characterization of a major aromatic dna adduct detected in human breast tissues
TRANSCRIPT

Characterization of a Major Aromatic DNA AdductDetected in Human Breast Tissues
Donghui Li,1* Mianying Wang,2 Pervez Firoz Firozi,1 Ping Chang,1
Weiqing Zhang,1 Wanda Baer-Dubowska,3 Bhagavatula Moorthy,4
Suryanarayana V. Vulimiri,3 Regine Goth-Goldstein,5 Eric H. Weyand,6
and John DiGiovanni31Department of Gastrointestinal Medical Oncology, University of Texas,
M.D. Anderson Cancer Center, Houston, Texas2Department of Biomedical Sciences, University of Illinois, College of Medicine at
Rockford, Rockford, Illinois3Department of Carcinogenesis, University of Texas, M.D. Anderson Cancer Center,
Houston, Texas4Department of Pediatrics, Baylor College of Medicine, Houston, Texas
5Lawrence Berkeley National Laboratory, Berkeley, California6Rutgers, The State University of New Jersey, College of Pharmacy, Department of
Pharmaceutical Chemistry, Piscataway, New Jersey
A bulky DNA adduct (Spot 1) was previously detectedin normal adjacent breast tissues of 41% (36/87) ofwomen with breast cancer and in none (0/29) of thenoncancer controls by 32P-postlabeling. To character-ize this adduct, it was chromatographically com-pared with DNA adduct profiles generated in severalin vitro and in vivo experimental systems. First, MCF-7cells were exposed to a number of chemical carcino-gens, that is, benzo[a]pyrene (B[a]P), 4-OH-B[a]P,9-OH-B[a]P, 11-OH-B[a]P, B[a]P-trans-4,5-dihydrodiol,1-nitropyrene, 6-nitrochrysene, dibenzo[a,l]pyrene,benzo[c]phenanthrene, dibenzo[a,h]anthracene,3-methylcholanthrene, and 2-amino-1-methyl-6-pheny-limidazo[4,5-b]pyridine. Spot 1 was detected as a mi-nor adduct in cells treated with B[a]P but not other com-pounds. Second, to determine whether Spot 1 is derivedfrom lipid peroxidation products or estrogen metabo-lites, it was compared with adduct profiles of cells orDNAs exposed to 17�-estradiol, 4-hydroxy estradiol,4-hydroxynonenal, or oxidized oat oil. Spot 1 was notdetectable in these samples. In addition, Spot 1 did notcomigrate with the 1,N2-ethenodeoxyguanosine ad-duct standard. Third, to explore the mechanism ofSpot 1 formation, it was compared with adduct pro-
files detected in DNA or mononucleotides reacted withBPDE, 1-OH-7,8-dihydrodiol of B[a]P, and 3-OH-7,8-dihydrodiol of B[a]P as well as in rats orally treated withB[a]P. Spot 1 comigrated with a minor adduct in BPDE-treated DNA during anion exchange rechromatogra-phy but these two adducts were separated by partitionchromatography. Spot 1 also behaved in a mannerthat was very similar to that of the polar B[a]P adductsdetected in rat liver, but the two adducts were sepa-rated by HPLC. Fourth, Spot 1 was compared withCD1 mice exposed to 7H-benzo[c]fluorene (B[c]F).Spot 1 from some patients comigrated with a majoradduct induced by B[c]F. Finally, we found that thepresence of Spot 1 in human breast tissues was notrelated to smoking status but, rather, with CYP1A1MspI polymorphism. The CYP1A1 mutant carriershad a significantly higher frequency of this adductthan did the wild-type genotypes. Furthermore, individ-uals with Spot 1 had a significantly higher staining in-tensity for BPDE–PAH adducts in their tissue sections thanthose without it. These results demonstrate that this majorbulky DNA adduct detected in human breast tissues isrelated to PAH exposure. Environ. Mol. Mutagen.39:193–200, 2002. © 2002 Wiley-Liss, Inc.
Key words: DNA adduct; rechromatography analysis; Spot 1; polycyclic aromatic hydrocar-bons; comigration; chemical carcinogens
INTRODUCTION
The etiology of the majority of human breast cancers isunknown. Several types of chemical carcinogens have beensuggested to be involved in human breast cancer, for exam-ple, polycyclic aromatic hydrocarbons (PAHs), nitro-PAHs,aromatic amines, and heterocyclic aromatic amines [El-Bayoumy, 1992]. Many of these compounds are knownmammary carcinogens in laboratory animals and are presentin the ambient environment or in the human diet. However,epidemiological studies of breast cancer have not implicatedany environmental carcinogens except radiation [John et al.,1993]. Epidemiological studies have found inconclusiveresults on the role of cigarette smoking, organochlorine
compounds, as well as well-done meat intake in breastcancer. Experimental evidence, on the other hand, supports
Grant sponsor: National Institutes of Health; Grant number: CA70264;Grant sponsor: NIEHS; Grant number: P30 ES07784; Grant sponsor:University of Texas M.D. Anderson Breast Cancer Research Program.
*Correspondence to: Donghui Li, Department of Gastrointestinal MedicalOncology, University of Texas, M.D. Anderson Cancer Center, HoustonTX 77030. E-mail: [email protected]
Received 25 September 2001; provisionally accepted 31 October 2001; andin final form 30 November 2001
Published online 4 March 2002 in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/em.10063
Environmental and Molecular Mutagenesis 39:193–200 (2002)
© 2002 Wiley-Liss, Inc.
a role of environmental carcinogens in breast cancer. Forexample, human mammary lipids have been shown to con-tain genotoxic compounds and to induce DNA damage[Martin et al., 1996, 1997]. The spectrum of p53 genemutation in breast cancer suggests the involvement of geno-toxic compounds [Biggs et al., 1993]. The anatomic featuresof mammary gland also make it a susceptible target forchemical carcinogens. Lipophilic compounds can be storedin the fat pads [Obana et al., 1981], and mammary epithelialcells have a high capacity to activate carcinogens in situ[Carmichael et al., 1996].
