bioe 109 summer 2009 lecture 6- part ii molecular evolution
Post on 20-Dec-2015
216 views
TRANSCRIPT

BIOE 109Summer 2009
Lecture 6- Part IIMolecular evolution

The mechanism of inheritance
circa 1865: Mendel’s work demonstrates “factors” in pea plants that are inherited independentlyfrom one another.
circa 1900: rediscovery of Mendel’s work
1910-1915: TH Morgan inferred existence of “genes”and mapped their locations on chromosomes

“Classical” versus “balanced” views of genome structure

“Classical” versus “balanced” views of genome structure
• controversy began in the 1920’s with the establishment of two schools of genetics.

“Classical” versus “balanced” views of genome structure
• controversy began in the 1920’s with the establishment of two schools of genetics.
• the “Naturalists” studied natural populations (e.g. Dobzhansky, Mayr).

“Classical” versus “balanced” views of genome structure
• controversy began in the 1920’s with the establishment of two schools of genetics.
• the “Naturalists” studied natural populations (e.g. Dobzhansky, Mayr).
• the “Mendelians” studied genetics exclusively in the laboratory (e.g., Morgan, Sturtevant, Muller).

Classical
+ + - + + +
+ + + + + +

Classical
+ + - + + +
+ + + + + +
+ = “wild type” allele- = deleterious recessive allele

Classical Balanced
+ + - + + +
+ + + + + +
A1 B2 C1 D4 E3 F6
A3 B2 C4 D5 E5 -
+ = “wild type” allele- = deleterious recessive allele

Classical Balanced
+ + - + + +
+ + + + + +
A1 B2 C1 D4 E3 F6
A3 B2 C4 D5 E5 -
Most loci homozygous Most loci heterozygousfor wild type alleles

Classical Balanced
+ + - + + +
+ + + + + +
A1 B2 C1 D4 E3 F6
A3 B2 C4 D5 E5 -
Most loci homozygous Most loci heterozygousfor wild type alleles
Polymorphism rare Polymorphism common

Why is this distinction important?
Classical Balanced

Why is this distinction important?
Classical Balanced
Speciation Difficult Easy(mutation- (opportunity- limited) limited)

Why is this distinction important?
Classical Balanced
Speciation Difficult Easy(mutation- (opportunity- limited) limited)
Selection Purifying Balancing

Why is this distinction important?
Classical Balanced
Speciation Difficult Easy(mutation- (opportunity- limited) limited)
Selection Purifying Balancing
Population Inter > Intra Intra > Intervariation

Why is this distinction important?
Classical Balanced
Speciation Difficult Easy(mutation- (opportunity- limited) limited)
Selection Purifying Balancing
Population Inter > Intra Intra > Intervariation
Polymorphism transient balanced(short-lived) (long-lived)

Allozyme electrophoresis setup

Starch gel stained for Phosphoglucomutase (Pgm)

Extensive allozyme variation exists in nature
Vertebrates(648 species)

Extensive allozyme variation exists in nature…
…so this confirms the balanced view?
Vertebrates(648 species)

Extensive allozyme variation exists in nature…
…so this confirms the balanced view?
Vertebrates(648 species)
NO! MOST POLYMORPHISMS ARE NEUTRAL!

The neutral theory of molecular evolution
• first proposed by Motoo Kimura in 1968.

The neutral theory of molecular evolution
• first proposed by Motoo Kimura in 1968.
• two observations led Kimura to develop neutral theory:

The neutral theory of molecular evolution
• first proposed by Motoo Kimura in 1968.
• two observations led Kimura to develop neutral theory:
1. “Excessive” amounts of protein (allozyme) polymorphism
2. Molecular clock

2. The molecular clock • first reported by Zuckerkandl and Pauling in 1962.

2. The molecular clock • first reported by Zuckerkandl and Pauling in 1962.

2. The molecular clock • first reported by Zuckerkandl and Pauling in 1962.
Age 7 Age 17 Age 22 Age 46

2. The molecular clock • first reported by Zuckerkandl and Pauling in 1962.
• refers to apparent constant rate of protein evolution over large periods of time.
http://www.blackwellpublishing.com/ridley/video_gallery/LP_What_is_the_molecular_clock.asp

2. The molecular clock • first reported by Zuckerkandl and Pauling in 1962.
Method:

2. The molecular clock • first reported by Zuckerkandl and Pauling in 1962.
Method:
1. Obtain homologous amino acid sequences from a group of taxa.

2. The molecular clock • first reported by Zuckerkandl and Pauling in 1962.
Method:
1. Obtain homologous amino acid sequences from a group of taxa. 2. Estimate divergence times (from the fossil record).

2. The molecular clock • first reported by Zuckerkandl and Pauling in 1962.
Method:
1. Obtain homologous amino acid sequences from a group of taxa. 2. Estimate divergence times (from the fossil record). 3. Assess relationship between protein divergence and evolutionary time.

