relation of methemoglobin to hernolysis - blood
TRANSCRIPT

Relation of Methemoglobin to Hernolysis
By BYRoN B. CLARK, Pii.D., AND ROBERT W. MORRISSEY, M.I).
Jl�ith the Tech nical A ssistance of DOROTHY BLAIR
IT IS \VELL KNO\VN that a number of chemical agents, w’hieh under certain
circumstances cause intravascular hemolvsis an(l the development of hem-
olytic anemia, also produce methemoglobinemia. Some examples of these agents
are stllfanilam ide, phenylhydrazine, aniline , acetanilid, p-aminophenol , plas-
mochin, etc. Such a correlation may lead to the inference that intracorpu.scular
methemoglobin is related in some way to the process of hemolysis. It has been
observed that. the dosage level of acetanilid and acetophenet.idin causing signifi-
cant anemia also exceeds the threshold for methemoglobin formation.’ The data
of Rimington and Hemmings2 might also he interpreted to lend some additional
support to the view’ that methemoglobin plays some positive role. In studies on
drugs related to sulfanilamide, they observed a close correlation between the
action of aromatic amino compounds to increase porphyrin excretion and to
form met.hemoglohin. Since the increased porphyrin excretion in rodent.s was
due largely to coproporphyrin III, it has been suggested that this represents
an abnormal pathw’ay for the degradation of hemoglobin possibly through met-
hemoglobin.2 ‘ � However, Rimington and Hemmings2 w’ere not certain as to the
role of the methemoglobin in either the increased erythrocyte destruction or the
increased porphyrin excretion since the latter w’as not observed after sodium
nitrite administration, which readily produces methemoglobin. Fox and Otten-
berg4 studying the hemolytic anemias from sulfonamides in man observed a
greater percentage of methemoglohin in the serum than in the erythrocytes and
suggest that it ��‘as t.he cells containing methemoglobin w’hich underwent he-
molysis. This discrepancy betw’een serum and cells could be readily accounted
for by the presence in serum of oxidants w’ith low’ penetrability into the eryth-
rocytes, or by different equilibrium conditions, as in fact the high percentage of
met hemoglobin observed in the urine suggests may have been the case.
On the other hand, there are some reasons to doubt that methemoglohin per
se favors hemolysis. Hemolytic anemias have not l)een reported as a feature
of nitrite toxicity, an(l as will be seen, w’e have not. observed it in experiments
designed to test this point. Also Keilin and Mann5 state that intracorpuscular
hemoglobin may be readily changed to methemoglobin with sodium nitrite with-
out altering the integrity of the cell membrane. It is w’ell known that certain
drugs may produce a hemolytic anemia without the formation of methemo-
globin. Furthermore, the alteration of hemoglobin to form carbon monoxide
hemoglobin has been show’n not to alter the osmotic fragility of erythroeytes.6
However, these are insufficient reasons to decide w’hether methemoglobin
From the Department of Pharmacology, Tufts College Medical School, Boston, Massa-chusetts, and the Department of Physiology and Pharmacology, Albany Medical College,
Union University, Albany, N. Y.
A preliminarY report on this subject appeared in Federation Proceedings, 4: 114, 1945.
532
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom

CLARK AND MORRISSEY 533
plays any definitive role in the process of hemolysis. There is a paucity of any
quantitative data on the sul)ject. Therefore a comprehensive study has been
ma(le in order to provide quantitative data with highly sensitive t.echnics. A
comparison has been made in vitn’o of the resistance of oxyhemoglobin and
methemoglobin containing ervthrocytes to hemolysis by saponin, hypotonic
saline, l)ile salts and lysolecithin. Also included are oi)servations on the hemato-
logic effects in vivo of chronic methemoglobinemia produced by sodium nitrite.
