developmental cell article - perelman school of medicine filedevelopmental cell article local crh...
TRANSCRIPT

Developmental Cell
Article
Local CRH Signaling Promotes Synaptogenesisand Circuit Integration of Adult-Born NeuronsIsabella Garcia,1,2 Kathleen B. Quast,3 Longwen Huang,4 Alexander M. Herman,1 Jennifer Selever,3 Jan M. Deussing,5
Nicholas J. Justice,6 and Benjamin R. Arenkiel1,3,4,7,*1Program in Developmental Biology, Baylor College of Medicine, Houston, TX 77030, USA2Medical Scientist Training Program, Baylor College of Medicine, Houston, TX 77030, USA3Department of Molecular and Human Genetics, Baylor College of Medicine, Houston, TX 77030, USA4Department of Neuroscience, Baylor College of Medicine, Houston, TX 77030, USA5Max Planck Institute of Psychiatry, 80804 Munich, Germany6Institute of Molecular Medicine, University of Texas Health Science Center at Houston, Houston, TX 77030, USA7Jan and Dan Duncan Neurological Research Institute at Texas Children’s Hospital, Houston, TX 77030, USA
*Correspondence: [email protected]
http://dx.doi.org/10.1016/j.devcel.2014.07.001
SUMMARY
Neural activity either enhances or impairs de novosynaptogenesis and circuit integration of neurons,but how this activity is mechanistically relayed inthe adult brain is largely unknown. Neuropeptide-ex-pressing interneurons arewidespread throughout thebrain and are key candidates for conveying neuralactivity downstream via neuromodulatory pathwaysthat are distinct from classical neurotransmission.With the goal of identifying signaling mechanismsthat underlie neuronal circuit integration in the adultbrain, we have virally traced local corticotropin-releasing hormone (CRH)-expressing inhibitory inter-neurons with extensive presynaptic inputs onto newneurons that are continuously integrated into theadult rodent olfactory bulb. Local CRH signalingonto adult-born neurons promotes and/or stabilizeschemical synapses in the olfactory bulb, revealing aneuromodulatory mechanism for continued circuitplasticity, synapse formation, and integration of newneurons in the adult brain.
INTRODUCTION
Synaptogenesis and circuit integration of neurons in the brain is
governed by diverse repertoires of synaptic and extrasynaptic
inputs. Excitatory input by principal neurons has profound
effects on sculpting and pruning synaptic connectivity (Favero
and Castro-Alamancos, 2013; Le Roux et al., 2013). However,
recent evidence suggests that interneurons also play significant
roles in modulating synapse formation (De Marco Garcıa et al.,
2011; Hensch et al., 1998; Le Magueresse and Monyer, 2013).
Inhibitory interneurons are highly heterogeneous and, depending
on the brain region, can vastly outnumber principal neurons
(Chen and Greer, 2004; Isaacson and Strowbridge, 1998; Lledo
et al., 2008). Neurochemical classification schemes have shown
that interneurons not only express GABA and calcium-binding
Developmen
proteins such as parvalbumin (PV), calretinin (CR), and calbindin
(Barinka et al., 2012; Kosaka and Kosaka, 2008; Rudy et al.,
2011), but also a cast of neuromodulatory peptides, including so-
matostatin (SST), cholecystokinin, oxytocin, and corticotropin-
releasing hormone (CRH) (Huang et al., 2013; Le Magueresse
and Monyer, 2013; Ma et al., 2006; Rudy et al., 2011; Xu et al.,
2013). Neuropeptidergic interneurons are promising candidates
for modulating changes in local synapse and circuit structure,
and are pervasive throughout the rodent olfactory bulb (OB).
Endowed with the feature of ongoing neurogenesis (Alvarez-
Buylla and Temple, 1998), the mouse olfactory system offers
an excellent in vivomodel to investigate mechanisms that under-
lie synaptogenesis, circuit plasticity, and the integration of new
neurons into existing networks (Abrous et al., 2005; Ming and
Song, 2005). Adult-born neurons are continuously generated in
the subventricular zone (SVZ), migrate via the rostral migratory
stream (RMS), and populate the OB where the vast majority
become inhibitory granule cells that form connections with OB
principal mitral and tufted cells (Mori et al., 1983; Price and Po-
well, 1970a, 1970b; Carleton et al., 2003; Shepherd and Greer,
2004). This interaction influences olfactory behaviors and odor-
related memories (Abraham et al., 2010; Breton-Provencher
et al., 2009; Mouret et al., 2009; Rochefort et al., 2002).
Studies have found that the survival and integration of adult-
born granule cells is activity-dependent during a developmental
critical period between 2 and 4 weeks after their birth (Kelsch
et al., 2009; Yamaguchi and Mori, 2005), when they receive in-
puts from local interneuron subtypes, including deep and super-
ficial short axon cells and Blanes cells (Arenkiel et al., 2011; Eyre
et al., 2008; Pressler and Strowbridge, 2006), as well as centrif-
ugal fibers from deeper regions of the brain (Arenkiel et al., 2011;
Balu et al., 2007; Panzanelli et al., 2009; Whitman and Greer,
2007). Maturing granule cells extend their dendrites into the
external plexiform layer (EPL), where they connect with principal
mitral cells. Interestingly, the EPL also harbors amore dispersed,
and heterogeneous population of neuropeptidergic interneurons
(Kosaka and Kosaka, 2008; Lepousez et al., 2010a, 2010b) that
also form reciprocal synaptic connectivity with mitral cells
(Huang et al., 2013; Kato et al., 2013; Miyamichi et al., 2013). Un-
like granule cells, EPL interneurons are generated in the early
postnatal period and remain stable throughout life (Batista-Brito
tal Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc. 645

(legend on next page)
Developmental Cell
Neuropeptide Signaling and Circuit Integration
646 Developmental Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc.

Developmental Cell
Neuropeptide Signaling and Circuit Integration
et al., 2008), but their potential neuromodulatory role in shaping
the integration of adult-born neurons is unknown.
We have previously shown that odor enrichment increases the
number of inputs onto adult-born neurons in the OB (Arenkiel
et al., 2011), causing enhanced cell survival and integration.
However, the precise signaling mechanisms between these
inputs and granule cells remain in question. In this study, we
have mapped local neuropeptidergic EPL interneurons with
anatomical and functional connectivity onto granule cells during
peak periods of synaptogenesis and circuit integration. Using
loss- and gain-of-function analyses, conditional viral-genetic
technologies, optical imaging, electrophysiological recordings,
and optogenetic stimulation, we have uncovered a neuro-
peptidergic signaling mechanism between local CRH+ EPL
interneurons and adult-born granule cells that plays an important
role in synapse formation, circuit plasticity, and the integration of
new neurons into the existing networks, revealing a dual func-
tional role for neuropeptidergic inhibitory interneurons in the
mouse OB.
RESULTS
Adult-Born Neurons Receive Inputs from Local EPLInterneuronsTo reveal the identities of the presynaptic inputs made onto
adult-born granule cells, we performed targeted monosynaptic
tracing using genetically engineered rabies virus (RV), SADG-
EGFP RV (Arenkiel et al., 2011; Wickersham et al., 2007a,
2007b). RV is a neurotropic virus that travels retrogradely be-
tween connected neurons. Endowing it with the avian coat pro-
tein EnvA provides selectivity of infection to source cells that ex-
press the TVA receptor, and replacing the glycoprotein G with
an EGFP reporter in the viral genome provides a fluorescently
labeled map of connectivity. Using a conditional knockin mouse
(ROSALSL-Rabies G-IRES-TVA, referred to herein asROSARITVA/RITVA)
that expresses the elements for conditional monosynaptic
tracing (rabies-G and TVA) (Takatoh et al., 2013), we selectively
targeted adult-born neurons in the OB for transsynaptic tracing.
ROSARITVA/RITVA mice were stereotaxically injected with a lenti-
virus that expressed Cre recombinase and a tdTomato reporter
into the RMS (Figure 1A). TdTomato marked adult-born neurons
red, and Cre recombinase allowed for conditional expression of
TVA and G. 28 days postlentiviral injection, we introduced RV
into the core of the OB to target newly integrated granule cells
for connectivity mapping. One week post-RV infection, the tar-
geted OB showed high-level EGFP expression (Figure 1B)
compared to negative controls in which no Cre was present (Fig-
ure S1A available online), suggesting that (1) adult-born neurons
Figure 1. Adult-Born Granule Cells Receive Presynaptic Inputs from E
(A) Genetic strategy for targeting adult-born neurons for transsynaptic tracing.
(B) Dorsal view of mouse brain showing a labeled OB 7 days post infection with
(C) Cross-section of (B). RMS, rostral migratory stream; GCL, granule cell layer; EP
(D) High-magnification view of (C). tdTomato+/EGFP+ cells (arrows) are newborn n
EPL presynaptic inputs; asterisks mark mitral cell inputs). MCL, mitral cell layer.
15 mm.
(E and F) EPL presynaptic inputs are PV+ (E, arrows; scale bar represents10
represents10 mm).
See also Figure S1.
Developmen
were successfully targeted for tracing, and (2) that retrograde
jumping of RV had occurred. Tissue sections revealed strong
expression of tdTomato in adult-born ‘‘source’’ cells, and high-
efficiency RV labeling in presynaptic inputs (Figures 1C and
1D). As positive controls, we identified presynaptic EGFP la-
beling in mitral cells (Figure 1D), validating RV jumping to known
synaptic partners. Interestingly, we found that adult-born
granule cells also received extensive inputs from both deep
and superficially located interneurons, with highest enrichment
from cells located in the EPL (Figure 1D).