Other investigators and we previously reported the detec-tion of aromatic DNA adducts in human breast tissues[Perera et al., 1995; Li et al., 1996]. However, none of theadducts detected by 32P-postlabeling has been chemicallyidentified. Therefore, the possible source of exposure andthe biological significance of these DNA adducts detected inhuman breast tissues remain unknown. Direct structuralidentification of DNA adducts detected in human tissues bymass spectrometry is often precluded because the amount ofDNA available is limited and the level of adduct is usuallyrelatively low. Characterization of the in vivo–detectedadducts, then, would rely on indirect methods such as chro-matographic comparisons with known adduct standards andimmunoreaction with antibodies recognizing specific DNAadducts.
In our previous study [Li et al., 1996], a bulky adduct(Spot 1) was detected in tissue DNA of 36 out of 87 (41%)breast cancer patients but in none of the 29 noncancercontrols. The level of this particular adduct was extremelyhigh in some patients (�1/106). In the attempt to character-ize this major adduct, the present study has compared Spot1 with adduct profiles generated in several in vitro and in
vivo experimental systems by cochromatography and re-chromatography analyses. Our data suggest that this adductis related to PAH exposure.
MATERIALS AND METHODS
Materials
All carcinogens were obtained from the National Cancer InstituteChemical Carcinogen Reference Standard Repository at the MidwestResearch Institute (Kansas City, MO). The benzo[a]pyrene (B[a]P)metabolites 1-hydroxy and 3-hydroxy 7,8-dihydrodiol of B[a]P wereobtained from Dr. R.G. Harvey at the University of Chicago. 1,N2-Ethenodeoxyguanosine was provided by Dr. F.-L. Chung from theAmerican Health Foundation. 4-Hydroxynonenal (4-HNE) was a gen-erous gift from Dr. Hermann Esterbauer at the Institute of Biochemis-try, University of Graz, Austria.
Nonmalignant breast tissue from breast cancer patients and normalbreast tissue from reduction mammoplasty were collected from Universityof Texas M.D. Anderson Cancer Center and a local clinic of plastic surgery(Houston, TX) during 1994 to 1996.
Cell Treatment With Carcinogens
The MCF-7 cell was selected as the test system because this cell line hasbeen used in many previous studies, demonstrating its high metaboliccapacity for the metabolism of PAH carcinogens and estrogens [Lau et al.,1995; Ralston et al., 1995; Einolf et al., 1996; Agarwal et al., 1997]. Cellswere grown to 50–80% confluence in DMEM with 10% fetal bovineserum. Cells were then treated with one of the following compounds:9-OH-B[a]P (2, 4, 8 �M), 11-OH-B[a]P (10 �M), 4-OH-B[a]P (10 �M),B[a]P-trans-4,5-dihydrodiol (10 �M), dibenzo[a,l]pyrene (10 �M), 6-ni-trochrysene (10 �M), benzo[c]phenanthrene (10 �M), dibenzo[a,h]anthra-cene (10 �M), 1-nitropyrene (2.5, 5, 10, and 20 mM), 2-amino-1-methyl-6-phenyl-imidazo[4,5-b]pyridine (PhIP, 40, 80, and 120 mM), and3-methylcholanthrene (1, 2, and 4 �M). Controls received the vehiclesolvent dimethylsulfoxide (DMSO) alone in the medium. After 7 to 24 hr
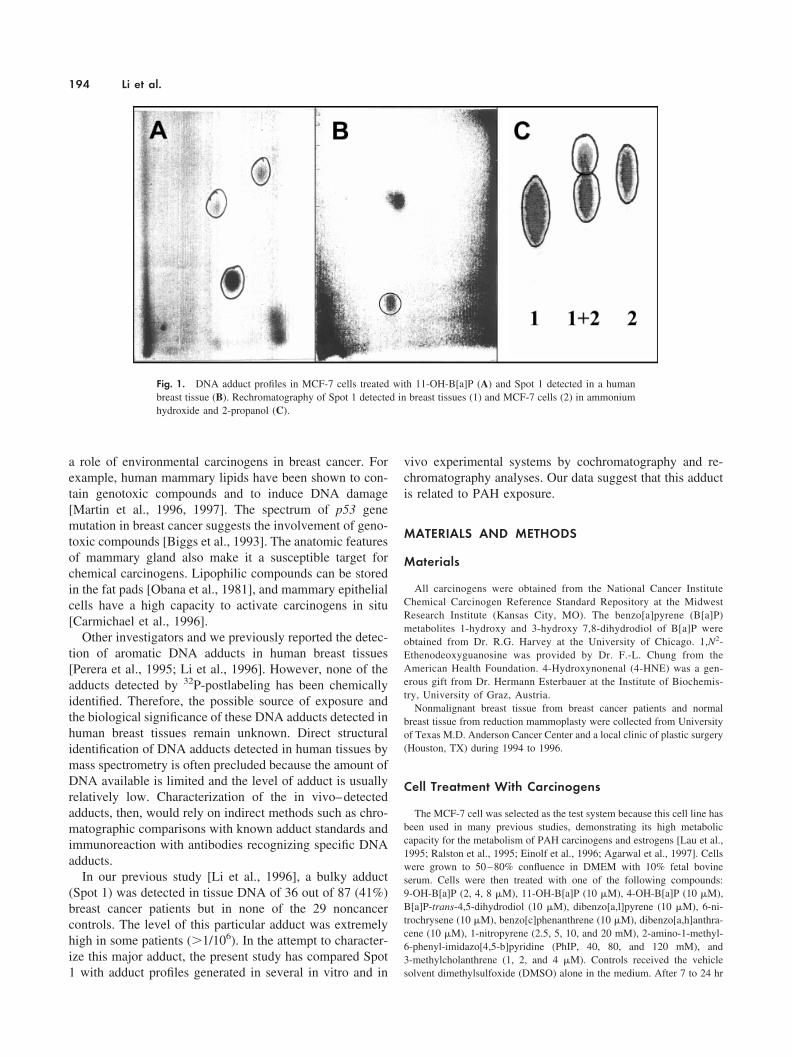
Fig. 1. DNA adduct profiles in MCF-7 cells treated with 11-OH-B[a]P (A) and Spot 1 detected in a humanbreast tissue (B). Rechromatography of Spot 1 detected in breast tissues (1) and MCF-7 cells (2) in ammoniumhydroxide and 2-propanol (C).
