the promoter of dna puff gene 11/9-1 of sciara
TRANSCRIPT

CELL REGULATION, Vol. 2, 875-888, November 1991
The promoter of DNA puff gene 11/9-1 of Sciaracoprophila is inducible by ecdysone in late prepupalsalivary glands of Drosophila melanogaster
Brigitta Bienz-Tadmor, Heidi S. Smith,and Susan A. GerbiBrown UniversityDivision of Biology and MedicineProvidence, Rhode Island 02912
DNA puffs occur in Sciarid salivary gland chromo-somes; they are sites of DNA amplification and in-tense transcription and they appear to encode se-creted structural proteins needed for pupation. Inthis report we have used P-element transformationof Drosophila to study regulation of a Sciara DNApuff gene. We found that a 718-bp promoter frag-ment of DNA puff gene 11/9-1 from Sciara coprophiladirects expression of the bacterial reporter geneCAT in late prepupal salivary glands of transgenicDrosophila melanogaster. The identical tissue andanalogous stage specificity indicate that some as-pects of the ecdysone response are evolutionarilyconserved between Drosophila and Sciara. Whentransgenic salivary glands are cultured in vitro, CATactivity is rapidly induced by ecdysone, suggestingdirect control of gene expression by the ecdysonereceptor. Putative stage-specific factors limitexpression of the chimeric Sciara-CAT gene intransgenic Drosophila to late prepupae but not tothird instar larvae when ecdysone titers are alsohigh.
Introduction
In many dipteran insects the polytene chro-mosomes of late larval and prepupal salivaryglands undergo decondensation at a few dis-tinct loci resulting in cytologically visible swell-ings or puffs that are tissue and stage specific(Beermann, 1952). Recently it was found thatpuffing and RNA transcription are independentevents despite their association (Korge, 1987;Meyerowitz etal., 1987). Sciarid flies are uniquein that they also contain DNA puffs that are sitesnot only of RNA synthesis (Gabrusewycz-Garciaand Kleinfeld, 1966) but also DNA amplification(Rudkin and Corlette, 1957; Crouse and Keyl,1968; Rasch, 1970; Glover et al., 1982). Because
puffing is under strict developmental control,genes within puffs are favorable candidates tostudy tissue and stage-specific control of geneexpression, and in the case of DNA puffs, geneamplification as well.
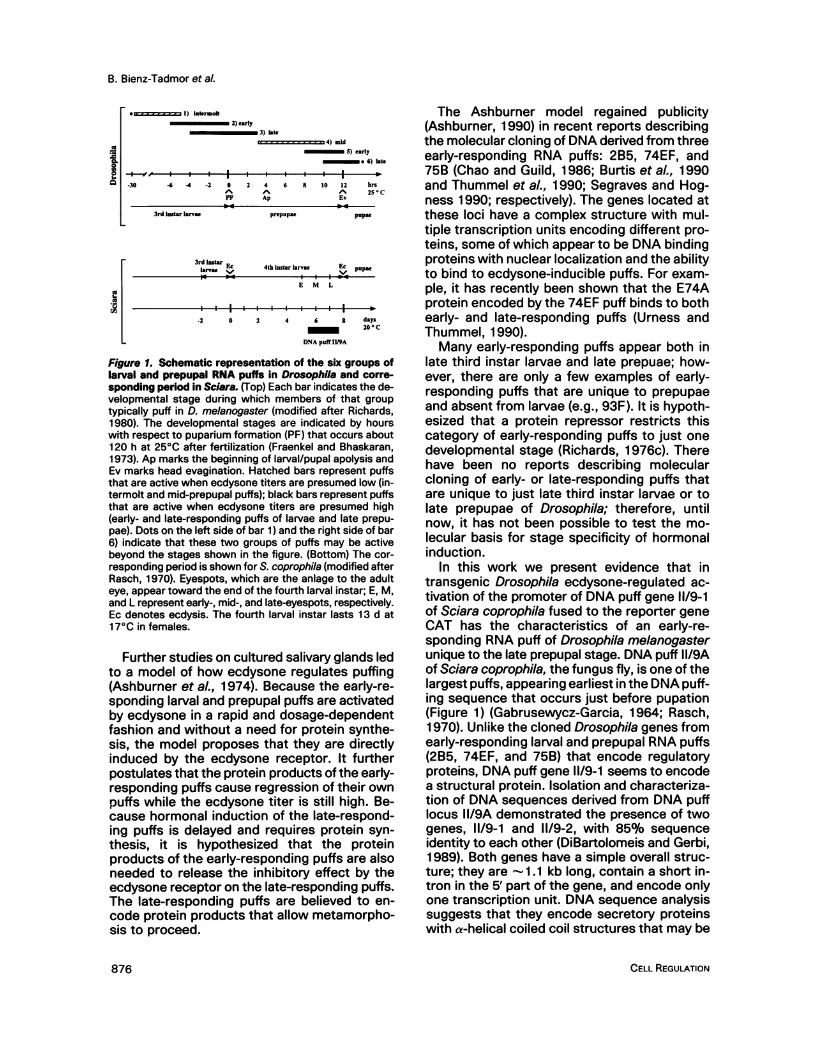
In Drosophila, the sequence of puffing ob-served in the salivary gland chromosomes oflate third instar larvae and prepupae is dividedinto 21 puff stages (Ashburner, 1967, 1969)comprising six main groups: the intermolt andthe early- and late-responding larval puffs andthe mid-prepupal and the early- and late-re-sponding late prepupal puffs (Figure 1). Thesmall group of intermolt puffs is active duringmost of the third larval instar and contains theglue protein genes; most of the intermolt puffsregress when the ecdysone titer starts to risenear the end of the third larval instar. At thattime, about half a dozen early-responding larvalpuffs appear and are followed by 100 late-responding larval puffs. A second cycle of puff-ing, comparable with the larval cycle, begins afew hours after puparium formation. This pre-pupal puffing sequence starts with the smallgroup of mid-prepupal puffs that, like the larvalintermolt puffs, are active while ecdysone levelsare low in the animal. A presumed rise in theecdysone titer toward the end of the prepupalstage initiates puffing in late prepupae, first ofthe early-responding puffs followed by the late-responding puffs.The entire sequence of larval puffing can be
induced in vitro when Drosophila intermolt sal-ivary glands are cultured in the presence of ec-dysone (Ashburner, 1972-1974; Ashburner etal., 1974; Ashburner and Richards, 1976). How-ever, transition from the larval to the prepupalpuff sequence does not occur in vitro unlesssalivary glands are placed in an ecdysone-freeculture medium and allowed to carry out proteinsynthesis. During this period, the mid-prepupalpuffs expand, and competence to respond toecdysone is acquired. A subsequent exposureof competent salivary glands to ecdysone in-duces puffing of the early- and late-respondingpuffs of late prepupae (Richards, 1 976a-c).
© 1991 by The American Society for Cell Biology 875
B. Bienz-Tadmor et al.
go
I
to
0 = 1 ) lntermot2) early
3) late- - - - - - 4) mld
S) early-. 6) late
.30 -6 -4 -2 0 2 4 6 8 10 12 hrs^ 25°C
PF Ap Ev
3rd latar larvae prepupae pupae
nst EC 4th instar larvae Ec pupae
E M L
-2 0 2 4 6 8 days
DNA puffI/9A
Figure 1. Schematic representation of the six groups oflarval and prepupal RNA puffs in Drosophila and corre-sponding period in Sciara. (Top) Each bar indicates the de-velopmental stage during which members of that grouptypically puff in D. melanogaster (modified after Richards,1980). The developmental stages are indicated by hourswith respect to puparium formation (PF) that occurs about120 h at 250C after fertilization (Fraenkel and Bhaskaran,1973). Ap marks the beginning of larval/pupal apolysis andEv marks head evagination. Hatched bars represent puffsthat are active when ecdysone titers are presumed low (in-termolt and mid-prepupal puffs); black bars represent puffsthat are active when ecdysone titers are presumed high(early- and late-responding puffs of larvae and late prepu-pae). Dots on the left side of bar 1) and the right side of bar6) indicate that these two groups of puffs may be activebeyond the stages shown in the figure. (Bottom) The cor-responding period is shown for S. coprophila (modified afterRasch, 1970). Eyespots, which are the anlage to the adulteye, appear toward the end of the fourth larval instar; E, M,and L represent early-, mid-, and late-eyespots, respectively.Ec denotes ecdysis. The fourth larval instar lasts 13 d at170C in females.