Local CRH+ EPL Interneurons Make Connections ontoAdult-Born Granule CellsImmunohistochemical characterization of the presynaptic EGFP+
EPL cells revealed that granule cell inputs constituted a subset of
nondopaminergic interneurons (GFAP, 0%; tyrosine hydroxylase,
0%; CR+, 92% ± 5%; Figures S1B–S1D), with strong colabeling
of PV (95% ± 3%; Figure 1E), and SST (45% ± 5%; Figure S1E).
Highest neuropeptide expression was observed with CRH
(68% ± 4%; Figure 1F), suggesting that adult-born neurons
received direct input from resident CRH+ interneurons in the
OB. Although immunohistochemistry identified enrichment of
CRH protein in EPL interneurons, we also detected substantial
extracellular CRH in the EPL (Figure 1F), suggesting that CRH is
locally secreted. To more precisely identify CRH+ interneurons
as bona fide inputs, we used genetic labeling techniques.
To this end, we crossed CRH-Cre mice to conditional
tdTomato reporter mice (CRH-Cre+/�; ROSALSL-tdTom+/�), andobserved strong tdTomato signal in the paraventricular nucleus
of the hypothalamus, a known hub for CRH synthesis and secre-
tion (Figure S2), and high levels of expression in EPL interneurons
of theOB (Figure 2A),whichwepreviously characterized asmulti-
polar and anaxonic fast-spiking PV+ interneurons (Huang et al.,
2013). To determine if adult-born granule cells received inputs
from CRH+ EPL interneurons, we performed transsynaptic
tracing in CRH-Cre+/�; ROSALSL-tdTom+/� mice and electropo-
rated the avian TVA receptor and rabies-G glycoprotein into the
SVZ of CRH-Cre+/�; ROSALSL-tdTom+/� pups (Figure 2B), tran-
siently targeting neural progenitors that give rise to adult-born
neurons. 28 days later, mice were injected with RV into the core
of the OB, and killed 7 days later. OB slices revealed strong
EGFP expression, with high efficiency infection of targeted
granule source cells and presynaptic inputs (Figure 2C). Both
source cells and all inputs were labeled EGFP+, whereas CRH+
inputs were EGFP+/tdTomato+. Through this differential label-
ing, we verified that new granule cells indeed received extensive
input from local CRH+ EPL interneurons with 86% (±5% SEM) of
EPL presynaptic inputs expressing CRH via lineage tracing.
xternal Plexiform Layer Interneurons
SADG-EGFP RV. Scale bar represents 500 mm.
L, external plexiform layer; GL, glomerular layer. Scale bar represents 300 mm.
euron source cells. EGFP+ cells are presynaptic inputs (open arrowheadsmark
Scale bar represents 80 mm. Inset shows a source cell. Scale bar represents
mm) and CRH+ (F, arrows; arrowheads mark extracellular CRH; scale bar
tal Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc. 647

Figure 2. Local CRH+ EPL Interneurons
Make Connections onto Adult-Born Granule
Cells
(A) Genetic lineage of CRH+ neurons in the OB of
CRH-Cre+/�; ROSALSL-tdTom+/� mice. GL, glomer-
ular layer; EPL, external plexiform layer; MCL,
mitral cell layer; GCL, granule cell layer; scale bars
represent 500, 150, and 25 mm).
(B) Experimental scheme to identify presynaptic
inputs in CRH lineage-traced mice.
(C) SADDG-EGFP RV transsynaptic tracing in
CRH-Cre+/�; ROSALSL-tdTom+/� mice (arrows
identify newborn neuron source cells; open
arrowheads mark CRH+ presynaptic EPL in-
terneurons; scale bars represent 150 and 80 mm).
See also Figure S2.
Developmental Cell
Neuropeptide Signaling and Circuit Integration
Adult-Born Granule Cells Dynamically Express CRHR1CRH is best known as a hypothalamic regulatory hormone
that mediates systemic stress responses (Vale et al., 1981;
Vale et al., 1983). In addition, CRH has been implicated both
as a neurotransmitter and neuromodulator in the hippocam-
pus, amygdala, and cerebellum (Maras and Baram, 2012; Roo-
zendaal et al., 2008; Schmolesky et al., 2007). CRH can bind to
two G protein coupled receptors, CRHR1 and CRHR2 (Perrin
et al., 1993, 1995; Perrin and Vale, 1999), but in the brain
shows higher affinity to CRHR1, which mediates many of its
physiological effects (Bale and Vale, 2004). When bound to
CRHR1, Gs-coupled signaling is activated (Berger et al.,
2006; Blank et al., 2003; Perrin et al., 1993; Thiel and Cibelli,
1999).
Having identified that CRH+ EPL interneurons provide inputs
onto granule cells, we next sought to investigate the expression
of CRH receptors in theOB. RT-PCR revealed that bothCRH and
CRHRmRNAswere present in the bulb, and that CRHR1was ex-
pressed at much higher levels than CRHR2 (Figure 3A). Because
available antibodies for CRHR are not useful for detecting
endogenous CRHR1 (Refojo et al., 2011) and the expression
648 Developmental Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc.
pattern is not conclusive in the OB (Fig-
ure S3A), we used genetic strategies
to determine the CRHR1 cell type-spe-
cific expression pattern. Consistent with
mRNA transcript detection, tissue sec-
tions from CRHR1-EGFP BAC transgenic
mice, whose expression pattern was pre-
viously validated to match endogenous
CRHR1 (Justice et al., 2008), revealed
high levels of spatially restricted CRHR1
in granule cells (Figure 3B). Finally, to
determine the precise spatial localiza-
tion of CRH and CRHR1 neurons within
the OB,CRHR1-EGFPmicewere crossed
to CRH-Cre+/��; ROSALSL-tdTom+/� mice
to generate CRHR1-EGFP+/�; CRH-
Cre+/�; ROSALSL-tdTom+/� double reporter
mice. OB sections showed a cell type-
specific juxtaposition between EGFP-
labeled CRHR1+ granule cells, and
tdTomato-labeled CRH+ EPL interneu-
rons (Figure 3C). Whereas CRHR1+ granule cell bodies were
located throughout the GCL with superficial enrichment, CRH+
EPL interneurons almost exclusively resided in the EPL, were
absent from the GCL (Figure S3B), and directly juxtaposed
CRHR1+ dendrites. These data support the idea that CRH ligand
is locally released by EPL interneurons and directly acts on
CRHR1+ granule cell dendrites in the EPL.
EGFP expression was nearly absent from the RMS and
became gradually enriched in the outer GCL, with strong enrich-
ment in superficial regions (Figures 3B and 3C). These data sug-
gested that CRHR1 might exhibit a dynamic expression pattern
with granule cell maturation, and that CRHR1 might be ex-
pressed both in early postnatal-born granule cells which pre-
dominantly localize to the superficial GCL (Lemasson et al.,
2005), as well as adult-born granule cells. To test if CRHR1 is
dynamically regulated during periods of newborn granule cell
synaptogenesis and to better determine its spatiotemporal
expression profile, CRHR1-EGFP mice were pulsed with the
proliferation marker EdU, and killed between 7 and 60 days,
spanning early, intermediate, and late phases of synaptogenesis
and circuit integration (Figure 3D; Carleton et al., 2003). We

Figure 3. Adult-Born Granule Cells Dynami-
cally Express CRHR1
(A) Semiquantitative RT-PCR for CRH and CRHR1/
2 of whole OB RNA.
(B) OB cross-section of CRHR1-EGFP BAC
transgenic mice. RMS, rostral migratory stream;
GCL,granule cell layer; EPL, external plexiform
layer; GL, glomerular layer; scale bar represents
200 mm.
(C) Reporter expression of CRHR1-EGFP; CRH-
Cre+/�; ROSALSL-tdTom+/� mice (arrows point to
CRHR1+ granule cells; open arrowheads mark
CRH+ EPL interneurons; scale bars represent 60
and 20 mm).
(D) Experimental scheme to determine the devel-
opmental expression profile of CRHR1 expression
in granule cells.
(E) CRHR1 expression in newborn neurons 28 days
post EdU injection (scale bar represents 60 mm).
(F) Quantification of CRHR1-expression in granule
cells (data points represent averages ± SEM, n = 3
animals per time point).
(G) CRHR1::EGFP expression in adult-born
granule cells (scale bars represent 100 and 20 mm).
See also Figure S3.
Developmental Cell
Neuropeptide Signaling and Circuit Integration
Developmental Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc. 649

(legend on next page)
Developmental Cell
Neuropeptide Signaling and Circuit Integration
650 Developmental Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc.

Developmental Cell
Neuropeptide Signaling and Circuit Integration
found that the ratio of EdU-labeled granule cells that expressed
CRHR1 was very low at 7 days of age (5.8% ± 3.9%; Figure 3F),
and substantially increased between 14 (34.5% ± 4.1%), 21
(59.2% ± 4.2%), and 28 days of age (81.3% ± 2.2%). This value
slightly increased further at 40 days post-EdU pulsing (87.8% ±
4.5%), and the number of colabeled neurons plateaued at
60 days (87.5% ± 3.2%). Intriguingly, dynamic enrichment of
CRHR1 coincided with critical periods of activity-dependent
survival, synaptogenesis, and circuit integration between 14
and 28 days of granule cell age (Kelsch et al., 2009; Mouret
et al., 2008; Yamaguchi and Mori, 2005).