194 Li et al.
of exposure, cells were washed with PBS and then harvested by a brieftreatment with trypsin:EDTA. Cell pellets were stored at �80°C untilDNA isolation.
To test whether Spot 1 is derived through an endogenous mechanismrelated to estrogen metabolism and oxidative stress, cells were exposed to
4 �M 17�-estradiol (E2), 20 nM 4-OH-E2, and 10 nM 13-hydroperoxy-9,11-octadecadienoid acid.
In Vitro Reaction of DNA With Carcinogens
DNA (from lung tissues of 6-week-old untreated rats) was reacted withone of the following chemicals: BPDE (0.02 and 0.1 mM), 1-hydroxy and3-hydroxy-7,8-dihydrodiol of B[a]P (B[a]P triol, 0.1 mM) in the presenceof human mammary microsomes (1 mg protein), 4-HNE and oxidized oatoil [Li et al., 1995]. Microsomes were isolated from normal breast tissuesof women receiving reduction mammoplasty. The reaction of DNA withcarcinogens used either 1 mM NADPH or 0.15 mM cumene hydroperoxideas cofactors. The chemical carcinogens were first reacted with microsomesat 37°C for 10 min and then the incubation continued for another 30 minafter addition of 750 �g DNA. DNA was purified by solvent extraction andRNAse treatment. 3�-Monophosphates of normal nucleosides were alsoreacted with BPDE and the modified nucleotides were extracted withbutanol.
Animal Treatment
Female Sprague–Dawley rats (150 g) were orally treated with 20 mg/kgB[a]P once daily for 1 or 4 days. Controls received the vehicle solventDMSO alone. Animals were euthanized 24 hr after the last dosing. Mam-mary gland and liver tissues were removed and kept at �80°C until DNAisolation. Female CD-1 mice were fed basal gel diet containing 7H-benzo[c]fluorene (B[c]F; 60 mg/kg) ad libitum for 14 days [Koganti et al.,2000]. Mice were euthanized and lungs quickly removed and stored at�20°C before DNA isolation.
DNA Adduct Analysis
DNA was extracted by the phenol/chloroform procedure and DNAadducts were analyzed by the nuclease P1-enhanced version of the 32P-postlabeling assay [Reddy et al., 1986]. Chromatography conditions wereas previously described [Li et al., 1996]. Spots that migrated to a locationsimilar to that of Spot 1 on the two-dimensional maps were eluted with 6N ammonium hydroxide/isopropanol (1:1, v:v) for rechromatography anal-ysis. (Solvents used for rechromatography are given in the figure legends.)
The HPLC analysis used a liquid chromatography system equipped witha radioactivity detector. Separations were performed on a Zorbax phenyl-modified reverse-phase column [Baer-Dubowska et al., 1995].
DNA adduct detection in breast tissue sections by the immunohisto-chemistry method was as previously described [Santella, 1999].
RESULTS AND DISCUSSION