EXPERIMENTAL METHODS
In order to carry out. these experiments, it waS necessary to use some chemical agent
to l)roduce intracorpuscular n�ethensoglobin. The requirenients for such an agent are (1)
that it forms n�ethemoglobin in vitro as well as iii vivo, (2) that it can readily penetrate theerytlirocyte niembrane and (3) that it Produces no injury to the nien�branse in the concen-
trrttions required to form relatively high concentrations of niethemoglobins. These require-
nienits eliniiniate many aromatic coniipounds, some of which at least act as sin�l)le h’sins or
accelerate certain lvsin.s. After i1U�iC�OUS �)relirnin1ary experiments sodium nitrite was
chosen as the niost suitable agent availal)le. The nit rite readily penetrates the erythiro-
cyte and fornis n�ethiemoglobin ra�)idly at loss- conicenitrations.7 In the procedure of pre-
�arinsg the erythrocyte suspenisionis, there was no gross evidence of hemolysis :Issociate(iwitl’i the presence of the nitrite ; after repeated washings si-it hi saline no t race of nit rite
could he detected.7’ 8 Furthern�ore, the intentional additions of low ct)nicenltrations of so-
diun� nit rite I 0 the lytic system (under t hose condit ions inn which oxyhenioglobin arid niet-
hen�oglohini cells hemolyzed in tine same time) did not significantly alter the time required
for hennolysis, indicating that this agent did not act to an iniportanit degree as an aced-
erator or inhibitor of the lvsins used. Keilin arid Mann5 have also presented some evi(lencethat siniilar treatn�ent of er�throc�tes does riot n�oclifv their J)ernseability.
Erythi’ocyte suspensions. Dog erythrocytes were eniployedi for all of the data to he re-
ported. However, human erythrocytes were also employedl in a sufficient number of experi-ments to establish the qualitative similarity of the results. The standard cell suspensions
��‘ere prepared ins the folloiving ‘�-:�‘. A sanll)le of 1)100(1, about 4 cc., s�as (Irawn into a ceni-
trifuge tube containing 25 cc. of 0.85 l)d� cent sodium chloride anti 250 mg. of sodium cit-
rate. This s:tline-cit.rate suspension was (livided into two portions; one l)ortion WHS treated
with sodium nitrite, arid 10 mg. �er cc. of blood was allowed! to stand for ten minutes dur-
ing which time about 80 to 90 per cent of the hemoglobin was converted to methemoglobin.
When lower percentages of met hemoglobin were desired proport ionately smaller amounts
of sodium nitrite were added. The dells of both portions were then washed three times by
cent rifugat ions and resuspension irs 30 cc. of saline per cc. of packed cells. The suspensions
were carefully standardized in the final dilution with saline to contain 0.8 Gm. of total
hemoglobin per 100 cc.,9 which corresponded approximately to 250,000 cells per cu.mm.These two cell suspensions will be referred to as “oxyhemoglobin cells” and “methemo-
globin cells.”
All experiments except t.hose with hypot.onic saline were carried out in buffered saline.
Sorensen’s M/l5 J)hiosphate mixtures were used in proportion of 1 l)art in 10 of 0.85 per
cent saline to obtain the desired p11 indicated in the experiments; the initial p11 before
hemolysis for a sufficient number of experiments was checked by the glass electrode.
Saponin. The time required for complete hernolysis was determined for a sUspension of
washieti erythrocytes and saponin ins a saline-phosphate buffer with the tubes in a constant
temperature water bath according to the general method described by Ponder.’#{176}�’t The
hemol�’tic system consisted of 2 cc. of buffered saline, 2 cc. of the saponin dilution in buf-
fered saline and 1 cc. of the standard cell suspension in soft. glass test tubes 12 X 100 mm.
A series of dilutions of saponin were employed to obtain time-dilution curves. The dilu-
tions of saponin indicated in the figures are the final dilutions iii the hemolytic system.Hypotonic Saline. Percentage hemolysis curves were obtained with a series of dilutions
of hypotonic saline using the general technic described by Hunter.” The erythrocyte sus-
pensions were prepared as described above except that the final dilution was that of the
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom

534 RELATION OF METHEMOGLOBIN TO HEMOLYSIS
original volume of blood. One-tenth cc. of such a suspension was ad(ledi to 10 cc. of each
dilution of hypotonsic saline. After standing for one hour at. room tensperature, the series
of tubes were centrifuged and the liberated hemoglobin determined9 in the supernatant.
Bile Salts. The same general technic for 100 per cent hemolysis as describetl for saponin
was used. Both sodium glycocholat.e and taurocholate were tested and yielded qualita-
lively similar results, but only the data on glycocholate will he presented.
Lysolecithin. The lysolecithin used was prepared by the actions of snake venom* on egg
yolk, aS described l)�’ King and Dolan.’2 The material obtained was l)rohahly somewhat
impure, since no attempt was made to separate out the lysocephalin, but nevertheless pro-
vided a preparation of constant activity. For use a stock solution of lysolecithin was pre-
pared each day containing 10 mg. per cc. in absolute nsethyl alcohol from which appropriatedilutions were made with saline and kept in paraffined test tubes.’3 The standiard suspen-
sions of ervthroc�tes were prepared in the same way as described ah)ove.