Finally, to determine the subcellular localization of CRHR1, we
expressed a CRHR1::EGFP fusion construct with a tdTomato
cell fill in new granule cells. We observed CRHR1::EGFP in den-
drites, with enriched subcellular localization in a subset of den-
dritic spines in the EPL (Figure 3G). Interestingly, CRHR1::EGFP
was also present in extrasynaptic dendritic regions (Figures
S3C–S3E), suggesting that CRH-mediated local neuropeptide
signaling might not occur exclusively at synapses, but also via
extrasynaptic mechanisms not restricted to dendritic spine
structures.
Together, these data support the idea that CRH is synthesized
locally by EPL interneurons and can signal to granule cells via
time-dependent expression of CRHR1, suggesting a possible
role for secreted CRH in the long-term survival and circuit inte-
gration of adult-born neurons.
CRH Signaling Is Required for Normal Levels of Adult-Born Granule Cell SurvivalTo determine how CRH signaling affects adult-born neurons,
mice lacking CRH or its receptor (CRH�/� or CRHR1�/�) were
pulsed with bromodeoxyuridine (BrdU) and killed either 24 hr
later to assay proliferation in the SVZ (BrdU+/Ki67+ cells), or
30 days later to assay survival in the GCL (BrdU+ cells; Fig-
ure 4A), focusing on deeper regions where adult-born granule
cells reside (Lemasson et al., 2005). Compared to wild-type lit-
termates, CRH�/� mutants showed increased SVZ proliferation
(p < 0.05, 4,942 �/� 302 cells in CRH�/�, compared to 3,906 ±
354 cells in CRH+/+ mice), but decreased cell survival in the OB
(p < 0.005, 25 ± 1 cell in CRH�/�, compared to 36 ± 2 cells in
CRH+/+ mice; Figures 4B and C). This increased proliferation in
CRH�/� mice is consistent with previous reports that stress
impairs neurogenesis both in the SVZ and in the hippocampus
(de Andrade et al., 2013; Hitoshi et al., 2007, Schoenfeld and
Gould, 2013), and that CRH�/� mice show decreased stress
levels (Jacobson et al., 2000). Cleaved caspase-3 and TUNEL
staining revealed increased apoptosis in the GCL of CRH�/�
Figure 4. CRH Signaling Is Required for Normal Levels of Adult-Born G
(A) Experimental scheme to determine cellular proliferation and granule cell surv
(B) Quantification of proliferating cells (BrdU and Ki67 double-positive cells) in th
(C) Quantification of adult-born granule cell survival in control and CRH�/� mice.
(D) Quantification of proliferating cells in the SVZ of CRHR1�/� mice (p > 0.05 St
(E) Quantification of adult-born granule cell survival in control and CRHR1�/� mi
(F) Representative images of the GCL of CRHR1+/+ and CRHR1f/f mice that expr
(G) Quantification of the ratio of Cre-EGFP+/tdTom+ granule cells (*p < 0.01 Stu
(H–P)Western blots of the synaptic proteins Synapsin, PSD95, andNR2B of OBs o
(*p < 0.05 Student’s t test). All data points represent averages ± SEM, n = 4 anim
See also Figure S4.
Developmen
mice (Figure S4A). To investigate whether granule cell apoptosis
was secondary to loss of CRH+ interneurons, we examined the
integrity of the EPL and performed cell counts using DAPI and
CR, which overlaps with EPL interneurons (Huang et al., 2013;
Figures S1D, S4B, and S4C) and found no difference. Moreover,
because CRH has important systemic effects as a regulatory
hormone, many of which are mediated by corticosteroids, we
questioned whether the attrition of granule cells in CRH�/�
mice was corticosteroid-dependent and supplemented adult
CRH�/� mice with corticosterone. Corticosteroid supplementa-
tion at a concentration that readily crosses the blood-brain bar-
rier and is capable of rescuing embryonic phenotypes in utero
(Muglia et al., 1995), did not change granule cell survival in
CRH�/� mice (Figure S4F), suggesting that adult-born neuron
survival was not mediated by systemic CRH signaling, but likely
through local CRH.
As loss of secreted CRH systemically in CRH�/� mice could
have secondary effects in the OB, we next assayed for prolifera-
tion and cell survival in CRHR1�/� mice, and saw no change in
SVZ-based proliferation compared to control littermates (p >
0.05, 4,172 ± 113 cells in CRHR�/� versus 4,337 ± 303 cells in
CRHR+/+ mice, Figure 4D). These data were consistent with
the observation that the SVZ lacked CRHR1 expression in
CRHR1-EGFP mice (data not shown), and that SVZ proliferation
might be mediated by systemic signaling rather than through
central CRH receptor activation. However, decreased numbers
of granule cells were noted in the OB 30 days post-BrdU pulsing
(p < 0.001, 24 ± 1 cells inCRHR�/� versus 32 ± 2 cells inCRHR+/+
mice, Figure 4E), as well as increased numbers of apoptotic cells
(Figure S4D). Moreover, this phenotype was also corticosteroid-
independent (Figure S4G).
To bypass any potential systemic effects of using germline
CRH loss-of-function alleles, we next conditionally removed
CRHR1 specifically from adult-born granule cells by injecting a
mixture of equal titers of AAV particles that expressed either
Cre-P2A-EGFP or tdTomato (control) into the RMS of
CRHR1+/+ or CRHR1flox/flox mice (Kuhne et al., 2012), and re-
vealed a 24.8% ± 2.8% decrease in the ratio of Cre-EGFP+/
tdTom+ granule cells between CRHR1+/+ and CRHR1flox/flox
mice (Figures 4F and 4G). Interestingly, morphological analysis
on the proportion of surviving granule cells showed no difference
between Cre and tdTomato+ neurons (data not shown).
Having found decreased numbers of surviving granule cells in
CRH loss-of-function mutants, we wondered if synaptic protein
expression was affected in these models and performed west-
ern blot analysis, which showed significantly decreased levels
of Synapsin, PSD95, and NR2B in the OBs of CRH�/�,
ranule Cell Survival and Synaptic Protein Expression
ival in CRH mutant alleles (scale bars represent 100 and 15 mm).
e SVZ of control and CRH�/� mice (*p < 0.05 Student’s t test).
udent’s t test).
ce (*p < 0.001 Student’s t test).
essed Cre-EGFP or tdTomato in granule cells (scale bar represents 50 mm).
dent’s t test).
fCRHR1�/�,CRHR1�/�, andCRHR1f/fmice injectedwith Cre or control viruses
als each.
tal Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc. 651

Figure 5. Constitutive CRHR Signaling in Granule Cells Promotes
Synaptogenic Changes in the OB
(A) Representative images of granule cells expressing tdTomato or tdTomato
and a constitutively active (CA) CRHR1-EGFP fusion construct (CA)
CRHR::EGFP. Scale bar represents 50 mm.
(B–D) Quantification of average granule cell length (B, p > 0.05 Student’s t test),
(C) mean total dendrite length (*p < 0.05), and (D) average dendritic branch
number in tdTomato and (CA)CRHR+ granule cells (*p < 0.01 Student’s t test).
(E) Scholl analysis of the number of intersections in tdTomato or (CA)CRHR+
granule cells (*p < 0.05 ANOVA).
Developmental Cell
Neuropeptide Signaling and Circuit Integration
652 Developmental Cell 30, 645–659, September 29, 2014 ª2014 Els
CRHR�/�, and CRHR1flox/flox mice compared to controls (Fig-
ures 4H–4P). Together, these data imply an important role for
CRH signaling in the maintenance and/or generation of synap-
ses in the OB.
We next asked if CRHR1 expression correlates with cell sur-
vival, and stained CRHR1-EGFP olfactory bulb tissue with acti-
vated caspase-3 (Figure S4H). Interestingly, we did not observe
any CRHR1-EGFP+/caspase-3+ cells. Together, these loss-of-
function and molecular marker data suggest that granule cells
require local CRH signaling for normal development andmatura-
tion, and that CRHR1 expression correlates with synapse for-
mation and/or survival, allowing granule cells to integrate into
existing brain circuits.
Gain-of-Function CRH Signaling Promotes IncreasedSynaptic Protein Expression in the OBHaving found that lack of CRH signaling leads to impaired
granule cell survival (Figure 4), we next queried the consequence
of enhanced CRH signaling. To determine a role for CRHR1 in
shaping synapse development and neuronal maturation, we ex-
pressed a constitutively active version of CRHR1 fused to EGFP,
(CA)CRHR1::EGFP (Nielsen et al., 2000) in newborn granule cells
(Figure 5A). Morphological characterization of granule cells
expressing (CA)CRHR1::EGFP revealed normal average neuron
length comparable to tdTomato controls (p > 0.05, 284 ± 21 mm
in (CA)CRHR1:EGFP versus 316 ± 19 mm in controls, Figure 5B),
but increased total dendrite length (p < 0.05, 894 ± 30 mm versus
672 ± 25 mm, Figure 5C), dendritic branch number (p < 0.01,
10.5 ± 0.65 mm versus 7 ± 0.7 mm, Figure 5D), and branch
intersections at both proximal and distal radii from the soma
(p < 0.05; Figure 5E). Next, we quantified dendritic spines
and found that total spine number was increased in (CA)
CRHR1:EGFP neurons (Figure 5F). Neurolucida reconstructions
revealed increased complexity in (CA)CRHR1::EGFP granule
cells compared to control neurons (Figure 5G).