In an attempt to characterize the major adduct Spot 1detected in human breast tissues, we previously showed thatthis adduct was present, as a minor adduct, in B[a]P-treatedMCF-7 cells [Li et al., 1996]. However, it is not clearwhether this adduct is derived from DNA-binding B[a]Pmetabolites or whether it is an indirect DNA modificationthat does not contain the B[a]P moiety. It is known thatindirect DNA adducts with unknown chemical identitycould be generated during metabolism of carcinogens andestrogens [Liehr et al., 1986, Marnett et al., 1993]. Todistinguish whether Spot 1 is directly derived from B[a]P oris an endogenous DNA adduct formed as a consequence ofB[a]P exposure, we compared it with adduct profiles de-
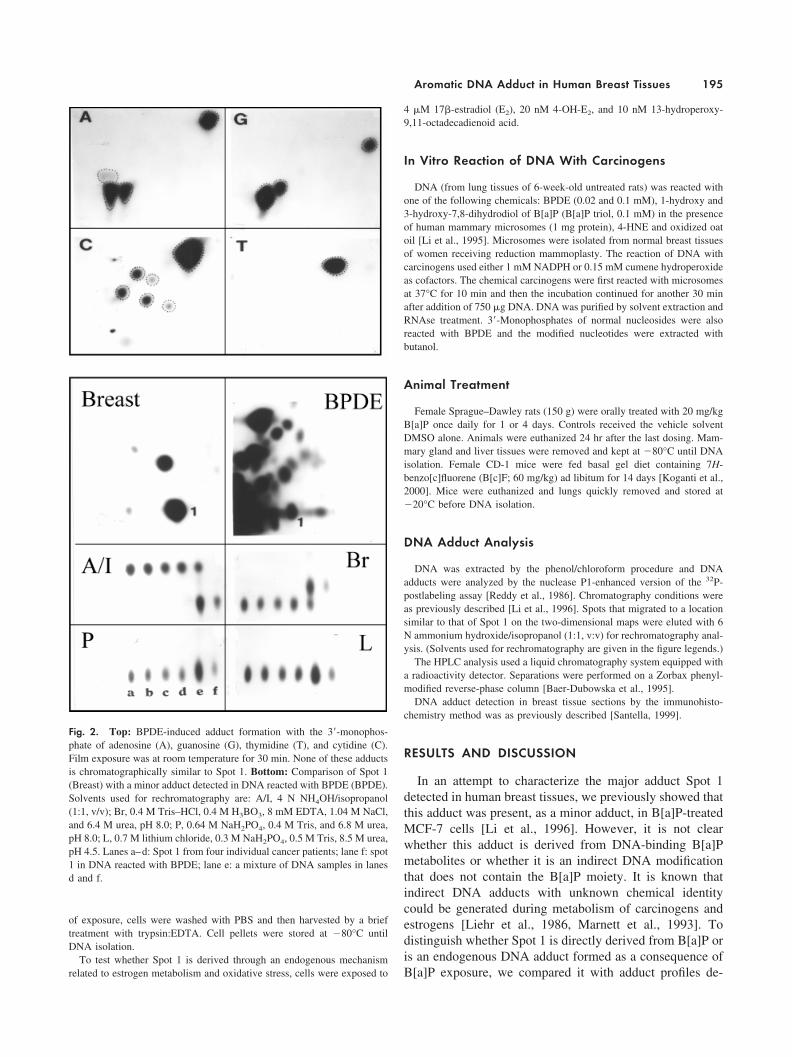
Fig. 2. Top: BPDE-induced adduct formation with the 3�-monophos-phate of adenosine (A), guanosine (G), thymidine (T), and cytidine (C).Film exposure was at room temperature for 30 min. None of these adductsis chromatographically similar to Spot 1. Bottom: Comparison of Spot 1(Breast) with a minor adduct detected in DNA reacted with BPDE (BPDE).Solvents used for rechromatography are: A/I, 4 N NH4OH/isopropanol(1:1, v/v); Br, 0.4 M Tris–HCl, 0.4 M H3BO3, 8 mM EDTA, 1.04 M NaCl,and 6.4 M urea, pH 8.0; P, 0.64 M NaH2PO4, 0.4 M Tris, and 6.8 M urea,pH 8.0; L, 0.7 M lithium chloride, 0.3 M NaH2PO4, 0.5 M Tris, 8.5 M urea,pH 4.5. Lanes a–d: Spot 1 from four individual cancer patients; lane f: spot1 in DNA reacted with BPDE; lane e: a mixture of DNA samples in lanesd and f.
Aromatic DNA Adduct in Human Breast Tissues 195
tected in MCF-7 cells exposed to several other carcinogensor B[a]P metabolites, that is, dibenzo[a,l]-pyrene, benzo[c]-phenanthrene, dibenzo[a,h]-anthracene, 1-nitropyrene, 6-ni-trochrysene, 2-amino-1-methyl-6-phenyl-imidazo[4,5-b]pyri-dine, 3-methylcholanthrene, 9-OH-B[a]P, 11-OH-B[a]P,4-OH-B[a]P, and B[a]P-trans-4,5-dihydrodiol. Most ofthese compounds induced DNA adduct formation but noneof these adducts is in a location similar to that of Spot 1under our chromatography conditions (films not shown).Nevertheless, 11-OH-B[a]P induced two DNA adducts, oneof which migrated in a manner similar to that of Spot 1 inboth two-dimensional anion exchange chromatography andpartition chromatography (Fig. 1). This observation sug-gests that Spot 1 was not derived from an endogenousmechanism after carcinogen exposure but was somehowrelated to B[a]P exposure.
The conclusion that Spot 1 was not derived from anendogenous mechanism is also supported by evidence froma series of experiments that compared Spot 1 with adductprofiles derived from estrogen exposure and lipid peroxida-tion. Spot 1 was not detected in MCF-7 cells treated with E2
and 4-OH-E2, an in vivo metabolite of E2 (film not shown).Spot 1 did not comigrate with the 1,N2-ethenodeox-yguanosine adduct or DNA adducts induced by 4-HNE,
13-hydroperoxy-9,11-octadecadienoid acid, or oxidized oatoil (film not shown).