The time required for Sf) per cent hemolysis was used as the end point for this lysin;
this could he estimated accurately from the time per cent hemolysis curves which were
obtained by an adaptation of the photoelectric method described by Wilbur and Collier.’3
The hemolytic system consisted of 8 cc. of buffered saline, 1 cc. of the standard cell sus-
pension and 1 cc. of the lysolecithin dilution in saline. The dilutions indicated in the data
are the final dilutions of the lysin in the colorimeter tube. The per cent hemolysis curves
were obtained using the Evelyn colorimeter with filter 660.� The galvanometer was set at
100 for the one hundred per cent hemolysis blank containing the above system of the cor-
responding type of cells which were completely hemolyzed with saponin. The initial opac-it)’ reading for zero per cent hemolysis was obtained for both kinds of cells with the above
system in which the lysolecithin solution was replaced by saline. Such control tubes were
prepared for each pH and temperature used. (In most. cases it was impractical to obtain
an initial opacity reading following the addition of the lysolecithin because of the initial
rapid rate of hemolysis with concentrations of lysin yielding fifty per cent hemolysis in a
reasonable time.) In the experimental tubes following the addition of lysolecithin, galva-
nometer readings were recorded at appropriate time intervals. The per cent of hemolysiswas read directly from semi-log graph paper, using the opacity reading of the unhemolyzed
control tube to establish the point for zero hemolysis.
RESULTS
Saponin. Time-dilution curves for 100 per cent hemolysis were determined
with saponin in parallel experiments on both oxyhemoglobin and methemo-
globin cells. The average results in 16 parallel experiments at. pH 7.3 and 38 C.
are show’n in figure IA, and indicate that the presence of methemoglobin in the
erythrocytes has no effect on their resistance to saponin lysis. Qualitatively
similar results were obtained at pH 6.2 and 38 C. and also at 25 C. pH 6.2 and
7.3, and with human eryt.hrocytes. The addition of free oxyhemoglobin and met-
hemoglobin initially to the system had a small hut equal inhibitory effect on the
subsequent saponin hemolysis.
Another series of experiments, using both the methemoglobin and oxyhemo-
globin cells, were performed in an identical manner except. that. chemical agents
know’n to accelerate saponin lysis were included in equal concentrations in both
systems. Thus, each experiment consisted of four time-dilution curves; that is,
oxyhemoglobin and methemoglobin cells each in the presence and absence of
t.he accelerator. The results are illustrated in figure 1B, C, and D. The accelera-
t.ors varied from the highly active quinine to the relatively weak acetanilid.’4
The curves obtained for the oxyhemoglobin and methemoglobin cells were es-
* Crotalus atrox venom kindly supplied by Sharpe and Dohme.
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom

A B
QUININE-O.125’,M/L
z
�
C D
ACETANILID- 12 MM/L
0
(‘LARK AND MORRISSEY 535
sentially identical. Similar data ss’ere obtained w’ith several other accelerators,
such as aniline, acetophenetidin, and p-aminophenol; the latter is the only agent
listed which forms met.hemoglobin in vitro under t.he condit.ions of these experi-
ment.s. (Sulfanilamide ss’as found to be a w’eak inhibitor and to affect both types
of cells to the same degree.) The results clearly indicate that t.he presence of
methemoglobin does not alter the time required for hemolysis of erythrocytes
by saponin alone or combined with an accelerator.
zz0Q.
U,
‘I.
0
B(NZCNE- 0-45 MM/L
2 4 6 8
TIME IN MINUTES
FIG. 1-Effect of methemoglobin on saponin hemolysis at. pH 7.3 and 38 C.
o Oxyhemoglobins cells#{149}Met.hemoglobin cells
L�s Oxyhemoglobin cells plus accelerator
A Methensoglobin cells plus accelerator
Ilypo#{252}inic saline. Percentage hemolysis curves w’ere obtained for a series of
dilutions of hypotonic saline at room temperature (about 25 C.). Ten parallel
experiments on both oxyhemoglohin and methemoglobin cells were made, and
the averaged percentage of hemolysis for each concentration of saline appears in
figure 2. These data indicate that t.he presence of methemoglobin does not
alter the resistance of the erythrocytes t.o hemolysis by hypotonic saline.