Then, we targeted granule cells in the OB for conditional
gain-of-function studies with spatiotemporal specificity during
periods of endogenous CRHR1 expression to determine if
CRHR1 activation is sufficient for modulating synaptogenic
changes. For this, we first generated a BAC transgenic allele
to drive Cre recombinase from the CRHR1 promoter. To vali-
date cell type specificity of Cre expression, we generated
CRHR1-EGFP;CRHR1-Cre+/;ROSALSL-tdTom+/� mice, and saw
Cre activity that matched the expression pattern of CRHR1-
EGFP transgenic mice in the OB (Figure S5A). We next gener-
ated a conditional adeno-associated virus that carries (CA)
(F) Quantification of the number of dendritic spines between tdTomato and
(CA)CRHR+ granule cells (*p < 0.05 ANOVA). N = 10 cells each from three
animals.
(G) Representative granule cell morphology reconstructions.
(H) Experimental scheme for targeting CRHR1+ OB granule cells for consti-
tutive CRHR1-activation.
(I) Expression pattern of AAV-flex-(CA)CRHR1::GFP in granule cells (arrows
point to (CA)CRHR1::GFP+ neurons; scale bars represent 1,000, 100, and
20 mm).
(J–O) Western blot analysis of synaptic protein expression of CRHR-Cre+/�
OBs injected with either flexed GFP or flexed-(CA)CRHR-GFP AAV (*p < 0.05
Student’s t test, n = 4 animals each). All data points are averages ± SEM.
See also Figure S5.
evier Inc.

Developmental Cell
Neuropeptide Signaling and Circuit Integration
CRHR1::EGFP in an inverted ‘‘flexed’’ configuration (Atasoy
et al., 2008) and stereotaxically targeted the RMS for infection
of granule cells in CRHR1-Cre+/� mice (Figures 5H and 5I). To
confirm enhanced signaling through CRHR1, we isolated whole
OBs for western blot analysis. Control lysate was obtained from
CRHR1-Cre+/� mice injected with serotype-matched flexed
GFP virus. Since CRHR1 is Gs-coupled and leads to activation
of CREB via phosphorylation (Blank et al., 2003; Thiel and Ci-
belli, 1999), we first assayed pCREB levels in (CA)CRHR1-ex-
pressing OBs (Figure 5J), which were significantly increased.
Interestingly, we also observed significant changes in levels of
synaptic protein expression, including a large increase in the
presynaptic protein Synapsin (Figure 5K), suggesting the forma-
tion of de novo synapses or the strengthening of existing synap-
ses with enhanced CRHR1 signaling. Intriguingly, however,
although the quantity of PSD95 showed an increased trend of
expression, these changes were not statistically significant (Fig-
ure 5L). Because granule cells are GABAergic and form recip-
rocal dendrodendritic synapses with mitral cells (Mori et al.,
1983; Panzanelli et al., 2009; Shepherd and Greer, 2004), we
also assayed for changes in both inhibitory and excitatory re-
ceptor expression. The mitral cell-specific GABA-Aa1 (Whitman
and Greer, 2007), as well as NR1, and NR2B receptor subunits
showed increased expression (Figures 5M, 5N, and S5B),
whereas levels of AMPA receptor subtypes were decreased
compared to GFP controls (Figures 5O and S5C). Immunohisto-
chemistry for the upregulated proteins revealed increased
levels of NMDA and GABA receptor expression in the EPL of
gain-of-function experiments compared to controls (Figures
S5D–S5F). Intriguingly, increased NR1 and NR2B expression
was localized to both (CA)CRHR1::EGFP+ as well as EGFP
lacking dendritic structures, which could be due to the
nature of the membrane-bound overexpression construct. For
example, the fusion protein is not exclusively targeted to all
synaptic structures within newborn granule cells, and its syn-
aptogenic properties may in fact influence the formation of syn-
aptic structures in a more widespread manner. Consistent with
this, we also noted significant upregulation of GABA-Aa1,
which is expressed by mitral/tufted cells in the OB, and is
not expected to colocalize with (CA)CRHR::EGFP structures.
Together, these data suggest an overall increase in synap-
tic connectivity within the OB circuitry with constitutive
CRHR1 signaling, which then also may lead to non-cell-auton-
omous and more widespread increases in synaptic protein
expression.
Together, increased spine numbers, upregulated Synapsin
and NMDA receptor expression, combined with decreased
AMPA receptors suggests that CRH signaling in the OB pro-
motes the formation of new immature synapses, and/or potential
synaptic scaling of existing excitatory synapses (Turrigiano et al.,
1998; Turrigiano and Nelson, 2004).
Gain-of-Function CRH Signaling Promotes FunctionalSynaptogenesis in the OBA hallmark of bona fide circuit integration is functional syn-
aptic connectivity. Having determined that constitutively active
CRHR1 signaling leads to synaptogenic changes in the OB (Fig-
ure 5), we next queried the functional consequence of gain-of-
function CRH signaling using electrophysiology.
Developmen
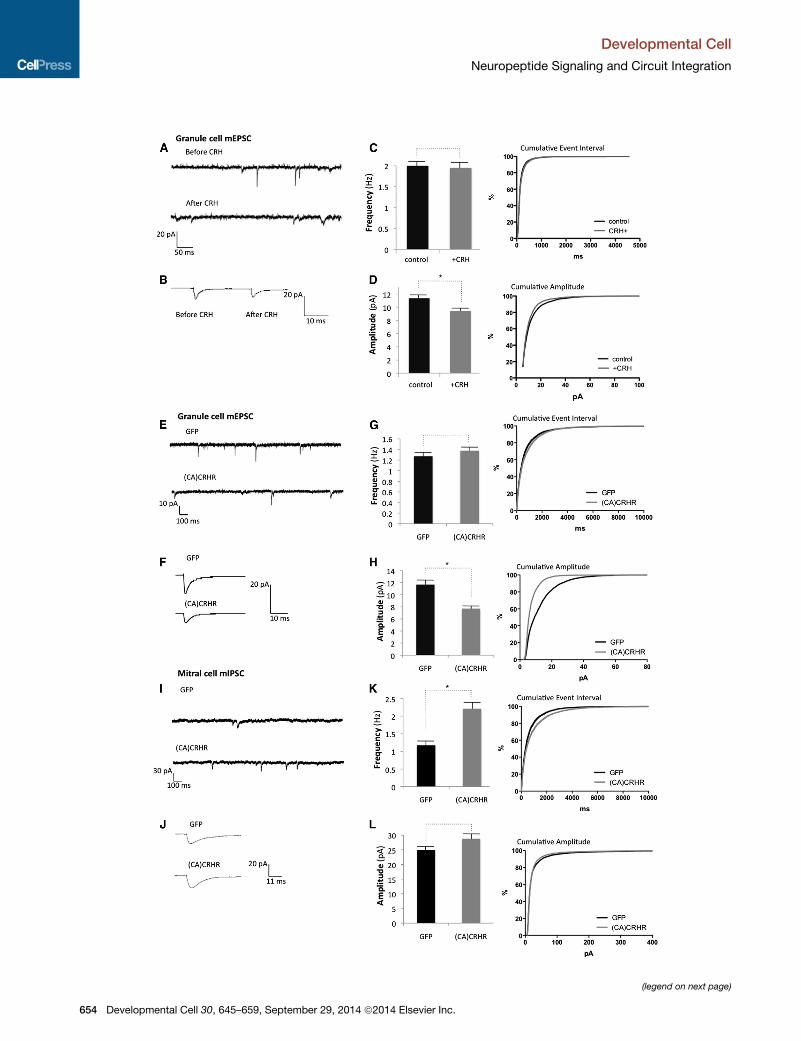
To determine if soluble CRH ligand influenced granule cell
electrophysiological properties, we made whole cell recordings
from CRHR1-EGFP+ granule cells while bath-applying CRH,
which showed no change in the frequency of miniature excit-
atory postsynaptic currents (mEPSCs), but induced a significant
decrease in amplitude (p < 0.05, 11.55 ± 0.72 pA before, and
8.8 ± 0.35 pA after CRH; Figures 6A–6D). Whole cell recordings
from granule cells that expressed (CA)CRHR1 compared to
EGFP controls showed no change in mEPSC frequency, but
significantly decreased mEPSC amplitudes (p < 0.05, 7.73 ±
1.38 pA in (CA)CRHR1 versus 11.69 ± 1.27 pA in EGFP controls;
Figures 6E–6H). These data were consistent with the observa-
tion that CRH gain-of-function signaling through activated
CRHR1 led to decreased AMPA receptor levels (Figures 5O
and S5C) and further suggested that upregulation of glutamater-
gic synapses via NMDA receptor expression likely reflects func-
tionally silent or immature synapses. Worth noting, we did not
observe any rapid changes in firing, passive membrane proper-
ties, or membrane potential following CRH application (data not
shown), suggesting that in the OB CRH does not act directly as a
neurotransmitter, but likely functions as a true neuromodulator.
