expression, localization, and...
TRANSCRIPT

JPET #162800
1
EXPRESSION, LOCALIZATION, AND PHARMACOLOGICAL ROLE OF KV7 POTASSIUM CHANNELS IN SKELETAL MUSCLE PROLIFERATION,
DIFFERENTIATION AND SURVIVAL AFTER MYOTOXIC INSULTS
Fabio Arturo Iannotti, Elisabetta Panza, Vincenzo Barrese, Davide Viggiano, Maria Virginia Soldovieri, and Maurizio Taglialatela*
Division of Pharmacology, Dept. of Neuroscience, University of Naples Federico II, Naples, Italy (F.A.I., E.P., V.B., and M.T.); and Dept of Health Sciences, University
of Molise, Campobasso, Italy (F.A.I., D.V., M.V.S. and M.T.)
JPET Fast Forward. Published on December 29, 2009 as DOI:10.1124/jpet.109.162800
Copyright 2009 by the American Society for Pharmacology and Experimental Therapeutics.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
2
Running head: Kv7 expression and function in skeletal muscle
*Correspondence should be addressed to: Maurizio Taglialatela MD PhD, Department of Health Sciences, University of Molise. Via De Sanctis, 86100 – Campobasso, Italy. Tel. (+39)0874-404851; Fax: (+39)0874-404778. Email: [email protected] Number of text pages: 28 Number of figures: 5 (plus 2 supplemental Figures) Number of Tables: 1 (plus 1 Supplemental Table) Word count:
Abstract: 199 Introduction: 717 Discussion: 1714
Non-standard abbreviations: K+, potassium; [Ca2+]i, intracellular Ca2+ concentration; IKM, M-current; LQTS, long QT syndrome; DFNA2, non-syndromic autosomal dominant hearing loss; BFNS, Benign Familial Neonatal Seizures; GM, growth medium; DM, differentiation medium; MyHC, myosin heavy chain; AnkG, Ankyrin-G; RT, retigabine; MEV, mevastatin; HMG-CoA, 3-hydroxy-3-methylglutaryl coenzyme A; DMEM, Dulbecco’s modified Eagle’s medium; FBS, Fetal Bovine Serum; CHO, Chinese Hamster Ovary; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; PBS, phosphate buffered saline; OD, optical density.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
3
ABSTRACT
Changes in the expression of potassium channels regulate skeletal muscle development. The
purpose of this study has been to investigate the expression profile and phamacological role of Kv7
voltage-gated potassium channels in skeletal muscle differentiation, proliferation, and survival after
myotoxic insults. Transcripts for all Kv7 genes (Kv7.1-5) were detected by PCR and/or real-time
PCR in murine C2C12 myoblasts; Kv7.1, Kv7.3 and Kv7.4 transcripts were up-regulated after
myotube formation. Western-blot experiments confirmed Kv7.2, Kv7.3 and Kv7.4 subunit
expression, and the up-regulation of Kv7.3 and Kv7.4 subunit during in vitro differentiation. In adult
skeletal muscles from mice and humans, Kv7.2 and Kv7.3 immunoreactivity was mainly localized at
the level of intracellular striations positioned between ankyrinG-positive triads, whereas that of
Kv7.4 subunits was largely restricted to the sarcolemmal membrane. In C2C12 cells, retigabine (10
μM), a specific activator of neuronally-expressed Kv7.2-5 subunits, reduced proliferation,
accelerated myogenin expression, and inhibited the myotoxic effect of mevastatin (IC50 ≈ 7 μM); all
these effects of retigabine were prevented by the Kv7 channel blocker XE-991 (10 μM).
Collectively, these data highlight neural Kv7 channels as significant pharmacological targets to
regulate skeletal muscle proliferation, differentiation, and myotoxic effects of drugs.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
4
INTRODUCTION
Skeletal muscle development is a multifactorial, highly controlled process involving the
coordinated regulation of a large number of genes which allow proliferating myoblasts to withdraw
from the cell cycle, to fuse in ordered arrays of large multinucleated myotubes, and to further
differentiate into mature muscle fibers (Walsh and Perlman, 1997).
Changes in the expression of potassium (K+) channels appear to be associated with the myoblast
to myotube transition (Cooper, 2001). Following earlier reports on distinct voltage-sensitive K+
channels being developmentally regulated in vivo (Lesage et al., 1992), K+ currents carried by
ether-à-go-go (Kv10.1) (Bijlenga et al., 1998) and KIR2.1 (Wieland and Gong, 1995; Fischer-
Lougheed et al., 2001) channels have been shown to play a specific role in determining the
membrane hyperpolarization which occurs during the early phases of myoblast commitment and
fusion in vitro. This phenomenon is believed to allow significant Ca2+ influx through T-type
voltage-gated Ca2+ channels, thereby triggering the increase in intracellular Ca2+ concentration
([Ca2+]i) necessary for myoblast fusion (Bijlenga et al., 2000). Other K+ channels, such as
intermediate-conductance Ca2+-activated K+ channels, appear to be down-regulated during
myogenesis, although they do not play a significant role in myoblast proliferation (Fioretti et al.,
2005).
Voltage-dependent K+ channels belonging to the Kv7 subclass have been implicated in resting
membrane potential and electrical excitability control in many cell types (Miceli et al., 2008). The
Kv7 family consists of five members (Kv7.1-Kv7.5), each showing distinct tissue distribution and
subcellular localization, as well as biophysical, pharmacological, and pathophysiological properties.
While Kv7.1 is mainly expressed in cardiac muscle, gastrointestinal epithelia, and inner ear, Kv7.2
and Kv7.3 are expressed in the central and peripheral nervous system (Cooper et al., 2000); on the
other hand, Kv7.4 subunits appear to be mainly expressed in the cochlea and central auditory
pathways (Kharkovets et al., 2000), whereas Kv7.5 transcripts have been detected in several brain
regions and in sympathetic ganglia (Lerche et al., 2000; Schroeder et al., 2000). Since Kv7.2-7.5
were first discovered in neurons, these are currently identified as neural Kv7 genes (Brown and
Passmore, 2009).
Kv7 channels give rise to outwardly rectifying voltage-dependent K+ currents with an
heterogeneous functional role. Kv7.1 subunits form the molecular basis of IKs, a cardiac current
involved in action potential repolarization (Sanguinetti et al., 1996), while Kv7.2 and Kv7.3 subunits
(possibly in association with Kv7.4 and Kv7.5 at specific neuronal sites) can form homomeric or
heteromeric K+ channels underlying the M-current (IKM) (Wang et al., 1998). IKM is a slowly
activating/deactivating, and non-inactivating subthreshold current which regulates neuronal
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
5
excitability, functioning as a brake for repetitive action potential firing (Brown and Adams, 1980).
In humans, mutations in the Kv7.1 gene are responsible for one form of long QT syndrome (LQTS-
1), whereas mutations targeting Kv7.4 cause a rare form of non-syndromic autosomal dominant
hearing loss (DFNA2); gene defects in Kv7.2 or Kv7.3 are responsible for Benign Familial Neonatal
Seizures (BFNS), an inherited epilepsy of the newborn (Miceli et al., 2008; Brown and Passmore,
2009).
In addition to neural tissue, Kv7.4 and Kv7.5 expression has been described also in smooth
muscle cells from several vascular beds, where they control reactivity to vasopressors (Yeung et al.,
2007; Mackie et al., 2008). In addition, Kv7.1 (Tsevi et al., 2005) and Kv7.5 (Lerche et al., 2000;
Schroeder et al., 2000) transcripts have been detected in adult skeletal muscle; changes in Kv7.1 and
Kv7.5 transcript levels occur during proliferation and differentiation of rat L6E9 myoblasts in vitro
(Roura-Ferrer et al., 2008). However, no data is yet available on other Kv7 family members;
moreover, the functional role of Kv7.1 and Kv7.5 in skeletal muscle is still poorly defined.
