teori dalam fisiologi reproduksi udang laut. Kiranya, hasil kajian ini menambah pemahaman dan pengalaman penulis sehingga kelak dapat menerapkannya.
Adapun sistematika pembahasan dalam paper ini mencakup: pendahuluan, morfologi dan karakteristik udang laut (windu), organ dan sistem reproduksi pada udang laut, tingkat kematangan gonad udang laut, kontrol aksi hormon pada fisiologi udang laut, peran hormon dalam sistem reproduksi udang laut, serta pengaruh faktor lingkungan pada kadar hormon dan sistem reproduksi udang laut.
Morfologi dan karakteristik udang laut (Penaeid)
Udang windu digolongkan ke dalam famili Peneidae, genus peneus pada
filum Anthropoda. Ada ribuan spesies di filum ini. Namun, yang mendominasi perairan
1

Gambar 1. Udang Penaeid (windu)
Gambar 2. Morfologi udang laut (windu)
2

berasal dari kelas Crustacea. Ciri-ciri kelas Crustacea yaitu memiliki 3 pasang kaki berjalan yang berfungsi untuk mencapit, terutama dari ordo Decapoda.
Secara morfologi, udang windu (Gambar 1 dan 2) terdiri dari kepala yang bergabung dengan dada (chepalothorax) dan perut (abdomen). Kepala udang windu terdiri dari antenula, antena, mandibula, dan sepasang maxillae. Kepala udang windu juga dilengkapi dengan 5 pasang kaki jalan (periopod) yang terdiri dari 2 pasang maxillae dan 3 pasang maxiliped. Perut udang windu terdiri dar 6 ruas dan juga terdapat pasang kaki renang (pleopod) serta sepasang uropod (mirip ekor) yang membentuk kipas bersama-sama telson. Sifat udang windu aktif pada kondisi gelap dan dapat hidup pada kisaran salinitas lebar dan suka memangsa sesama jenis (kanibal), tipe pemakan lambat tapi terus menerus (continous feeder) serta mencari makan lewat organ sensor. Spesies ini memiliki 6 stadia naupli, 3 stadia zoea, 3 stadia mysis dan stadia post larva dalam siklus hidupnya. Stadia post larva berkembang menjadi juvenil dan akhirnya menjadi dewasa (Haliman dan Dian, 2005).
Siklus hidup udang laut (Penaeid)
Secara umum udang kawin di daerah lepas pantai yang dangkal. Prosesnya sangat sederhana meliputi pemasukkan/penitipan spermatophore dari induk jantan ke betina, terlihat pada gambar 3 yaitu siklus kehidupan dari udang Penaeid. Dimana setelah dapat sinyal dari alam maka dengan segera terjadi pemijahan. Sedangkan untuk proses pembuahan atau peneluran biasanya pada daerah lepas pantai yang lebih dalam. Telur-telur dengan segera dikeluarkan dan difertilisasi secara eksternal dalam air. Seekor induk betina mampu menghasilkan sampai satu juta telur setiap bertelur.
Dalam waktu 13-14 jam, telur kecil tersebut berkembang menjadi larva berukuran mikroskopik yang disebut nauplii/ nauplius (Perry, 2008). Pada tahap nauplii tersebut larva akan memakan kuning telur yang tersimpan dalam tubuhnya, kemudian terjadi metamorfosis menjadi zoea. Pada tahap zoea, terlihat larva mulai ke arah permukaan, memakan alga dan setelah beberapa hari bermetamorfosis lagi menjadi mysis. Tahap Mysis mulai terlihat bentuk menyerupai udang kecil dan memakan alga dan zooplankton, dan berenang menuju daerah asuhan yaitu ekosistem mangrove atau estuarin Selanjutnya mysis akan mengalami metamorfosis menjadi postlarva sekitar 3 sampai 4 hari. Tahap postlarva adalah tahap dimana udang sudah mulai memiliki karakteristik seperti udang dewasa. Sedangkan secara keseluruhan proses dari tahap nauplii sampai postlarva membutuhkan waktu sekitar 12 hari. Setelah itu postlarva akan bermigrasi menuju estuarin yang kaya nutrisi dan bersalinitas rendah, dan tumbuh di sana kemudian akan kembali ke laut terbuka saat berukuran dewasa (FAO, 2011).
Organ dan sistem reproduksi udang laut Udang betina memiliki organ eksternal sistem reproduksi yang disebut telikum.
Telikum berguna sebagai tempat untuk menampung sperma yang akan dilepaskan pada saat pemijahan. Telikum terletak antara pangkal kaki jalan ke-4 dan ke-5. Udang windu memiliki telikum yang tidak tertutup oleh lempeng karapas yang keras atau biasa disebut telikum terbuka, sehingga proses perkawinannya tidak didahului oleh molting (Bailey-Brock dan Moss 1992). Udang betina juga memiliki organ internal sistem reproduksi
3

Gambar 3. Siklus hidup Udang Penaeid (Stewart, 2005).
Gambar 4. Organ reproduksi udang laut. A. Jantan. T= testis; VD= vas deferens. B. Betina. O = ovari.
(King, 1948)
4

Gambar 5. Organ repoduksi jantan (lateral view), A. T= testis VD = vas deferens; TA= terminal ampoule.
B. Spermatozoa. C. Petasma (King, 1948)
Gambar 6. Sistem reproduksi udang jantan. A. T=testis; PVD= proximal vas deferens; MVD= medial vas
deferens; DVD= distal vas deferens: TA= terminal ampoule. B. Permukaan ventral udang dewasa. P=petasma; 0=opening of vas deferens.C. Petasma. D. pleopods udang jantan pasangan kedua. AM=appendixmusculina.
5
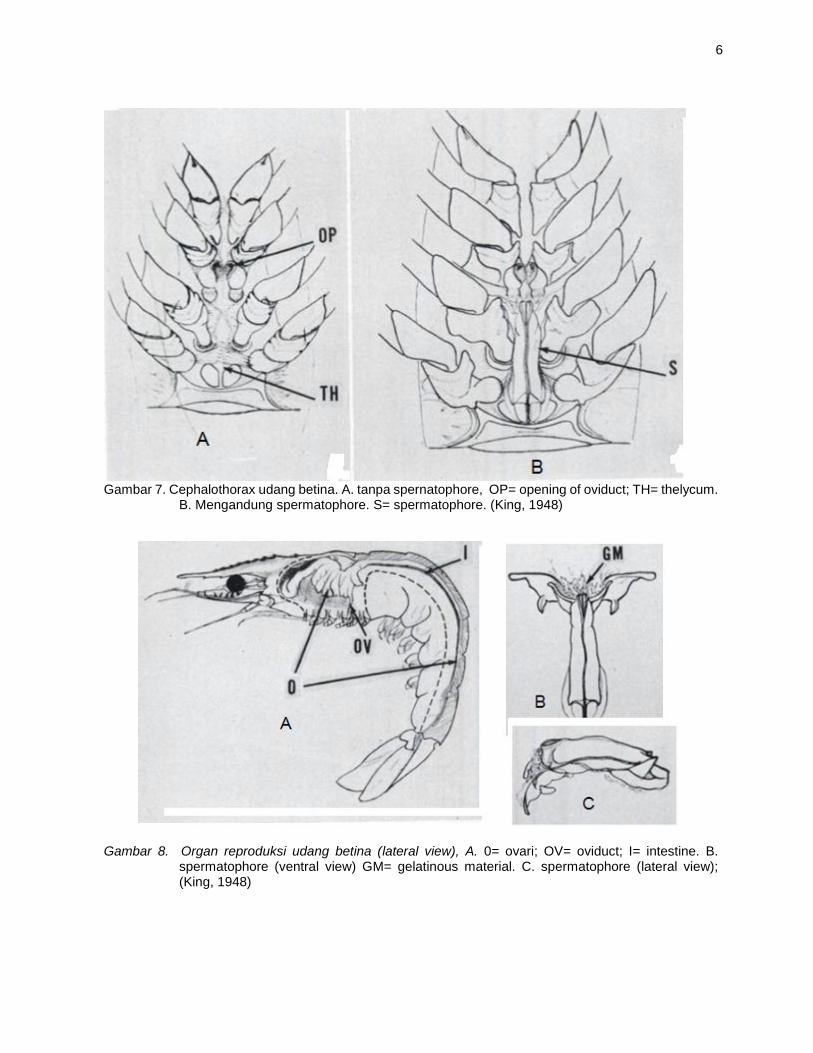
Gambar 7. Cephalothorax udang betina. A. tanpa spernatophore, OP= opening of oviduct; TH= thelycum.
B. Mengandung spermatophore. S= spermatophore. (King, 1948)
Gambar 8. Organ reproduksi udang betina (lateral view), A. 0= ovari; OV= oviduct; I= intestine. B.
spermatophore (ventral view) GM= gelatinous material. C. spermatophore (lateral view); (King, 1948)
6