With the observation that GABA-Aa1 receptor subunit expres-
sion increased with CRH signaling (Figure 5M), we next hypoth-
esized that functional GABAergic synapses from granule cells
onto mitral cells might be upregulated. To address this, we
recorded miniature inhibitory postsynaptic currents (mIPSCs)
from mitral cells in OBs where CRHR+ granule cells expressed
(CA)CRHR1::EGFP, and found a significant increase in fre-
quency of mIPSCs (p < 0.05, 2.21 ± 0.39 Hz in (CA)CRHR1
versus 1.17 ± 0.18 pA in EGFP controls, Figures 6I–6L), sug-
gesting either increased formation or stabilization of granule
cell-mitral cell synapses or changes in presynaptic release prop-
erties. Hence, biochemical and electrophysiological evidence
suggests that enhanced CRHR signaling in granule cells leads
to increased functional synaptogenesis and circuit plasticity in
the OB.
Acute Optogenetic Activation of CRH+ EPL InterneuronsPromotes Release of CRH in the OBCRH+ EPL interneurons make connections onto granule cells,
which in concert dynamically express CRHR1 (Figures 1, 2,
and 3). Lack of CRHR1 expression during a critical time window
in granule cell maturation and circuit integration leads to
decreased cell survival (Figure 4), and constitutively active
CRHR1 enhances synapse formation and circuit integration of
adult-born neurons (Figures 5 and 6). Together these data
support a mechanism by which activity-induced release of
CRH from EPL interneurons may influence granule cell synapto-
genesis. We next questioned if manipulating the activity of CRH+
EPL interneurons directly and acutely could dynamically recapit-
ulate the physiological effects observedwith constitutive CRHR1
activation.
To activate CRH+ EPL interneurons with spatiotemporal spec-
ificity, CRH-Cre+/� mice were crossed to ROSALSL-ChR2 mice
to obtain CRH-Cre+/�; ROSALSL-ChR2 mice that selectively ex-
pressed the light gated cation channel channelrhodopsin 2
(ChR2; Boyden et al., 2005; Nagel et al., 2003) in CRH+ EPL in-
terneurons in theOB. From thesemice, wemade slices of theOB
(Figure 7A) and hypothalamus (Figure S6), which were acutely
tal Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc. 653
(legend on next page)
Developmental Cell
Neuropeptide Signaling and Circuit Integration
654 Developmental Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc.

Developmental Cell
Neuropeptide Signaling and Circuit Integration
photostimulated ex vivo in small volumes of artificial cerebral spi-
nal fluid (ACSF). Light-stimulated depolarization of CRH+ neu-
rons led to CRH release in both hypothalamic control (Figure 7B)
and OB slices (Figure 7C), whereas soluble CRH levels in light-
stimulated controls were unchanged. Together, these data sug-
gest that the targeted depolarization of CRH+ neurons elicited
the release of stored CRH neuropeptide from EPL interneurons.
Finally, to determine the effects of CRH+ EPL interneuron
activation on granule cell synapses in vivo, CRH-Cre+/�;ROSALSL-ChR2+/� mice were chronically implanted with fiberop-
tics directly over the olfactory bulb (Ung and Arenkiel, 2012)
and photostimulated with blue laser light (Figure 7D). Acute
in vivo photostimulation in awake and behaving mice led to
enhanced olfactory bulb expression of pCREB (Figure 7E), reca-
pitulating the effect seen in constitutive CRHR1 activation (Fig-
ure 5J). Western blots of stimulated animals showed significant
upregulation of the synaptic proteins Synapsin, PSD95, and
NR2B (Figures 7F–7H), suggesting a conserved synaptogenic
effect of photostimulated release of CRH from EPL interneurons
in vivo, which mirrored constitutively active CRHR1 signaling
(Figure 5).
DISCUSSION
Neuropeptides, including CRH, have been implicated in a variety
of neuronal processes, ranging from neuromodulation and den-
dritic outgrowth, to neuroprotection (Chen et al., 2004; Hanstein
et al., 2008; Sheng et al., 2012). However, the precise role of CRH
in the maturation and integration of granule cells in the OB has
not been investigated. Here, we describe a neuropeptidergic
function for inhibitory interneurons in shaping cell survival, syn-
aptogenesis, and circuit integration of new neurons in the adult
brain that is distinct from classical neurotransmitter signaling.
Together, our data support a potential mechanism for a tripar-
tite-like interaction between mitral cells, EPL interneurons, and
granule cells (Figure 7I). We and others have recently shown
that mitral cells and EPL interneurons exhibit reciprocal connec-
tivity (Huang et al., 2013, Kato et al., 2013; Miyamichi et al.,
2013). It is conceivable that neuronal activity via olfactory sen-
sation is conveyed to mitral cells, which in turn stimulate EPL
interneurons to both shapemitral cell output, and release soluble
CRH. In this way, activity-dependent release of CRH, and
subsequent reception by granule cells that express CRHR1
both synaptically and/or extrasynaptically, may function as
a key mechanism toward modulating synaptogenesis and
plasticity via neuropeptide signaling, linking activity to circuit
integration via neuropeptidergic interneurons. Ultimately, this
interaction has the potential to shape whole circuit activity and
Figure 6. Constitutive CRHR Signaling in Granule Cells Promotes Syna
(A) Representative trace of CRHR1-EGFP+ granule cell before and after CRH ap
(B) Average mEPSCs before and after CRH.
(C and D) Quantification of the mEPSC frequency and amplitude before (C) and
(E) Representative mEPSC traces of granule cells from CRHR-Cre+/� mice inject
(F) Average mEPSCs of EGFP and (CA)CRHR1::EGFP+ granule cells.
(G and H) Quantified granule cell mEPSC frequency and amplitude (*p < 0.05 St
(I) Representative mIPSC traces of mitral cells from CRHR-Cre+/� mice injected
(J) Average mitral cell mIPSCs.
(K and L) Average frequency and amplitude of mitral cell mIPSCs in CRHR-Cre+
CRHR1::EGFP (*p < 0.05 Student’s t test, n = 13 mitral cells per group from thre
Developmen
plasticity. Although this model supports a role for local interac-
tions, it remains to be determined if other forms of neuromodu-
latory signals are conveyed specifically to EPL interneurons via
other local or centrifugal inputs.
Dual Roles of InterneuronsNeuropeptides are traditionally acknowledged for roles in
shaping whole-body physiological responses and/or modulating
systemic homeostatic mechanisms (Vale et al., 1981). Recently,
neuropeptides have received increasing attention for their roles
in shaping synapses and facilitating neuronal plasticity (Bayatti
et al., 2003; Fenoglio et al., 2006; Lipschitz et al., 2005). Inter-
estingly, however, interneurons have been most extensively
studied with respect to their traditional, GABAergic inhibitory
function, and the neuropeptides they express have served pri-
marily in categorizing the many different interneuron subtypes
throughout the brain (Ma et al., 2006; Rudy et al., 2011; Xu
et al., 2013). Although granule cells make up the majority of
inhibitory interneurons in the OB, previous work from our lab
and others has identified other types of interneurons with
GABAergic connections onto mitral cells that likely serve impor-
tant olfactory functions (Huang et al., 2013; Kato et al., 2013;
Miyamichi et al., 2013; Lepousez et al., 2010a, 2010b; Kosaka
and Kosaka, 2008). Here, we have identified a population of
inhibitory interneurons in the OB with a neuropeptidergic role
in promoting synaptic protein expression and circuit integration.
Interestingly, this interneuron population has a dual role in
shaping the OB circuitry and likely, olfaction. This interaction is
inhibitory onto mitral cells and neuromodulatory onto granule
cells, suggesting increased connectivity and/or signaling not
only between excitatory principal cells and inhibitory interneu-
rons, but also between different subtypes of interneurons. This
apparent dual functionality may indeed exemplify a prominent
mechanism to support neuronal plasticity, and a way to shape
circuitries by inhibitory neurotransmitter signaling onto mitral
cells, and neuromodulatory signaling onto granule cells. EPL in-
terneurons form reciprocal GABAergic connections with mitral
cells, and are in turn depolarized following odor stimulation
(Huang et al., 2013; Kato et al., 2013; Miyamichi et al., 2013).
This activation further strengthens mitral cell inhibition, but
may also facilitate CRH release, linking olfactory-dependent
activity to neuromodulation, and ultimately synaptogenesis
and circuit integration.
Local Neuropeptide Signaling, Synaptogenesis, andCircuit IntegrationOur data suggest that local neuromodulatory signaling by CRH+
EPL interneurons in the OB aids in granule cell circuit integration
ptic and Circuit Plasticity in the OB
plication (500 nM CRH).
after (D) CRH bath application (*p < 0.05, n = 13 cells from three animals).
ed with either AAV-flexed EGFP or AAV-flexed-(CA)CRHR1::EGFP.
udent’s t test, n = 11 granule cells per group from three animals).
with either AAV-flexed EGFP or AAV-flexed-(CA)CRHR1::EGFP.
/� OBs in which CRHR-expressing granule cells express either EGFP or (CA)
e animals). All data points represent mean ± SEM.
tal Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc. 655

Figure 7. Optogenetic Activation of CRH+ EPL Interneurons Induces Synaptogenesis in the OB
(A) ChR2 expression pattern of CRH-Cre+/�; ROSALSL-ChR2+/� mice (scale bars represent 200 and 50 mm).