In this study, using murine C2C12 cells as an experimental model for in vitro skeletal
myogenesis, we have investigated the expression pattern of Kv7.1-5 transcripts and proteins during
skeletal muscle differentiation. Moreover, the expression of Kv7 subunits in adult skeletal muscle
from mouse and humans, as well as their possible role in C2C12 cell proliferation, differentiation,
and protection from drug-induced myotoxicity, has also been investigated. To this aim, channel
blockers (XE-991) and activators (retigabine) of Kv7 channels (Miceli et al., 2008) were used. The
results obtained indicate that neural Kv7 channels regulate skeletal muscle cell proliferation,
differentiation, and response to myotoxic stimuli, thus highlighting novel potential therapeutic
opportunities for Kv7 channel modulators in skeletal muscle disorders including drug-induced
myopathies.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
6
METHODS
CELL CULTURE AND REAGENTS. Murine C2C12 myoblasts were propagated in a growth medium
(GM) composed of Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% Fetal
Bovine Serum (FBS), 50 U/ml penicillin plus 50 μg/ml streptomycin, and 1% L-glutamine (Gibco-
Italia, Milan, Italy), in a humidified atmosphere of 95% air/5% CO2 at 37°C. Exposure of
proliferating C2C12 cells for 24-72 hours to a lower (from 10% to 0.1%) FBS concentration, plus the
addition of 5 μg/ml insulin and 5 μg/ml transferrin (differention medium, DM), induced their
differentation into myotubes, as revealed by phase-contrast light transmission analysis showing an
increased cell size and an elongated fiber-like morphology (Supplemental Fig. 1A, panels a and b).
Confocal immunofluorescence analysis revealed the induction in C2C12 myotubes of myosin heavy
chain (MyHC), a marker of late skeletal muscle cells maturation (Bennett and Tonks, 1997)
(Supplemental Fig. 1A, panels c and d), a result also confirmed by Western-blot analysis
(Supplemental Fig. 1B). Staining with chromomycin A3, a fluorescent DNA-binding antibiotic
labelling cell nuclei, confirmed in vitro myotube formation, as multiple nuclei were evident in a
single fiber (Supplemental Fig. 1A, panels c and d).
Chinese Hamster Ovary cells (CHO cells) were grown in 60mm plastic Petri dishes in DMEM
containing 10% FBS, non-essential amino acids (0.1 mM), penicillin (50 U/ml), and streptomycin
(100 μg/ml) in a humidified atmosphere at 95% O2/5% CO2 at 37°C. After plating, the cells were
transfected on the next day with plasmids encoding Kv7 cDNAs using Lipofectamine 2000
(Invitrogen, Milan, Italy); a plasmid encoding for the Enhanced Green Fluorescent Protein
(Clontech, Palo Alto, CA, USA) was used as a transfection marker; total cDNA in the transfection
mixture was kept constant at 4 μg.
RNA EXTRACTION, SEMIQUANTITATIVE RT-PCR, AND QUANTITATIVE REAL-TIME PCR. Total RNA
was isolated from C2C12 myoblasts and myotubes or from native tissues using the TRI-Reagent
(Sigma-Aldrich, Milan, Italy), reacted with DNAse-I (1U/μl; Sigma-Aldrich, Milan, Italy) for 15
min at room temperature, followed by spectrophotometric quantification. Final preparation of RNA
was considered DNA- and protein-free if the ratio between readings at 260/280 nm was > 1.7.
Isolated mRNA was reverse-transcribed using MuLV high-capacity reverse transcriptase (50 U/μl;
Applied Biosystem, Monza, Italy) in RT buffer containing 4 mM dNTP, RT Random Primers, 1μl
RNAse Inhibitor at 37°C for 120 min; cDNA obtained was amplified in PCR gold buffer, also
containing 25 mM MgCl2, 10 mM dNTP mix, 0.25 mM forward and reverse primers (see
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
7
Supplemental Table 1 for primer sequences), and 5U/μl Amplitaq Gold (Applied Biosystem,
Monza, Italy). The protocol used for PCR amplification was the following: denaturation at 95°C for
1 min, annealing at 54-60°C (template-dependent) for 1 min, elongation at 72°C for 1 min (35
cycles, one every 13 min). To test the ability of the Kv7 primers to specifically recognize the
mRNA target against which they were designed, RT-PCR experiments were performed using
cDNA templates from adult mouse brain and heart mRNAs (Supplemental Fig. 2). Neural Kv7.2-
Kv7.5 transcripts were effectively detected in brain tissue (left panel); a very faint band
corresponding to Kv7.1 mRNA could also be identified, consistent with previous results (Ohya et
al., 2003; Yeung et al., 2007). Moreover, in mouse heart, only Kv7.1 and Kv7.4 transcripts (the latter
of presumed vascular origin) showed significant expression (right panel).
Quantitative real-time PCR was carried out in a 7500 fast real-time PCR system (Applied
Biosystem, Monza, Italy) using the Kv7 primers using SYBR Green detection. Samples were
amplified simultaneously in triplicate in one assay run, and the ct (threshold cycle) value for each
experimental group was determined. Data normalization was performed using as control the ct from
S16, a constitutively-expressed ribosomal protein; differences in mRNA content between groups
were calculated as normalized values using the 2-Δct formula.
CELL PROLIFERATION. To assess C2C12 cell proliferation, myoblasts were trypsinized and seeded
onto 24-well plastic plates at a 2x103 cells/cm2 density. After adhesion, the cells were synchronized
in DM for 30h; cells were then washed twice with phosphate buffered saline (PBS), and 1μCi/ml of
[methyl-3H] thymidine (GE Healthcare, Milan, Italy) was added in fresh GM or DM medium
containing the drug(s) of interest. After 24h, the cells were fixed in cold methanol, washed three
times in ice-cold 10% trichloracetic acid, and solubilised in 1% SDS and 0.3% NaOH (Roura-Ferrer
et al., 2008). The radioactivity in each sample was determined by liquid scintillation counting in a
LS/5000/CE system (Beckman Analytical Instruments, Milan, Italy).
CELL VIABILITY. C2C12 cells were seeded at 2x103 cells/cm2 density in 24-well plastic plates. One
day after plating, mevastatin (alone or in the presence of Kv7 channel modulators) was added to the
culture medium for the indicated times (24, 48 and 72 hours). Cell viability was evaluated with the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; 10 mg/ml; Sigma-Aldrich,
Milan, Italy) reduction assay; formazan salts formation upon MTT reduction by mitochondria of
living cells was detected spectrophotometrically at 595 nm.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
8
CONFOCAL IMMUNOFLUORESCENCE ANALYSIS. Cells. C2C12 cells, plated on poly-L-lysine-coated
coverslips, were washed three times with PBS and incubated at room temperature (20-22°C) with
freshly-made paraformaldehyde (4% w/v) for 10 min. The cells were then incubated for 5 min with
0.1 M glycine, washed in PBS, and incubated for 1 hour with anti-mouse monoclonal anti-MyHC
(dil. 1:500; Sigma, Milan, Italy). Subsequently, the cells were incubated at room temperature for 1h
with a fluorescent secondary antibody (Jackson Laboratories, Bar Harbor, ME, USA) anti-mouse
IgG conjugated to CY3 for anti-MyHC, diluted 1:200 in PBS containing 10% (v/v) fetal bovine
serum (FBS) and 0.1% Triton X-100. Chromomycin A3 was used as nuclear marker. Coverslips
were mounted in SlowFadeTM (Invitrogen – Molecular Probes, San Giuliano Milanese, Italy).
