bioe 109 summer 2009 lecture 10-part ii life history evolution
Post on 22-Dec-2015
223 views
TRANSCRIPT

BIOE 109Summer 2009
Lecture 10-Part IILife history evolution

What is “life history”?
A typical life history of any animal-
Birth
maturity
Reproduction Death
Reproductionceases

What are “life history” characters?

What are “life history” characters?
• from a “fitness” perspective, there are only two important events in life: reproduction and death.

What are “life history” characters?
• from a “fitness” perspective, there are only two important events in life: reproduction and death.
• traits that determine the timing and details of these events are termed life history characters:

What are “life history” characters?
• from a “fitness” perspective, there are only two important events in life: reproduction and death.
• traits that determine the timing and details of these events are termed life history characters:
1. Age at first reproduction (time to maturity)
2. Total life span
3. Mode and frequency of reproduction
4. Fecundity (no. of offspring produced)
5. Parental care
6. Viability of off-springs

Darwinian Demon
• Mature at birth
• Reproduce frequently and in large numbers
• Produce high quality offspring always
• Live forever (at least live very long)
Do Darwinian Demons exist in nature?

Do Darwinian Demons exist in nature?
• Mature at birth- yes!• Produce frequently and in large amounts-No!• Live forever-No!
Thrips egg mite (Adactylidium sp.)

X-ray of female brown kiwi
1/6 its weight!!
• Mature at birth- No!• Produce high quality offspring frequently- yes!• Live long-sort of! (20 years)

Remember!
• Amount of energy an organism can harvest is finite and biological processes take time!
• trade-offs between life history traits are unavoidable!
• variation in life-histories are due to differences in the allocation of energy. • Organisms that find “optimal balance” between costs and benefits are favored by natural selection.
• The “optimal” solution might be different in different environments.

Some life history questions:
1. Why do individuals age and die?

Some life history questions:
1. Why do individuals age and die?
2. How many offspring should an individual produce in any given year?

Some life history questions:
1. Why do individuals age and die?
2. How many offspring should an individual produce in any given year?
3. How large should each offspring be?

Why do organisms age and die?

Why do organisms age and die?
• Aging or senescence, is a late life decline in an individual’s fertility and probability of survival.
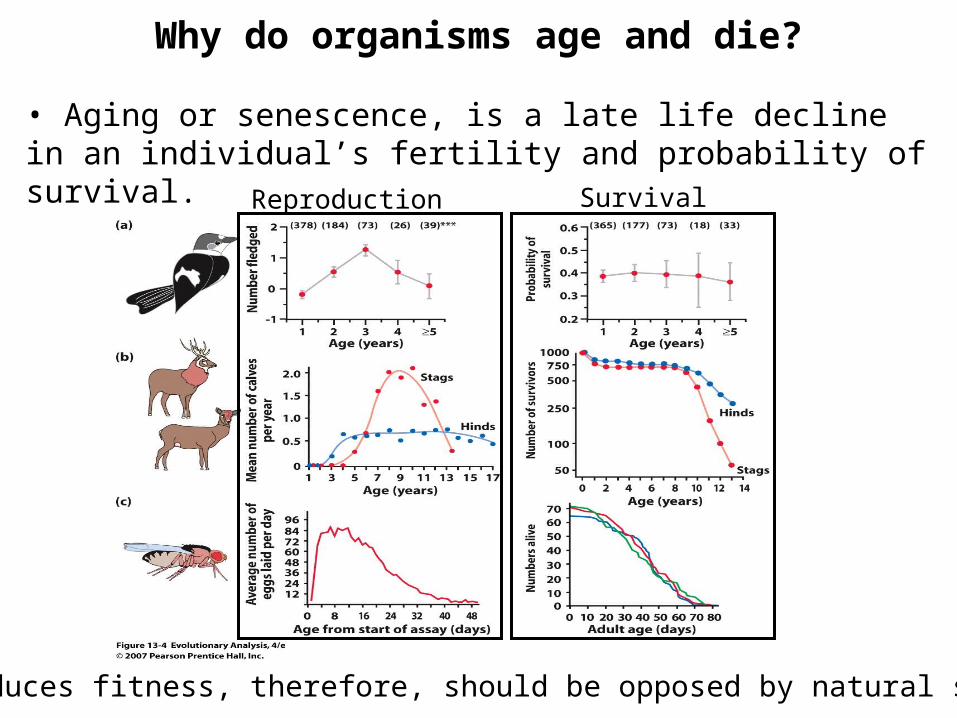
Reproduction Survival
Why do organisms age and die?
• Aging or senescence, is a late life decline in an individual’s fertility and probability of survival.
Aging reduces fitness, therefore, should be opposed by natural selection

Why do organisms age and die?
• two main theories that try to explain aging are “rate-of-living” theory and the evolutionary theory.