(B and C) Quantification of CRH concentration with hypothalamic or OB optogenetic activation in ROSALSL-ChR2+/� (control) or CRH-Cre+/�; ROSALSL-ChR2+/�
mice (*p < 0.05 Student’s t test, n = 3 animals per group).
(D) Experimental scheme for in vivo photostimulation of CRH+ EPL interneurons.
(legend continued on next page)
Developmental Cell
Neuropeptide Signaling and Circuit Integration
656 Developmental Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc.

Developmental Cell
Neuropeptide Signaling and Circuit Integration
by increasing synaptic protein expression and/or stabilizing
existing synapses. Removing CRH signaling decreases synaptic
protein expression, whereas enhancing CRH signaling induces
synaptic protein expression in theOB. Perhapsmore intriguingly,
the result that CRHR loss-of-function does not completely
abolish adult-born neuron survival and integration poses some
additional questions. Given that numerous neuropeptides,
including aomatostatin, vasointestinal peptide, cholecystokinin,
oxytocin, and neuropeptide Y are present in the olfactory bulb
(Gracia-Llanes et al., 2003; Lepousez et al., 2010a, 2010b; Ma
et al., 2006; Tobin et al., 2010), and that granule cells in turn
appear to express the cognate receptors, it remains to be seen
if other neuromodulators have the same effect on the OB cir-
cuitry, and if these neuropeptides act in concert to promote
synaptogenesis and circuit integration in the adult brain.
EXPERIMENTAL PROCEDURES
Experimental Animals
All experimental animals were treated in compliance with the United States
Department of Health and Human Services and the Baylor College of Medicine
IUACUC guidelines. ROSARITVA/RITVA (Takatoh et al., 2013), ROSALSL-tdTom
(Arenkiel et al., 2011), CRHR1-EGFP mice (Justice et al., 2008), and
CRHR1flox/flox mice (Kuhne et al., 2012) were previously described. CRH-Cre
(Taniguchi et al., 2011), ROSALSL-ChR2 (Madisen et al., 2012), CRH�/� (Muglia
et al., 1995), and CRHR1�/� (Smith et al., 1998) were obtained from Jackson
Laboratories and maintained on a C57BL/6 background. Generation of the
CRHR1-Cre line is described in the Supplemental Experimental Procedures.
Transsynaptic Tracing
Briefly, adult (6–8weeks old)ROSARITVA/RITVAmicewere injectedwith high titer
lentivirus encoding tdTomato-IRES-Cre stereotaxically into the RMS. Twenty-
eight days later, mice were injected with low titer SADG-EGFP RV (Wicker-
sham et al., 2007b) into the core of the OB and killed after 7 days for mapping
studies. Next, CRH-Cre+/�; ROSALSL-tdTom+/� pups were electroporated with
an expression construct for rabies-G and TVA as previously described (Are-
nkiel et al., 2011). Twenty-eight days postelectroporation, mice were injected
with SADG-EGFP RV as described above. See also Supplemental Experi-
mental Procedures.
Immunohistochemistry and Imaging
OB tissues were processed for immunohistochemistry as previously
described (Huang et al., 2013). Briefly, free-floating sections were stained
with the following primary antibodies: guinea pig anti-Parvalbumin, and rabbit
anti-CRH (kindly provided by Nick Justice), followed by washing and incuba-
tion with species-specific Alexa-633 secondary antibodies. Confocal image
analysis was performed using a Leica TCS SPE confocal microscope. See
also Supplemental Experimental Procedures
CRH–/– and CRHR1–/– Adult-Born Neuron Proliferation, Survival,
and Apoptosis
Age-matched adult (6–8 weeks old) male mice were injected with BrdU. For
SVZ-based proliferation studies, animals were killed 24 hr later as and 50-
mm-thick sections were taken throughout the SVZ and stained with mouse
anti-BrdU and rabbit anti-Ki67 to assess proliferating cells. To assay granule
cell survival, mice were killed 30 days later and processed for immunohisto-
chemistry using mouse anti-BrdU antibody. Images were taken in the middle
and outer granule cell layer, avoiding the RMS and IPL/MCL regions, and total
(E–H) Western blot analysis of synaptic protein expression of ROSALSL-ChR2+/� (c
animals per group).
(I) Model: CRH signaling between local EPL interneurons and granule cells prom
mean ± SEM.
See also Figure S6.
Developmen
cell numbers were counted through serial sections. See also Supplemental
Experimental Procedures.
Constitutively Active CRHR1 Overexpression
A constitutively active CRHR1::EGFP fusion construct (Nielsen et al., 2000)
was electroporated into the SVZ of P2 pups for morphology analysis using
Neurolucida software. The construct was further subcloned into a conditional
flexed adeno-associated-viral vector (Atasoy et al., 2008) and packaged into
viral particles. Virus was stereotaxically injected into the RMS of CRHR1-Cre
mice, and OB tissue was harvested 14 days postinjection. See also Supple-
mental Experimental Procedures.
Electrophysiology
Animals (P21–P35) were deeply anesthetized using isoflurane, and perfused
intracardially with ice-cold ACSF. Coronal olfactory bulb slices (300 mm)
were placed in a room-temperature chamber mounted on an Olympus upright
microscope (BX50WI), and perfused with oxygenated ACSF. Cells were
depicted using fluorescence and differential interference contrast imaging.
Synaptic currents were recorded using cesium-methanesulfonate based inter-
nal solutions. See also Supplemental Experimental Procedures.
In Vivo Photostimulation
Fiberoptics were generated and implanted directly over the olfactory bulb as
previously described (Ung and Arenkiel, 2012). Animals were allowed to
recover from the surgery for 3 days prior to photostimulation. Photostimulation
was performed for 3 hr using a blue laser source (CrystaLaser) controlled by a
Master-8 (A.M.P.I.). See also Supplemental Experimental Procedures.
Statistical Methods
Unless otherwise indicated, statistical comparisons between experimental
groups were made using Student’s t test and all error bars represent SEM.
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures
and six figures and can be found with this article online http://dx.doi.org/10.
1016/j.devcel.2014.07.001.
ACKNOWLEDGMENTS
This work was supported by the McNair Medical Institute, NINDS grant
F31NS081805 to I.G. and NINDS R01NS078294 to B.R.A. We thank Drs.
Dona Murphey, Ian Davison, and Steve Shea for critical review of this
manuscript.
Received: December 8, 2013
Revised: May 2, 2014
Accepted: June 30, 2014
Published: September 4, 2014
REFERENCES
Abraham, N.M., Egger, V., Shimshek, D.R., Renden, R., Fukunaga, I.,
Sprengel, R., Seeburg, P.H., Klugmann, M., Margrie, T.W., Schaefer, A.T.,
and Kuner, T. (2010). Synaptic inhibition in the olfactory bulb accelerates
odor discrimination in mice. Neuron 65, 399–411.
Abrous, D.N., Koehl, M., and LeMoal, M. (2005). Adult neurogenesis: from pre-
cursors to network and physiology. Physiol. Rev. 85, 523–569.
Alvarez-Buylla, A., and Temple, S. (1998). Stem cells in the developing and
adult nervous system. J. Neurobiol. 36, 105–110.
ontrol) or CRH-Cre+/�; ROSALSL-ChR2+/� mice (*p < 0.05 Student’s t test, n = 4
otes synapse formation and stabilization in the OB. All data points represent
tal Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc. 657

Developmental Cell
Neuropeptide Signaling and Circuit Integration
Arenkiel, B.R., Hasegawa, H., Yi, J.J., Larsen, R.S., Wallace, M.L., Philpot,
B.D., Wang, F., and Ehlers, M.D. (2011). Activity-induced remodeling of olfac-
tory bulb microcircuits revealed by monosynaptic tracing. PLoS ONE 6,
e29423.
Atasoy, D., Aponte, Y., Su, H.H., and Sternson, S.M. (2008). A FLEX switch
targets Channelrhodopsin-2 to multiple cell types for imaging and long-range
circuit mapping. J. Neurosci. 28, 7025–7030.
Bale, T.L., and Vale, W.W. (2004). CRF and CRF receptors: role in stress
responsivity and other behaviors. Annu. Rev. Pharmacol. Toxicol. 44,
525–557.
Balu, R., Pressler, R.T., and Strowbridge, B.W. (2007). Multiple modes of
synaptic excitation of olfactory bulb granule cells. J. Neurosci. 27, 5621–5632.
Barinka, F., Salaj, M., Ryba�r, J., Kraj�covi�cova, E., Kubova, H., and Druga, R.
(2012). Calretinin, parvalbumin and calbindin immunoreactive interneurons in
perirhinal cortex and temporal area Te3V of the rat brain: qualitative and
quantitative analyses. Brain Res. 1436, 68–80.
Batista-Brito, R., Close, J., Machold, R., and Fishell, G. (2008). The distinct
temporal origins of olfactory bulb interneuron subtypes. J. Neurosci. 28,
3966–3975.
Bayatti, N., Zschocke, J., and Behl, C. (2003). Brain region-specific neuropro-
tective action and signaling of corticotropin-releasing hormone in primary
neurons. Endocrinology 144, 4051–4060.
Berger, H., Heinrich, N., Wietfeld, D., Bienert, M., and Beyermann, M. (2006).
Evidence that corticotropin-releasing factor receptor type 1 couples to Gs-
and Gi-proteins through different conformations of its J-domain. Br. J.