yang terdiri dari sepasang ovari. Ovari tersebut berbentuk tubular, simetrik bilateral, terletak di bagian ventral hingga rongga dada dan berkembang ke arah posterior hingga hepatopankreas (Gambar 4). Cuping abdominal berdampingan dengan usus dan cuping anterior terdapat di cepalotorax. Cuping lateral berkembang menyamping seperti jari dan terletak antara cuping anterior dan posterior (Tarsim 2007). Oviduk berada diantara kedua sisi ovari dan memanjang hingga organ genital eksternal, yaitu pada koksapodit pasangan kaki jalan ke-3 (Bailey-Brock dan Moss 1992). Pada saat matang, ovari akan tampak berkembang dan memanjang hingga beberapa segmen abdominal.
Sistem reproduksi udang betina terdiri dari sepasang ovarium, oviduk, lubang genital, dan thelycum (Gambar 6 dan 7). Oogonia diproduksi secara mitosis dari epitelium germinal selama kehidupan reproduktif dari udang betina. Thelychum sebagai organ reproduksi betina umumnya terdapat diantara pangkal kaki jalan ke 4 dan 5, yang berfungsi untuk menyimpan spermatofor pada saat terjadi proses pembuahan. Thelycum bersifat terbuka yaitu tidak tertutup oleh lempengan karapas yang keras. Sedangkan petasma pada jantan terdapat pada pangkal kaki jalan ke 5 (Gambar 5). Pada thelycum terbuka proses perkawinannya tidak didahului dengan molting. Organ reproduksi (Gambar 7) betina terdiri dari sepasang ovari berbentuk tubular, simetrik bilateral, terletak pada bagian ventral sampai rongga dada dan berkembang ke arah posterior hingga hepatopankreas (Bailey-Brock dan Moss, 1992).
Perkembangan kematangan gonad udang laut
Tingkat kematangan telur dapat diketahui dari perubahan warna pada ovarinya (kandungan telur), yang berada di bagian punggung udang mulai darichepalotorax (karapas) hingga ke telson (pangkal ekor). Ovari akan berkembang dari yang semula berwarna putih hingga berwarna merah kekuningan (orange) ketika matang gonad. Perubahan warna ovari pada udang windu mudah diketahui, hal ini dikarenakan udang windu merupakan salah satu jenis udang yang memiliki kulit tipis dan jernih. Secara visual, kematangan telur udang windu dapat dibedakan dalam empat tingkatan yaitu: tingkat 1: Gonad tipis, transparan dan tidak terlihat melalui eksoskeleton dorsal (punggung kerangka luar). tingkat 2: Merupakan tingkat kematangan awal dengan terlihat adanya benang halus di bagian punggung udang. Tingkat 3: Gonad tampak lebih tebal dan semakin jelas berwarna merah kekuningan (orange). Tingkat 4: Tingkat kematangan gonad yang siap memijah ditandai dengan ovari yang berwarna merah kekuningan (orange) pada punggung udang darichepalotorax hingga telson.
Sedangkan secara histologi, tingkat kematangan gonad udang jantan dan betina dapat dijelaskan pada Gambar 9 dan 10.
Struktur dan sistem endokrin pada udang laut
Menurut Sunee Wanlem et all (2011), proses fisiologis krustasea, termasuk udang laut diatur oleh neuroendocrines peptida di alam. Crustasea Hyperglycemic Hormon (CHH), sebagai suatu neuroendokrin banyak disintesis oleh organ-X (XO) dan disimpan dalam kelenjar sinus (SG) sebelum dilepaskan langsung ke hemolymph. Organ-X dan kelenjar sinus (XO-SG) yang terletak di ganglia optik di eyestalks udang mensintesis dan mengkontrol hormon neuropeptida untuk pengaturan fisiologi dan
7
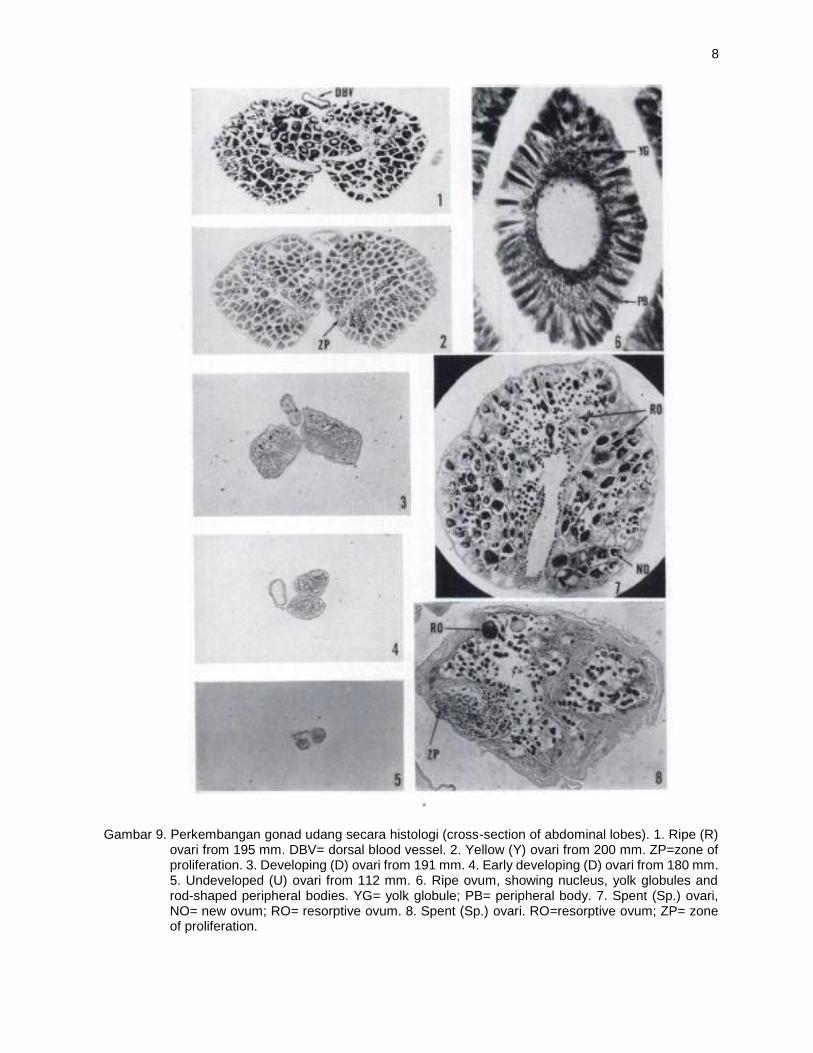
Gambar 9. Perkembangan gonad udang secara histologi (cross-section of abdominal lobes). 1. Ripe (R) ovari from 195 mm. DBV= dorsal blood vessel. 2. Yellow (Y) ovari from 200 mm. ZP=zone of proliferation. 3. Developing (D) ovari from 191 mm. 4. Early developing (D) ovari from 180 mm. 5. Undeveloped (U) ovari from 112 mm. 6. Ripe ovum, showing nucleus, yolk globules and rod-shaped peripheral bodies. YG= yolk globule; PB= peripheral body. 7. Spent (Sp.) ovari, NO= new ovum; RO= resorptive ovum. 8. Spent (Sp.) ovari. RO=resorptive ovum; ZP= zone of proliferation.
8