Further studies on cultured salivary glands ledto a model of how ecdysone regulates puffing(Ashburner et al., 1974). Because the early-re-sponding larval and prepupal puffs are activatedby ecdysone in a rapid and dosage-dependentfashion and without a need for protein synthe-sis, the model proposes that they are directlyinduced by the ecdysone receptor. It furtherpostulates that the protein products of the early-responding puffs cause regression of their ownpuffs while the ecdysone titer is still high. Be-cause hormonal induction of the late-respond-ing puffs is delayed and requires protein syn-thesis, it is hypothesized that the proteinproducts of the early-responding puffs are alsoneeded to release the inhibitory effect by theecdysone receptor on the late-responding puffs.The late-responding puffs are believed to en-code protein products that allow metamorpho-sis to proceed.
The Ashburner model regained publicity(Ashburner, 1990) in recent reports describingthe molecular cloning of DNA derived from threeearly-responding RNA puffs: 2B5, 74EF, and75B (Chao and Guild, 1986; Burtis et al., 1990and Thummel et al., 1990; Segraves and Hog-ness 1990; respectively). The genes located atthese loci have a complex structure with mul-tiple transcription units encoding different pro-teins, some of which appear to be DNA bindingproteins with nuclear localization and the abilityto bind to ecdysone-inducible puffs. For exam-ple, it has recently been shown that the E74Aprotein encoded by the 74EF puff binds to bothearly- and late-responding puffs (Urness andThummel, 1990).Many early-responding puffs appear both in
late third instar larvae and late prepuae; how-ever, there are only a few examples of early-responding puffs that are unique to prepupaeand absent from larvae (e.g., 93F). It is hypoth-esized that a protein repressor restricts thiscategory of early-responding puffs to just onedevelopmental stage (Richards, 1976c). Therehave been no reports describing molecularcloning of early- or late-responding puffs thatare unique to just late third instar larvae or tolate prepupae of Drosophila; therefore, untilnow, it has not been possible to test the mo-lecular basis for stage specificity of hormonalinduction.
In this work we present evidence that intransgenic Drosophila ecdysone-regulated ac-tivation of the promoter of DNA puff gene 11/9-1of Sciara coprophila fused to the reporter geneCAT has the characteristics of an early-re-sponding RNA puff of Drosophila melanogasterunique to the late prepupal stage. DNA puff 11/9Aof Sciara coprophila, the fungus fly, is one of thelargest puffs, appearing earliest in the DNA puff-ing sequence that occurs just before pupation(Figure 1) (Gabrusewycz-Garcia, 1964; Rasch,1970). Unlike the cloned Drosophila genes fromearly-responding larval and prepupal RNA puffs(2B5, 74EF, and 75B) that encode regulatoryproteins, DNA puff gene 11/9-1 seems to encodea structural protein. Isolation and characteriza-tion of DNA sequences derived from DNA pufflocus 11/9A demonstrated the presence of twogenes, 11/9-1 and 11/9-2, with 850/° sequenceidentity to each other (DiBartolomeis and Gerbi,1989). Both genes have a simple overall struc-ture; they are -1.1 kb long, contain a short in-tron in the 5' part of the gene, and encode onlyone transcription unit. DNA sequence analysissuggests that they encode secretory proteinswith a-helical coiled coil structures that may be
CELL REGULATION
-
876

Sciara DNA puff promoter in Drosophila
-616 +102
P rosy Scioro SU40-CRT P
Figure 2. Schematic representation of the Sciara-CAT P-element construct. The Sciara promoter fragment (-616to +102 respective to the cap site indicated by an open box)was linked to the SV40-CAT gene (the CAT gene, markedby a black box, is flanked by 5' and 3 untranslated sequencesof SV40 marked by a solid line, and includes an intron). TheP-element ends (P) are shown as vertically hatched boxes,and the selectable marker gene rosy is depicted by a di-agonally hatched box. Directions of transcription of the rosygene and the hybrid SV40-CAT gene are indicated by arrows.This schematic map is not drawn to scale.
needed for pupation (DiBartolomeis and Gerbi,1989). During the late fourth larval instar, thetwo genes undergo an -20-fold amplificationbefore a burst in transcription in the salivaryglands just before pupation resulting in massiveamounts of puff-specific mRNA (DiBartolomeis,Wu, Liang, Smith, and Gerbi, unpublished data).We report here that the promoter of Sciara
DNA puff gene 11/9-1 drives expression of thereporter gene CAT exclusively in late prepupalsalivary glands of Drosophila P-element trans-formants. In vitro studies on isolated salivaryglands from transgenic Drosophila show a rapidinduction of CAT activity by ecdysone, sug-gesting direct control of gene expression by theecdysone receptor. In addition, stage-specifictrans-acting factors are proposed to restricthormonal induction to the late prepupal stagein transgenic Drosophila. These stage-specifictrans-acting factors as well as the ecdysone re-ceptor of Drosophila must interact specificallywith the 718-bp promoter fragment of SciaraDNA puff gene 11/9-1, resulting in appropriatetissue and stage-specific expression in trans-genic prepupae.
ResultsThe Sciara DNA puffpromoter shows stagespecificity in DrosophilaA 718-bp long promoter fragment of DNA puffgene 11/9-1 from Sciara coprophila (-616 to+102, relative to the cap site) was linked to thebacterial reporter gene CAT and inserted intoa P-element transformation vector (Figure 2).The recombinant plasmid was stably introducedinto germ line chromosomes of Drosophila mel-anogaster by P-element transformation, andseveral homozygous transformed lines wereestablished. All lines carried a single copy in-sertion as shown by Southern blot analysis of
genomic DNA (Figure 3, shown only for the A-line and C-line) and by in situ hybridization forone line (Figure 3, C-line).To determine the time period during which
the chimeric Sciara-CAT gene is expressed inDrosophila, CAT assays were carried out onwhole transgenic animals at various develop-mental stages. Figure 4 shows that in two dif-ferent lines (A-line and C-line) high levels of CATactivity first appear in 9-h prepupae (PP9) andcontinue at the same high levels throughout thelate prepupal and early pupal stages (P15) untilthey decrease and disappear in 36- to 48-h pu-pae (P36 and P48). Data mentioned later sug-gest that the continued high levels of CAT ac-tivity after its induction at PP9 reflect CATstability rather than de novo synthesis through-out this period. No CAT activity can be detectedduring the early larval stages (L1,2), the late pu-
- C. :C. C C.
0) 0) 0
_. _J
23.1L _9.4- 3__6.6_
4.4-
2.3 -
2.0 -
9*-."*'- ,.24
,., .6'*6;,
* . h
.,. 'i's *.I,/4 *'.f.w -. -
. A .
- ek1 21).1 '*
4>'
I
Figure 3. Single insertion site of the Sciara-CAT P-elementconstruct in transgenic Drosophila. Southern blot analysis(left panel) was carried out by hybridizing a labeled P-elementprobe (derived from the downstream P-element end, Figure2) to Xho I digested genomic DNA. The DNA was preparedfrom 9-h prepupae (PP9) or adult flies (Adult) transformedwith the Sciara-CAT P-element construct. The single hy-bridizing Xho I fragment is -8 kb long for the C-line and 10kb for the A-line, reflecting the location of the next Xho Isite in the host's DNA beyond the insertion. The fragmentsizes of HindlIl digested X DNA are shown on the left. Thestronger signal for transgenic adult flies (A-line) is due toloading more genomic DNA rather than amplification ofSciara specific DNA in adult Drosophila. The right panelshows a representative in situ hybridization of polytenechromosomes of transgenic Drosophila (C-line) hybridizedto a biotin-labeled 5.5-kb probe containing Sciara gene Iand upstream sequences (see map in Figure 6). The site ofinsertion at locus 56-AB on chromosome ll/R is indicatedby an arrow.
Vol. 2, November 1991 877

B. Bienz-Tadmor et al.
*A r t
1_2 I 3l 1'PO P1'6PIP Pj5 P3 }P4X \
^I e
i c -
a-
1I1 II'flI'1P('P6 IN P1I P36 P491 P-2
Figure 4. Temporal specificity of gene expression intransgenic Drosophila. CAT assays are shown for two linesof transgenic Drosophila (A-line and C-line). CAT activity wasdetermined in extracts prepared at various developmentalstages each from 10 total animals: first and second instarlarvae (Li, 2), late third instar larvae (L3, 115 h old), pre-pupae (PP), pupae (P), and adults (A). The numbers for theprepupal and pupal stages represent hours after pupariumformation (which occurs at PPO) at 25 ± 1 'C. Positions ofunacetylated chloramphenicol (CAM) and its acetylatedforms (ac-1 and ac-3) as well as the origin (ori) on the chro-matogram are indicated on the right.