Pharmacol. 149, 942–947.
Blank, T., Nijholt, I., Grammatopoulos, D.K., Randeva, H.S., Hillhouse, E.W.,
and Spiess, J. (2003). Corticotropin-releasing factor receptors couple to
multiple G-proteins to activate diverse intracellular signaling pathways in
mouse hippocampus: role in neuronal excitability and associative learning.
J. Neurosci. 23, 700–707.
Boyden, E.S., Zhang, F., Bamberg, E., Nagel, G., and Deisseroth, K. (2005).
Millisecond-timescale, genetically targeted optical control of neural activity.
Nat. Neurosci. 8, 1263–1268.
Breton-Provencher, V., Lemasson, M., Peralta, M.R., 3rd, and Saghatelyan, A.
(2009). Interneurons produced in adulthood are required for the normal func-
tioning of the olfactory bulb network and for the execution of selected olfactory
behaviors. J. Neurosci. 29, 15245–15257.
Carleton, A., Petreanu, L.T., Lansford, R., Alvarez-Buylla, A., and Lledo, P.M.
(2003). Becoming a new neuron in the adult olfactory bulb. Nat. Neurosci. 6,
507–518.
Chen, W.R., and Greer, C.A. (2004). Olfactory bulb. In The Synaptic
Organization of the Brain, Fifth Edition, G.M. Shepherd, ed. (New York:
Oxford University Press), pp. 165–216.
Chen, Y., Bender, R.A., Brunson, K.L., Pomper, J.K., Grigoriadis, D.E., Wurst,
W., and Baram, T.Z. (2004). Modulation of dendritic differentiation by cortico-
tropin-releasing factor in the developing hippocampus. Proc. Natl. Acad. Sci.
USA 101, 15782–15787.
de Andrade, J.S., Cespedes, I.C., Abrao, R.O., Dos Santos, T.B., Diniz, L.,
Britto, L.R., Spadari-Bratfisch, R.C., Ortolani, D., Melo-Thomas, L., da Silva,
R.C., and Viana, M.B. (2013). Chronic unpredictable mild stress alters an
anxiety-related defensive response, Fos immunoreactivity and hippocampal
adult neurogenesis. Behav. Brain Res. 250, 81–90.
De Marco Garcıa, N.V., Karayannis, T., and Fishell, G. (2011). Neuronal activity
is required for the development of specific cortical interneuron subtypes.
Nature 472, 351–355.
Eyre, M.D., Antal, M., and Nusser, Z. (2008). Distinct deep short-axon cell sub-
types of the main olfactory bulb provide novel intrabulbar and extrabulbar
GABAergic connections. J. Neurosci. 28, 8217–8229.
Favero, M., and Castro-Alamancos, M.A. (2013). Synaptic cooperativity regu-
lates persistent network activity in neocortex. J. Neurosci. 33, 3151–3163.
Fenoglio, K.A., Chen, Y., and Baram, T.Z. (2006). Neuroplasticity of the hypo-
thalamic-pituitary-adrenal axis early in life requires recurrent recruitment of
stress-regulating brain regions. J. Neurosci. 26, 2434–2442.
658 Developmental Cell 30, 645–659, September 29, 2014 ª2014 Els
Gracia-Llanes, F.J., Crespo, C., Blasco-Ibanez, J.M., Marques-Marı, A.I., and
Martınez-Guijarro, F.J. (2003). VIP-containing deep short-axon cells of the
olfactory bulb innervate interneurons different from granule cells. Eur. J.
Neurosci. 18, 1751–1763.
Hanstein, R., Lu, A., Wurst, W., Holsboer, F., Deussing, J.M., Clement, A.B.,
and Behl, C. (2008). Transgenic overexpression of corticotropin releasing
hormone provides partial protection against neurodegeneration in an in vivo
model of acute excitotoxic stress. Neuroscience 156, 712–721.
Hensch, T.K., Fagiolini, M., Mataga, N., Stryker, M.P., Baekkeskov, S., and
Kash, S.F. (1998). Local GABA circuit control of experience-dependent plas-
ticity in developing visual cortex. Science 282, 1504–1508.
Hitoshi, S., Maruta, N., Higashi, M., Kumar, A., Kato, N., and Ikenaka, K. (2007).
Antidepressant drugs reverse the loss of adult neural stem cells following
chronic stress. J. Neurosci. Res. 85, 3574–3585.
Huang, L., Garcia, I., Jen, H.I., and Arenkiel, B.R. (2013). Reciprocal connec-
tivity between mitral cells and external plexiform layer interneurons in the
mouse olfactory bulb. Front Neural Circuits 7, 32.
Isaacson, J.S., and Strowbridge, B.W. (1998). Olfactory reciprocal synapses:
dendritic signaling in the CNS. Neuron 20, 749–761.
Jacobson, L., Muglia, L.J., Weninger, S.C., Pacak, K., and Majzoub, J.A.
(2000). CRH deficiency impairs but does not block pituitary-adrenal responses
to diverse stressors. Neuroendocrinology 71, 79–87.
Justice, N.J., Yuan, Z.F., Sawchenko, P.E., and Vale, W. (2008). Type 1 corti-
cotropin-releasing factor receptor expression reported in BAC transgenic
mice: implications for reconciling ligand-receptor mismatch in the central
corticotropin-releasing factor system. J. Comp. Neurol. 511, 479–496.
Kato, H.K., Gillet, S.N., Peters, A.J., Isaacson, J.S., and Komiyama, T. (2013).
Parvalbumin-expressing interneurons linearly control olfactory bulb output.
Neuron 80, 1218–1231.
Kelsch, W., Lin, C.W., Mosley, C.P., and Lois, C. (2009). A critical period for
activity-dependent synaptic development during olfactory bulb adult neuro-
genesis. J. Neurosci. 29, 11852–11858.
Kosaka, T., and Kosaka, K. (2008). Heterogeneity of parvalbumin-containing
neurons in the mouse main olfactory bulb, with special reference to short-
axon cells and betaIV-spectrin positive dendritic segments. Neurosci. Res.
60, 56–72.
Kuhne, C., Puk, O., Graw, J., Hrab�e de Angelis, M., Schutz, G., Wurst, W., and
Deussing, J.M. (2012). Visualizing corticotropin-releasing hormone receptor
type 1 expression and neuronal connectivities in the mouse using a novel
multifunctional allele. J. Comp. Neurol. 520, 3150–3180.
LeMagueresse, C., and Monyer, H. (2013). GABAergic interneurons shape the
functional maturation of the cortex. Neuron 77, 388–405.
Le Roux, N., Cabezas, C., Bohm, U.L., and Poncer, J.C. (2013). Input-specific
learning rules at excitatory synapses onto hippocampal parvalbumin-express-
ing interneurons. J. Physiol. 591, 1809–1822.
Lemasson, M., Saghatelyan, A., Olivo-Marin, J.C., and Lledo, P.M. (2005).
Neonatal and adult neurogenesis provide two distinct populations of newborn
neurons to the mouse olfactory bulb. J. Neurosci. 25, 6816–6825.
Lepousez, G., Csaba, Z., Bernard, V., Loudes, C., Videau, C., Lacombe, J.,
Epelbaum, J., and Viollet, C. (2010a). Somatostatin interneurons delineate
the inner part of the external plexiform layer in the mouse main olfactory
bulb. J. Comp. Neurol. 518, 1976–1994.
Lepousez, G., Mouret, A., Loudes, C., Epelbaum, J., and Viollet, C. (2010b).
Somatostatin contributes to in vivo gamma oscillation modulation and odor
discrimination in the olfactory bulb. J. Neurosci. 30, 870–875.
Lipschitz, D.L., Crowley, W.R., Armstrong, W.E., and Bealer, S.L. (2005).
Neurochemical bases of plasticity in themagnocellular oxytocin system during
gestation. Exp. Neurol. 196, 210–223.
Lledo, P.M., Merkle, F.T., and Alvarez-Buylla, A. (2008). Origin and function of
olfactory bulb interneuron diversity. Trends Neurosci. 31, 392–400.
Ma, Y., Hu, H., Berrebi, A.S., Mathers, P.H., and Agmon, A. (2006). Distinct
subtypes of somatostatin-containing neocortical interneurons revealed in
transgenic mice. J. Neurosci. 26, 5069–5082.
evier Inc.

Developmental Cell
Neuropeptide Signaling and Circuit Integration
Madisen, L., Mao, T., Koch, H., Zhuo, J.M., Berenyi, A., Fujisawa, S., Hsu,
Y.W., Garcia, A.J., 3rd, Gu, X., Zanella, S., et al. (2012). A toolbox of Cre-
dependent optogenetic transgenic mice for light-induced activation and
silencing. Nat. Neurosci. 15, 793–802.
Maras, P.M., and Baram, T.Z. (2012). Sculpting the hippocampus from within:
stress, spines, and CRH. Trends Neurosci. 35, 315–324.
Ming, G.L., and Song, H. (2005). Adult neurogenesis in the mammalian central
nervous system. Annu. Rev. Neurosci. 28, 223–250.
Miyamichi, K., Shlomai-Fuchs, Y., Shu, M., Weissbourd, B.C., Luo, L., and
Mizrahi, A. (2013). Dissecting local circuits: parvalbumin interneurons underlie
broad feedback control of olfactory bulb output. Neuron 80, 1232–1245.