100 200 300 400 500
Time (millions of years)
No. of aminoacid substitutions
The molecular clock
-globin gene in vertebrates

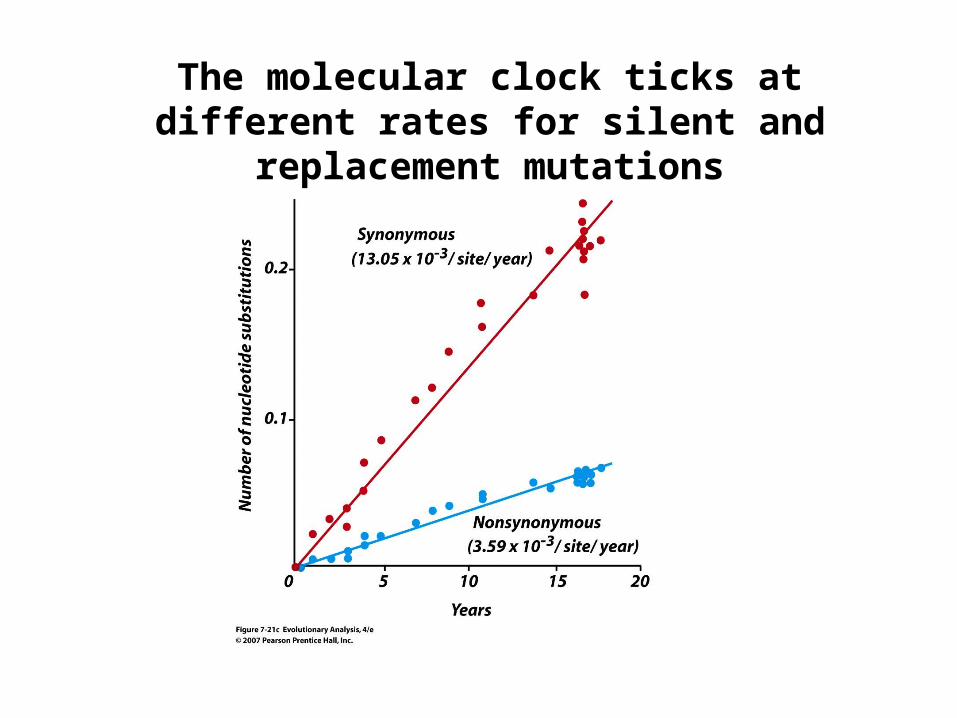
The molecular clock ticks at different rates for silent and replacement mutations

Kimura argued that the molecular clock reflects the action of random drift, not selection!
100 200 300 400 500
Time (millions of years)
No. of aminoacid substitutions
-globin gene in vertebrates

Main features of the neutral theory
1. The rate of protein evolution is roughly constant per site per year.

Main features of the neutral theory
1. The rate of protein evolution is roughly constant per site per year.
- this is the "molecular clock" hypothesis.

Main features of the neutral theory
1. The rate of protein evolution is roughly constant per site per year.
- this is the "molecular clock" hypothesis.
- per site PER YEAR, not per site PER GENERATION
Shorter generation time faster rate of molecular evolutionLonger generation time slower rate of molecular evolution

2. Rate of substitution of neutral alleles equals the mutation rate to neutral alleles.

2. Rate of substitution of neutral alleles equals the mutation rate to neutral alleles. • this rate is unaffected by population size!

3. Rates of protein evolution vary with degree of selective constraint.

3. Rates of protein evolution vary with degree of selective constraint.
• “selective constraint” represents the ability of a protein to “tolerate” random mutations.

3. Rates of protein evolution vary with degree of selective constraint.
• “selective constraint” represents the ability of a protein to “tolerate” random mutations.
• for highly constrained molecules, most mutations are deleterious and few are neutral.

3. Rates of protein evolution vary with degree of selective constraint.
• “selective constraint” represents the ability of a protein to “tolerate” random mutations.
• for highly constrained molecules, most mutations are deleterious and few are neutral.
• for weakly constrained molecules, more mutations are neutral and few are deleterious.
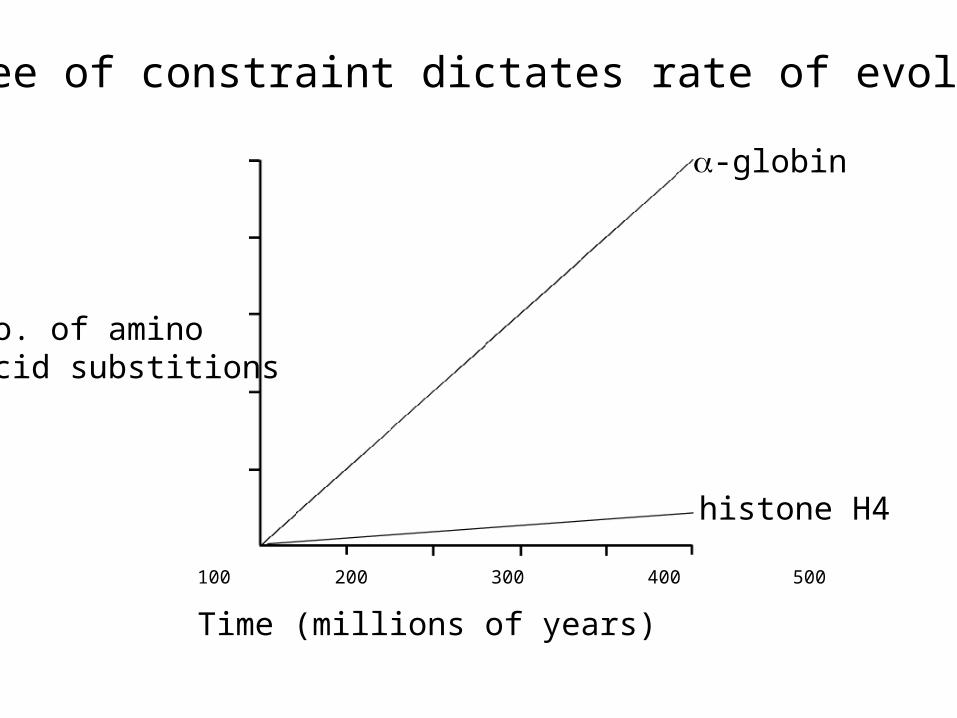
100 200 300 400 500
Time (millions of years)
No. of aminoacid substitions
-globin
histone H4
Degree of constraint dictates rate of evolution

high constraint low slow rate of evolution

high constraint low slow rate of evolution
low constraint high fast rate of evolution