The Sciara DNA puffpromoter shows tissuespecificity in DrosophilaIn Sciara coprophila, mRNA expression of DNApuff gene 11/9-1 is seen only in salivary glandsand not in the remaining carcasses (Di-Bartolomeis, Wu, Liang, Smith, and Gerbi, un-published data). To test whether the chimericSciara-CAT gene is expressed with the sametissue specificity in transgenic Drosophila as inSciara, CAT activity was determined in tissuesdissected from 9-h prepupae, the stage whenhigh levels of CAT gene expression first appear.Figure 5 shows that CAT activity is located inthe salivary glands of 9-h prepupae in bothtransformed lines (A-line and C-line). CAT ac-tivity is not detectable in other tissues: cuticlestructures and muscles, imaginal discs andbrain, and fat body; very low levels of CAT ac-tivity were found in the Malpighian tubules andgut. CAT activity is also absent in the salivaryglands of 9-h prepupae transformed with a pro-moterless CAT construct or a CAT constructdriven by the promoter of the late chorion genesi5 of Drosophila (B. Bienz-Tadmor, Wu, Liang,Smith, and Gerbi, unpublished data). This indi-cates that CAT activity seen in transformed sal-ivary glands is due to the presence of the SciaraDNA puff sequences rather than vector or hostsequences. In 24-h pupae, the stage when his-tolysis of the salivary glands is completed (Bod-
pal (P72), or adult stages (A). In the A-line, verylow levels of CAT activity are seen during thelate third larval instar and the early- and mid-prepupal stages (L3, 115 h old; PPO and PP6).In the C-line, this low level expression is onlyvisible after longer exposure times.The sharp increase in CAT activity in late pre-
pupae of transgenic Drosophila coincides withthe proposed increase in ecdysone levels duringthe prepupal period that induces the early-re-sponding RNA puffs (Richards, 1981). This sug-gests a positive and stage-specific hormonalregulation of hybrid gene expression in Dro-sophila. As will be discussed later, the prepupalexpression of the chimeric gene in Drosophilaappears to be analogous with the temporalspecificity in Sciara where there is a rapid in-crease in mRNA from DNA puff gene 11/9-1shortly before pupation (Figure 1) (DiBartolo-meis, Wu, Liang, Smith, and Gerbi, unpublisheddata).
*000*000000~~~~~~~~('0l
*-----------(.'\I N1'T(l IDII Hi .(" hit ( a SG. Iu >11 >1
-4 _- _ _ _ _ _--0 .* + +
\ -lirit ( l inte 1lP ,0
Figure 5. Tissue specificity of gene expression in trans-genic Drosophila. CAT assays were carried out on extractsprepared from various dissected tissues from 10 transgenicprepupae that were 9 h old. For the A-line, CAT activity wasdetermined in tissues such as the cuticle structures andmuscles (CM), Malpighian tubules and gut (MTG), imaginaldiscs and brain (IDB), fat body (FB), and salivary glands (SG)and in total animals (Tot). For the C-line, CAT activity wasdetermined in carcasses (Ca, prepupae from which the sal-ivary glands were removed), in the salivary glands (SG), andin total animals (Tot). CAT activity was also determined insalivary glands of prepupae transformed with a promoterlessCAT construct (SGo) or with a CAT construct carrying apromoter of the chorion gene s15 of Drosophila (SGp).
CELL REGULATION878

Sciara DNA puff promoter in Drosophila
enstein, 1965), CAT activity is associated withundefined cell masses and body fluids and couldnot be attributed to any of the tissues still dis-cernable at this stage of general metamorphosis(data not shown). Hence, CAT activity seen in24-h and older pupae is unlikely to be causedby de novo expression of the CAT gene in anovel tissue; rather, CAT activity in intact pupaecan be explained by a high stability of the CATprotein before and during the final stages ofhistolysis of the salivary glands, as will be dis-cussed below.
Therefore, it appears that hybrid geneexpression in transgenic Drosophila is primarilyrestricted to the salivary glands, the same andonly tissue in which DNA puff gene 11/9-1 is ex-pressed in Sciara. It should be pointed out thatthe Sciara DNA puff promoter retains the sametissue and analogous stage specificity in trans-genic Drosophila as in Sciara, even thoughSouthern blot analysis (Figure 6) and in situ hy-bridization (Figures 3 and 10) show no heter-ologous hybridization between Sciara DNA puffgene 11/9-1 and the Drosophila genome. None-theless, hybrid gene expression driven by theSciara DNA puff promoter in transgenic Dro-sophila coincides with the appearance of theearly-responding RNA puffs of Drosophila duringthe late prepupal period (Richards, 1 976c).The stage and tissue specificity of hybrid gene
expression illustrated for two lines of transgenicDrosophila is identical in all four lines that weanalyzed (data shown only for the A-line and C-line in Figures 4 and 5), but the quantitative lev-els of CAT activity varied from line to line. Allfour lines have independent positions of auto-somal integration. This suggests that the ob-served pattern of expression is due to SciaraDNA rather than flanking host sequences, al-though it appears that there is a position effecton the level of expression of the hybrid genedependent on its site of chromosomal integra-tion. A position effect on quantitative levels ofgene expression has been reported for manyother gene constructs in transgenic Drosophila(e.g., Mitsialis et al., 1987).To extend our analysis from the protein to the
RNA level, Northern blot analysis and primerextension experiments were carried out. How-ever, neither of the two techniques detectedCAT-specific mRNA at any developmental stage(preliminary data not shown) including late pre-pupae (PP6-9), a time when high levels of CATactivity are present. The inability to detect CAT-specific mRNA in late prepupae may in part bedue to a much higher sensitivity of the enzy-matic assay for CAT as compared with the sen-
sitivity of the RNA detection experiments. Ad-ditionally, it may also indicate that CAT mRNAis expressed indeed only very briefly and henceis present only in a fraction of the 6- to 9-h pre-pupae from which the RNA was prepared.
DNA from Sciara DNA puffgene 11/9-1 is notamplified in transgenic DrosophilaIt is unknown where the amplification controlelements reside in Sciara DNA puff 11/9A. How-ever, it seemed likely that the observed CATactivity in transgenic Drosophila was due to hy-brid gene expression from nonamplified DNA.We have used quantitative Southern blots toshow that in fact even the 8.8-kb region of SciaraDNA from which the 71 8-bp promoter fragmentwas derived does not become amplified intransgenic Drosophila. This 8.8-kb region con-tains DNA puff genes 11/9-1 and 11/9-2 and flank-ing sequences. As can be seen in Figure 6, thereis no amplification of this DNA in salivary glandsfrom 0- to 1-h prepupae (PP0-1) or from 8- to9-h prepupae (PP8-9) as compared with car-casses of the corresponding stages or adultflies.
The Sciara DNA puffpromoter shows astage-specific response to ecdysone in vitroTo validate the notion that DNA puff gene 11/9-1of Sciara is active in the same manner as thelate prepupal RNA puffs of Drosophila, its in vitroresponse to ecdysone was examined in culturedsalivary glands of transgenic Drosophila. Asmentioned earlier, it has been demonstratedthat puffing of the late prepupal RNA puffs ofDrosophila can be induced in competent pre-pupal salivary glands that are cultured in thepresence of 20-OH ecdysone (Richards, 1 976c),shown to be the most active form of ecdysone(Ashburner, 1971). Competence to respond toecdysone is reached in vivo in the salivaryglands of 6-h prepupae, a stage when ecdysonelevels are deduced to be extremely low (Rich-ards, 1976a). Competence to respond to ec-dysone can also be reached in vitro if early pre-pupal salivary glands are incubated in anecdysone-free medium before hormonal induc-tion (Richards, 1976b,c).To test the Sciara promoter for a stage-spe-
cific response to ecdysone in vitro, salivaryglands of transgenic Drosophila (A-line) wereisolated from several developmental stages andincubated for various periods in the presence(+E) or absence (-E) of 20-OH ecdysone. CATactivity was determined in these glands afterthe in vitro incubation was completed and com-
Vol. 2, November 1991 879

B. Bienz-Tadmor et al.
tIi! 1. ., II il1i
t .- r!L
1F(I S t
1"__S4_*i!