Tissue Slices. Human muscle preparations were obtained from a 30 year-old male’s biopsy of the
brachial biceps, after informed consent approved by the local Ethical Committee was given. Animal
muscle preparations derived from adult male C57/BL6 mice (50-60 g) housed under constant
conditions of temperature (22 ± 1°C) and relative humidity (50%) with a regular light-dark schedule
(lights on from 7 AM to 7 PM) and free access to food and water. All experiments were carried out
in accordance with the European Community Council Directive (86/609/EEC) and were approved
by the Ethics Committees of the Institutions where the experiments were performed. All efforts
were taken to minimize animal suffering and to use the minimum number of animals necessary to
obtain reliable results. Animals were deeply anesthetized, perfused with saline (10 ml) followed by
50 ml of freshly-made paraformaldehyde (4% w/v in PBS). From these animals, soleus muscles
were removed and post-fixed in the same fixative for 2 h at 4°C, and washed with 0.1 M glycine for
5-10 min. Mouse or fixed human tissue was cut with a cryostat into 20 μm thick sections which
were mounted onto pre-treated Superfrost slides, stored at -80°C until further processing. Mouse
and human sections were incubated overnight at 4°C with the following primary antibodies: a)
rabbit-polyclonal anti-Kv7.2 (dil. 1:100; a gift from dr. Holger Lerche, University of Ulm,
Germany) (Luisi et al., 2009); b) rabbit polyclonal anti-Kv7.3 (dil. 1:500; a gift from Dr. Alvaro
Villarroel–CSIC Universidad del Paìs Vasco, Bilbao, ES); c) mouse monoclonal anti Kv7.4 (dil.
1:100; Neuromab, Davis, CA, USA); d) mouse monoclonal anti Ankyrin-G (dil 1:50; Abcam plc,
Cambridge, UK). The primary antibodies were diluted in PBS containing 10% (v/v) fetal bovine
serum (FBS) and 0.1% Triton X-100 and incubated overnight at 4°C. Slices were then washed in
PBS and incubated at room temperature for 1h with anti-mouse IgG conjugated to CY3 for Kv7.4,
or anti-rabbit IgG conjugated to CY3 for Kv7.2 or Kv7.3 (Jackson Laboratories, Bar Harbor, ME,
USA), or anti-mouse IgG conjugated to CY5 for AnkG); secondary antibodies were diluted 1:100 in
PBS containing 10% (v/v) fetal bovine serum (FBS) and 0.1% Triton X-100. Chromomycin A3 was
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
9
used as a nuclear marker. Slices were allowed to air dry before mounting in SlowFadeTM
(Invitrogen – Molecular Probes, San Giuliano Milanese, Italy).
Cells and tissues were analyzed using a Zeiss LSM 510 Meta argon/krypton laser scanning
confocal microscope (Carl Zeiss, Jena, Germany). Images were acquired using the multi-track
system to avoid crosstalk among channels. The excitation/emission settings were as follows:
430/505-550 nm for chromomycin, 543/560-615 nm for CY3, 633/650 nm for CY5. Images were
confocally captured using a 63x oil immersion objective (PlanApochromat, NA = 1.4), with a
maximal confocal zoom factor of 3, fixed box sizes of 512 x 512 pixels, pinhole below 1 Airy unit.
Each image was acquired 4 times and the signal averaged to improve the signal to noise ratio.
WESTERN BLOT. Cells were washed two times in cold PBS and lysed with lysis solution (150 mM
NaCl, 1 mM EDTA pH 8, 10 mM Tris HCl, pH 8, 1% SDS and protease inhibitors). Lysates (100-
150 μg) were boiled 5 min in Laemmli SDS loading buffer and separated by 8% SDS-PAGE. Filters
were incubated overnight at 4°C with following antibodies: (a) rabbit anti-Kv7.2 (1:2000 dilution, dr.
H. Lerche, University of Ulm, Germany) (Luisi et al., 2009); (b) rabbit anti-Kv7.3 (dil. 1:1000; Dr.
Alvaro Villarroel – CSIC Universidad del Paìs Vasco, Bilbao, ES); (c) mouse anti Kv7.4 (dil. 1:500;
Neuromab, Davis, CA-USA); (d) mouse anti-Myosin heavy chain (MyHC) (dil. 1:1000; Sigma,
Milan, Italy). An anti-α-tubulin antibody (Sigma, Milan, Italy) was used to check for equal protein
loading. Reactive bands were detected by chemiluminescence (ECL-plus, Amersham Bioscience, NJ,
USA). Images were analyzed on a ChemiDoc station with Quantity-one software (BioRad, Segrate,
Italy).
MATERIALS. N-(2-amino-4-(4-fluorobenzylamino)-phenyl)-carbamic acid ethyl ester (retigabine)
was from ASTA Medica (Radebeul, Germany) or Valeant Pharmaceuticals (Aliso Viejo, CA,
USA); 10,10-bis(4-pyridinylmethyl)-9(10H)-anthracenone (XE-991) was from Tocris Bioscience
(Bristol, UK). Fura-2 (Calbiochem-Inalco, Milan, Italy) was kept as 1 mM stock solutions in
DMSO at -20°C. All the remaining material was from Sigma Chemicals (Milan, Italy).
STATISTICS. Data are expressed as means±SEM of the given number of experiments (n). Data sets
were compared using matched Student’s t tests or, if necessary, with one-way ANOVA, followed
by the Newman Keul’s test. Statistically significant differences were accepted when p was <0.05.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
10
RESULTS
EXPRESSION OF KV7 K+ CHANNEL TRANSCRIPTS AND PROTEINS DURING C2C12 CELLS
DIFFERENTIATION. To investigate the relative changes in the expression of transcripts encoding for
distinct classes of Na+, Ca2+, and K+ channels in in vitro differentiating C2C12 cells, RT-PCR
experiments were performed using gene-selective primers (Supplemental Table 1). For each gene,
its expression was quantified as relative to that of the constitutive ribosomal protein S16. The data
obtained (shown in Table 1) revealed that, while the mRNAs from most of the ion channel genes
presently tested could not be detected at either myoblast or myotube stage, other mRNAs, such as
those encoding for Kir 2.1, Kir2.2, and Kv10.1, were already expressed at the myoblast stage.
Among Kv7 members, the present RT-PCR results revealed that, in addition to Kv7.1 and Kv7.5
transcripts whose skeletal muscle expression has been previously described (Roura-Ferrer et al.,
2008), those encoding for Kv7.2, Kv7.3, and Kv7.4 were also detected in C2C12 myoblasts cultured
in growth medium (GM) (Table 1 and Fig. 1A).
C2C12 cells incubation for 72 hours in differentiation medium (DM), an experimental
condition promoting myotube formation (Supplemental Fig. 1), induced the novel expression of
transcripts encoding for other ion channels such as Cav1.1, Cav3.1, Nav1.4, Nav1.9, and, among
voltage-gated K+ channels, Kv1.5, Kv3.1, and Kv3.4. In addition, the expression levels of the
majority of ion channel transcripts detected at the myoblast stage did not increase during the
differentiation process, rather showing in some cases a substantial decrease (KCa3.1, Kv10.1). By
contrast, time-course experiments revealed that the abundance of the mRNAs encoding for Kv7.1,
Kv7.3 and Kv7.4, but not for Kv7.2 and Kv7.5, showed a time-dependent increase after 48 hours of
cell exposure to DM, reaching a plateau at 72 hours (Table 1 and Fig. 1A). The fact that Kv7 genes
appear as the only the ion channel subclass whose mRNAs can be detected in C2C12 myoblasts and
whose transcript levels are increased upon myotube formation, is suggestive of their potential role
during skeletal myogenesis.