In a second series of experiments a sub-lytic concentration of quinine (i.e., a
concent.ration producing no hemolysis in 0.85 per cent. saline in one hour) s�’as
included in the hypotonic saline before the addition of the erythrocytes. As
shown in figure 2, the presence of quinine increases the percentage of hemolysis
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom

536 RELATION OF METHEMOGLOBIN TO HEMOLYSIS
for a given concentration of saline, but again no difference is evident. in the per-
centage of hemolysis of the oxyhemoglobin and methemoglobin cells.
Bile salts. The experiments with the bile salts w’ere carried out using the same
technic for 100 per cent hemolysis as s�’as described for saponin. Most of the ex-
periments were made at 38 C. w’ith sodium glycocholate. In preliminary experi-
ments, it was found that the relative rates of hemolysis of the oxyhemoglobin
and methemoglobin cells differed according to the pH of the system. This rela-
tionship is illustrated in figure 3, which represent.s times for hemolysis for the
tw’o types of cells by 1: 2000 glycocholate over a range of pH values. Below’ pH
‘00
80 //
En /
> /-J #{163}0� 60 /
uJ /
I Es’
O 40 #{163}5,,’
/� ./
.62 � .46 .42 1
PER CENT SALT CONCENTRATION
FIG. 2.-Effect of methemoglobin on hypotonic saline hemolysis at 25 C.
o Oxyhemoglobin cells#{149}Methemoglobin cells
� Oxyhemoglohin cells plus quinine
A Methemoglohin cells plus quinine
6.6, hemolysis of the methemoglobin cells takes place at. a slightly iiicreased
rate. On the other hand, above pH 7.0 the time required for the hemolysis of
the methemoglobin cells becomes excessively long as compared to the oxyhemo-
globin cells. The two curves cross at approximately pH 6.7. The general rela-
t.ionship evident at pH 6.2 and 7.3 was also observed to hold for a series of dilu-
tions of both glycocholate and taurocholate. The inhibition of methemoglohin
cells at pH 7.3 and their slight acceleration at pH 6.2 was found to be roughly
proportional to the methemoglobin concentration in the cells.
In view’ of the reversal of the rates of hemolysis of the two types of cells with
respect to pH, it seemed advisable to determine w’hether solutions of free oxy-
hemoglobin and methemoglobin differed in their inhibiton’y activity. A portion
of the buffered saline in the hemolytic system was replaced by buffered hemo-
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom
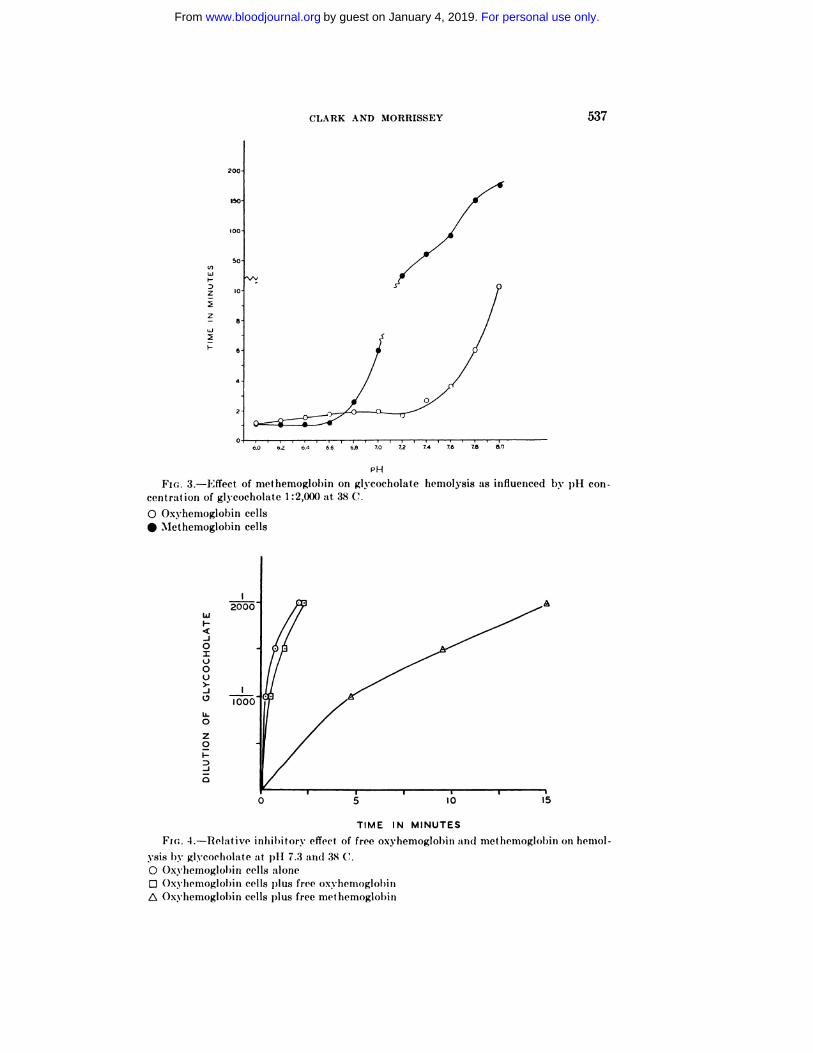
CLARK AND MORRISSEY 537
200
150
00
50
I0
InL�7
z
2z
U2I-
8
6
2
60 62 6.4 6.6 6.8 7.0 7.2 7.4 7.6 7.8 60
hemolysis as influenced by pH con-
wI-
-J
0I000
-I
0LI.