Gambar 10. Tingkat kematangan gonad udang jantan 1. Section of miniature testis containing spermatogonia and follicle cells. SG= spermatogonium: NC= nutritive cell. 2. Section of mature testis, each tubule containing spermatogonia, primary spermatocytes and secondary spermatocytes. SG= spermatogonium; PS= primary spermatocyte; SS= secondary spermatocyte: NC=nutritive cell. 3. Section of mature testis, primary spermatocytesand secondary spermatocytesin separate but adjoining tubules. SG= spermatogonium; PS= primary spermatocyte; SS= secondary sperniatocyte; F= follicle cell. 4. Section of mature testis, showing spermatids. ST= spermatid. 5. Spermatozoa from medial region of vas deferens. SP= spermatozoan; TP= tailpiece. (King, 1948)
9

metabolisme udang windu. Hormon yang dihasilkan dari XO-SG terdiri dari CHH, MIH, GIH atau VIH dan MOIH. CHH berfungsi terutama dalam regulasi kadar glukosa, metabolisme karbohidrat dan lipid, proses reproduksi dan osmoregulasi. Hormon-hormon yang merangsang sebenarnya dapat menghambat proses-proses tertentu seraya merangsang proses-proses lain. Hormon dapat memberikan efeknya pada struktur-struktur target dengan cara (1) mengubah fungsi gen, (2) mempengaruhi jalur-jalur metabolik secara langsung, (3) mengontrol perkembangan organ-organ spesifik (George et al, 2006).
Secara umum struktur dan sistem endokrin udang laut yang mensintesis hormon pada udang laut ditunjukkan pada gambar 11.
Kontrol aksi hormon pada sistem fisiologi udang laut
Hormon memberikan efek-efeknya pada jaringan-jaringan targetnya, langsung ataupun tidak langsung, melalui pergiliran aktivitas metabolik sel-sel spesifik atau melalui interaksi dengan genom untuk mengaktifkan atau menonaktifkan gen ataupun untuk memodulasi aktivitasnya. Demi melakukan tugas-tugas fisiologis tersebut, hormone harus melakukan penetrasi kedalam selalu menggerakkan serangkaian peristiwa kimiawi setelah melekat ke membran. Sejumlah hormon dapat langsung menyebrangi membran luar dan internal sel, sedangkan sejumlah hormon lain melintasi saluran-saluran yang sudah ada sebelumnya atau menciptakan saluran-saluran baru saat melekat ke sel.
Banyak hormon yang melekat ke reseptor-reseptor spesifik pada membrane sel dari sel-sel target dan memanggil bantuan ‘pembawa pesan kedua’ (second messenger), yakni ‘asisten’ di sitoplasma sel. Ion-ion kalsium diketahui berperan sebagai pembawa pesan kedua. AMP siklik (cyclic AMP, cAMP) adalah contoh lain molekul semacam itu. Ketika sebuah hormone (pembawa pesan pertama) melekat ke sebuah reseptor, hormon itu menyebabkan enzim adenilat siklase mengkonversi ATP menjadi cAMP. Kemudian cAMP tersebut mengaktivasi atau mendeaktivasi system-sistem enzim tertentu yang spesifik bagi sel yang dimaksud, dan dengan demikian mewujudkan fungsi hormon yang melekat ke reseptor sel itu. Sebagai akibatnya, fosfodiesterase memecah cAMP menjadi AMP tunggal, dan karenanya kerja hormonal pun diakhiri. AMP lalu didaur ulang menjadi ATP. Pembawa pesan kedua seringkali mengaktivasi enzim yang merupakan bagian dari sebuah system yang langkah terakhirnya menghasilkan kerja akhir yang sebenarnya dari hormone. Umumnya mekanisme untuk kerja hormon yang satu itu lebih cepat daripada mekanisme yang melibatkan modulasi genom. Modulasi kerja gen mungkin melibatkan peningkatan transkipsi ataupun translasi. Steroid, misalnya glukokortikoid, biasanya berikatan dengan sebuah reseptor protein di sitoplasma, dan kompleks tersebut pun bergerak kedaam nucleus. Didalam nukleus, terjadilah efek yang mempengaruhi mekanisme genetik.
Aksi hormon pada sistem reproduksi udang. Hormon-hormon yang mengatur diferensiasi sifat seksual udang jantan dan betina muncul dari ovari dan kelenjar androgen. Neurosekresi kompleks ganglionik X-organ dan kelenjar sinus dapat menghambat pemasakan ovari dan aktivitas sekretori kelenjar androgen. Diferensiasi normal dari ovari dan testis juga dipengaruhi oleh Y-organ dan hormon molting.
10

Gambar 11. Struktur dan sistem Endokrin pada Udang Windu (Golding dan Pow 1988).
11
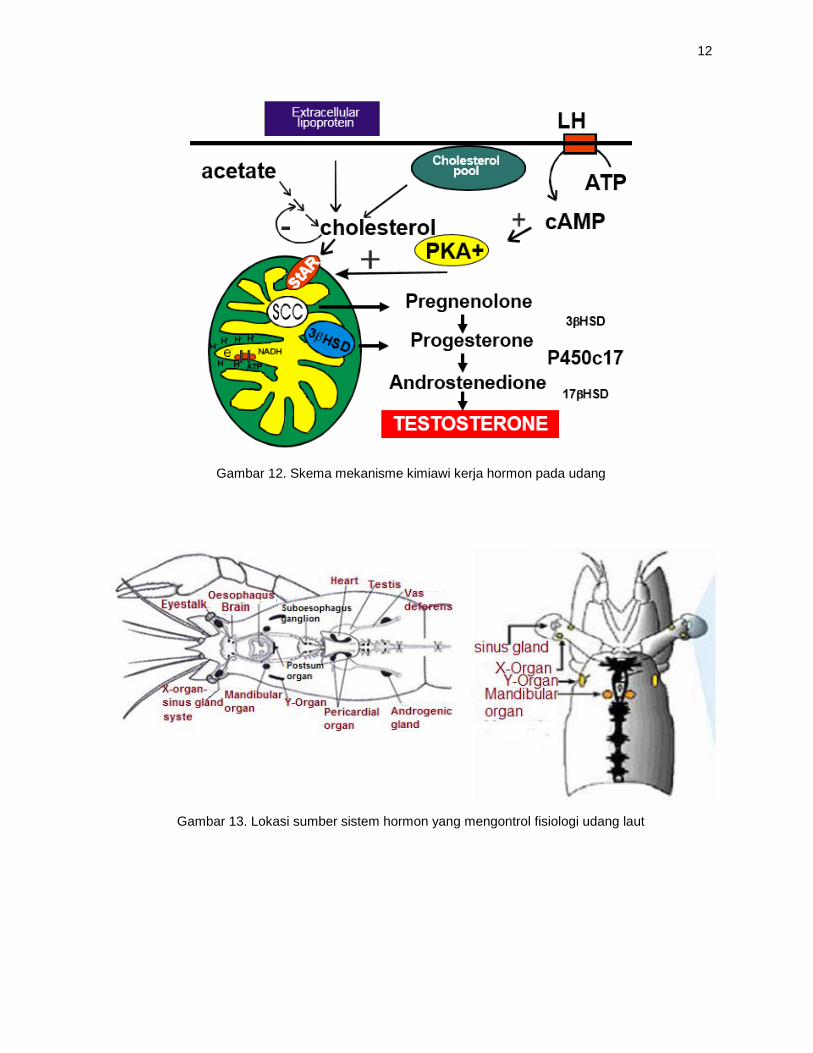
Gambar 12. Skema mekanisme kimiawi kerja hormon pada udang
Gambar 13. Lokasi sumber sistem hormon yang mengontrol fisiologi udang laut
12