11 --W -_
._----
H IO O.FH21
Figure 6. DNA from Sciara DNA puff 11/9A does not amplifyin transgenic Drosophila. A map of the 8.8-kb Sciara DNA(1.6-kb and 7.2-kb fragments) integrated into the Drosophilagenome is shown at the top. The 8.8-kb Sciara DNA containsthe DNA puff genes 11/9-1 and 11/9-2 ( ) and flanking se-quences (LI). The labeled DNA probes are shown below themap (the dotted line represents Sciara genomic sequencespresent in the 5.5-kb probe but not included in the transgenicinsertion). Southern blot analysis was carried out with Xhodigested genomic DNA from three transgenic Drosophila
lines (Al184, A200, S191) and nontransformed flies (cn,ry).The DNA was prepared from 0- to 1-h prepupal salivaryglands and carcasses (PPO-1 SG and Ca) or 8- to 9-h pre-pupal salivary glands and carcasses (PP8-9 SG and Ca), aswell as from adults of the same lines. Genomic DNA fromlines Al 84 and A200 was hybridized to probes derived fromgene 1 and gene 2. The hybridizing Xho fragments are 2.4,2.7, and 7.7 kb long, as indicated in the left-most lane bythe hybridization pattern of the Xho digested Sciara-P ele-ment construct and on the schematic map above (the Xhosite in parentheses was destroyed during cloning). For line191 the 5.5-kb gene 1 fragment was used as a probe re-
sulting in only one hybridizing 7.7-kb Xho fragment (indi-cated by a dot). (in addition to this single band, two bandsof X DNA used as a carrier are also visible in the salivarygland sample.) A labeled probe derived from the DrosophilaAlcohol dehydrogenase gene (Adh) was used to normalizethe amount of loaded DNA; it detected an 8-kb Xho frag-ment.
pared with the enzymatic activity of salivaryglands taken directly from animals at the cor-responding developmental stage. As shown inFigure 7, CAT activity can be induced by incu-bating competent salivary glands in a mediumcontaining 5 x 10-6 M 20-OH ecdysone. Com-petence to respond to ecdysone was reachedeither in vivo or in vitro, but the induced levels
of CAT activity in 6-h prepupal salivary glandsthat reached competence in vivo (PP6, +E2 and+E4) are higher than in 3-h prepupal salivaryglands that reached competence in vitro duringa 3-h preincubation in ecdysone-free mediumbefore induction (PP3, 3/+E2). Note that aboutthe same levels of induced CAT activity are ob-served if competent salivary glands are incu-bated for 2 (PP6, +E2) or 4 h (PP6, +E4) in thepresence of ecdysone. This indicates thatsteady-state levels of CAT activity have beenreached essentially within the first 2 h of an invitro exposure to ecdysone, as will be shownconclusively below.
In the absence of ecdysone, induction of CATactivity does not take place; the low levels ofCAT activity found in salivary glands culturedwithout ecdysone (PP6, -E2 and PP3, 3/-E2)are not higher than the levels seen in salivaryglands directly isolated from prepupae of thecorresponding stages (PP6, salivary gland andPP3, salivary gland) that are before in vivo in-duction by ecdysone. In addition, CAT activitycannot be induced by ecdysone in salivaryglands of -1 10-h third instar larvae (L3, +E4)or in incompetent salivary glands of 3-h pre-pupae (PP3, +E2). Taken together, the resultsfrom these in vitro studies indicate that the chi-meric Sciara-CAT gene can be induced by 20-OH ecdysone in competent prepupal Drosophilasalivary glands. This indicates a stage-specifichormonal induction restricted to the late pre-pupal period. Note that steady-state levels of
4- -1 12 - F: F.. -
Figure 7. Stage specificity of hormonal induction in vitro.CAT assays were performed on extracts prepared from fivepairs of transgenic Drosophila salivary glands (A-line) cul-tured in vitro in the presence (+E) or absence (-E) of ec-dysone for 2 (+E2 or -E2) or 4 h (+E4 or -E4) and fromsalivary glands that were not cultured (SG). A 3-h preincu-bation in ecdysone-free medium before transfer in ecdysoneor control medium is indicated by 3/+E2 or 3/-E2, respec-tively. The developmental stages at which the salivary glandswere isolated are as follows: late third instar larvae (L3,- 1 10 h old), 3-h prepupae (PP3), 6-h prepupae (PP6), and9-h prepupae (PP9).
CELL REGULATION880

Sciara DNA puff promoter in Drosophila
protein activity (CAT) were measured ratherthan mRNA initiation because nuclear run-onexperiments are not feasible in this system.However, hormonal induction seems to takeplace at the transcriptional level because in vitroinduction of CAT activity in 6-h prepupal salivaryglands does not take place in the presence ofactinomycin D (data not shown).
The Sciara DNA puffpromoter shows a quickresponse to ecdysone in vitroAs mentioned earlier, in Drosophila the larvaland late prepupal RNA puffs are divided intoearly- and late-responding puffs. One differencebetween these groups of puffs is seen in therapidity by which they respond to ecdysone invitro. In late third instar larvae early-respondingpuffs respond to ecdysone within minutes witha maximal response between 1 and 4 h, whereaslate-responding puffs show a delay in their re-sponse to ecdysone for .4 h (Ashburner, 1972).The puffing sequence in late prepupae in vivois much shorter than in larvae, and although theearly-responding prepupal puffs are inducedquicker than the late-responding RNA puffs, thedifferences are less pronounced in late prepu-pae than in larvae (Richards, 1976c).To determine the rapidity by which the Sciara
DNA sequences respond to ecdysone in vitro,competent salivary glands of 6-h prepupae werecultured in the presence of ecdysone for variousperiods of time. As shown in Figure 8, a stronginduction of CAT activity was detected in sali-vary glands that were cultured in ecdysone-containing medium for 1 h or longer. Theamount of induced CAT activity is almost as highas that found in vivo for salivary glands of 9-hprepupae (PP9, salivary gland) and is essentiallythe same regardless of whether the salivaryglands were incubated for 1 (PP6, +E1), 2 (PP6,+E2), or 6 h (PP6, +E6). However, shorter in-cubation periods of half an hour or less do notresult in a significant induction of CAT activity;only low levels of CAT activity are found in sal-ivary glands incubated for a few minutes (PP6,+EO; incubation shorter than 1 0 min) or half anhour (PP6, +E1 /2). Similar low levels of CAT ac-tivity are also present in 6-h prepupal salivaryglands that were either cultured for 6 h in theabsence of ecdysone (PP6, -E6) or that werenot cultured at all (PP6, salivary gland).These results indicate that steady-state levels
of induced CAT activity are reached betweenhalf an hour and 1 h of exposure to ecdysonein vitro, suggesting a rapid hormonal inductionof the chimeric Sciara-CAT gene. Note that
ac-3
ac- I
* * * * * **@ (C.A I
Ori
1'S6
Si; SGP_4
"P9
Figure 8. rime-course of hormonal induction in vitro. CATactivity was determined in five pairs of transgenic Drosophilasalivary glands isolated from 6-h prepupae (A-line). The sal-ivary glands were cultured in the presence of ecdysone forvarious periods of time (PP6, +EO; +E1/2; +E1; +E6). Theincubation time is given in hours with an accuracy of ±10min (except the incubation time for +EO was 0-10 min).CAT activity was also determined in 6-h (PP6, SG) and 9-h(PP9, SG) prepupal salivary glands that were not culturedand in 6-h prepupal salivary glands that were cultured for 6h in the absence of ecdysone (PP6, -E6).
protein levels, rather than mRNA levels or puffsizes, were determined in this experiment, sothe presumed transcriptional response musthave been even more rapid. Hence, the SciaraDNA sequences show a response to ecdysonein transgenic Drosophila like that expected foran early-responding gene of a Drosophila RNApuff.
Gene expression driven by the Sciara DNApuffpromoter is terminated in vitro while stillin the presence of ecdysoneIn vitro studies on isolated Drosophila salivaryglands showed that although ecdysone inducespuffing of the early-responding larval or prepupalRNA puffs, these puffs typically regress rapidlywhile ecdysone is still present (Ashburner, 1972;Ashburner and Richards, 1976; Richards,1976c). To test the behavior of the chimericSciara-CAT gene after high levels of CAT activityhave been reached in vivo, salivary glands of 9-h prepupae were cultured for 12 h with andwithout ecdysone. As shown in Figure 9, levelsof CAT activity are approximately the same dur-ing a 12-h incubation in the presence or absenceof ecdysone (PP9, +E12 and PP9, -E12, re-spectively). Therefore, once CAT activity hasbeen induced in vivo in 9-h prepupae, this ac-tivity is maintained for an additional 12 h evenin the absence of ecdysone. This steady-statelevel of CAT activity for 12 h could reflect either(1) continued synthesis of CAT mRNA and/or
Vol. 2, November 1991
E 6 EO -F. -I'll -E2 -E6
881

B. Bienz-Tadmor et al.
protein coupled with an equal rate of turnoveror (2) no further synthesis of CAT mRNA and/or protein after the initial burst but a stable CATproduct. To distinguish between these possi-bilities we did a 12-h incubation of 9-h prepupalsalivary glands in the presence of actinomycinD (PP9, +Am1 2) or cycloheximide (PP9, +Ch1 2).As seen in Figure 9, neither drug significantlychanges CAT activity levels from that alreadypresent in salivary glands of 9-h prepupae (PP9,salivary gland). This indicates that maintenanceof CAT activity for an additional 12 h does notrequire de novo synthesis of either CAT mRNAor CAT protein, suggesting that the CAT proteinhas virtually no turnover in Drosophila salivaryglands for .12 h.When these findings are applied to the in vivo
situation, they suggest that most expression ofthe CAT gene occurs only for a short period inlate prepupae that are probably between 6 and9 h old and that CAT activity seen thereafterprimarily is due to high stability of the CAT pro-tein rather than de novo expression of the CATgene. Similarly, the early-responding RNA puffsof Drosophila larvae and late prepupae are onlytransiently induced by ecdysone, and they re-gress rapidly (Ashburner, 1972; Richards,1 976c).