0z0
I-
-J
5 0
FIG. 3.-Effect of nsethensoglobini on glycocholate
centration of glycocholate 1:2,000 at 38 C.
o Oxyhemoglohin cells
#{149}Methemoglobin cells
TIME IN MINUTES
FIG. 4.-Relative inhihitor�’ effect of free oxyhemoglobin arid met hensoglohin on hennol-
ysis by glycocholate at p11 7.3 arid 38 C.
o Oxyhennoglobin cells aloneD Oxyhennoglobi ni cells plus free oxyhemoglobi n
� Oxyhemoglobi n cells plus free met hemoglobin
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom

538 RELATION OF METHEMOGLOBIN TO HEMOLYSIS
glohin solutions equivalent to 10 per cent of the hemoglobin contained in the
cells in the system. It was found as show’n in figure 4 that at pH 7.3, where the
methemoglobin cells were apparently resistant to hemolysis, that t.he addition
of a solution of methemoglobin to the oxyhemoglohin cells caused a much greaterinhibition than the addition of oxyhemoglohin in a similar system. At. pFI 6.2
the tw’o types of hemoglobin had very little inhibitory effect. The same qualita-
tive results for pH 6.2 and 7.3 were also obtained at 25 C. It therefore seems
probable that the apparent increased resistance of the methemoglobin contain-
200
80
60
UI 140
-j
0
120
I
U� 100
0
0 80z0U
60
2
I- 40
20
20 25 30 35 38
TEMPERATURE
FIG. 5.-Effect of methemoglobin on lysolecithin hemolysis as influenced by pH and tern-
l)erature.
o Oxyhernoglobin cells at pH 7.3
L�s Oxyhemoglobin cells at pH 6.2
#{149}Methemoglobin cells at pH 7.3
A Methernoglohin cells at pH 6.2
ing cells at pH 7.3 is due to the greater inhibitory effect of the init.ially released
methemoglobin on further hemolysis.
Lysolecithin. In these experiments, the time required for fifty per cent he-
molysis was determined as described in the section on methods. The results with
lysolecithin were found to be influenced in a rather complex way by pH and tem-
perature, w’ith t.hese factors determining whether the oxyhemoglobin and met-
hemoglobin cells hemolyzed in the same or different times.
The influence of varying the pH from 6.0 to 8.0 at 25 and 38 C. was deter-
mined for a 1:200,000 dilution of lysolecithin. At 25 C. and between pH 6.8 to 7.6
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom

CLARK AND MORRISSEY 539
the oxyhemoglobin and methemoglohin cells hemolyzed at. the same times. How’-
ever, at pH 6.0 to 6.8 at. 25 C. and at all pHs tested at 38 C. the methemoglobin
cells consistently hemolyzed more rapidly than the oxyhemoglobin cells. From
this data pH 6.2 and 7.3 were selected for a further study of the effect of tem-
perature; an example of the results obtained with a 1 :200,000 dilution of lysolec-
it.hin are illustrated in figure 5. From the above data and figure 5, it. may be said
that around pH 7.3 and 25 C. 1)0th the oxyhemoglobin and methemoglobin
cells hemolyzed in approximately the same time, and more extensive data show
that there is no significant. difference. How’ever, under several other conditions
of pH and temperature, the methemoglobin cells hemolyzed more rapidly.