Perusakan atau pemotongan tangkai mata dapat mengakibatkan pembesaran ovari dan deposisi kuning telur di dalam oosit. Pemberian ekstrak yang dibuat dari tangkai mata, ganglionik X-organ, atau kelenjar sinus dapat menghambat pembesaran ovari pada udang betina yang memasuki periode aktivitas produksi. Organ X terdapat di dalam tangkai mata (eyestalk). Sedangkan kelenjar sinus merupakan cadangan untuk penyimpanan dan pencurahan neurohormon yang berasal dari akson-akson neurosekretori. Kelenjar sinus utamanya terdiri atas terminal akson, dan erat hubungannya dengan banyak saluran vaskular. Dalam sistem reproduksi, kelenjar sinus menghasilkan Vitellogenesis Inhibiting Hormone (VIH) dan Mandibular Organ Inibiting Hormone (MOIH).
Aksi hormon pada sistem migrasi pigmen retina udang. Mata udang terdiri dari banyak unit yang disebut ommatidia. Secara fungsional, ommatidium memiliki tiga kelompok pigmen yang berbeda yaitu pigmen retina distal, pigmen retina proksimal, dan pigmen putih pemantul. Ekstrak tangkai mata yang dibuat dari udang yang telah diadaptasikan kepada cahaya mengakibatkan adaptasi cahaya pada pigmen distal dan pigmen pemantuk apabila disuntikkan ke dalam resipien yang telah diadaptasikan dalam kegelapan .
Aksi hormon pada sistem ganti kulit (Molting) udang. Pada udang, ganti kulit dapat bersifat musiman atau terus-menerus, tergantung kondisi lingkungan yang beraneka ragam. Perusakan tangkai mata (eyestalk) dapat mempercepat ganti kulit dan pertumbuhan prekoks. Ini terjadi karena ganglionik organ X dan kelenjar sinus yang terdapat di tangkai mata beraksi mencegah ganti kulit. Hormon yang dihasilkan oleh kelenjar sinus dalam sistem ganti kulit adalah Molt Inhibiting Hormone (MIH). Dalam proses ganti kulit, organ Y memproduksi suatu hormon yang melakukan peranan positif. Jumlah air yang digunakan pada saat ganti kulit Water Balance Hormon (WBH) yang dihasilkan oleh tangkai mata.
Aksi hormon pada sistem akselerasi jantung udang. Frekuensi dan amplitudo denyut jantung bertambah seiring dengan pelepasan neurohormon oleh organ perikardia.
Aksi hormon pada sistem kromatoforotrofin dan perubahan warna udang. Kromatoforotrofin merupakan zat pengatur sel pigmen udang di dalam darah. Peran kelenjar sinus dalam sistem kromatoforotrofin dan perubahan warna adalah menghasilkan Red Pigment Concentrating Hormone (RPCH) dan Pigment Dispersing Hormone (PDH). Sistem ini sangat dipengaruhi oleh cahaya Hormon penyebar pigmen (PDH) dan faktor penyebar pigmen (PDF) neuropeptida disimpan dalam tubuhn udang laut. Baik PDF dan PDH diperkirakan hadir sepanjang hidup invertebrata, seperti mereka terdeteksi dari embrio tahap menjadi dewasa. Sistem jaringan PDH/PDF selalu berhubungan dengan visual sistem. Oleh karena itu, tidak mengherankan bahwa tingkat ekspresi biasanya terkait dengan kondisi perubahan pencahayaan lingkungan. Dalam tubuh udang, PDH terlibat dalam perubahan warna yang terkait dengan dispersi pigmen pigmen integumen dan perisai pigmen dalam mata majemuk. PDH tampaknya juga akan terlibat dalam mengatur sensitivitas respon listrik untuk yang berhubungan dengan stimulasi cahaya. Aksi hormon pada sistem metabolisme pada udang. Selama siklus ganti kulit, terjadi variasi mencolok di dalam metabolisme jaringan. Hal ini melibatkan faktor-faktor tangkai mata yang bisa jadi merupakan neurosekresi. Pengambilan tangkai mata mengakibatkan penurunan kadar gula darah dan peningkatan kandungan glikogen
13

hipodermis. Pemberian ekstrak tangkai mata atau kompleks organ X kelenjar sinus dapat menginduksi hiperglikemia . Pada sistem metabolisme, kelenjar sinus berperan dalam menghasilkan Crustacean Hyperglycemic Hormone (CHH) .
Mekanisme kontrol hormonal pada pertumbuhan dan fisiologi reproduksi Udang Windu
Pertumbuhan dan reproduksi udang laut diatur oleh kombinasi hormon neuropeptida, ecdysteroids (hormon molting) dan metil farnesoate isoprenoid (MF). Molting pada udang windu dikendalikan oleh kelenjar kompleks pada tangkai mata (X-organ/ sinus) yang menghasilkan moulting inhibiting hormone (MIH, sebuah neuropeptida yang berperan menghambat produksi ekdisteroid oleh sepasang Y-organ yang terletak dicephalothorax (Mykles et al. 2010). Y-organ merupakan sumber hormon molting, disekresikan sebagai ecdysone, prekursor, untuk hemolymph yang akan dikonversi menjadi hormon yang aktif, 20-OH-ecdysone, oleh aktivitas 20-hidroksilase pada epidermis, jaringan serta organ lainnya (Huberman 2000). Pengaruh MIH di organ Y udang telah banyak diteliti, termasuk pada udang pantai Eropa (Carcinus maenas), udang tanah punggung hitam (Gecarcinus lateralis), dan lobster berduri Afrika Selatan (Jasus lalandii) (Mykles et al. 2010). Ecdysteron.
Ecdysteron merupakan hormon streoid utama pada krustase termasuk udang laut, yang memiliki fungsi utama sebagai hormon moulting (penggantian kulit) dan mengontrol pembentukan exoskeleton baru untuk menggantikan exoskeleton yang lama, selain itu, juga mengatur fungsi fisiologi, seperti pertumbuhan, metamorfosis, dan reproduksi. Penggantian kulit udang laut dapat terjadi secara alami atau dapat diinduksi melalui rangsangan dan penambahan hormon ini (Lafont et al., 2003; Gunamalai et al., 2003).
Ganti kulit (moulting/ecdysis) adalah proses yang sangat penting dalam siklus hidup udang laut sebab pelepasan cangkang adalah suatu keharusan untuk proses pertumbuhan dan metamorfosis (Dorrington, 1979; Fattah, 1998), Rekayasa fisiologi dibutuhkan untuk mempercepat fase moulting dan mendapatkan waktu moulting yang bersamaan. Penelitian untuk menentukan dosis yang dapat memacu terjadinya pergantian kulit pada udang telah banyak dipublikasikan. Informasi ini sangat menunjang dalam pengembangan teknologi budidaya udang windu terutama dalam hal merekayasa fisiologi hormonal untuk mempercepat moulting khususnya dalam produksi skala massal.
Pemberian ECD sebagai moulting stimulan dengan konsentrasi 100 mg/L menghasilkan moulting sebanyak 50%, sedangkan pada konsentrasi 125 mg/L dan 150 mg/L hanya menghasilkan 33,3% saja (Herlinah et al., 2014). Hal ini disebabkan karena kebutuhan ECD untuk stimulasi moulting cukup 100 mg/L saja dan kemungkinan lebih rendah dari konsentrasi tersebut, karena di dalam tubuh udang sendiri sudah memiliki ECD yang dapat digunakan untuk memacu moulting. Gunamalai et al. (2003) mempublikasikan bahwa ecdysterone, sesuai namanya merupakan steroid tulang belakang (steroid backbone) dan sangat berkaitan dengan proses moulting atau yang
14

Gambar 14. Skema kontrol vitellogenesis pada udang betina. MF: methyl farnesoate, MOIH: mandibular
organinhibiting hormone, Vg: vitellogenin, VIH: vitellogenesis-inhibiting hormone, VSH: vitellogenesis stimulating hormone.
15