DNA from Sciara DNA puff 11/9A does notpuff in transgenic Drosophila salivary glandchromosomesBecause the chimeric Sciara-CAT gene is ex-pressed in late prepupal salivary glands oftransgenic Drosophila, we wondered if expres-sion of the inserted DNA is also associated withpolytene chromosome puffing. It is not knownwhere the sequences responsible for puffingoccur in the Sciara 11/9A locus, so rather thanjust testing the limited 71 8-bp promoter region,we tested the larger 8.8-kb genomic regioncontaining it (see map in Figure 6). In situ hy-bridization of salivary gland chromosomes fromDrosophila transformed with the 8.8-kb SciaraDNA (line Al 84) showed a single insertion siteat locus 1 C on the X chromosome (Figure 10,top). When this locus was examined in polytenechromosome squashes from 6- to 9-h (PP6-PP9) prepupal salivary glands, no puff was foundat the site of the inserted Sciara DNA (Figure10, middle). However, the early-responding RNApuffs of Drosophila late prepupae were clearlypresent (e.g., RNA puff 2B5). Because the ex-pected time interval of transcriptional activityand hence potential puffing activity is only for
1 h during this 3-h prepupal period (see pre-
E12 -H 2 S --km 12l h I'
Figure 9. Stability of CAT proteins in transgenic salivaryglands in vitro. CAT activity was determined in five pairs oftransgenic Drosophila salivary glands isolated from 9-h pre-pupae (PP9, A-line) that were not cultured (SG) or were cul-tured for 12 h in the presence (+El 2) or absence of ecdysone(-El 2) or cultured for 12 h in the presence of actinomycinD (+Aml 2) or cycloheximide (+Chl 2). In this particular ex-periment, CAT activity of salivary glands that were not cul-tured (SG) was slightly lower than usual. As a control, bothdrugs were shown to be active because they inhibited theinduction of CAT activity when added to the medium con-taining ecdysone for culture of 6-h prepupal salivary glands(data not shown).
ceding section), we also examined squashes ofsalivary gland chromosomes from competentprepupae after a 30- to 60-min in vitro incuba-tion with ecdysone. Even in this case, no puffwas apparent at the site of insertion (Figure 10,bottom), although other early-responding pre-pupal RNA puffs of Drosophila were induced invitro by ecdysone (e.g., RNA puff 2B5).
Discussion
Expression of the Sciara DNA puffgenesoccurs in the absence ofDNA amplificationor puffingIn this work we present evidence that a 71 8-bplong promoter fragment of DNA puff gene 11/9-1of Sciara coprophila directs expression of thereporter gene CAT in transgenic Drosophila withcorrect tissue and analogous stage specificity.However, expression occurs in the absence ofDNA amplification or puffing of an 8.8-kb frag-ment of Sciara DNA that contains the 71 8-bppromoter fragment. This indicates that neitheramplification nor puffing is required for expres-sion of the chimeric Sciara-CAT gene. Similarobservations have been previously reported;expression of the Drosophila glue proteins doesnot require RNA puffing at the site of gene in-
CELL REGULATION882

Sciara DNA puff promoter in Drosophila
tegration (Korge, 1987; Meyerowitz et al., 1987),and correct temporal and tissue-specificexpression of the Drosophila chorion genes doesnot require major amplification of the chorionlocus (Orr et al., 1984; Parks et al., 1986; Wak-imoto et al., 1986). Future studies will be neededto define the positions of control elementsfor amplification and puffing in Sciara DNApuff 11/9A. However, it is possible that thesecontrol elements are present in the tested 8.8-kb Sciara DNA but are not recognized or thephysiological conditions (e.g., ecdysone levels)are not appropriate in transgenic Drosophila.
The Sciara DNA puffpromoter retains tissueand stage specificity in DrosophilaNorthern blots show that in Sciara coprophila,DNA puff gene 11/9-1 is expressed only in sali-vary glands of fourth instar larvae (Di-Bartolomeis, Wu, Liang, Smith, and Gerbi, un-published data). The experiments we reporthere demonstrate that this tissue specificity isretained in transgenic Drosophila in which the71 8-bp-long promoter fragment of Sciara DNApuff gene 11/9-1 directs expression of the re-porter gene CAT exclusively in the late prepupalsalivary glands (Figure 5). Sequences within the718 bp of Sciara DNA responsible for this tissuespecificity have not yet been identified.The retention of stage specificity for expres-
sion of the chimeric gene in transgenic animals(Figure 4) is harder to ascertain because of de-velopmental differences between Sciara andDrosophila, although both are holometabolousorganisms with a complete metamorphosis.Sciara is a lower dipteran fly (suborder Nema-tocera) with four larval instars preceding pu-pation (Figure 1); each stage is demarcated byecdysis, that is shedding of the cuticle, presum-ably triggered by an increase in ecdysone titer,though this has not been measured directly.Drosophila, instead, is a higher dipteran fly(suborder Cyclorrapha) that has evolved a pu-parium (Fraenkel and Bhaskaran, 1973). At theend of the third larval instar, the larval cuticleis not shed in response to the increased ecdy-sone titer (unlike the third/fourth larval instarecdysis of Sciara) but instead forms the barrel-like puparium (PF in Figure 1). Four hours afterpuparium formation, larval/pupal apolysis be-gins (Ap in Figure 1) in which the puparium sep-arates from the underlying epidermis. Twelvehours after puparium formation, the head evag-inates (Ev in Figure 1); this event was suggestedto be analogous to larval/pupal ecdysis in lowerdiptera and marks the onset of pupation (Fraen-
kel and Bhaskaran, 1973). There has been de-bate whether the term "molt" should refer toapolysis or ecdysis and which of these two pro-cesses should be used to define an instar (Hin-ton, 1976; Whitten, 1976). There also has beenambiguity in how to use the term "prepupa." Inthis study we use "prepupa" in the same wayused by Ashburner and Richards to refer to thestage of Drosophila extending from pupariumformation to head evagination (PP0-PP1 2;"prepupa" also used in this sense by Fraenkeland Bhaskaran, 1973; Bainbridge and Bownes,1981, 1988). Our results therefore suggest thatDNA puff gene 11/9-1 sequences are active inboth fly species (Sciara fourth larval instar orDrosophila prepupa) at the developmental stagepreceding pupation.The retention of tissue and analogous stage
specificity of the Sciara DNA puff promoter intransgenic Drosophila is the first indication thatsome aspects of the mechanism by which ec-dysone controls gene expression are evolution-arily conserved. The Sciaridae family belongs tothe suborder Nematocera for which the fossilrecord dates back to the Triassic, 220 million yago; the Drosophilidae family belongs to thesuborder Brachycera for which the fossil recorddates back to the Jurassic, 180 million y ago(Carpenter, 1991). Therefore, the taxonomicsplit between Sciara and Drosophila was quiteancient, having occurred > 180 million y ago. Theevolutionary conservation of the ecdysone re-sponse with respect to tissue and stage spec-ificity between these dipteran flies is consistentwith the conservation seen for several receptorsof other steroid hormones (for reviews see Ya-mamoto, 1985; Evans, 1988; Green and Cham-bon, 1988; Beato, 1989).