Why do organisms age and die?
• two main theories that try to explain aging are “rate-of-living” theory and the evolutionary theory.
1. The “rate-of-living” theory (ROL)
• aging caused by the accumulation of irreparable damage to cells and tissues.

Why do organisms age and die?
• two main theories that try to explain aging are “rate-of-
living” theory and the evolutionary theory.
1. The “rate-of-living” theory (ROL)
• aging caused by the accumulation of irreparable damage to cells and tissues.
• organisms have reached their limit of biologically possible repair.

Why do organisms age and die?
• two main theories that try to explain aging are “rate-of-living” theory and the evolutionary theory.
1. The “rate-of-living” theory (ROL)
• aging caused by the accumulation of irreparable damage to cells and tissues.
• organisms have reached their limit of biologically possible repair (they have been already selected enough!)
• populations lack the genetic variation that would enable them to evolve more effective repair mechanisms

The ROL theory makes two important predictions:

The ROL theory makes two important predictions:
1. Lifespan should correlate negatively with metabolic rate.

The ROL theory makes two important predictions:
1. Lifespan should correlate negatively with metabolic rate.
• predicts that by-products of metabolism cause cell and tissue damage (and ultimately death).
Met
abol
ic r
ate
Lifespan

Testing the rate-of-living theory
Wide variation of energy expenditures among mammals!
amount of energy expended/gram of tissue in lifetime

The ROL theory makes two important predictions:
1. Lifespan should correlate negatively with metabolic rate.
• predicts that by-products of metabolism cause cell and tissue damage (and ultimately death).
2. Longevity should not respond to selection.

The ROL theory makes two important predictions:
1. Lifespan should correlate negatively with metabolic rate.
• predicts that by-products of metabolism cause cell and tissue damage (and ultimately death).
2. Longevity should not respond to selection.
• predicts no genetic variation should exist for genes increasing longevity.

Selection for increased life span in Drosophila
Luckinbill et al. 1984

The ROL theory makes two important predictions:
1. Lifespan should correlate negatively with metabolic rate. NO
• predicts that by-products of metabolism cause cell and tissue damage (and ultimately death).
2. Longevity should not respond to selection. NO
• predicts no genetic variation should exist for genes increasing longevity.

2. The evolutionary theory of aging

2. The evolutionary theory of aging
• aging and death caused by incomplete repair of cell and tissue damage throughout life that leads to gradual decay and eventually complete collapse of metabolic system.

2. The evolutionary theory of aging
• aging and death caused by incomplete repair of cell and tissue damage throughout life that leads to gradual decay and eventually complete collapse of metabolic system.
Why repair is incomplete?

2. The evolutionary theory of aging
• aging and death caused by incomplete repair of cell and tissue damage throughout life that leads to gradual decay and eventually complete collapse of metabolic system. Why repair is incomplete?
1. Late onset of deleterious mutations

2. The evolutionary theory of aging
• aging and death caused by incomplete repair of cell and tissue damage throughout life that leads to gradual decay and eventually complete collapse of metabolic system.
Why repair is incomplete?
1. Late onset of deleterious mutations
• many examples known in humans (Huntington’s, some cancers), Drosophila, houseflies.

2. The evolutionary theory
• aging and death caused by incomplete repair of cell and tissue damage throughout life that leads to gradual decay and eventually complete collapse of metabolic system. Why repair is incomplete?
1. Late onset of deleterious mutations
• many examples known in humans (Huntington’s, some cancers), Drosophila, houseflies.
2. Trade-offs between fecundity and longevity

2. The evolutionary theory
• aging and death caused by incomplete repair of cell and tissue damage throughout life that leads to gradual decay and eventually complete collapse of metabolic system. Why repair is incomplete?
1. Late onset of deleterious mutations
• many examples known in humans (Huntington’s, some cancers), Drosphila, houseflies.
2. Trade-offs between fecundity and longevity
• genes that have a beneficial effect early in life but a detrimental effect later are said to exhibit “antagonistic pleiotropy”.

Evidence for antagonistic pleiotropy:the methuselah locus in Drosophila

How many offspring should an individual produce in a given reproductive attempt?

How many offspring should an individual produce in a given reproductive attempt?
-produce 100 eggs-diameter= 300 micrometers
-produce 10-50 million eggs-diameter= 50-55 micrometers

How many offspring should an individual produce in a given reproductive attempt?
• individuals of some species reproduce only once and then die (e.g. pacific salmon).
• this is called semelparity.