Since the RT-PCR technique does not allow one to quantify in absolute terms the extent of gene
expression changes, quantitative real time PCR experiments, using a relative-comparative method,
were performed; this technique, by comparing the cycle-dependent increase in the relative
fluorescence of the amplified product, allows one to define, for each experimental condition, a cycle
threshold (ct); this parameter is then normalized to the ct value of a constitutive gene (S16) whose
expression does not change during the experimental procedures, thus defining a Δct value (Fig 1B).
Comparison of Δct values obtained in non-differentiated versus differentiated C2C12 cells (ΔΔct),
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
11
revealed that Kv7.1, Kv7.3 and Kv7.4 mRNA expression was increased 15-, 10-, and 25-fold after 72
hours of myoblasts exposure to DM, respectively; real-time PCR experiments, similarly to semi-
quantitative RT-PCR, did not reveal significant changes in Kv7.2 and Kv7.5 mRNA abundance
during C2C12 cell differentiation in vitro (data not shown).
To evaluate the expression of the proteins encoded by the Kv7.2, Kv7.3, and Kv7.4 mRNAs
herewith described for the first time, Western-blot analysis using subunit-specific antibodies were
performed in cell lysates from C2C12 myoblasts and myotubes (72 hours of DM exposure) (Fig. 2).
Antibody specificity was tested in parallel Western-blot experiments using membrane fractions
from control CHO cells and from CHO cells transfected with the respective Kv7 cDNA. Each anti-
Kv7 antibody revealed a single subunit-specific band only in CHO cells transfected with the
corresponding cDNA; untransfected CHO cells did not show any specific signal. In C2C12 cells
(Fig. 2A, right panels), anti-Kv7.2, -Kv7.3 and -Kv7.4 antibodies recognized a single band of 95, 97
and 77 kDa, respectively; the size of these bands was in accordance with the predicted molecular
weight of the recognized proteins, and identical to those identified in transfected CHO cells.
Moreover, C2C12 cell differentiation significantly increased the intensities of the bands
corresponding to Kv7.3 and Kv7.4, but not that of Kv7.2. Fig. 2B quantifies the pooled results of the
densitometric analysis of Kv7-specific bands in non-differentiated and differentiated C2C12 cells.
These results suggest that, in addition to Kv7.1 and Kv7.5 subunits (Roura-Ferrer et al., 2008),
Kv7.2, Kv7.3, and Kv7.4 subunits are also expressed in C2C12 cells, with Kv7.3, and more so Kv7.4,
being up-regulated during myotube formation in vitro.
EXPRESSION OF KV7.2, KV7.3 AND KV7.4 K+ CHANNEL TRANSCRIPTS AND PROTEINS IN ADULT
SKELETAL MUSCLE FROM MICE AND HUMANS. In mice skeletal muscle tissue, mRNAs encoding for
all Kv7 subunits could be detected by RT-PCR (Fig. 3A). Furthermore, confocal
immunofluorescence experiments on mouse skeletal muscle cryosections revealed that Kv7.2 and
Kv7.3 immunofluorescence was mainly associated with intracellular striations; in addition, Kv7.2,
but not Kv7.3, immunofluorescence also appeared to be expressed along the sarcolemmal
membrane. By contrast, Kv7.4 immunoreactivity appeared to be exclusively localized at the level of
the plasma membrane (Fig. 3B). To gain further insight into the subcellular localization of Kv7.2,
Kv7.3 and Kv7.4 subunits, experiments were performed using antibodies against Ankyrin-G
(AnkG), a scaffold protein localized within the sarcoplasmic reticulum in adult skeletal muscle
fibers (Kordeli et al., 1995; Kordeli et al., 1998); in these experiments, the nuclear marker
chromomycin A3 was also used. In longitudinal sections, AnkG immunofluorescence was
associated with both the sub-plasmalemmal region of the sarcomere and its intracellular striations,
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
12
labelling two bands per sarcomere. Each of these bands is likely to correspond to a triad running on
each side of a Z-line; within each triad, AnkG has been shown to be concentrated along
plasmalemmal T-tubules (Flucher et al., 1990). A similar distribution pattern has been shown for
ryanodine receptors-1 (RyR-1) (Salanova et al., 2002). In the same sections, Kv7.2- and Kv7.3-
positive striations were thin and non-overlapping with those recognized by anti-AnkG antibodies,
being instead positioned between each pair of thick AnkG-labeled triads; these results suggest the
possible expression of Kv7.2 and Kv7.3 subunits in a sarcomeric pattern at the level of the Z-line.
While Kv7.2 and Kv7.3 exhibited intracytoplasmic immunoreactivity, Kv7.4 immunostaining did not
co-localize with AnkG at the cytoplasmic level, being only detected along the plasmalemmal
membrane, as also revealed by the subsarcolemmal position of the myotube nuclei.
In human skeletal muscle, Kv7.2, Kv7.3, and Kv7.4 mRNAs were detected by RT-PCR (Fig.
3C); moreover, an expression pattern similar to that previously described in mouse muscle tissue
was observed with confocal immunohistochemistry (Fig. 3D); while Kv7.2 and Kv7.3
immunoreactivity appeared to be prevalently associated with intracytoplasmic striations,
sarcolemmal labelling was instead revealed by the anti-Kv7.4 antibody.