Gene expression driven by the Sciara DNApuffpromoter is inducible by ecdysoneA rise in salivary gland ecdysone titer is corre-lated with DNA puffing (Stocker et al., 1984),and hormone injection and ligation experimentshave shown that ecdysone controls DNA puffformation in Sciarids (Crouse, 1968; Stocker andPavan, 1974; Amabis and Amabis, 1 984a,b). Wehave now extended these cytological observa-tions to the molecular level. Our in vitro exper-iments with cultured Drosophila salivary glandsindicate that the chimeric Sciara-CAT gene isinducible by 20-OH ecdysone as it shows thata quick response to ecdysone and steady-statelevels of CAT activity are reached between halfan hour and 1 h of exposure to ecdysone. Therapid response of the hybrid gene parallels the
Vol. 2, November 1991 883

B. Bienz-Tadmor et al.
4004
~~~~~N~~~~~~t%~s
#* wA"At *A<
,,:+sSi ti'I0~~.# #~
rapidity by which ecdysone induces the early-responding RNA puffs of Drosophila. Thesepuffs respond to ecdysone within minutes andreach their maximal size between 1 and 4 h inlarval salivary glands (Ashburner, 1972) and af-ter 1 h in prepupal salivary glands (Richards,1 976c). According to Ashburner's model, early-responding RNA puffs of Drosophila are directlyregulated by the ecdysone receptor (Ashburneret al., 1974). This model is supported by theexample of puff 74EF, an early-responding RNApuff of Drosophila, to which a cloned ecdysonereceptor is able to bind (cited in Burtis et al.,1 990).Promoters of genes that are directly regulated
by steroid receptors contain a hormone re-sponsive element, which is a short palindromicsequence with a variable degree of conservationbetween different steroid hormones (Evans,1988; Green and Chambon, 1988; Beato, 1989).The DNA binding site (EcRE) of the ecdysonereceptor appears to be an imperfect palin-drome, the more conserved right side of whichhas the consensus sequence of TGA A/C CY(Cherbas et al., 1991). There are three se-quences (starting at -500, -421, and -271)(DiBartolomeis and Gerbi, 1989) in the 718-bppromoter fragment of DNA puff gene 11/9-1identical to this putative EcRE consensus se-quence except that Y is A/T in the Sciara se-quences. This further agrees with the notionthat the Sciara DNA puff promoter of gene11/9-1 is directly controlled by the ecdysone re-ceptor. Sites in DNA puff gene 11/9-1 that binda labeled ecdysone receptor will be experimen-tally determined, but note that our present re-
Figure 10. DNA from Sciara DNA puff 11/9A does not puffin transgenic Drosophila salivary gland chromosomes. Thepolytene chromosome squashes were prepared from pre-pupal salivary glands of Drosophila (line Al 84) transformedwith 8.8 kb of Sciara genomic DNA (see map in Figure 6).(Top) In situ hybridization to salivary gland chromosomesfrom larvae just before puparium formation using a biotin-ylated probe of the 5.5-kb EcoRl fragment that containsSciara DNA puff gene 11/9-1 and upstream DNA. This probebinds only to locus 1 C on the X chromosome (arrow). RNApuff 2B5 of Drosophila is nearby and is puffed at this stage(Ashburner, 1969). (Middle) Polytene chromosomes weresquashed from salivary glands of 6- to 9-h transgenic pre-pupae (PP6-PP9). The arrow marks the locus of Sciara DNAinsertion (1 C) as determined by in situ hybridization (shownabove); the open arrowhead indicates the early-respondingRNA puff 2B5 of Drosophila. (Bottom) Polytene chromo-somes were squashed from salivary glands isolated fromtransgenic 0 h prepupae (PPO). These salivary glands werepreincubated for 6 h at 25 ± 1 °C in Robbs' medium andincubated for 30-60 min in the presence of ecdysone. Ar-rows same as middle panel.
CELL REGULATION
Y
iF
I- m :. -
,t J-. ..,
;.; ..i,
;"§U" 14K.,
*--W
"W4"''
" .1.:,i:
884
Am1 .,.if.I
ik *.I" ..,4-61-

Sciara DNA puff promoter in Drosophila
sults predict that at least some of the bindingsites for the ecdysone receptor must residewithin the 718-bp long promoter fragment ofgene 11/9-1 tested in transgenic Drosophila.Similarly, an ecdysone responsive element wasidentified in the 5' flanking promoter region ofthe Drosophila hsp27 gene (Riddihough andPelham, 1987). However, the E74A gene ofDrosophila seems instead to contain the ecdy-sone responsive element(s) within the first in-tron, -8-14 kb downstream of the start site oftranscription (cited in Burtis et al., 1990).
Induction of gene expression is astage-specific eventIn vitro, in transgenic Drosophila, induction ofthe chimeric Sciara-CAT gene by ecdysone is astage-specific event occurring only in compe-tent prepupal salivary glands and not in larvalor early prepupal salivary glands. In vivo, intransgenic Drosophila, a burst in CAT activity isseen only during the late prepupal and not dur-ing the late larval peak in the ecdysone titer.Because hormonal induction of the hybrid genedoes not occur in transgenic salivary glands oflate third instar larvae in vitro and in vivo, theremust be either a repressor-like factor that pre-vents induction of hybrid gene expression in latethird instar larvae or, alternatively, a proteinfactor that facilitates induction of hybrid geneexpression in late prepupae. The latter possi-bility of a prepupal co-inducer may be suggestedby the necessity of protein synthesis to achievecompetence (Richards, 1976b,c). The formerpossibility of a larval repressor is supported bythe observation that puffing of 93F, an early-responding puff found only in late prepupae ofDrosophila, could be prematurely induced in lar-val salivary glands if they were preincubated inthe presence of a protein synthesis inhibitor(Richards, 1 976c). Similar experiments are dif-ficult to carry out in our system: we have notbeen able to show a significant premature in-duction of CAT activity by preincubating larvalsalivary glands in the presence of cycloheximideeven if the drug was rigorously washed out be-fore hormonal induction (data not shown).
Regardless of the nature of such stage-spe-cific regulatory proteins, they should show somedegree of evolutionary conservation betweenthe two fly species because the Sciara promoterof DNA puff gene 11/9-1 was assayed in trans-genic Drosophila. This evolutionary conservationis remarkable in light of the obvious morpho-logical differences seen at the late larval, pre-pupal, and pupal stages between Sciara and
Drosophila. Our construct of the promoter fromSciara coprophila DNA puff gene 11/9-1 is thefirst description of sequences that are signifi-cantly active only in Drosophila salivary glandsfrom late prepupae but not from larvae, therebyallowing future studies in Drosophila on the mo-lecular mechanism restricting the hormonal re-sponse to just one specific stage.
Materials and methods
Plasmid construction and P-elementtransformationTo study transcriptional control, a promoter fragment ofSciara DNA puff gene 11/9-1 was linked to the reporter geneCAT (Figure 2). The Sciara DNA puff promoter was isolatedas a 718-bp long EcoRV-Bal I fragment. It contains DNAsequences from -616 to +102 relative to the cap site ofthe gene (DiBartolomeis and Gerbi, 1989). This promoterfragment was ligated to the unique Sal I site (filled in byreverse transcriptase) of the polylinker of the P-elementvector CarCAT-1 previously described by Mitsialis et al.(1987). The CarCAT-1 vector is composed of the reportergene CAT flanked by SV40 sequences (providing RNA pro-cessing signals) that were inserted as a 1.7-kb Bgl l-BamHlfragment into the Hpa I site of the P-element vector Carnegie20 (Rubin and Spradling, 1983). The Sciara DNA fragmentwas linked to the reporter gene CAT within the 5' untrans-lated sequences of SV40. Sequencing the junction indicateda short open reading frame from the Sciara ATG before theATG start codon for CAT protein which occurs 74 nucleo-tides further downstream. Carnegie 20 also contains a 7.2-kb Drosophila fragment with the rosy gene (Rubin andSpradling, 1983).
For studies on amplification or puffing, an 8.8-kb fragmentof Sciara genomic DNA containing DNA puff genes 11/9-1and 11/9-2 and flanking DNA was used (Figure 6). The 8.8-kb-long fragment of Sciara DNA puff 11/9A was isolated asa 1 .6-kb Xho I-Sal I fragment and a 7.2-kb Sal I-Sal I fragment(the downstream Sal I site comes from the X vector) (Di-Bartolomeis and Gerbi, 1989). The 1 .6-kbXho I-Sal I fragmentwas ligated into the unique Sal I site of the Carnegie 20vector (Rubin and Spradling, 1983). The unique Sal I site ofthis hybrid plasmid was used to insert the 7.2-kb Sal I-Sal Ifragment resulting in a P-element vector carrying 8.8-kbSciara DNA sequences.The resulting constructs were injected into embryos of
the strain cn; ry2 of Drosophila melanogaster, together withthe helper plasmid pir25.7w.c. (Karess and Rubin, 1984) atDNA concentrations of 0.5 and 0.1 mg/ml, respectively. P-element transformation was carried out as previously de-scribed (Rubin and Spradling, 1982; Spradling and Rubin,1982). Surviving Go flies were individually mated with theparental strain and individual G, transformants of separateGo crosses were used to establish homozygous lines.