biasa dikenal dengan ecdysis. Manfaat dari hormon ini selain sebagai hormon moulting juga dapat meningkatkan sintasan karena memberi efek positif terhadap proses metabolisme karbohidrat, lemak, protein, dan tulang.
Penentuan dosis yang tepat sangat penting untuk mendapatkan hasil yang optimal, hal ini sebagaimana yang dijelaskan oleh Fujaya et al. (2012) mengemukakan bahwa hormon dilepaskan pada saat yang tepat dalam jumlah yang wajar pada organisme normal. Kesalahan dalam pemilihan waktu, serta jumlah yang kurang atau lebih akan menimbulkan konsekuensi yang hebat terhadap organisme. Secara spesifik, Dorrington (1979) menjelaskan bahwa konsentrasi hormon yang tinggi menyebabkan produksi reseptor hormon tersebut menjadi terhambat dan kemampuan reseptor sel sasaran dalam mengikat hormon menurun. Reseptor adalah suatu perangkat biologi dalam tubuh yang bertugas mengenali kode yang dibawa oleh suatu hormon. Bila kinerja reseptor menurun maka pembentukan produk baru seperti protein akan terhambat pula. Akibatnya, pertumbuhan dan moulting akan terhambat. Proses pergantian kulit atau moulting merupakan suatu fenomena yang mutlak terjadi pada krustase termasuk udang windu. Namun peristiwa ini tidak berlangsung dalam waktu dan jumlah yang sama dikarenakan adanya perbedaan mekanisme fisiologis yang dipengaruhi faktor internal dan eksternal masing-masing spesies. Dalam kondisi normal, udang muda tentunya akan memiliki kemungkinan yang lebih besar dan sering untuk moulting dibanding udang yang besar ataupun yang telah mencapai kematangan meskipun tidak terlepas dari siklus hormonal (proses fisiologis) yang ada terjadi di dalam tubuh udang tersebut.
Hormon yang berperan dalam siklus pematangan gonad udang windu adalah gonado inhibiting hormone (GIH), yang dihasilkan oleh organ-X serta dilepaskan oleh sinus gland ke sirkulasi darah, dan gonado stimulating hormone (GSH), yang dihasilkan oleh thoracic ganglion dan otak (Sirojini et al. 1995). Gonad-inhibiting hormon (GIH), juga disebut vitellogenesis-inhibiting hormon (VIH) penting dalam menghambat proses vitellogenesis. Bersama-sama dengan molt-inhibiting hormon (MIH), neurohormon ini termasuk dalam keluarga neuropeptida CHH/MIH/VIH. Semuanya diproduksi di sel neuroendokrin dari terminalis medula organ X, terlokalisasi dalam tangkai mata udang (Gambar 13), dan diangkut ke ujung akson cluster dari sel-sel yang membentuk organ aneurohemal, kelenjar sinus. Detail tentang hibridisasi in-situ dan studi immunocytochemical mengungkapkan bahwa sering terjadi co-localization dari dua neurohormonnya dalam tangkai mata (eyestalk) udang (De Kleijn et al. 1998).
Fungsi GIH (Gonad-inhibiting hormon) adalah secara langsung menghambat perkembangan kelenjar androgen pada individu jantan dan ovarium pada individu betina, sehingga spermatozoa atau ovum terhambat perkembangannya. Selain itu, GIH juga dapat mempengaruhi perkembangan gonad secara langsung, dengan cara menghambat aktivitas organ-Y. Sebab apabila organ-Y bekerja aktif, akan menghasilkan GSH yang berfungsi merangsang kelenjar androgen untuk menghasilkan hormon androgen, yang berfungsi merangsang testis untuk menghasilkan spermatozoa pada individu jantan, dan merangsang ovum untuk menghasilkan telur pada individu betina. Apabila konsentrasi GSH meningkat dan konsentrasi GIH menurun dalam sirkulasi darah (hemolymph) udang, maka pematangan ovum akan segera berlangsung.
MIH rekombinan atau sintetis yang diinjeksikan secara in vivo secara signifikan memperpanjang durasi siklus molting, menurunkan titer hemolymph ekdisteroid, atau keduanya (Chen et al. 2007). Analisis sekuen MIH yang dimurnikan dari berbagai
16
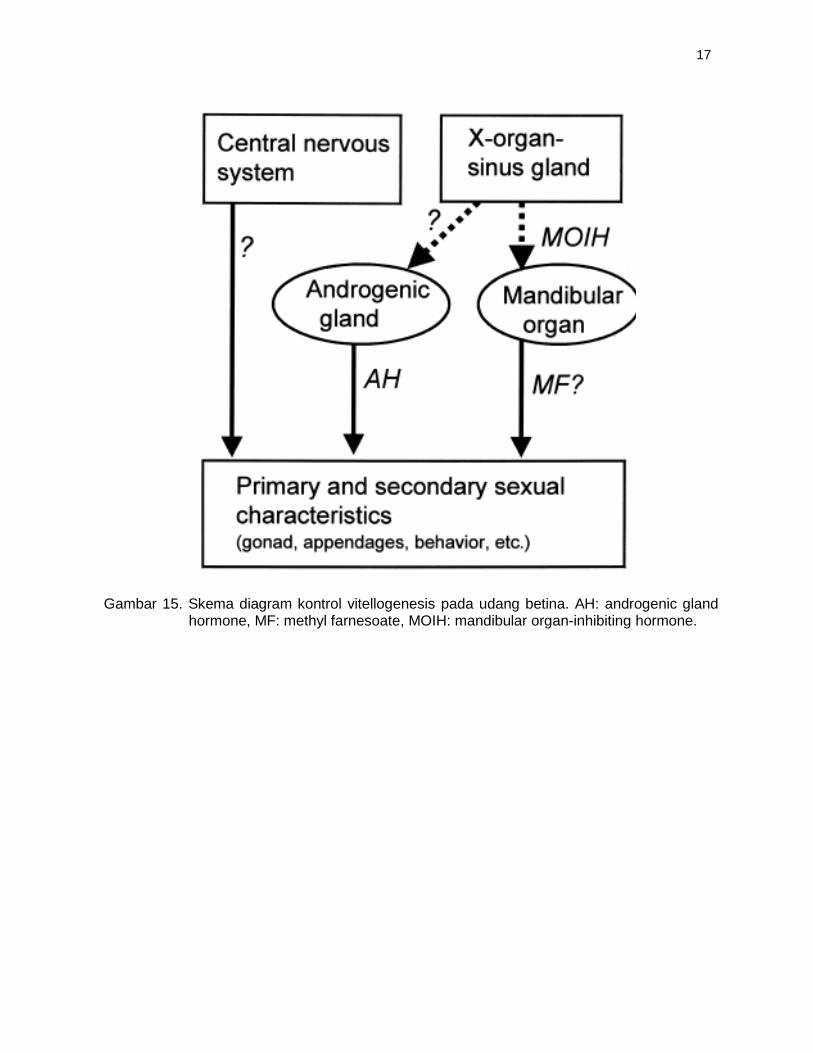
Gambar 15. Skema diagram kontrol vitellogenesis pada udang betina. AH: androgenic gland
hormone, MF: methyl farnesoate, MOIH: mandibular organ-inhibiting hormone.
17

spesies menunjukkan bahwa MIH ini mencapai panjang 74-78 residu dan memiliki 6 residu sistein yang conserved. Berdasarkan urutan homologi, MIH dibagi dalam family crustacea hyperglychemichormone (CHH), yang juga termasuk CHH, vitellogenesis-inhibiting hormone (VIH) atau gonad-inhibiting hormone (GIH), dan mandibular organ-inhibiting hormone (MOIH), serta anggota peptide lain yang ditemukan dalam spesies non-crustacea. Karakterisasi molekuler prekursor MIH menunjukkan bahwa prekursor ini terdiri dari sinyal peptida dan MIH dewasa. Berdasarkan hal ini dan karakteristik sekuen lainnya, diusulkan bahwa MIH, dengan VIH dan MOIH, merupakan sub kelompok tipe II, sedangkan CHH, prekursor yang terdiri dari sinyal peptida, peptida CHH terkait, dan CHH dewasa, membentuk tipe subkelompok (Chen et al. 2007). Hormon gonadotropin (GTH) dan antidopamin (AD)
Pongtippatee et al. (2007) melaporkan bahwa rendahnya tingkat perkawinan udang secara alami yang menyebabkan rendahnya telur yang dibuahi berhubungan dengan kondisi fertilitas induk jantan. Salah satu teknik yang saat ini diaplikasikan untuk meningkatkan tingkat pembuahan telur udang induk tambak adalah dengan inseminasi buatan (Bart et al., 2006; Coman et al., 2007a; Arnold et al., 2012). Teknik ini dapat menghasilkan telur yang fertil (terbuahi) dan larva G2, akan tetapi jumlah larva yang dihasilkan sangat sedikit diduga karena spermatofor yang diimplantasikan terlepas dari telikum induk betina sebelum proses pembuahan terjadi (Laining et al., 2013). Beberapa penelitian melaporkan bahwa pematangan gonad pada udang dan ikan dapat dilakukan melalui rangsangan hormonal seperti gonadotropin-releasing hormones (Tinikul et al., 2014), kombinasi antara gonadotropin dan antidopamin pada udang windui (Yusuf, 2011; Ramdani, 2013), kombinasi antara human chorionic gonadotropin (hCG), salmon pituitary extract (SPE), dan gonadotropin-releasing hormone analogue (GnRHa) pada ikan sidat (Kagawa et al., 2013).
Rangsangan hormonal pada induk udang windu asal tambak untuk merangsang perkembangan gonad dan perkawinan secara alami belum banyak dilakukan. Penggunaan hormon methyl farnesoate (MF) dikombinasikan dengan ablasi mata belum memberikan hasil yang signifikan dalam pematangan gonad dan tingkat perkawinan udang windu hasil domestikasi (Marsden, 2008). Saat ini, rekayasa hormonal pada udang khususnya induk yang berasal dari alam dilakukan khususnya sebagai upaya alternatif dari teknik ablasi mata dalam mempercepat proses pematangan gonad. Kombinasi hormon gonadotropin (GTH) dan antidopamin (AD) secara oral sebagai pengganti ablasi pada induk udang windu menghasilkan 57% induk dapat memijah, masih relatif rendah dibandingkan dengan ablasi yaitu 69% (Yusuf, 2011). Tingkat pemijahan meningkat hingga 67% jika GTH dan AD dikombinasikan lagi dengan rGH, meskipun tingkat pemijahan pada ablasi masih lebih tinggi yaitu 71% (Ramdani, 2013).
Antidopamin adalah bahan kimia yang dapat menghentikan kerja dopamin (prolactin-inhibiting factor, prolactin inhibiting hormone, prolactostatin) yaitu neurotransmitter yang berperan dalam menghambat pematangan gonad udang (Chen & Zhuang, 2003) dan menghambat pelepasan prolaktin pada mamalia. Aplikasi kombinasi GTH dan AD sebagai pengganti ablasi pada udang windu alam perlu dilakukan untuk mengetahui efek hormon ini terhadap pematangan dan produksi naupli. Hasil dari penelitian ini diharapkan dapat dijadikan acuan dalam aplikasinya pada induk
18