ACTIVATION OF KV7 CHANNELS REGULATES C2C12 CELL PROLIFERATION AND DIFFERENTIATION IN
VITRO. To investigate the possible functional role of Kv7 channels in skeletal muscle proliferation
and differentiation, C2C12 cells were exposed to retigabine (RT), a compound acting as a selective
activator of neural Kv7 channels, but unable to modulate cardiac Kv7.1 channels (Miceli et al.,
2008), and to XE-991, a Kv7 channel blocker which does not discriminate among channels formed
by different Kv7 subunits (Wang et al., 1998). Proliferation was measured by 3H-thymidine
incorporation, whereas differentiation was quantified by the transcripts levels of myogenin, a
muscle-specific transcription factor required for myotube formation (Bennett and Tonks, 1997). RT
(10 μM) reduced C2C12 myoblasts proliferation to values similar to those observed in cells exposed
to DM. The Kv7 blocker XE-991 (10 μM) failed to interfere with C2C12 cell proliferation, although
it prevented the inhibitory effect exerted by RT (Fig. 4A). In RT-PCR experiments, RT (10 μM)
increased the mRNA expression level for myogenin after 24 and 48 hours of cell exposure to DM,
an effect prevented by XE-991 (10 μM) (Fig. 4B); similar results were also obtained with real-time
PCR experiments (data not shown). These results suggest that the reduced C2C12 myoblasts
proliferation caused by RT may be attributed, at least in part, to an enhanced or anticipated
differentiation of myoblasts into myotubes triggered by the pharmacological activation of neural
Kv7 channels.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
13
KV7 CHANNEL ACTIVATION BY RT PREVENTS STATIN-INDUCED C2C12 CELLS TOXICITY. Skeletal
muscle toxicity is a serious although uncommon side-effect of therapy with 3-hydroxy-3-
methylglutaryl coenzyme A (HMG-CoA) reductase inhibitors of the statin class. Since statin-
induced myotoxicity involves an inhibition of myotube formation in vitro (Baba et al., 2008), an
effect opposite to that prompted by neural Kv7 channel activation in the present experiments, the
consequences of the pharmacological manipulation of Kv7 channels on C2C12 myoblast viability
after exposure to mevastatin (MEV) were evaluated. When proliferating C2C12 myoblasts were
exposed to MEV (0.1-100 μM), a dose- and time-dependent reduction in cell viability was
observed. The IC50 for MEV-induced toxicity was time-dependent, being ~20 μM after 24 hrs, ~6
μM after 48 hours, and ~2 μM after 72 hours of drug exposure (Fig. 5A). When compared to
myoblasts, differentiated C2C12 myotubes appeared considerably less sensitive to statin-induced
toxicity; in fact, the percent of survival after exposure to 3 μM MEV for 48 hours was 57.5±4.4%
(n=8) and 93±6% (n=4) in myoblasts and myotubes, respectively (p<0.05); even more dramatic
differences were observed when MEV concentrations were increased to 100 μM (cell survival was
16±0.5%, n=6, and 75±4%, n=5, in C2C12 myoblasts and myotubes, respectively; p<0.05). The
potential myoprotective actions of Kv7 modulators was assessed upon simultaneous incubation of
proliferating C2C12 cells with RT, XE-991, and RT+XE-991 together with 3 μM MEV for 48 hours,
a value close to the calculated EC50 for the statin. RT (1-100 μM) dose-dependently and fully
protected C2C12 cells from MEV-induced cytotoxicity; the IC50 for RT-induced myoprotection was
~7 μM (Fig. 5B). At each time point (24, 48, and 72 hours), XE-991 (10 μM; Fig. 5C) counteracted
the protective effect of RT on MEV-induced toxicity, strongly suggesting that the Kv7-activating
potential of the drug was responsible for this pharmacological action.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
14
DISCUSSION
EXPRESSION AND MODULATION OF KV7 CHANNEL MEMBERS IN DEVELOPING AND ADULT SKELETAL
MUSCLE. Changes in K+ channels function exert a complex regulation of cell fate, depending on cell
type and differentiation stage, as well as on the specific channel subtype. Indeed, in neurons, an
increased activity of plasma membrane K+ channels is one of the earliest events triggered by
neurodegenerative stimuli; in most of these cases, the inhibition of K+ efflux prevents cell death
(Yu, 2003). Enhanced K+ channel function decreases [K+]i, causing cell shrinkage and cytosolic
acidification, activates some key enzymes involved in apoptosis such as caspases and
endonucleases, and enhances the formation of cytochrome c-dependent apoptosomes (Yu, 2003).
By contrast, in skeletal muscle cells, the early phases of myotube formation are dependent on the
coordinated expression of K+ channels of the delayed-rectifier (Kv10.1) (Bijlenga et al., 1998) and
inward-rectifier (KIR2.1) (Wieland and Gong, 1995; Fischer-Lougheed et al., 2001) families; this
phenomenon leads to a marked hyperpolarization of the cell membrane potential, a necessary
rerequisite for Ca2+-dependent myoblast acquisition of fusion competence (Bijlenga et al., 2000).
In order to identify additional ion channels which might participate in the early phases of
skeletal muscle development, an extensive analysis of the changes in the expression levels of an
array of ion channel transcripts during in vitro C2C12 myoblasts differentiation into myotubes was
carried out in the present study. The results obtained reveal that, in addition to Kv7.1and Kv7.5
transcripts, whose expression in skeletal muscle tissue had been reported previously (Lerche et al.,
2000; Schroeder et al., 2000; Tsevi et al., 2005), Kv7.2, Kv7.3 and Kv7.4 mRNAs are also present in
skeletal muscle cells. Moreover, Kv7.1, Kv7.3, and Kv7.4 transcripts, but not Kv7.2 and Kv7.5, were
up-regulated upon myotube formation, arguing in favour of a potential role for their protein
products during myogenesis. Roura-Ferrer et al. (2008) recently investigated the expression of
Kv7.1 and Kv7.5 transcripts and proteins in differentiating rat L6E9 myoblasts; the present results
confirm that Kv7.5 mRNAs did not increase during myotube formation in vitro, and reveal a
previously un-noticed increase in Kv7.1 mRNA.
The expression of Kv7.2, Kv7.3 and Kv7.4 proteins in C2C12 myoblasts, as well as the up-
regulation of Kv7.3 and Kv7.4 (but not of Kv7.2) in C2C12 myotubes was also confirmed by Western
blot experiments. The subcellular localization of these proteins in adult skeletal muscle from mice
and humans was studied by immunohistochemistry. In longitudinal sections from mouse muscle,
Kv7.2 subunits appeared to be located along the plasmamembrane as well as within the cytoplasm,
at the boundary between sarcomeres, flanked on both sides by AnkG-positive triads; this region
appears to correspond to the Z-line, a multiprotein complex playing an integral role in maintaining
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
15
striated muscle structure and function, and showing a crucial role in the pathogenesis of human
skeletal muscle myopathies (Sheikh et al., 2007). A similar Z-line distribution to Kv7.2 has been
also observed for Kv7.3, whereas Kv7.4 expression seems restricted to the plasmamembrane.
Interestingly, Kv7.2 and Kv7.3 subunits localize in intracellular regions where InsP3R and the
scaffolding protein Homer have been identified (Salanova et al., 2002); although the precise
functional and structural inter-relationships between these Z-line and sarcoplasmic reticulum
proteins is still debated (Volpe et al., 2004), it seems plausible that they may regulate the function
of the sarcoplasmic reticulum, thereby controlling skeletal muscle Ca2+ homeostasis and excitation-
contraction coupling.
FUNCTIONAL ROLE OF KV7 CHANNEL MEMBERS IN SKELETAL MUSCLE PROLIFERATION AND
DIFFERENTIATION. Retigabine (RT) is a novel IKM activator under clinical scrutiny for activity
against hyperexcitability diseases, such as epilepsy, migraine and chronic pain (Miceli et al., 2008).
RT activates with different apparent affinity homomeric or heteromeric channels formed by Kv7
neural subunits, but is unable to potentiate the currents carried by cardiac Kv7.1 channels. RT
enhances neural Kv7 currents by causing an hyperpolarizing shift of the voltage-dependence of
activation and/or by increasing the maximal amount of current elicited by strong depolarizations
(Miceli et al., 2008). In the present study, RT markedly reduced C2C12 myoblast proliferation and
enhanced myotube formation, possibly because the drug enhanced the contribution of Kv7 channels
to C2C12 membrane potential control, thereby rendering more negative the cell membrane potential,
a phenomenon known to facilitate skeletal muscle differentiation and halt cell proliferation
(Wieland and Gong, 1995; Bijlenga et al., 1998; Bijlenga et al., 2000; Fischer-Lougheed et al.,
2001). Noticeably, Kv7.1, Kv7.3, and Kv7.4 transcripts appear to be up-regulated at a differentiation
stage at which Kv10.1 transcripts show a decreased expression (Table 1); the down-regulation of the
Kv10.1 channel results in an hyperpolarization to ~-65 mV of the membrane potential of myoblasts
as a result of the major remaining influence of KIR channels; this would permit an enhanced impact
of subthreshold Kv7 channels under moderately depolarizing conditions.