Southern blot analysisGenomic DNA was isolated from transgenic Drosophila es-sentially as described by Jowett (1986), restricted, and sub-jected to gel electrophoresis on 1% agarose gels run at 70V in 40 mM Tris-acetate, 1 mM EDTA (TAE) buffer (Sambrooket al., 1989). After this the DNA was transferred to nylon(Gene Screen Plus, New England Nuclear-DuPont, Boston,MA) or nitrocellulose filters (Sartorius, distributed by Van-guard Intl., Neptune, N.J.). Probes were labeled with a-32PdATP (New England Nuclear-DuPont) using a random prim-
Vol. 2, November 1991 885

B. Bienz-Tadmor et al.
ing kit (Boehringer Mannheim, Indianapolis, IN). Hybridiza-tion to nitrocellulose filters followed Sambrook et al., (1989)with hybridization at 650C and the final posthybridizationwash in 0.1X SSC at 610C. Hybridization to nylon filtersfollowed the manufacturer's preferred method (Sartorius).Subsequent autoradiography used XAR 5 X-ray film (East-man Kodak, Rochester, NY) and Cronex Lightening-Plus in-tensifying screens (Eastman Kodak) at -700C.
Polytene chromosome analysisDetails of in situ hybridization were basically as describedby Engels et al. (1 986). A 5.5-kb EcoRI fragment containingSciara DNA puff gene 11/9-1 and upstream DNA was labeledfor use as a probe with biotin-1 6-dUTP (Boehringer Mann-heim) using a random priming kit (Boehringer Mannheim).The probe was subsequently detected with streptavidin-al-kaline phosphatase (Gibco, BRL, Gaithersburg, MD), stainingwith nitro-blue tetrazolium (NBT) and 5-bromo-4 chloro-3indolyl phosphate (BCIP) as described by Engels etal. (1986).TMAX-1 00 film (Eastman Kodak) was used for photography.
For in vitro induction studies, salivary glands from 0-hprepupae (PPO) of transgenic Drosophila were isolated andincubated for 6 h at 25 ± 1 0C in Robbs' tissue culture me-dium (Robbs, 1969) followed by a 30- to 60-min incubationin 400 Al Robbs' medium containing 1 Al of 1 mg/ml 20-OHecdysone. Salivary glands were squashed in 45% acetic acidfor polytene chromosome analysis.
CAT assaysTransformed animals were grouped into developmentalstages according to Bodenstein (1965). For the larval stages,hours were counted from the time of oviposition. Well-con-ditioned flies were allowed to lay eggs for 1 h and thenremoved from the vials. Larvae were harvested after thetime indicated for each experiment. For the prepupal andpupal stages, hours were counted from the time of pupariumformation (Fraenkel and Bhaskaran, 1973; Bainbridge andBownes, 1981). Positions of individual prepupae weremarked on the food vials at the time of puparium formation,and prepupae or pupae were collected after the number ofhours indicated for each experiment. All vials were incubatedat 25 ± 1 °C. Animals were arrested at the desired devel-opmental stage by putting them on ice for up to severalhours.Whole animals or dissected tissues were used to prepare
extracts for CAT assays. Tissues were dissected in an ice-cold TSEL buffer 10.25 M tris(hydroxymethyl)aminomethane-HCI, pH 7.8,0.4 M sucrose, 5 mM EDTA, 0.5 mM leupeptin].Typically, 10 whole animals or dissected tissues were ho-mogenized in 150 MI TSEL buffer containing 1 mg/ml bovineserum albumin (BSA) to stabilize the CAT protein. The pres-ence of BSA precluded measurement of the total proteinamount in the homogenate. For each sample, 100 Al of ho-mogenate were assayed for CAT activity in an overnightincubation at 370C as described by Mitsialis et al. (1987).
In vitro experimentsTransformed animals were staged as described above, ex-cept that they were never held on ice. Salivary glands ofthree to five larvae or prepupae of the indicated develop-mental stage were dissected in a drop of Robbs' tissue cul-ture medium (Robbs, 1969) and immediately transferred intodissection wells containing 400 AI of Robbs' medium with1 ztL of ethanol or 1 Al of 1 mg/ml 20-OH ecdysone (dissolvedin ethanol; Sigma, St. Louis, MO) to result in a final ecdysoneconcentration in the medium of 5 x 10-6 M. The mediumused for preincubation of the salivary glands did not containethanol or ecdysone. For the inhibitor studies, cycloheximide
(Sigma) and actinomycin D (Sigma) were dissolved in waterand added to the medium in a volume of 4 jA to give finalconcentrations of 7 x 10-5 and 9 x 10' M, respectively.The salivary glands were incubated for the indicated amountof time (±10 min) at 25 ± 10C by covering the dissectionwell with a microscope slide. After the incubation period,the glands were immediately transferred into 150 ul of TSELbuffer (Mitsialis et al., 1987). The entire homogenate wasassayed for CAT activity, as described above. Each in vitroexperiment was repeated two to three times, and the resultswere reproducible in these independent experiments.
AcknowledgmentsWe thank Peter Cherbas for his helpful advice regarding ourin vitro experiments, Michael Conboy for technical assis-tance with the injections, Nan Wu and Fedya Urnov forcomputer help, and Donna Meinville for typing assistance.We also thank Susan DiBartolomeis, Barbara Stebbins-Boaz,Rocco Savino, and Josephine Spitzer for thoughtful com-ments on this manuscript. This work was supported by NIHGM-35929 to S.A.G.
Received: March 4, 1991.Revised and accepted: August 6, 1991.
References
Amabis, D.C., and Amabis, J.M. (1984a). Effects on ecdys-terone in polytene chromosomes of Trichosia pubescens.Dev. Biol. 102, 1-9.
Amabis, D.C., and Amabis, J.M. (1984b). Hormonal controlof gene amplification and transcription in the salivary glandchromosomes of Trichosia pubescens. Dev. Biol. 102, 1 0-20.Ashburner, M. (1967). Patterns of puffing activity in the sal-ivary gland chromosomes of Drosophila. Autosomal puffingpatterns in a laboratory stock of Drosophila melanogaster.Chromosoma 21, 398-428.
Ashburner, M. (1969). Patterns of puffing activity in the sal-ivary gland chromosomes of Drosophila. The X-chromosomepuffing patterns of D. melanogaster and D. simulans. Chro-mosoma 27, 47-63.
Ashburner, M. (1971). Induction of puffs in polytene chro-mosomes of in vitro cultured salivary glands of Drosophilamelanogaster by ecdysone and ecdysone analogues. Nature230, 222-224.
Ashburner, M. (1972). Patterns of puffing activity in the sal-ivary gland chromosomes of Drosophila. Induction by ec-dysone in salivary glands of Drosophila melanogaster cul-tured in vitro. Chromosoma 38, 255-281.
Ashburner, M. (1973). Sequential gene activation by ecdy-sone in polytene chromosomes of Drosophila melanogaster.Dependence upon ecdysone concentration. Dev. Biol. 35,47-61.
Ashburner, M. (1974). Sequential gene activation by ecdy-sone in polytene chromosomes of Drosophila melanogaster.The effects of inhibitors of protein synthesis. Dev. Biol. 39,141-157.
Ashburner, M. (1990). Puffs, genes and hormones revisited.Cell 61, 1-3.
Ashburner, M., Chihara, C., Meltzer, P., and Richards, G.(1974). Temporal control of puffing activity in polytene chro-
CELL REGULATION886

Sciara DNA puff promoter in Drosophila
mosomes. Cold Spring Harbor Symp. Quant. Biol. 38, 655-662.Ashburner, M., and Richards, G. (1976). Sequential geneactivation by ecdysone in polytene chromosomes of Dro-sophila melanogaster. Consequences of ecdysone with-drawal. Dev. Biol. 54, 241-255.
Bainbridge, S.P., and Bownes, M. (1981). Staging the meta-morphosis of Drosophila melanogaster. J. Embryol. Exp.Morphol. 66, 57-80.
Bainbridge, S.P., and Bownes, M. (1988). Ecdysteroid titersduring Drosophila metamorphosis. Insect Biochem. 18, 185-197.
Beato, M. (1989). Gene regulation by steroid hormones. Cell56, 335-344.Beermann, W. (1952). Chromomerenkonstanz und spezi-fische Modifikation der Chromosomenstruktur in der En-twicklung und Organdifferenzierung von Chironomus ten-tans. Chromosoma 5, 139-198.
Bodenstein, D. (1965). The postembryonic development ofDrosophila. In: Biology of Drosophila, ed. M. Demerec, NewYork: Hafner Publishing Company, 275-367.