Because Spot 1 was detected in B[a]P-treated MCF-7cells [Li et al., 1996] as a minor and more polar adduct thanthe BPDE–deoxyguanosine (dG) adduct (a major B[a]Padduct detected in animal tissues and cells exposed toB[a]P), we asked whether Spot 1 is derived from BPDE-modified bases other than dG. In vitro reaction of BPDEwith four individual 3�-phosphate mononucleosides inducedthe formation of specific adduct profiles (Fig. 2, top). How-ever, none of these adducts is located in the proximity ofSpot 1. Reaction of BPDE with DNA, on the other hand,induced the formation of a minor adduct in a locationsimilar to that of Spot 1 [Fig. 2, bottom, lane f (BPDE)].This in vitro adduct was barely separable from Spot 1during anion exchange rechromatography [Fig. 2, bottom,panels Br, P, L] but was clearly separated by partitionrechromatography [Fig. 2, bottom, A/I]. In fact, the in vitroBPDE-induced minor adduct was slightly more polar thanSpot 1. Next we examined several B[a]P derivatives, that is,1-hydroxy and 3-hydroxy B[a]P triols, which might givemore polar adducts than the parent compound [Baer-Dubowska et al., 1995]. In the presence of microsomesisolated from human breast tissues, reaction of DNA with
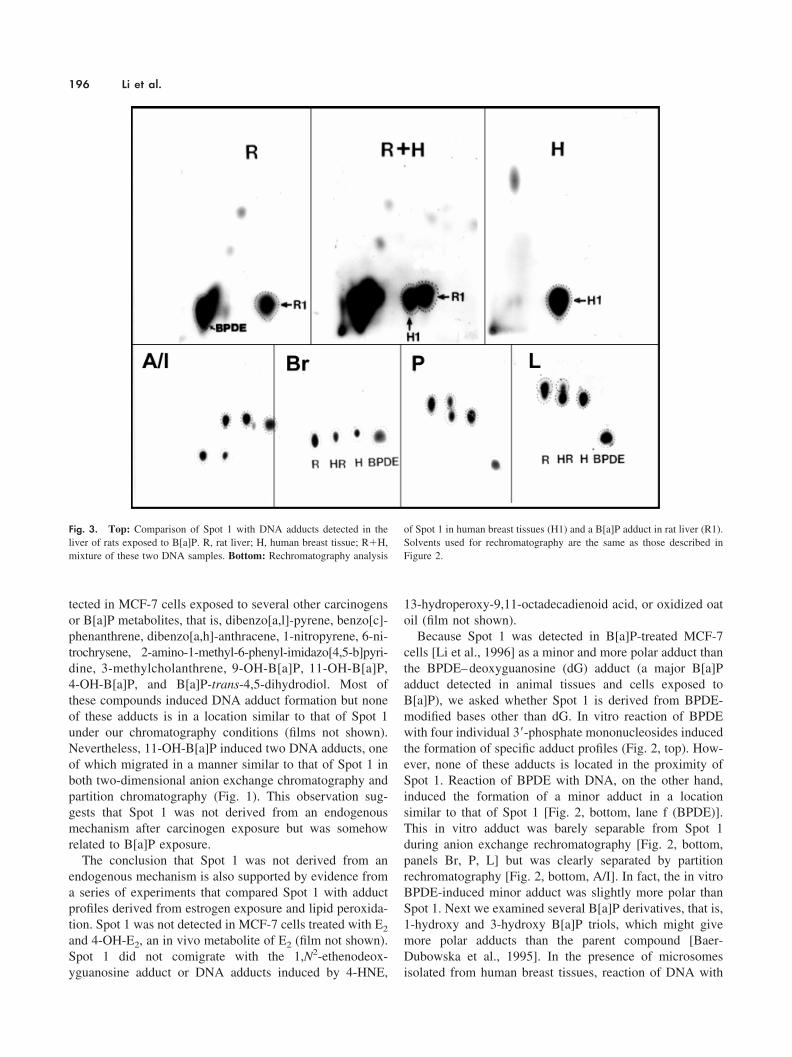
Fig. 3. Top: Comparison of Spot 1 with DNA adducts detected in theliver of rats exposed to B[a]P. R, rat liver; H, human breast tissue; R�H,mixture of these two DNA samples. Bottom: Rechromatography analysis
of Spot 1 in human breast tissues (H1) and a B[a]P adduct in rat liver (R1).Solvents used for rechromatography are the same as those described inFigure 2.
196 Li et al.
these two compounds resulted in formation of a number ofadducts that are highly lipophilic bulky adducts, given thatthe BPDE-induced major adducts and no polar adducts werefound to comigrate with Spot 1 (film not shown). Theseresults suggest that Spot 1 was not derived from BPDE-modified DNA bases other than dG and it was not derivedfrom 1-OH- and 3-OH-B[a]P triols. Because we could notdirectly generate this adduct in vitro, we examined the
possibility whether this adduct can be generated in an invivo system using an animal model.
Previous studies have shown that liver DNA from B[a]P-treated rats or rat mammary epithelial cells in vitro reactedwith B[a]P display some polar DNA adducts in addition tothe BPDE–dG adduct [Dock et al., 1986; Ross et al., 1990].To test whether Spot 1 comigrates with any of these morepolar B[a]P adducts in rats, Spot 1 was compared with
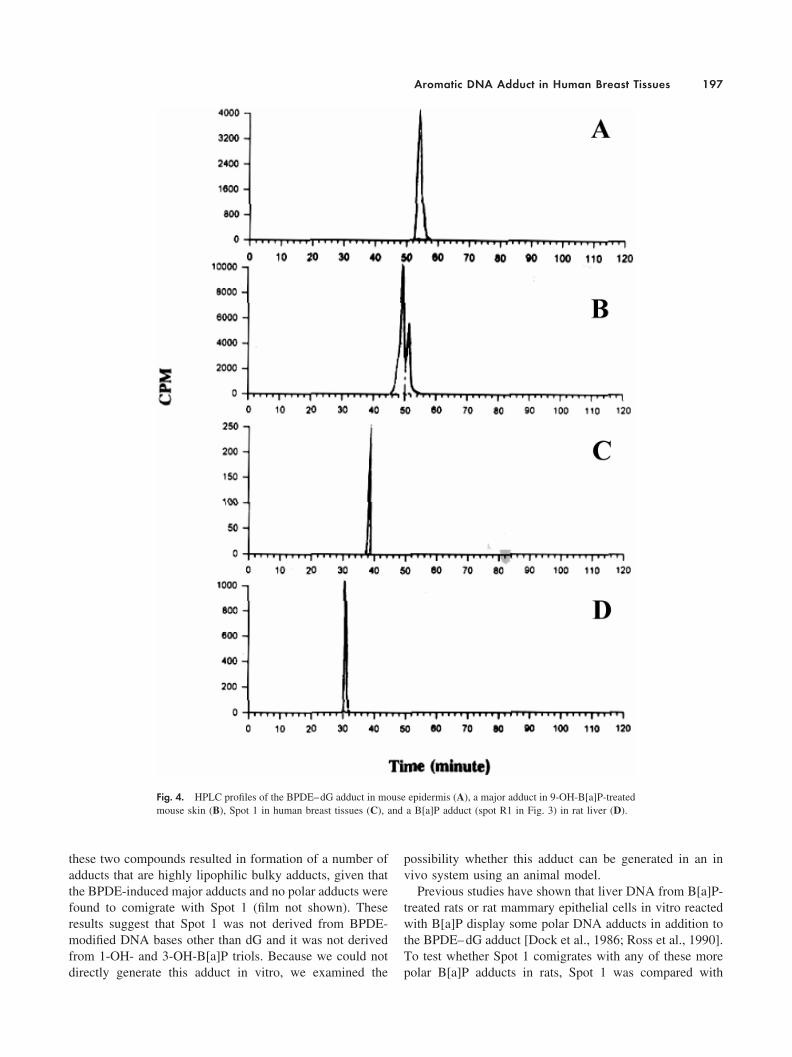
Fig. 4. HPLC profiles of the BPDE–dG adduct in mouse epidermis (A), a major adduct in 9-OH-B[a]P-treatedmouse skin (B), Spot 1 in human breast tissues (C), and a B[a]P adduct (spot R1 in Fig. 3) in rat liver (D).