In other experiments these differences in the hemolysis times of oxyhemoglobin
and methemoglohin cells w’ere found to he proportional to the methemoglohin
concentration. As might have been predicted, at pH 7.3 and 25 C., varying the
methemoglol)in concentration resulted in no significant difference in the he-
molysis time. However, for the other conditions tested (pH 6.2 at. 25 C. and pH
TABLE 1 -Effect of Free Henioqlobin on Lijsolecithin Heinolysis
(Lvsolecithini dilutions 1 : 200,000)
25U 38USystem � � � � �. � . ________
_______________________________ I pH 6.2 � pH 7.3 � pH 6.2 � pH 7.3
Hb02 Cells � 77 � 24 � 47 � 90
11l)O� Cells + 11h00 120 35 � 80 � 180
Hb02 Cells + MlIb � 65 � 35 I 45 � 100
MIIb Cells � 22 � 28 � 13 � 20
Time ins seconds for 50 per cent hemolysis.
6.2 and 7.3 at 38 C.), increasing the methemoglobin concentration caused a
proport ional acceleration of hemolysis.
An attempt. to explain these varying results suggested the possibility that free
oxyhemoglobin and methemoglobin initially released might cause differing (IC-
grees of inhibition of subsequent hemolysis, especially since an example of this
had already been found in the case of the bile salts. This possibility was tested
by comparing the relative inhibitory effects produced by replacing a portion of
the buffered saline in the system by buffered hemoglobin solutions equivalent
to 20 per cent of the hemoglobin contained in the cells in the syst.em. Represen-
tative results are show’n in table 1. It is evident that at 25 C. and pH 7.3, where
the oxyhemoglobin and methemoglobin cells hemolyze at approximately the same
rate, the addition of free oxyhemoglohin and methemoglohin caused the same
degree of inhibition. However, for the other conditions under which the methemo-
globin cells hemolyzed more rapidly, free oxyhemoglobin caused a much greater
degree of inhibiton than did free met.hemoglobin; in fact., the latter at times
caused some acceleration, especially at 25 C. and pH 6.2. These differences in
the inhibitory action of the tw’o kinds of hemoglobin were more easily demon-
strat.e(I using oxyhemoglobin cells, although t.hey were evident in a certain num-
her of the experiments using methemoglobin cells. These data suggest that the
observed differences in the inhibitory action of the two kinds of hemoglobin
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom

540 RELATION OF METHEMOGLOBIN TO HEMOLYSIS
may account for the difference noted in the rates of hemolysis of the oxyhemo-
globin and methemoglobin cells.
Some experiments were also made with lysolecithin prepared from dog blood
as described by Singer’5 using the method w’e have outlined for “synthetic”
lysolecithin. In a general sense the results w’ere similar, but. the conditions under
which they occurred w’ere different. At 38 C. and pH 6.2 and 7.3, there ss’as no
definite difference in the rate of hemolysis of oxyhemoglohin and methemoglobin
:: �
04
F-LUa 0
� : ________________ ________ ____
40 #{149}1��
0 0 20 30
DAYS
FIG. 6.-Flematologic effects of chronic niethemoglohinemia maintainned by the daily
administration of sodium nitrite.
Each character represents an individual dog. The vertically arranged characters at the
left are control determinations.
cells. However, at 25 C. and both pH 6.2 and 7.3, the methemoglobin cells
hemolyzed more rapidly than the oxyhemoglobin cells.
J7lernak3logic effects of methemoglobinem.ia in rico. Chronic methemoglobinemia
was maintained in 5 dogs over a period of thirty-five days by the administration
of sodium nit.rite subcutaneously as a 5 per cent solution once daily. The hemo-
globin pigments w’ere measured by the method of Evelyn and Malloy.9 The
dosage of nit.rite ��‘as adjusted to obt.ain a maximal met.hemoglobinemia ranging
between 50 and 70 per cent each day. These maximal values w’hich occurred
about tw’o hours after the administration of nitrite are represented in figure 6;
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom

CLARK AND MORRISSEY 541
thereafter, the methemoglobinemia progressively decreased so that none was
present. the following morning when the blood samples were obtained for the
other ol)servations recorded. The data summarized in figure 6 indicate that no
significant alterations occurred in the values for total hemoglobin, erythrocyt.es,
and leukocvtes. There s�’as a small l)ut significant increase in ret.iculocytes, and
in some cases a small increase in cell volume. The reticulocytosis indicates an
increased erythropoiesis that may be a response to increased erythrocyte (lestruc-
tion. If so, it must have been so small as to be covered by an increased erythro-
(yte l)rO(Iuctiofl, for there is no evidence of anemia. An alternative explanation
for the reticulocytosis appears more likely. It has been previously observed in
this lahorator’�’16 that this degree of methemoglobinemia causes some reduction
in the venous oxygen tension, and this could act as a stimulus to the hone mar-
l�O\\’. Ibis view is strengthened i�y the observation that an additional increase
in l)��)th total hemoglobin and reticulocytes occurs shortly after the nitrite ad-
ministration when the methemoglobinemia is greatest. Furthermore, Rimington
and Hemmings2 have ol)served no increased porphyrin excretion follow’ing nitrite
a(lminist ration . �I�If , t hese (lata indicate that chronic methemoglobinemia
maintained by nitrite cause no significant anemia, and suggest that no significant
decrease in erythrocyte resistance to the natural in vivo hemolysins occurred.