udang windu hasil domestikasi baik tambak maupun bak terkontrol untuk menstimulasi terjadinya proses perkawinan alami sehingga jumlah telur fertil yang dihasilkan dapat meningkat. Efek hormon GTH+AD
Penelitian tentang efek GTH+AD dalam perfoma reproduksi udang windu telah dilakukan oleh (Chen & Zhuang, 2003). Tingkat pematangan gonad pascainduksi mencapai 80% pada penelitian ini dan lebih tinggi jika dibandingkan dengan pemberian hormon secara oral pada udang windu (Yusuf, 2011). Hal ini menunjukkan bahwa aplikasi hormon pada udang lebih efektif melalui teknik injeksi dibandingkan secara oral, akan tetapi cara injeksi membutuhkan keahlian tersendiri terutama dalam meminimalkan tingkat stres udang selama injeksi dilakukan. Dengan kata lain, aplikasi hormon secara oral pada skala komersil diduga lebih efisien karena tekniknya mudah dan tidak memberikan dampak stres pada induk, meskipun secara umum pemberian hormon secara oral diperlukan dosis yang lebih tinggi.
Induksi GTH+AD pada dosis 0,3 mL/100 g bobot udang menghasilkan tingkat pemijahan 92% pada injeksi ke-3 dan 80% setelah proses injeksi 4x berakhir menunjukkan bahwa dosis tersebut sudah mampu memacu tingkat kematangan gonad bahkan lebih tinggi jika dibandingkan dengan tingkat pemijahan 60% pada ablasi mata. Dari pengamatan yang dilakukan ternyata induk yang diinduksi hormon, beberapa ekor di antaranya melakukan pemijahan secara parsial. Tingginya tingkat pembuahan telur pada induk pascainduksi GTH+AD (76% vs 54% pada kontrol) mengindikasikan bahwa rangsangan hormonal dapat meningkatkan terjadinya perkawinan alami pada udang windu dalam bak, sehingga hasil yang didapatkan dapat diaplikasikan pada induk udang windu hasil budidaya yang masih mengalami kendala rendahnya tingkat perkawinan alami yang selanjutnya menyebabkan rendahnya tingkat pembuahan telur.
Meskipun jumlah telur yang terbuahi dari induk yang diinduksi hormon lebih tinggi dari ablasi, tingkat penetasan telurnya (%) memperlihatkan bahwa jumlah naupli yang dihasilkan pada ablasi lebih banyak karena tingkat penetasan telurnya lebih tinggi yaitu 52% dibandingkan 39% pada induk yang diinduksi hormon. Hal ini berbeda dengan penelitian yang menggunakan hormon serotonin (5-hydroxytryptamine, 5-HT) pada induk udang windu hasil domestikasi yang menghasilkan tingkat penetasan lebih tinggi dari induk yang diablasi, diduga karena ablasi mata merusak optic lobe dan mengganggu proses fisiologi reproduksi sedangkan efek dari hormon lebih spesifik sehingga tidak mengganggu proses fisiologis lainnya (Wongprasert et al., 2006). Diduga pula 5HT berperan dalam optic lobe untuk menghambat sintesis dan sekresi gonadotrophine inhibitory hormone (GIH) atau berperan di otak dan thoracic ganglion untuk menstimulasi sintesis gonadotrophine stimulating hormone (GSH) (Sarojini et al., 1995; Fingerman, 1997). Selain itu, tingginya total asam amino dalam karkas udang windu yang diinduksi hormon dibandingkan kontrol diduga karena terjadinya perubahan fisiologis pada induk yang diablasi sehingga kemampuan untuk mensintesis asam amino menurun. Secara umum, hasil yang diperoleh pada penelitian beberapa penliti menunjukkan bahwa kombinasi hormon GTH dan AD pada dosis 0,3 mL/100 g induk dapat dilakukan hanya 3x induksi, serta dapat dijadikan alternatif pemacu pematangan gonad induk udang windu selain teknik ablasi mata. Crustacea Hyperglychemic Hormone (CHH)
19

Neuropeptida lain yang terdapat pada tangkai mata yang berhubungan dengan
dengan aktivitas MIH adalah crustacea hyperglycemic hormon (CHH), dinamakan demikian karena perannya dalam meningkatkan kadar glukosa dalam hemolymph. CHH dapat menghambat molting sebagai respon terhadap tekanan lingkungan tertentu. CHH/MIH-like peptide tidak sama seperti yang terdapat pada serangga. Transpor ion oleh CHH-like peptida hanya untuk transportasi Cl di epitel hindgut (Mykles et al. 2010). Sedangkan pada serangga transpor ion peptida (ITP) terjadi untuk Cl-, Na+, K+, dan penyerapan cairan serta menghambat sekresi H+ terjadi pada cardiacum corpus (Huberman 2000). Antara CHH dan MIH saling berkaitan dalam hal pengikatan reseptor independen
Reproduksi crustacea diatur oleh rantai kompleks interaksi hormonal dimana yang memainkan peranan dalam proses ini adalah hormon krustasea hyperglycaemic A dan B (CHH-A dan CHH-B) dan gonad inhibiting hormone (GIH). Neurohormonnya diproduksi di sel neuroendokrin yang sama dengan kelenjar kompleks sinus organ X, yang terletak di tangkai mata. CHH-A dan-B yang terlibat memicu timbulnya vitellogenesis dan CHH-B sendiri, khusus bertanggung jawab untuk merangsang pematangan oosit sebelum pemijahan, sedangkan GIHmencegah awal vitellogenesis di ovarium (De Kleijn et al. 1998).
CHH merupakan neuropeptida paling melimpah di sinus gland. Peran sentral mereka pada pengaturan metabolisme karbohidrat serta berperan dalam metabolisme lemak. Pada udang dan pada lobster, ablasi tangkai mata menyebabkan penurunan lipid total hemolymph, penurunan asam lemak bebas. CHH meningkatkan pelepasan in vitro asam lemak) bebas dan fosfolipid dari hepatopankreas . Sebuah peran yang lebih kompleks CHH di kontrol metabolik ini dibuktikan dengan signifikan mengikat organ-organ yang berbeda seperti hepatopankreas, jantung, epidermis dan Y-organ. Hal ini juga kemungkinan bahwa isomorphs berbeda dari CHH memiliki fungsi dan reseptor yang berbeda. Dalam otot lobster, menunjukkan bahwa CHH menghasilkan ketinggian GMP siklik, dengan aktivasi adenilat membran dan bukan oleh penghambatan fosfodiesterase (Huberman 2000). Adapun urutan sekuen asam amino pada CHH dapat dilihat pada gambar 4. berikut :
CHH memiliki multifungsi karena semua isoform CHH memiliki efek hyperglycaemic, CHH-B dapat merangsang pertumbuhan oosit dan CHH-A dapat menampilkan aktivitas molt-inhibiting. Selain itu,efek ablasi tangkai mata dan implantasi ganglia toraks/abdomen menunjukkan keberadaan suatu hormonvitellogenic-stimulating. mRNA CHH-A dan-B juga terdapat pada bagian sistem saraf selain ganglia optik, yangmenunjukkan bahwa CHH mungkin memiliki peran tambahan dalam kontrol reproduksi dan molting (De Kleijn et al. 1998). Crustacea hyperglycaemic hormon (CHHs) dari kelenjar sinus sistem X-organ (SG) neurosecretory di tangkai mata crustacea terlibat dalam pengaturan glukosa darah dan lipid, sekresi enzim hepatopancreatic, produksi Y-organ ekdisteroid dan transportasi insang ion.
Gonad-Inhibiting Hormone (GIH)
Reproduksi udang windu betina dikendalikan oleh sistem endokrin yang rumit. Kegiatan selular yang terjadi selama perkembangan ovarium disebut proses vitellogenesis, yang merupakan proses dimana vitellogenin (Vg), suatu prekursor
20