It should be mentioned that, in addition to activating neural Kv7 channels, RT also affects
other voltage- and ligand-gated channels; however, these pharmacological actions require
concentrations higher than those used in the present study (Rundfeldt and Netzer, 2000). Moreover,
the fact that XE-991 fully counteracted RT-induced effects on C2C12 cell proliferation and
differentiation strongly suggests that neuronal Kv7 channels act as molecular targets for RT. While
the pharmacological activation of neural Kv7 channels by RT effectively interfered with C2C12
myoblast proliferation and differentiation, both phenomena were not affected by the Kv7 channel
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
16
blocker XE-991. This result is apparent contrast with that of Roura-Ferrer et al. (2008) showing that
the IKM blocker linopiridine caused a 60% reduction of C2C12 cell proliferation. However, in this
study, linopiridine was used at a concentration (100 μM) which does not discriminate among
voltage-gated potassium channel subfamilies (Wang et al., 1998); therefore, further experiments are
needed before this pharmacological effect may be attributed to a selective involvement of Kv7
channels.
Several plausible explanations might be given to interpret the inability of XE-991 to
interfere with C2C12 cell proliferation and differentiation in the present experiments. The first is that
Kv7 channels only contribute to a limited amount of outward current, and therefore their blockade
does not significantly influence C2C12 cell membrane potential. The fact that Kv7 channels are not
strongly active at the resting membrane potential, but only become so when activated by RT, has
already been described in peripheral un-myelinated C-type nerve fibers (Lang et al., 2008) as well
as in central myelinated axons (Rivera-Arconada and Lopez-Garcia, 2006; Vervaeke et al., 2006).
Moreover, it should be underlined that XE-991 has been used in the present experiments at 10 μM;
while this concentration causes an almost complete block of homomeric or heteromeric Kv7.2 or
Kv7.3 channels (Wang et al., 1998), Kv7.4 (Sogaard et al., 2001), and even more so Kv7.5 (Yeung et
al., 2008a) channels display a much lower sensitivity to blockade by XE-991. Therefore, the
possibility exists that the described insensitivity to XE-991 of C2C12 cell proliferation and
differentiation might be ascribed to preferential the involvement of Kv7.4 and Kv7.5 subunits.
Although further experiments using antisense (Fischer-Lougheed et al., 2001) or RNA interference
(RNAi) are needed to clarify the role of specific Kv7 subunits, the present results showing a
selective up-regulation of Kv7 mRNAs during C2C12 cell differentiation argue in favour of a
significant role of neural Kv7 subunits in skeletal muscle maturation.
THE PHARMACOLOGICAL ACTIVATION OF NEURAL KV7 CHANNELS PROTECTS SKELETAL MUSCLE
CELLS FROM STATIN-INDUCED TOXICITY. Myopathy involving skeletal muscle weakness, tenderness
and pain, possibly associated to plasma creatine kinase elevation is among the rare but potentially
severe toxicities associated with statins, widely-prescribed lipid-lowering agents acting as HMG-
CoA reductase inhibitors (Sirvent et al., 2008). Despite the widespread concern over this clinically-
relevant side effect, knowledge of its molecular pathogenesis is still a matter of debate; statins have
been reported to affect mitochondrial function, thereby leading to a Ca2+ leak that directly interferes
with the regulation of sarcoplasmic reticulum Ca2+ cycling (Klopstock, 2008; Sirvent et al., 2008).
As a matter of fact, electron microscopy studies in skeletal muscles from statin-treated individuals
have revealed ultrastructural changes mostly located at the T-tubular level, even in the absence of
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
17
clinical and biochemical evidence of skeletal muscle damage (Mohaupt et al., 2009). In
consideration of the close proximity of RT-sensitive Kv7.2, Kv7.3 and Kv7.4 subunits to key sites
for [Ca2+]i handling in adult skeletal muscle fibers, the potential myoprotective effect of RT on
statin-induced C2C12 cell death was investigated. Mevastatin-induced myotoxicity was fully
counteracted by RT in a XE-991-sensitive manner, strongly supporting the hypothesis that Kv7-
opening actions were critically involved in RT-induced myoprotection. The apparent IC50 for RT-
induced reversal of mevastatin-triggered myotoxicity was approximately 7 μM; although this value
is significantly higher than the EC50 reported for RT-induced activation of homomeric Kv7.2 or
Kv7.3, or heteromeric Kv7.2/Kv7.3 or Kv7.3/Kv7.5 channels (0.3-2 μM), it is very close to that
described for Kv7.4 homomeric channels (5 μM) (Tatulian et al., 2001; Yeung et al., 2008b). This
observation suggests that Kv7.4 subunits may be the molecular targets responsible for the
myoprotective actions of RT in the present experimental model. Noticeably, among Kv7 transcripts,
Kv7.4 shows the largest increase following in vitro differentiation of C2C12 cells. The molecular
mechanism(s) underlying RT-induced myoprotection are still under investigation; nevertheless, in
view of the fact that, when compared to myotubes, myoblasts appear significantly more sensitive to
mevastatin-induced myotoxic actions, an important role seems to be attributable to the ability of RT
to promote C2C12 cell differentiation and withdrawal from the cell cycle, an effect opposite to that
prompted by statins in the same cells (Baba et al., 2008).
Collectively, the present results indicate that the pharmacological activation of neural Kv7
subunits regulates skeletal muscle cells proliferation, differentiation, and survival after myotoxic
stimuli, thus revealing a novel role for these channels as potential therapeutic targets in drug-
induced myopathies. Moreover, myoprotective effects triggered by the pharmacological activation
of neural Kv7 subunits may be of clinical relevance also in other skeletal muscle disorders; the RT
structural analogue flupirtine, a non-opioid analgesic which also acts as a neural Kv7 channel
opener (Miceli et al., 2008), appears effective against different types of musculoskeletal pains
(Friedel and Fitton, 1993), and displays significant skeletal muscle-relaxing effects in humans
(Lobish et al., 1996). It remains to be investigated whether, in addition to its effects on neural
sensory information processing, flupirtine-induced opening of Kv7 channels at the skeletal muscle
level is also involved in these pharmacological effects.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
18
ACKNOWLEDGEMENTS
Authors express their gratitude to Prof. Andrea Graziani, University of Piemonte Orientale
A. Avogadro, Novara, Italy, and Prof. Fabio Franciolini, University of Perugia, Perugia, Italy, for
sharing C2C12 cells; T.J. ZMNH Hamburg, Germany, for Kv7.2, Kv7.3, and Kv7.4 cDNAs; Drs.
Holger Lerche and Snezana Maljevic (University of Ulm, Germany) and Dr. Michael Schwake
(Christian-Albrechts-University Kiel, Germany) for their anti-Kv7.2 antibody; Dr. A. Villarroel,
CSIC Universidad del Paìs Vasco, Bilbao, Spain for his anti-Kv7.3 antibody; Prof. Lucio Santoro,
Univ. Federico II, Naples, Italy for human skeletal muscle specimens. The technical help from Dr.
Vincenzo Romaniello, Univ. of Naples Federico II and Dr. Erika Di Zazzo, University of Molise is
kindly acknowledged.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
19
REFERENCES
Baba TT, Nemoto TK, Miyazaki T, and Oida S (2008) Simvastatin suppresses the differentiation of
C2C12 myoblast cells via a Rac pathway. J Muscle Res Cell Motil 29: 127-134.
Bennett AM and Tonks NK (1997) Regulation of distinct stages of skeletal muscle differentiation
by mitogen-activated protein kinases. Science 278: 1288-12891.
Bijlenga P, Occhiodoro T, Liu JH, Bader CR, Bernheim L and Fischer-Lougheed J (1998) An ether
-à-go-go K+ current, Ih-eag, contributes to the hyperpolarization of human fusion-competent
myoblasts. J Physiol 512 ( Pt 2): 317-323.