Aromatic DNA Adduct in Human Breast Tissues 197
adduct profiles in B[a]P-treated rats. As reported previously[Ross et al., 1990], a fast moving B[a]P–DNA adduct (R1),in addition to the BPDE–dG adduct, was detected in mam-mary gland (data not shown) and liver from rats exposed toB[a]P (spot R1 in Fig. 3, panel R). This fast moving adduct(R1) migrated in a manner similar to that of Spot 1 under thesame two-dimensional chromatography conditions (Fig. 3,panels R and H). However, cochromatography (Fig. 3, panelR�H) and rechromatography analysis (Fig. 3, panels I, B,P, and L) demonstrated that adduct R1 is also more polar
than Spot 1. The two adducts were clearly separated bypartition chromatography (Fig. 3, panel I) and reverse-phaseHPLC (Fig. 4).
The results from comparisons of Spot 1 with DNA adductprofiles in MCF-7 cells exposed to B[a]P and 11-OH-B[a]P,in rats treated with B[a]P, and in DNA reacted with BPDEwere intriguing. Spot 1 was chromatographically similar tosome of the adducts detected in these experiments, althoughnone of them was a perfect match.
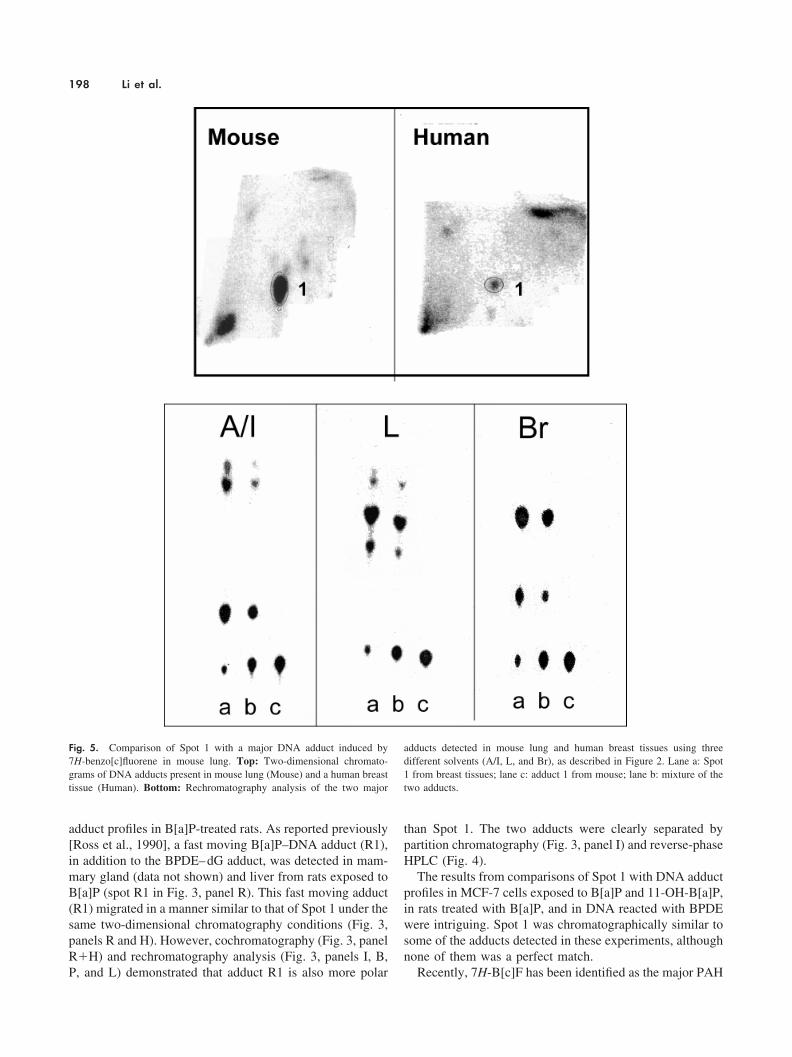
Recently, 7H-B[c]F has been identified as the major PAH
Fig. 5. Comparison of Spot 1 with a major DNA adduct induced by7H-benzo[c]fluorene in mouse lung. Top: Two-dimensional chromato-grams of DNA adducts present in mouse lung (Mouse) and a human breasttissue (Human). Bottom: Rechromatography analysis of the two major
adducts detected in mouse lung and human breast tissues using threedifferent solvents (A/I, L, and Br), as described in Figure 2. Lane a: Spot1 from breast tissues; lane c: adduct 1 from mouse; lane b: mixture of thetwo adducts.
198 Li et al.
compound present in coal tar to induce DNA adduct forma-tion in mouse lung [Koganti et al., 2000]. The major adductinduced by 7H-B[c]F behaved in a manner similar to that ofB[a]P adducts in multidimensional chromatography. With alimited amount of DNA samples on hand, we comparedSpot 1 (pooled DNA samples from three patients) with the7H-B[c]F–induced adduct in mouse lung. As shown inFigure 5, Spot 1 and the major 7H-B[c]F adduct migrated tothe same location on the TLC map after two-dimensionalchromatography. However, Spot 1 was separated into threefractions during rechromatography and only one of thefractions comigrated with the B[c]F adduct in all the sol-vents used. It is possible that the three fractions of Spot 1detected during rechromatography actually represent differ-ent adducts in each of the three patients. Nevertheless, thematch between the B[c]F adduct and one fraction of Spot 1is still an important observation. It suggests that exposure tocoal tar may induce DNA adduct formation in human breasttissues.