protein kuning telur, diakumulasikan di dalam oosit yang sedang berkembang. Vitellogenesis merupakan langkah penting dalam pematangan ovarium. Vg dapat disintesis di ovarium dan / atau situs nonovarian lain seperti hepatopankreas. Sintesis Vg dan pematangan ovarium diatur oleh faktor endokrin tangkai mata disebut sebagai vitellogenesis-inhibiting hormon (VIH) atau gonad-inhibiting hormon (GIH) (Treerattrakool et al. 2008).
Gonad-inhibiting hormon (GIH), juga disebut vitellogenesis-inhibiting hormon (VIH) penting dalammenghambat proses vitellogenesis. Bersama-sama dengan hormon molt inhibiting (MIH), neurohormon initermasuk dalam keluarga neuropeptida CHH/MIH/VIH. Semuanya diproduksi di sel neuroendokrin dari terminalis medula organ X, terlokalisasi dalam tangkai mata crustacea, dan diangkut ke ujung akson cluster dari sel-sel yang membentuk organ aneurohemal, kelenjar sinus. Detail tentang hibridisasi in situ dan studi immunocytochemical mengungkapkan bahwa sering terjadi co-localization dari dua neurohormonnya dalam tangkai mata lobster (De Kleijn et al. 1998).
Dibandingkan dengan CHH dan MIH, hanya sejumlah terbatas GIH telah diketahui. Peptida GIH yang pertama diisolasi secara in vivo berasal dari lobster Amerika Homarus americanus. Peptida lain yang mampu untuk menekan ekspresi Vg mRNA dalam fragmen ovarium adalah Pej-SGP-III Marsupenaeus japonicus. Demikian pula, pendekatan yang sama digunakan untuk pengujian aktivitas VIH pada lobster Procambarus bouvieri. MIH-B dari udang Metapenaeus ensis, meskipun mampu memperluas siklus molting, dapat dianggap sebagai kandidat lain untuk GIH karena tingkat mRNA dari peptida ini menurun tajam selama fase awal pematangan gonad dan meningkatkan terus-menerus saat proses vitellogenic. Encoding cDNA GIH-like peptida juga ditemukan dalam beberapa spesies lain seperti lobster Norwegia Nephropsnorvegicus dan udang Macrobrachium rosenbergii. Hasil penelitian terhadap beberapa jaringan pada P. monodon menunjukkan bahwa terdapat persamaan terhadap ekspresi Pem-GIH pada udang jantan dewasa dan betina dewasa serta pada udang remaja Mandibular Organ-Inhibiting Hormone (MOIH)
Pertumbuhan dan reproduksi udang laut juga diatur oleh kombinasi hormon neuropeptida, ecdysteroids (molting hormon) dan metil farnesoate isoprenoid (MF). MF disintesis dan dikeluarkan dari pasangan organ mandibula, sintesis yang sedang dimodulasi oleh satu atau lebih neuropeptida diproduksi dan dirilis dari X-organ kelenjar kompleks tangkai mata. Pemurnian dan penentuan struktur primer dari 78-residu sinus neuropeptida, mandibular organ-inhibiting hormon (MO-IH-1) dan varian, MO-IH-2, yang menghambat sintesis MF pada udang windu. Peptida ini adalah bagian dari molting-inhibiting hormon (MIH) kelompok dalam crustacea hyperglycaemic hormon (CHH), keluarga neuropeptida. MIH secara negatif mengatur produksi ekdisteroid dalam Y-organ (Tang et al. 1999). Hasil penelitian Tang et al. (1999) yang menganalisis distribusi dan ukuran MO-IH pada jaringan C. pagurus menunjukkan bahwa MO-IH paling banyak ditemukan pada bagian mata dan X-organ sebanyak 950 pasang basa (bp).
21

Interaksi faktor lingkungan, aksi hormonal dan pematangan gonad udang laut
Selain faktor aksi hormon, fisiologi reproduksi udang laut juga sangat dipengaruhi oleh beberapa faktor lingkungan seperti suhu air, kedalaman, intensitas cahaya, fotoperiodisme, dan beberapa faktor biologis seperti densitas aerial dan rasio kelamin (Gambar 16). Perilaku kawin pada udang disebabkan adanya feromon. Udang jantan hanya akan kawin dengan udang betina yang memiliki ovarium yang sudah matang atau matang gonad.
Habitat atau lingkungan merupakan salah satu faktor eksternal yang memiliki peranan penting di dalam reproduksi, terutama pada saat perkembangan gonad. Kondisi lingkungan sangat berpengaruh terhadap metabolisme tubuh. Sinyal dari lingkungan akan diterima oleh susunan saraf pusat dan komplek kelenjar sinus organ X yang ada pada tangkai mata (Gambar 16). Kemudian sistem saraf akan mensekresi hormon-hormon yang berperan dalam reproduksi. Faktor suhu, panjang hari, kualitas dan kuantitas cahaya serta aspek kualitas air mempengaruhi pematangan gonad, kualitas telur dan larva yang dihasilkan. Bahkan aktivasi dari telur-telurnya terjadi akibat kontak dengan air laut (Rojas dan Alvaro, 2006). Proses kawin alami pada kebanyakan udang biasanya terjadi pada waktu malam hari. Peneluran terjadi saat udang betina mengeluarkan telurnya yang sudah matang. Proses tersebut berlangsung kurang lebih selama dua menit. Ibarra et al (2007) melaporkan bahwa udang mampu memijah enam sampai tujuh kali dalam waktu 29 sampai 36 hari.
Dalam budidaya manipulasi lingkungan adalah suatu cara efektif dalam merangsang sekresi hormon untuk mempercepat kematangan gonad. Hoang et al, (2002) menyatakan dalam reproduksi udang telah diketahui bahwa fotoperiodisitas dan suhu berpengaruh terhadap kecepatan perkembangan gonad tetapi hasilnya belum cukup optimal.
Faktor lingkungan seperti salinitas, suhu dan musiman berperan penting dalam mengatur fisiologi udang laut, termasuk reproduksi, perilaku, molting, morfogenesis dan makanan. Peningkatan suhu (32°C), anoksia (0,25 ppm. O2) dan penurunan salinitas diikuti oleh peningkatan yang signifikan dalam kadar hemolymph MF udang laut. Demikian pula, udang stenohaline, menunjukkan peningkatan kadar hemolymph MF saat ditransfer ke salinitas rendah (20 ppt.). Sebaliknya, udang euryhaline, air salinitas rendah (15 ppt.) tidak meningkatkan kadar hemolymph MF (Sagi, 2001).
Koordinasi neuroendokrin dengan faktor lingkungan yang sesuai sangat penting untuk memastikan bahwa sistem reproduksi dan molting tahunan tetap berlangsung. Penyinaran juga berperan kuat dalam reproduksi beberapa spesies udang laut. Pematangan ovarium dan Vg sintesis meningkat secara signifikan di H. americanus ketika penyinaran diberikan. Penelitian menunjukkan bahwa suhu tinggi menyebabkan stres beberapa udang jantan. Suhu 24-27°C menunda spermatophore dan melanisasi, dan 33°C terbukti menjadi akut bagi kualitas sperma (Wilder et.al., 1994).
22
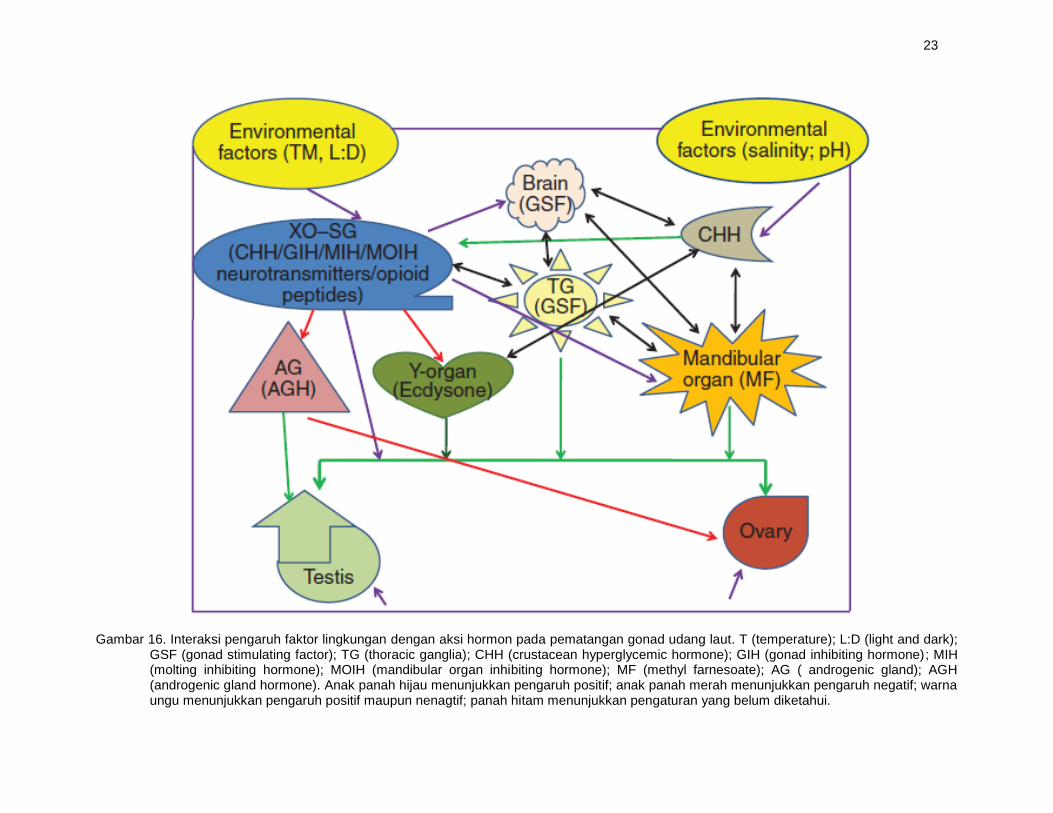
Gambar 16. Interaksi pengaruh faktor lingkungan dengan aksi hormon pada pematangan gonad udang laut. T (temperature); L:D (light and dark); GSF (gonad stimulating factor); TG (thoracic ganglia); CHH (crustacean hyperglycemic hormone); GIH (gonad inhibiting hormone); MIH (molting inhibiting hormone); MOIH (mandibular organ inhibiting hormone); MF (methyl farnesoate); AG ( androgenic gland); AGH (androgenic gland hormone). Anak panah hijau menunjukkan pengaruh positif; anak panah merah menunjukkan pengaruh negatif; warna ungu menunjukkan pengaruh positif maupun nenagtif; panah hitam menunjukkan pengaturan yang belum diketahui.
23