Bijlenga P, Liu JH, Espinos E, Haenggeli CA, Fischer-Lougheed J, Bader CR and Bernheim L
(2000) T-type alpha 1H Ca2+ channels are involved in Ca2+ signaling during terminal
differentiation (fusion) of human myoblasts. Proc Natl Acad Sci USA 97: 7627-7632.
Brown DA and Adams PR (1980) Muscarinic suppression of a novel voltage-sensitive K+ current in
a vertebrate neurone. Nature 283: 673-676.
Brown DA and Passmore GM (2009) Neural KCNQ (Kv7) channels. Br J Pharmacol 156: 1185-
1195.
Cooper E (2001) A new role for ion channels in myoblast fusion. J Cell Biol 153: 677-686.
Cooper EC, Aldape KD, Abosch A, Barbaro NM, Berger MS, Peacock WS, Jan YN and Jan LY
(2000) Co-localization and co-assembly of two human brain M-type potassium channel
subunits that are mutated in epilepsy. Proc Natl Acad Sci USA 97: 4914-4919.
Fioretti B, Pietrangelo T, Catacuzzeno L and Franciolini F (2005) Intermediate-conductance Ca2+-
activated K+ channel is expressed in C2C12 myoblasts and is downregulated during myogenesis.
Am J Physiol Cell Physiol 289: C89-C96.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
20
Fischer-Lougheed J, Liu JH, Espinos E, Mordasini D, Bader CR, Belin D and Bernheim L (2001)
Human myoblast fusion requires expression of functional inward rectifier Kir2.1 channels. J
Cell Biol 153: F9-F12.
Flucher BE, Morton ME, Froehner SC and Daniels MP (1990) Localization of the alpha 1 and alpha
2 subunits of the dihydropyridine receptor and ankyrin in skeletal muscle triads. Neuron 5: 339-
351.
Friedel HA and Fitton A (1993) Flupirtine. A review of its pharmacological properties, and
therapeutic efficacy in pain states. Drugs 45: 548-569.
Kharkovets T, Hardelin JP, Safieddine S, Schweizer M, El-Amraoui A, Petit C and Jentsch TJ
(2000) KCNQ4, a K+ channel mutated in a form of dominant deafness, is expressed in the inner
ear and the central auditory pathway. Proc Natl Acad Sci USA 97: 4333-4338.
Klopstock T (2008) Drug-induced myopathies. Curr Opin Neurol 21: 590-595.
Kordeli E, Lambert S and Bennett V (1995) AnkyrinG. A new ankyrin gene with neural-specific
isoforms localized at the axonal initial segment and node of Ranvier. J Biol Chem 270: 2352-
2359.
Kordeli E, Ludosky MA, Deprette C, Frappier T and Cartaud J (1998) AnkyrinG is associated with
the postsynaptic membrane and the sarcoplasmic reticulum in the skeletal muscle fiber. J Cell
Sci 111: 2197-2207.
Lang PM, Fleckenstein J, Passmore GM, Brown DA and Grafe P (2008) Retigabine reduces the
excitability of unmyelinated peripheral human axons. Neuropharmacol 54: 1271-1278.
Lerche C, Scherer CR, Seebohm G, Derst C, Wei AD, Busch AE and Steinmeyer K (2000)
Molecular cloning and functional expression of KCNQ5, a potassium channel subunit that may
contribute to neuronal M-current diversity. J Biol Chem 275: 22395-22400.
Lesage F, Attali B, Lazdunski M and Barhanin J (1992) Developmental expression of voltage-
sensitive K+ channels in mouse skeletal muscle and C2C12 cells. FEBS Lett 310: 162-166.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
21
Lobisch M, Schaffler K, Wauschkuhn H and Nickel B (1996) Clinical pilot study of the myogenic
effects of flupirtine in comparison to tetrazepam and placebo. Arzneimittelforschung 46: 293-
298.
Luisi R, Panza E, Barrese V, Iannotti FA, Viggiano D, Secondo A, Canzoniero LM, Martire M,
Annunziato L and Taglialatela M (2009) Activation of pre-synaptic M-type K+ channels
inhibits [3H]D-aspartate release by reducing Ca2+ entry through P/Q-type voltagegated Ca2+
channels. J Neurochem 109: 168-181.
Mackie AR, Brueggemann LI, Henderson KK, Shiels AJ, Cribbs LL, Scrogin KE, and Byron KL
(2008) Vascular KCNQ potassium channels as novel targets for the control of mesenteric artery
constriction by vasopressin, based on studies in single cells, pressurized arteries, and in vivo
measurements of mesenteric vascular resistance. J Pharmacol Exp Ther 325: 475-483.
Miceli F, Soldovieri MV, Martire M and Taglialatela M (2008) Molecular pharmacology and
therapeutic potential of neuronal Kv7-modulating drugs. Curr Opin Pharmacol 8: 65-74.
Mohaupt MG, Karas RH, Babiychuk EB, Sanchez-Freire V, Monastyrskaya K, Iyer L, Hoppeler H,
Breil F and Draeger A (2009) Association between statin-associated myopathy and skeletal
muscle damage. Can Med Assoc J 7: E11-E18.
Ohya S, Sergeant GP, Greenwood IA and Horowitz B (2003) Molecular variants of KCNQ
channels expressed in murine portal vein myocytes: a role in delayed rectifier current. Circ Res
92: 1016-1023.
Rivera-Arconada I and Lopez-Garcia GA (2006) Retigabine-induced population primary afferent
hyperpolarisation in vitro. Neuropharmacol 51: 756–763.
Roura-Ferrer M, Solé L, Martínez-Mármol R, Villalonga N and Felipe A (2008) Skeletal muscle
Kv7 (KCNQ) channels in myoblast differentiation and proliferation. Biochem Biophys Res
Commun 369: 1094-1097.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
22
Rundfeldt C and Netzer R (2000) Investigations into the mechanism of action of the new
anticonvulsant retigabine. Interaction with GABAergic and glutamatergic neurotransmission
and with voltage gated ion channels. Arzneimittelforschung 50: 1063-1070.
Salanova M, Priori G, Barone V, Intravaia E, Flucher B, Ciruela F, McIlhinney RA, Parys JB,
Mikoshiba K and Sorrentino V (2002) Homer proteins and InsP(3) receptors co-localise in the
longitudinal sarcoplasmic reticulum of skeletal muscle fibres. Cell Calcium 32: 193-200.
Sanguinetti MC, Curran ME, Zou A, Shen J, Spector PS, Atkinson DL and Keating MT (1996)
Coassembly of K(V)LQT1 and minK (IsK) proteins to form cardiac I(Ks) potassium channel.
Nature 384: 80-83.
Schroeder BC, Hechenberger M, Weinreich F, Kubisch C and Jentsch TJ (2000) KCNQ5, a novel
potassium channel broadly expressed in brain, mediates M-type currents. J Biol Chem 275:
24089-24095.
Sheikh F, Bang ML, Lange S and Chen J (2007) "Z"eroing in on the role of Cypher in striated
muscle function, signaling, and human disease. Trends Cardiovasc Med 17: 258-262.
Sirvent P, Mercier J and Lacampagne A (2008) New insights into mechanisms of statin-associated
myotoxicity. Curr Opin Pharmacol 8: 333-338.
Søgaard R, Ljungstrøm T, Pedersen KA, Olesen SP and Jensen BS (2001) KCNQ4 channels
expressed in mammalian cells: functional characteristics and pharmacology. Am J Physiol Cell
Physiol 280: C859-C866
Tatulian L, Delmas P, Abogadie FC and Brown, DA (2001) Activation of expressed KCNQ
potassium currents and native neuronal M-type potassium currents by the anti-convulsant drug
retigabine. J Neurosci 21, 5535-5545.