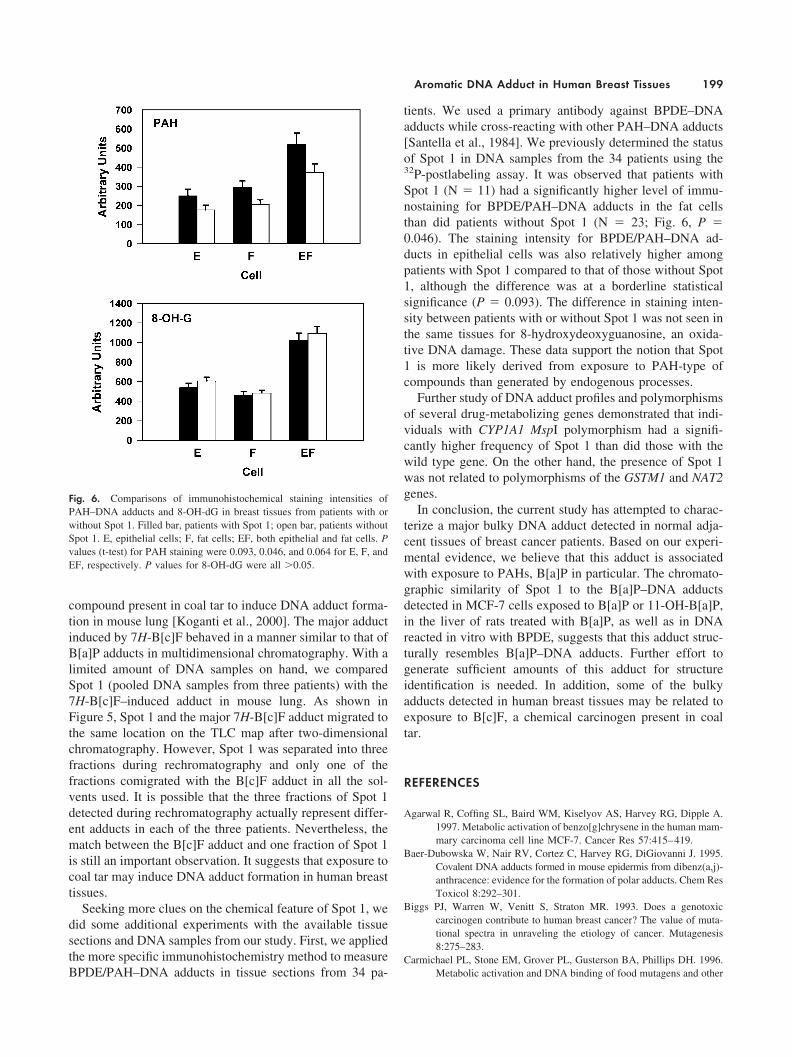
Seeking more clues on the chemical feature of Spot 1, wedid some additional experiments with the available tissuesections and DNA samples from our study. First, we appliedthe more specific immunohistochemistry method to measureBPDE/PAH–DNA adducts in tissue sections from 34 pa-
tients. We used a primary antibody against BPDE–DNAadducts while cross-reacting with other PAH–DNA adducts[Santella et al., 1984]. We previously determined the statusof Spot 1 in DNA samples from the 34 patients using the32P-postlabeling assay. It was observed that patients withSpot 1 (N � 11) had a significantly higher level of immu-nostaining for BPDE/PAH–DNA adducts in the fat cellsthan did patients without Spot 1 (N � 23; Fig. 6, P �0.046). The staining intensity for BPDE/PAH–DNA ad-ducts in epithelial cells was also relatively higher amongpatients with Spot 1 compared to that of those without Spot1, although the difference was at a borderline statisticalsignificance (P � 0.093). The difference in staining inten-sity between patients with or without Spot 1 was not seen inthe same tissues for 8-hydroxydeoxyguanosine, an oxida-tive DNA damage. These data support the notion that Spot1 is more likely derived from exposure to PAH-type ofcompounds than generated by endogenous processes.
Further study of DNA adduct profiles and polymorphismsof several drug-metabolizing genes demonstrated that indi-viduals with CYP1A1 MspI polymorphism had a signifi-cantly higher frequency of Spot 1 than did those with thewild type gene. On the other hand, the presence of Spot 1was not related to polymorphisms of the GSTM1 and NAT2genes.
In conclusion, the current study has attempted to charac-terize a major bulky DNA adduct detected in normal adja-cent tissues of breast cancer patients. Based on our experi-mental evidence, we believe that this adduct is associatedwith exposure to PAHs, B[a]P in particular. The chromato-graphic similarity of Spot 1 to the B[a]P–DNA adductsdetected in MCF-7 cells exposed to B[a]P or 11-OH-B[a]P,in the liver of rats treated with B[a]P, as well as in DNAreacted in vitro with BPDE, suggests that this adduct struc-turally resembles B[a]P–DNA adducts. Further effort togenerate sufficient amounts of this adduct for structureidentification is needed. In addition, some of the bulkyadducts detected in human breast tissues may be related toexposure to B[c]F, a chemical carcinogen present in coaltar.
REFERENCES
Agarwal R, Coffing SL, Baird WM, Kiselyov AS, Harvey RG, Dipple A.1997. Metabolic activation of benzo[g]chrysene in the human mam-mary carcinoma cell line MCF-7. Cancer Res 57:415–419.
Baer-Dubowska W, Nair RV, Cortez C, Harvey RG, DiGiovanni J. 1995.Covalent DNA adducts formed in mouse epidermis from dibenz(a,j)-anthracence: evidence for the formation of polar adducts. Chem ResToxicol 8:292–301.