Burtis, K.C., Thummel, C.S., Jones, C.W., Karim, F.D., andHogness, D.S. (1990). The Drosophila 74EF early puff con-tains E74, a complex ecdysone-inducible gene that encodestwo ets-related proteins. Cell 61, 85-99.Carpenter, F. (1991). Treatise on Invertebrate Paleontology:Hexapoda, Insecta. Lawrence, KS: Geological Society ofAmerica and the University of Kansas-Museum of Inver-tebrate Paleontology (in press).Chao, A.T., and Guild, G.M. (1986). Molecular analysis ofthe ecdysterone-inducible 2B5 early puff in Drosophila mel-anogaster. EMBO J. 5, 143-150.
Cherbas, L., Lee, K., and Cherbas, P. (1991). Identificationof ecdysone response elements by analysis of the DrosophilaEip 28/29 gene. Genes and Dev. 5, 120-131.
Crouse, H.V. (1968). The role of ecdysone in DNA-puff for-mation and DNA synthesis in the polytene chromosomesof Sciara coprophila. Proc. Natl. Acad. Sci. USA 61, 971-978.
Crouse, H.V., and Keyl, H.G. (1968). Extra replications inthe DNA puffs of Sciara coprophila. Chromosoma 25, 357-364.
DiBartolomeis, S.M., and Gerbi, S.A. (1989). Molecularcharacterization of DNA puff 11/9A genes in Sciara copro-phila. J. Mol. Biol. 210, 531-540.
Engels, W.R., Preston, C.R., Thompson, P., and Eggleston,W.B. (1986). In situ hybridization to Drosophila salivarychromosomes with biotinylated DNA probes and alkalinephosphatase. Focus (Bethesda Research Lab) 8, 6-8.
Evans, R.M. (1988). The steroid and thyroid hormone re-ceptor superfamily. Science 240, 889-895.
Fraenkel, G., and Bhaskaran, G. (1973). Pupariation and pu-pation in cyclorrhaphous flies (Diptera): terminology and in-terpretation. Ann. Entomol. Soc. Am. 66, 418-422.
Gabrusewycz-Garcia, N. (1964). Cytological and autoradio-graphic studies in Sciara coprophila salivary gland chro-mosomes. Chromosoma 15, 312-344.
Gabrusewycz-Garcia, N., and Kleinfeld, R.G. (1966). A studyof the nucleolar material of Sciara coprophila. J. Cell Biol.29, 347-359.
Glover, D.M., Zaha, A., Stocker, A.J., Santelli, R.V., Pueyo,M.T., deToledo, S.M., and Lara, F.J.S. (1982). Gene ampli-fication in Rhynchosciara salivary gland chromosomes. Proc.Natl. Acad. Sci. USA 79, 2947-2951.
Green, S., and Chambon, P. (1988). Nuclear receptors en-hance our understanding of transcription regulation. TrendsGenet. 4, 309-314.
Hinton, H.E. (1976). Notes on neglected phases in meta-morphosis, and a reply to J.M. Whitten. Ann. Entomol. Soc.Am. 69, 560-566.
Jowett, T. (1986). Preparation of nucleic acids. In: Drosophila,a Practical Approach, ed. D.B. Roberts, Oxford: IRL Press,275-286.
Karess, R.E., and Rubin, G.M. (1984). Analysis of P trans-posable element functions in Drosophila. Cell 38, 135-146.
Korge, G. (1987). Polytene chromosomes. In: Results andProblems in Cell Differentiation, vol. 14, Structure andFunction of Eukaryotic Chromosomes, ed. W. Hennig, Berlin:Springer-Verlag, 27-58.
Meyerowitz, E.M., Rahavan, K.V., Mathers, P.H., and Roark,M. (1987). How Drosophila larvae make glue: control of Sgs-3 expression. Trends Genet. 3, 288-292.
Mitsialis, S.A., Spoerel, N., Leviten, M., and Kafatos, F.C.(1987). A short 5'-flanking region is sufficient for develop-mentally correct expression of moth chorion genes in Dro-sophila. Proc. NatI. Acad. Sci. USA 84, 7987-7991.
Orr, W., Komitopoulou, K., and Kafatos, F.C. (1984). Mutantssuppressing in trans chorion gene amplification in Drosoph-ila. Proc. NatI. Acad. Sci. USA 81, 3773-3777.
Parks, S., Wakimoto, B., and Spradling, A. (1986). Replicationand expression of an X-linked cluster of Drosophila choriongenes. Dev. Biol. 117, 294-305.
Rasch, E.M. (1970). Two-wavelength cytophotometry ofSciara salivary gland chromosomes. In: Introduction toQuantitative Cytochemistry, vol. 2, eds. G.L. Wied and G.F.Bahr, New York: Academic Press Inc., 335-355.
Richards, G. (1976a). The control of prepupal puffing pat-terns in vitro: implications for prepupal ecdysone titers inDrosophila melanogaster. Dev. Biol. 48, 191-195.
Richards, G. (1976b). Sequential gene activation by ecdy-sone in polytene chromosomes of Drosophila melanogaster.The mid prepupal puffs. Dev. Biol. 54, 256-263.
Richards, G. (1 976c). Sequential gene activation by ecdysonein polytene chromosomes of Drosophila melanogaster. Thelate prepupal puffs. Dev. Biol. 54, 264-275.
Richards, G. (1980). Ecdysteroids and puffing in Drosophilamelanogaster. In: Progress in Ecdysone Research. Devel-opments in Endocrinology, vol. 7, ed. J.A. Hoffmann, Am-sterdam: Elsevier/North-Holland, Biomedical Press, 363-378.
Richards, G. (1981). The radioimmune assay of ecdysteroidtiters in Drosophila melanogaster. Mol. Cell. Endocrinol. 21,181-197.
Riddihough, G., and Pelham, H.R.B. (1987). An ecdysoneresponse element in the Drosophila hsp27 promoter. EMBOJ. 6, 3729-3734.
Robbs, J.A. (1969). Maintenance of imaginal discs of Dro-sophila melanogaster in chemically defined media. J. CellBiol. 41, 876-885.
Vol. 2, November 1991 887

B. Bienz-Tadmor et al.
Rubin, G.M., and Spradling, A.C. (1982). Genetic transfor-mation of Drosophila with transposable element vectors.Science 218, 348-353.
Rubin, G.M., and Spradling, A.C. (1983). Vectors for P-ele-ment mediated gene transfer in Drosophila. Nucleic AcidsRes. 11, 6341-6351.
Rudkin, G.T., and Corlette, S.L. (1957). Disproportionatesynthesis of DNA in a polytene chromosome region. Proc.NatI. Acad. Sci. USA 43, 964-968.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). MolecularCloning, a Laboratory Manual, Second edition, Cold SpringHarbor: Cold Spring Harbor Laboratory Press.
Segraves, W.A., and Hogness, D.S. (1990). The E75 ecdy-sone-inducible gene responsible for the early puff in Dro-sophila encodes two new members of the steroid receptorsuperfamily. Genes Dev. 4, 204-219.
Spradling, A.C., and Rubin, G.M. (1982). Transposition ofcloned P-elements into Drosophila germ line chromosomes.Science 218, 341-347.
Stocker, A.J., and Pavan, C. (1974). The influence of ecdys-terone on gene amplification, DNA synthesis and puff for-
mation in the salivary gland chromosomes of Rhynchosciarahollaenderi. Chromosoma 45, 295-319.Stocker, A.J., Troyano-Pueyo, M., Pereira, C.D., and Lara,F.J.S. (1984). Ecdysteroid titers and changes in chromosomalactivity in the salivary glands of Rhynchosciara americana.Chromosoma 90, 26-38.Thummel, C.S., Burtis, K.C., and Hogness, D.S. (1990). Spa-tial and temporal pattern of E74 transcription during Dro-sophila development. Cell 61, 101-115.Urness, L.D., and Thummel, C.S. (1990). Molecular inter-actions within the ecdysone regulatory hierarchy: DNAbinding properties of the Drosophila ecdysone-inducibleE74A protein. Cell 63, 47-61.Wakimoto, B.T., Kalfayan, L.J., and Spradling, A.C. (1986).Developmentally regulated expression of Drosophila choriongenes introduced at diverse chromosomal positions. J. Mol.Biol. 187, 33-45.Whitten, J.M. (1976). Definition of insect instars in terms of"apolysis" or "ecdysis." Ann. Entomol. Soc. Am. 69, 556-559.Yamamoto, K.R. (1985). Steroid receptor transcription ofspecific genes and gene networks. Annu. Rev. Genet. 19,209-252.
CELL REGULATION888