Mori, K., Kishi, K., and Ojima, H. (1983). Distribution of dendrites of mitral, dis-
placed mitral, tufted, and granule cells in the rabbit olfactory bulb. J. Comp.
Neurol. 219, 339–355.
Mouret, A., Gheusi, G., Gabellec, M.M., de Chaumont, F., Olivo-Marin, J.C.,
and Lledo, P.M. (2008). Learning and survival of newly generated neurons:
when time matters. J. Neurosci. 28, 11511–11516.
Mouret, A., Lepousez, G., Gras, J., Gabellec, M.M., and Lledo, P.M. (2009).
Turnover of newborn olfactory bulb neurons optimizes olfaction. J. Neurosci.
29, 12302–12314.
Muglia, L., Jacobson, L., Dikkes, P., and Majzoub, J.A. (1995). Corticotropin-
releasing hormone deficiency reveals major fetal but not adult glucocorticoid
need. Nature 373, 427–432.
Nagel, G., Szellas, T., Huhn, W., Kateriya, S., Adeishvili, N., Berthold, P., Ollig,
D., Hegemann, P., and Bamberg, E. (2003). Channelrhodopsin-2, a directly
light-gated cation-selective membrane channel. Proc. Natl. Acad. Sci. USA
100, 13940–13945.
Nielsen, S.M., Nielsen, L.Z., Hjorth, S.A., Perrin, M.H., and Vale, W.W. (2000).
Constitutive activation of tethered-peptide/corticotropin-releasing factor re-
ceptor chimeras. Proc. Natl. Acad. Sci. USA 97, 10277–10281.
Panzanelli, P., Bardy, C., Nissant, A., Pallotto, M., Sassoe-Pognetto,M., Lledo,
P.M., and Fritschy, J.M. (2009). Early synapse formation in developing inter-
neurons of the adult olfactory bulb. J. Neurosci. 29, 15039–15052.
Perrin, M.H., and Vale, W.W. (1999). Corticotropin releasing factor receptors
and their ligand family. Ann. N Y Acad. Sci. 885, 312–328.
Perrin, M.H., Donaldson, C.J., Chen, R., Lewis, K.A., and Vale, W.W. (1993).
Cloning and functional expression of a rat brain corticotropin releasing factor
(CRF) receptor. Endocrinology 133, 3058–3061.
Perrin, M., Donaldson, C., Chen, R., Blount, A., Berggren, T., Bilezikjian, L.,
Sawchenko, P., and Vale, W. (1995). Identification of a second corticotropin-
releasing factor receptor gene and characterization of a cDNA expressed in
heart. Proc. Natl. Acad. Sci. USA 92, 2969–2973.
Pressler, R.T., and Strowbridge, B.W. (2006). Blanes cells mediate persistent
feedforward inhibition onto granule cells in the olfactory bulb. Neuron 49,
889–904.
Price, J.L., and Powell, T.P. (1970a). An electron-microscopic study of the
termination of the afferent fibres to the olfactory bulb from the cerebral hemi-
sphere. J. Cell Sci. 7, 157–187.
Price, J.L., and Powell, T.P. (1970b). The synaptology of the granule cells of the
olfactory bulb. J. Cell Sci. 7, 125–155.
Refojo, D., Schweizer, M., Kuehne, C., Ehrenberg, S., Thoeringer, C., Vogl,
A.M., Dedic, N., Schumacher, M., von Wolff, G., Avrabos, C., et al. (2011).
Glutamatergic and dopaminergic neurons mediate anxiogenic and anxiolytic
effects of CRHR1. Science 333, 1903–1907.
Rochefort, C., Gheusi, G., Vincent, J.D., and Lledo, P.M. (2002). Enriched odor
exposure increases the number of newborn neurons in the adult olfactory bulb
and improves odor memory. J. Neurosci. 22, 2679–2689.
Roozendaal, B., Schelling, G., and McGaugh, J.L. (2008). Corticotropin-
releasing factor in the basolateral amygdala enhances memory consolidation
via an interaction with the beta-adrenoceptor-cAMP pathway: dependence on
glucocorticoid receptor activation. J. Neurosci. 28, 6642–6651.
Developmen
Rudy, B., Fishell, G., Lee, S., and Hjerling-Leffler, J. (2011). Three groups of
interneurons account for nearly 100% of neocortical GABAergic neurons.
Dev. Neurobiol. 71, 45–61.
Schmolesky, M.T., De Ruiter, M.M., De Zeeuw, C.I., and Hansel, C. (2007). The
neuropeptide corticotropin-releasing factor regulates excitatory transmission
and plasticity at the climbing fibre-Purkinje cell synapse. Eur. J. Neurosci.
25, 1460–1466.
Schoenfeld, T.J., and Gould, E. (2013). Differential effects of stress and gluco-
corticoids on adult neurogenesis. Curr. Top. Behav. Neurosci. 15, 139–164.
Sheng, H., Xu, Y., Chen, Y., Zhang, Y., Xu, X., He, C., and Ni, X. (2012). CRH-R1
and CRH-R2 differentially modulate dendritic outgrowth of hippocampal neu-
rons. Endocrine 41, 458–464.
Shepherd, G.M., and Greer, C.A. (2004). Olfactory Bulb in the Synaptic
Organization of the Brain, G.M. Shepherd, ed. (New York: Oxford University
Press), pp. 159–204.
Smith, G.W., Aubry, J.M., Dellu, F., Contarino, A., Bilezikjian, L.M., Gold, L.H.,
Chen, R., Marchuk, Y., Hauser, C., Bentley, C.A., et al. (1998). Corticotropin
releasing factor receptor 1-deficient mice display decreased anxiety, impaired
stress response, and aberrant neuroendocrine development. Neuron 20,
1093–1102.
Takatoh, J., Nelson, A., Zhou, X., Bolton, M.M., Ehlers, M.D., Arenkiel, B.R.,
Mooney, R., and Wang, F. (2013). New modules are added to vibrissal premo-
tor circuitry with the emergence of exploratory whisking. Neuron 77, 346–360.
Taniguchi, H., He, M., Wu, P., Kim, S., Paik, R., Sugino, K., Kvitsiani, D., Fu, Y.,
Lu, J., Lin, Y., et al. (2011). A resource of Cre driver lines for genetic targeting of
GABAergic neurons in cerebral cortex. Neuron 71, 995–1013.
Thiel, G., and Cibelli, G. (1999). Corticotropin-releasing factor and vasoactive
intestinal polypeptide activate gene transcription through the cAMP signaling
pathway in a catecholaminergic immortalized neuron. Neurochem. Int. 34,
183–191.
Tobin, V.A., Hashimoto, H., Wacker, D.W., Takayanagi, Y., Langnaese, K.,
Caquineau, C., Noack, J., Landgraf, R., Onaka, T., Leng, G., et al. (2010). An
intrinsic vasopressin system in the olfactory bulb is involved in social recogni-
tion. Nature 464, 413–417.
Turrigiano, G.G., and Nelson, S.B. (2004). Homeostatic plasticity in the devel-
oping nervous system. Nat. Rev. Neurosci. 5, 97–107.
Turrigiano, G.G., Leslie, K.R., Desai, N.S., Rutherford, L.C., and Nelson, S.B.
(1998). Activity-dependent scaling of quantal amplitude in neocortical neu-
rons. Nature 391, 892–896.
Ung, K., and Arenkiel, B.R. (2012). Fiber-optic implantation for chronic optoge-
netic stimulation of brain tissue. J. Vis. Exp. e50004.
Vale, W., Spiess, J., Rivier, C., and Rivier, J. (1981). Characterization of a 41-
residue ovine hypothalamic peptide that stimulates secretion of corticotropin
and beta-endorphin. Science 213, 1394–1397.
Vale, W., Rivier, C., Brown, M.R., Spiess, J., Koob, G., Swanson, L., Bilezikjian,
L., Bloom, F., and Rivier, J. (1983). Chemical and biological characterization of
corticotropin releasing factor. Recent Prog. Horm. Res. 39, 245–270.
Whitman, M.C., and Greer, C.A. (2007). Synaptic integration of adult-gener-
ated olfactory bulb granule cells: basal axodendritic centrifugal input precedes
apical dendrodendritic local circuits. J. Neurosci. 27, 9951–9961.
Wickersham, I.R., Finke, S., Conzelmann, K.K., and Callaway, E.M. (2007a).
Retrograde neuronal tracing with a deletion-mutant rabies virus. Nat.
Methods 4, 47–49.
Wickersham, I.R., Lyon, D.C., Barnard, R.J., Mori, T., Finke, S., Conzelmann,
K.K., Young, J.A., and Callaway, E.M. (2007b). Monosynaptic restriction of
transsynaptic tracing from single, genetically targeted neurons. Neuron 53,
639–647.
Xu, H., Jeong, H.Y., Tremblay, R., and Rudy, B. (2013). Neocortical somato-
statin-expressing GABAergic interneurons disinhibit the thalamorecipient
layer 4. Neuron 77, 155–167.
Yamaguchi, M., and Mori, K. (2005). Critical period for sensory experience-
dependent survival of newly generated granule cells in the adult mouse olfac-
tory bulb. Proc. Natl. Acad. Sci. USA 102, 9697–9702.
tal Cell 30, 645–659, September 29, 2014 ª2014 Elsevier Inc. 659