DIscussIoN
rI�le l)niflcilMtl l)UEI)OSC of this paper was to determine whether the change in
the composition of inti�acorpuscular hemoglobin to methemoglobin may influence
the response of the erythrocyte to hemolysins. As outlined in the introduction,
the reasons for assuming that met.hemoglobin may have some influence are chiefly
circumstantial ; it s�’as also indicated that there were insufficient data on which
to base a (lecision.
The in VitV() experiments with saponin and hypotonic saline indicate that
intracorpuscular methemoglobin does not influence hemolysis by these lysins.
Neither (lid methemoglobin appear to alter the susceptibility of the erythrocytes
to the (oml)ined action of these lysins plus chemical accelerators.
The in vitro results w’ith bile salts and lysolecithin hemolysis w’ere more com-
plex; it s�’as found that the presence of methemoglobin either did or did not in-
fluence hemolysis depending upon the experimental conditions of pH and tem-
perature. It seems significant, how’ever, that. for 1)0th lysins there are conditions
(pH and temperature) under which the times required for hemolysis of oxy-
hemoglobin and methemoglobin containing eryt.hrocytes are the same. Also
significant are the findings that for those conditions under which the hemolysis
times were different., free oxhemoglobin and methemoglobin (extracorpuscular
hemoglobin added initially to the hemolyt.ic system) had different inhibitory
effects on sul)sequeflt hemolysis, and that these differences �s’ere directly cor-
related with the differences in the behavior of the oxyhemoglobin and methemo-
globrn cells. It is difficult to quantitate these effects. However, qualitatively such
results do permit. the reasonable assumption that the difference in hemolysis
of the oxyhemoglobin and methemoglobin containing erythrocytes may be ex-
plained On the basis of the different degrees to which the originally released
oxyhemoglobin and methemoglobin inhibits subsequent. hemolysis. If this is an
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom

542 RELATION OF METHEMOGLOBIN TO HEMOLYSIS
adequate explanation, then the observed data with bile salts and lysolecithin
could be accounted for; and the interpretation is permitted that intracorpuscular
met.hemoglobin has not altered the resistance of the erythrocytes to hemolysis
by bile salts and lysolecithin. A full explanation of the different degrees of in-
hibitory effect of oxyhemoglobin and methemoglobin for these lysins under cer-
tam experimental conditions cannot be given at present.
rFhe in vitro results w’ith saponin and hypotonic saline seem to indicate very
clearly that intracorpuscular methemoglobin does not. alter the resistance of
erythrocytes to hemolysis by these lysins. The results with bile salts and lyso-
lecithin are capable of a similar interpretation for the reasons outlined in the
preceding paragraph, although admittedly the results are more complex, and
there may be other interpretations, but the one given seems the most reasonable
from a consideration of the experimental data as a whole. The in vivo experiments
with chronic met.hemoglobinemia produced by the daily administration of sodium
nitrite did not produce a significant anemia, and this together w’ith the reported
absence of increased porphyrin excretion2 suggests that no significant decrease
in erythrocyte resistance to the intravascular lysins occurred.
rfhe experiments present strong prima-facie evidence that int.racorpuscular
methemoglobinemia per se is not primarily concerned in the process of int.ra-
vascular hemolysis. It is suggested that in those cases ��‘here methemoglobin
formation is associated w’ith the action of chemical agents producing hemolysis
or acceleration of lysins that methemoglobin formation is not an integral part
of the hemolytic action, but is an independent effect indicating the presence of
oxidants capable of penetrating the erythrocyt.e. Several such substances have
been studied in this laboratory and found to he active accelerators of variouslysins; hsow’ever, under specified conditions there �s’as no difference in the degree
of action on erythrocytes containing principally oxyhemoglobin as compared to
erythrocytes containing 80 to 90 per cent methemoglobin.