Biggs PJ, Warren W, Venitt S, Straton MR. 1993. Does a genotoxiccarcinogen contribute to human breast cancer? The value of muta-tional spectra in unraveling the etiology of cancer. Mutagenesis8:275–283.
Carmichael PL, Stone EM, Grover PL, Gusterson BA, Phillips DH. 1996.Metabolic activation and DNA binding of food mutagens and other
Fig. 6. Comparisons of immunohistochemical staining intensities ofPAH–DNA adducts and 8-OH-dG in breast tissues from patients with orwithout Spot 1. Filled bar, patients with Spot 1; open bar, patients withoutSpot 1. E, epithelial cells; F, fat cells; EF, both epithelial and fat cells. Pvalues (t-test) for PAH staining were 0.093, 0.046, and 0.064 for E, F, andEF, respectively. P values for 8-OH-dG were all �0.05.
Aromatic DNA Adduct in Human Breast Tissues 199

environmental carcinogens in human mammary epithelial cells.Carcinogenesis 17:1769–1972.
Dock L, Waern F, Martinez M, Grover PL, Jernstrom B. 1986. Studies onthe further activation of benzo[a]pyrene diol epoxide by rat livermicrosomes and nuclei. Chem–Biol Interact 58:301–318.
Einolf HJ, Amin S, Yagi H, Jerina DM, Baird WM. 1996. Benzo[c]phenan-threne is activated to DNA-binding diol epoxides in the humanmammary carcinoma cell line MCF-7 but only limited activationoccurs in mouse skin. Carcinogenesis 17:2237–2244.
El-Bayoumy K. 1992. Environmental carcinogens that may be involved inhuman breast cancer etiology. Chem Res Toxicol 5:585–590.
John EM, Kelsey JL. 1993. Radiation and other environmental exposuresand breast cancer. Epidemiol Rev 15:157–162.
Koganti A, Singh R, Rozett K, Modi N, Goldstein LS, Roy TA, Zhang FJ,Harvey RG, Weyand EH. 2000. 7H-Benzo[c]fluorene: a majorDNA adduct-forming component of coal tar. Carcinogenesis 21:1601–1609.
Lau HH, Coffing SL, Lee H, Harvey RG, Baird WM. 1995. Stereoselec-tivity of activation of 7,12-dimethylbenz[a]anthracene-3,4-dihydro-diol to the anti-diol epoxide metabolite in a human mammarycarcinoma MCF-7 cell-mediated V79 cell mutation assay. ChemRes Toxicol 8:970–978.
Li D, Wang MY, Liehr JG, Randerath K. 1995. DNA adducts induced bylipids and lipid peroxidation products: possible relationships toI-compounds. Mutat Res 344:117–126.
Li D, Wang MY, Dhingra K, Hittelman WN. 1996. Aromatic DNA adductsin adjacent tissues of breast cancer patients: clues to breast canceretiology. Cancer Res 56:287–293.
Liehr JG, Avitts TA, Randerath E, Randerath K. 1986. Estrogen-inducedendogenous DNA adduction: possible mechanism of hormonalcancer. Proc Natl Acad Sci USA 83:5301–5305.
Marnett LJ, Burcham PC. 1993. Endogenous DNA adducts: potential andparadox. Chem Res Toxicol 6:771–785.
Martin FL, Carmichael PL, Crofton-Sleigh C, Venitt S, Phillips DH,Grover PL. 1996. Genotoxicity of human mammary lipid. CancerRes 56:5342–5346.
Martin FL, Venitt S, Carmichael PL, Crofton-Sleigh C, Stone EM, ColeKJ, Gusterson BA, Grover PL, Phillips DH. 1997. DNA damage inbreast epithelial cells: detection by the single-cell gel (comet) assayand induction by human mammary lipid extracts. Carcinogenesis18:2299–2305.
Moore CJ, Pruess-Schwartz D, Mauthe RJ, Gould MN, Baird WM.1987. Interspecies differences in the major DNA adducts formedfrom benzo[a]pyrene but not 7,12-dimethylbenzo[a]anthracencein rat and human mammary cell cultures. Cancer Res 47:4402–4406.
Obana H, Hori S, Kashmoto L, Kunita N. 1981. Polycyclic aromatichydrocarbons in human fat and liver. Bull Environ Contam Toxicol27:23–27.
Perera FP, Estabrook A, Hewer A, Channing K, Rundle A, Mooney L,Wayatt R, Phillips DH. 1995. Carcinogen-DNA adducts in humanbreast tissue. Cancer Epidemiol Biomarkers Prev 4:233–238.
Ralston SL, Seidel A, Luch A, Platt KL, Baird WM. 1995. Stereoselectiveactivation of dibenzo[a,l]pyrene to (�)-anti (11R,12S,13S,14R)-and (�)-syn(11S,12R,13S,14R)-11,12-diol-13,14-epoxides whichbind extensively to deoxyadenosine residues of DNA in the humanmammary carcinoma cell line MCF-7. Carcinogenesis 16:2899–2907.
Reddy MV, Randerath K. 1986. Nuclease P1-mediated enhancement ofsensitivity of 32P-postlabeling test for structurally diverse DNAadducts. Carcinogenesis 7:1543–1551.
Ross J, Nelson G, Kligerman A, Erexson G, Bryant M, Early K, Gupta R,Nesnow S. 1990. Formation and persistence of novel benzo-[a]pyrene adducts in rat lung, liver, and peripheral blood lympho-cyte DNA. Cancer Res 49:5088–5094.
Santella RM. 1999. Immunological methods for detection of carcinogen-DNA damage in humans. Cancer Epidemiol Biomarkers Prev8:733–739.
Santella RM, Lin CD, Cleveland WL, Weinstein IB. 1984. Monoclonalantibodies to DNA modified by a benzo[a]pyrene diol epoxide.Carcinogenesis 5:373–377.
200 Li et al.