How many offspring should an individual produce in a given reproductive attempt?
• individuals of some species reproduce only once and then die (e.g. pacific salmon).
• this is called semelparity.
• individuals of most species reproduce several times throughout life.
• this is termed iteroparity.

How many offspring should an individual produce in a given reproductive attempt?
• individuals of some species reproduce only once and then die (e.g. pacific salmon).
• this is called semelparity.
• individuals of most species reproduce several times throughout life.
• this is termed iteroparity.
• for iteroparous species, how many offspring should an individual produce in a given attempt?

How many offspring should an individual produce in a given reproductive attempt?
• individuals of some species reproduce only once and then die (e.g. pacific salmon).
• this is called semelparity.
• individuals of most species reproduce several times throughout life.
• this is termed iteroparity.
• for iteroparous species, how many offspring should an individual produce in a given attempt?
• most intensively studied in birds – what is the optimal clutch size?

Lack’s hypothesis for the evolution of clutch size

Lack’s hypothesis for the evolution of clutch size
• proposed by David Lack in 1947.
David Lack (1910 – 1973)

Lack’s hypothesis for the evolution of clutch size
• proposed by David Lack in 1947. • assumes a trade-off between probability of individual survival and clutch size:

Lack’s hypothesis for the evolution of clutch size
• proposed by David Lack in 1947. • assumes a trade-off between probability of individual survival and clutch size:

Lack’s hypothesis for the evolution of clutch size
• proposed by David Lack in 1947.• assumes a trade-off between probability of individual survival and clutch size:
Selection will favor clutch size that produces max. surviving offspring

One test of Lack’s hypothesis
• Boyce and Perrins (1987) studied 4,489 clutches of great tits (Parus major) from 1960-1982.

One test of Lack’s hypothesis
• Boyce and Perrins (1987) studied 4,489 clutches of great tits (Parus major) from 1960-1982. Range in clutch size 1-17

One test of Lack’s hypothesis
• Boyce and Perrins (1987) studied 4,489 clutches of great tits (Parus major) from 1960-1982. Range in clutch size 1-17 Mean clutch size 8.53

One test of Lack’s hypothesis
• Boyce and Perrins (1987) studied 4,489 clutches of great tits (Parus major) from 1960-1982. Range in clutch size 1-17 Mean clutch size 8.53 Most productive 12clutch size

One test of Lack’s hypothesis
Most productive clutch size= 12
Actual mean clutch size= 8.53
Why such discrepancy?

What other assumptions underlie Lack’s hypothesis?

What other assumptions underlie Lack’s hypothesis?
1. No trade-offs between reproductive efforts across years.

What other assumptions underlie Lack’s hypothesis?
1. No trade-offs between reproductive efforts across years.
• many studies have added extra eggs to nests and found adverse effects on subsequent reproduction in the following year.

What other assumptions underlie Lack’s hypothesis?
2. Clutch size only affects viability of offspring.

What other assumptions underlie Lack’s hypothesis?
2. Clutch size only affects viability of offspring. • large clutches may reduce reproductive success of progeny.

What other assumptions underlie Lack’s hypothesis?
2. Clutch size only affects viability of offspring. • large clutches may reduce reproductive success of progeny. Example: Schluter and Gustafsson’s (1996) study on collared flycatchers

Clutch size can affect offspring fitness

What other assumptions underlie Lack’s hypothesis?
3. No year-to-year variation in optimal clutch size.

What other assumptions underlie Lack’s hypothesis?
3. No year-to-year variation in optimal clutch size.
• clutch size is not fixed for given genotype. It is phenotypically plastic.

What other assumptions underlie Lack’s hypothesis?
3. No year-to-year variation in optimal clutch size.
• clutch size is not fixed for given genotype. It is phenotypically plastic.
• thus, optimal clutch size may vary from year-to-year depending on availability of resources, etc.
Lack’s hypothesis is a null model!

How big should each offspring be?

How big should each offspring be?
• many studies have established a trade-off between egg size and egg number.

How big should each offspring be?
• many studies have established a trade-off between egg size and egg number.
26 species of fish 49 species of fruit flies

A model by Smith and Fretwell (1974)
Assumption 1: A trade-off between number and size of offspring

A model by Smith and Fretwell (1974)
Assumption 2: Individual offspring will have better chance of survivalIf they are larger.

A model by Smith and Fretwell (1974)
Result: The optimal offspring size is intermediate

A model by Smith and Fretwell (1974)
this is parent-offspring conflict!
Result: The optimal offspring size is intermediate

Recap
• differences in life-history traits involve differences in the allocation of energy. • trade-offs between life history traits are unavoidable! Questions:
1. Why do individuals age and die?
2. How many offspring should an individual produce in any given year?
3. How large should each offspring be?