Kesimpulan dan rekomendasi Sistem hormon pada udang laut meliputi: Gonad Inhibiting Hormin (GIH) yang
dilepaskan dai kelenjar sinus, dan Gonad Stimulating hormone (GSH) yang ditemukan pada otak dan thorac ganglion crustacea, MOIH, MF, DA, streroid. Interaksi faktor lingkungan dengan mekanisme kontrol hprmon dalam proses pematangan gonad udang laut sangat penting dipahami dalam menerpakan teknik-teknik reproduksi massal udang laut.
Kontrol sistem endokrin (neuroendokrin dan non-neuroendokrin) dalam sistem reproduksi krustasea menjadi dasar penelitian dan topik yang terus diperdebat. Selain itu, Hormon peptida dan neurotransmiter berpengaruh dalam sistem reproduksi dan non-reproductive udang laut. Fungsi dan perilakunya yang beragam pada organ target, masih belum jelas. Pendekatan dsRNAi /microRNAi memberikan perspektif baru pada GIH atau gonad dari rangsangan peptida dan protein. Penulis inginkan ke depan, perlu studi untuk membandingkan secara integrasi fisiologi reproduksi (seperti salinitas, suhu, dll) dan pengembangan pendekatan klasik untuk menjawab banyak pertanyaan menarik tentang peptida dan protein yang berasal eyestalk, otak dan TG.
DAFTAR PUSTAKA
Azzouna, 1912. Serotonin-likeimmunoreactivityof the ventralnerve cord of the shrimp
Palaemonetesmesogenitor Laboratoire de Biologie de la Reproduction et du Développement, département de Biologie, Faculté Ilmu des de Tunis, 1060 Belvedere, Tunis, Tunisia
Bailey-Brock JH and Moss SM. 1992. Peneid taxonomy, biology, and zoogeography, p. 9-27. In Fast A.W. and L.J. Lester. (Eds). Marine shrimp culture: principles and practices. Development in aquaculture and fisheries science, volume 23. Elsevier Science Publisher. B.V. Netherlands.
Charniaux-Cotton, H. & Payen, G., 1985. Sexual differentiation. In The biology of Crustacea Vol. 9, eds. Bliss, D. E. & Mantel, L. H., Academic Press, Orlando, Florida, USA, 217–299.
De Kleijn, D. P. V. & Van Herp, F., 1998. Involvement of the hyperglycemic neurohormone family in the control of reproduction in decapod crustaceans. Invertebr. Reprod. Dev., 33, 263–272.
Fingerman, M., 1997. Crustacean endocrinology: a retrospective, prospective, and introspective analysis. Physiol. Zool., 70, 257–269.
Hartenstein V. 2006. The neuroendocrine system of invertebrates: a developmental and evolutionary perspective. Department of Molecular, Cell and Developmental Biology, University of California Los Angeles, Los Angeles, California 90095, USA.
24

Huberman, A., 2000. Shrimp endocrinology. A review. Aquaculture, 191, 191–208.
Keller, R., 1992. Crustacean neuropeptides: structures, functions and comparative aspects. Experientia, 48, 439–448.
Laufer H, Takac P, Ahl JSB and Laufer MR. 1997. Methyl farnesoate and the effect of eyestalk ablation on the morphogenesis of the juvenile female spide carb Libinia emarginata. Invertebrate Reproduction Developmant. 31 : 63-68.
Meelkop, L. Temmerman, L. Scchoofs, T. Janssen, 2010. Signalling through Pigment Dispersing Hormone-Like Peptides In Invertebrates. Laboratory of Functional Genomoc and Proteomic, Zoological Institut, K.U. Leuven, Naamsestraat 59, B-3000 Leuve, Belgium.
Meusy, J. J. & Payen, G. G., 1988. Female reproduction in malacostracan Crustacea. Zool. Sci., 5, 217–265.
Nakamura, K., Okumura, T. & Aida, K. 1991. Identification of the Y organ in the kuruma prawn Penaeus japonicus. Nippon Suisan Gakkaishi, 57, 1463–1468.
Ohira, T. et al. 1999. Expression of a recombinant moltinhibiting hormone of the kuruma prawn Penaeus japonicus in Escherichia coli. Biosci. Biotechnol. Biochem., 63, 1576–1581.
Okumura, T. & Aida, K.. 2000 Hemolymph vitellogenin levels and ovarian development during the reproductive and non-reproductive molt cycles in the giant freshwater prawn, Macrobrachium rosenbergii. Fish. Sci., 66, 678–685.
Okumura, T. et al. 2003. In vitro secretion of ecdysteroid by Y-organ during molt cycle and evidence for secretion of 3-dehydroecdysone in the giant freshwater prawn, Macrobrachium rosenbergii (Crustacea: Decapoda: Caridea). Invertebr. Reprod. Dev., 44, 1–8.
Okuno, A. et al. 2002. Preparation of an active recombinant peptide of crustacean androgenic gland hormone. Peptides, 23, 567–572.
Sagi, A. & Khalaila, I. 2001. The crustacean androgen: a hormone in an isopod and androgenic activity in decapods. Am. Zool., 41, 477–484.
Sartaj Ahmad Allayie, S. Ravichandran, Bilal Ahmad Bhat, 2011. Hormonal Regulatory Role of Eyestalk Factors On Growth of Heart in Mud Rrab, Scylla serrata. Marine Biology, Annamalai University, Parangipettai 608502, India.
Tsukimura, B. 2001. Crustacean vitellogenesis: its role in oocyte development. Am. Zool., 41, 465–476.
Wilder, M. N. et al. 1994. Vitellogenin production induced by eyestalk ablation in juvenile giant freshwater prawn Macrobrachium rosenbergii and trial methyl farnesoate administration. Zool. Sci., 11, 45–53.
25