SUMMARY
Experiments are described which attempt to define the relation of met.hemo-
globin to hemolysis. A comparison w’as made in vitro of t.he resistance of eryth-
rocytes containing chiefly oxyhemoglobin or methemoglobin to hemolysis l)y
saponin, hypotonic saline, bile salts and lysolecithin. These data are interpreted
to indicate that int.racorpuscular methemoglobin does not alter the resistance
of erythrocytes to hemolysis by these lysins, although there are certain peculi-
arities in the results with bile salts and lysolecithin.
In the case of bile salts and lysolecithin, oxyhemoglobin or methemoglobin
may have an equal or different inhibitory effect on hemolysis depending upon
the conditions of pH and temperature.
Chronic methemoglobinemia in dogs maintained by the administration of
sodium nitrite does not produce an anemia.
These data suggest that the formation of methemoglobin is an independent
effect. and not a part of the hemolytic action of certain chemical agents.
REFERENCES
1 VAN Loox, E. J. AND CLARK, B. B.: Observations on the hernatological actions of acetani-
lid and acet.ophenetidin in the dog. J. Lab. & Clin. Med. �9: 942, 1944.
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom

CLARK AND MORRISSEY 543
2 Rn�nrNGToN, C. AND HEMMINGS, A. W. : CXVIII. Porphyrinuric action of drugs related to
sulfanilamide. Comparison with reported toxicity, therapeutic efficiency and causations
of methemoglobinemia. Definition of the structure responsible for l)orl)h�’rinuric act ion.Biochem. J. 38: 960, 1939.
3 BROWNLEE, G. : The role of the aromatic amino group in deranged l)igflieflt metal)olism.
Biochem. J. 33: 697, 1939.
4 Fox, C. L. AND OTTENBERG, R. : Acute hemolytic anemia from the sulfonsamides. J. Clin.Investigation 20: 593, 1941.
5 KEILIN, D. AND MANN, T. : Activity of carbonic anihydrase within red blood corpuscles.
Nature, Lonidon 148: 493, 1941.
6 MAYERS, M. R., RIVKIN, H. AND KRASNOW, F.: The effect of chemically pure carbon
monoxide, illuminating gas, and automobile exhaust gas �poni the fragility of the
red blood cells. J. Indust. Hyg. & Toxicol. 12: 300, 1930.
GREENBERG, L. A., LESTER, D. AND hAGGARD, 11. W.: The reaction of hemoglobin withnitrite. J. Biol. Chem. 151: 665, 1943.
8 STIEGLITZ, E. J. AND PALMER, A. E.: A colorimetric method for the determination of
nitrite in blood. J. Pharmacol. & Exper. Therap. 51: 398, 1934.
� EVELYN, K. A. AND MALLOY, 11. T.: Microdetermination of oxyhennoglobin, methemoglo-
bin and sulfhemoglobin in a single sample of blood. J. Biol. Chem. 126: 655, 1938.
ha PONDER, E.: The mammalian red cell and the properties of haernolytic systems, Berlin,
Gebrssder Borntraeger, 1934.
b -: Acceleration of hemolysis in relation to chemical structure. I. Benzene dierivatives.
J. Exper. Biol. 16: 38, 1939.
“HUNTER, F. T.: A photoelectric method for the quantitative determination of erythrocyte
fragility. J. Curs. Investigation, 19: 691, 1940.
12 KING, E. J. AND DOLAN, M.: LIX. Enzynsic hydrolysis of phosphatides. II. Lysoleci-thins. Biochem. J. 27: 403, 1933.
‘� Win�nua, K. M. AND COLLIER, H. B.: A comparison of the hemolytic actions of lysolecithin
and saponin. J. Cell. & Comp. Physiol. 22: 233, 1943.
14 CLARK, B. B., MORRISSEY, R. W. AND BLAIR, D.: Some observations on the mechanism
of hemolytic anemias produced by drugs. Federation Proc. 4: 114, 1945.
‘� SINGER, K.: The lysolecithin fragility test. Am. J. M. Sc. 199: 466, 1940.
16 CLARK, B. B., VAN LOON, E. J. AND ADAMS, W. L.: Respiratory anti circulatory responsesto acute methemoglobinemia produced by aniline. Am. J. Physiol. 189: 64, 1943.
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom

1951 6: 532-543
BYRON B. CLARK, ROBERT W. MORRISSEY and DOROTHY BLAIR Relation of Methemoglobin to Hemolysis
http://www.bloodjournal.org/content/6/6/532.full.htmlUpdated information and services can be found at:
Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society of
For personal use only.on January 4, 2019. by guest www.bloodjournal.orgFrom