Tsevi I, Vicente R, Grande M, López-Iglesias C, Figueras A, Capellà G, Condom E and Felipe A
(2005) KCNQ1/KCNE1 channels during germ-cell differentiation in the rat: expression
associated with testis pathologies. J Cell Physiol 202: 400-410.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
23
Vervaeke K, Gu N, Agdestein C, Hu H and Storm JF (2006) Kv7/KCNQ/M-channels in rat
glutamatergic hippocampal axons and their role in regulation of excitability and transmitter
release. J Physiol 576: 235–256.
Volpe P, Sandri C, Bortoloso E, Valle G and Nori A (2004) Topology of Homer 1c and Homer 1a
in C2C12 myotubes and transgenic skeletal muscle fibers. Biochem Biophys Res Commun 316:
884-892.
Walsh K and Perlman H (1997) Cell cycle exit upon myogenic differentiation. Curr Opin Genet
Dev 7: 597-602.
Wang HS, Pan Z, Shi W, Brown BS, Wymore RS, Cohen IS, Dixon JE and McKinnon D (1998)
KCNQ2 and KCNQ3 potassium channel subunits: molecular correlates of the M-channel.
Science 282: 1890-1893.
Wieland SJ and Gong QH (1995) Modulation of a potassium conductance in developing skeletal
muscle. Am J Physiol 268 (2 Pt 1): C490-C495.
Yeung SY, Pucovský V, Moffatt JD, Saldanha L, Schwake M, Ohya S and Greenwood IA (2007)
Molecular expression and pharmacological identification of a role for K(v)7 channels in murine
vascular reactivity. Br J Pharmacol 151: 758-770.
Yeung SY, Lange W, Schwake M and Greenwood IA (2008a) Expression profile and
characterisation of a truncated KCNQ5 splice variant. Biochem Biophys Res Comm 371: 741–
746.
Yeung SY, Schwake M, Pucovský V and Greenwood IA (2008b) Bimodal effects of the Kv7
channel activator retigabine on vascular K+ currents. Br J Pharmacol 155: 62-72.
Yu SP (2003) Regulation and critical role of potassium homeostasis in apoptosis. Prog Neurobiol
70: 363-386.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
24
FOOTNOTES
This study was supported by: E-Rare 2007 (EUROBFNS); Telethon (GGP07125); Regione Molise;
and Italian Ministry of University, Education, and Research (MIUR, PRIN 2007) to MT.
Reprint requests: Maurizio Taglialatela MD PhD, Department of Health Sciences, University of
Molise. Via De Sanctis, 86100 – Campobasso, Italy ([email protected]; [email protected])
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
25
LEGENDS FOR FIGURES
Figure 1. Expression of Kv7 transcripts during C2C12 cells differentiation. (A) Time-course of Kv7
mRNA expression during C2C12 cell in vitro differentiation. C2C12 myoblasts were cultured in GM or
exposed to DM for 24, 48, and 72 hours; total mRNA was extracted, retrotranscribed, and the
resulting cDNA amplified with Kv7-selective primers. For each gene, the top panels show agarose
gel electrophoresis of the RT-PCR products at each time point (MW of the standards is shown on
the left), whereas the quantification of the OD relative to that of the reference gene S16 are reported
in the lower panels. Each point is the Mean ± S.E.M. of 3 separate determinations. *=p<0.05 vs
C2C12 myoblasts (GM). Amplicon size was: 143 bp for Kv7.1; 490 bp for Kv7.2; 238 bp for Kv7.3;
100 bp for Kv7.4; 180 bp for Kv7.5. (B) Quantification of transcripts for Kv7 members using
quantitative real-time PCR. Data are expressed as 2-Δct relative to S16, as described in the Methods
section. Empty columns: myoblasts (GM); filled columns: myotubes (DM for 72 hours). Each bar is
the Mean ± S.E.M. of at least 4 separate determinations. *=p<0.05 vs myoblasts.
Figure 2. Expression of Kv7 subunits during C2C12 cells differentiation. (A) Western-blots on total
cell lysates from CHO (left panels) and C2C12 cells (right panels); Kv7 subunits were detected using
anti-Kv7-specific antibodies. C2C12 myoblasts were grown in GM, whereas myotubes were obtained
upon C2C12 cell exposure to DM for 72 hours. The panels at the bottom of each image show the
expression, on the same filters, of the intracellular protein α-tubulin, used as an internal standard for
equal protein loading. (B) Quantification of the data shown in panel A. For each experiment, the
optical density (OD) of the Kv7.x bands was obtained by densitometric analysis; this value was
divided by that of each respective α-tubulin band intensity. Data are expressed after normalization
to the ratio OD Kv7.x/OD α-tubulin in myoblasts. Empty columns: myoblasts (GM); filled columns:
myotubes (DM for 72 hours). Each data point is the Mean±S.E.M calculated from 4 separate
experiments. Asterisks denote values significantly different (p<0.05) from C2C12 myoblasts.
Figure 3. Expression of Kv7 genes and proteins in adult skeletal muscle from mice and humans.
(A) Agarose gel electrophoresis of RT-PCR products obtained from cDNA amplification of mouse
muscle mRNA using Kv7-selective primers (Supplementary Table 1). Amplicon sizes are reported
in the Legend for Fig. 1. (B) Confocal immunofluorescence images of mouse soleus muscle
longitudinal cryosections. Red: Kv7 immnoreactivity (top panels: Kv7.2; middle panels: Kv7.3,
lower panels: Kv7.4); green: AnkG immunoreactivity; blue: nuclei identified with chromomycin
A3. The scale bar is 10 μm for Kv7.2; 5 μm for Kv7.3; and 5 μm for Kv7.4. The insert in the panels
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
26
corresponding to Kv7.3 have been enlarged with a x3 magnification. (C) Agarose gel
electrophoresis of RT-PCR products obtained from cDNA amplification of human brachial biceps
mRNA using Kv7-selective primers. Amplicon size was: 115 bp for Kv7.2; 121 bp for Kv7.3;192 bp
for Kv7.4. (D) Confocal immunofluorescence images of human brachial biceps muscle cryosections.
Kv7 immunoreactivity is shown in red, and nuclei are identified in green with chromomycin A3.
The scale bar is 5 μm for Kv7.2; 5 μm for Kv7.3; and 10 μm for Kv7.4.
Figure 4. Effect of the pharmacological activation of Kv7 channels on C2C12 cell proliferation
and differentiation in vitro. (A) For proliferation assays, C2C12 myoblasts growth was synchronized
by cell exposure to DM for 30-36 hours; after this period, cell proliferation was assessed by [3H]-
thymidine incorporation in cells grown for further 24 hours in DM (empty bar) or in GM (with or
without the indicated drugs, filled bars). RT or XE-991 were each used at 10 μM. Each point is the
Mean ± S.E.M. of 3-4 separate determinations performed in quadruplicate. *=p<0.05 vs GM
controls (no drug added). (B) C2C12 cell differentiation was evaluated by measuring myogenin
mRNA expression with RT-PCR during cell exposure to DM for the indicated times. When
necessary, drugs were added at time 0. Myogenin expression was quantified densitometrically and
ODs were expressed relative to the reference gene S16. Each point is the Mean ± S.E.M. of 3
separate determinations performed in triplicate. *=p<0.05 vs C2C12 cells grown in DM (no drug
added).
Figure 5. Effect of Kv7 channel modulators on mevastatin-induced cytotoxicity in C2C12 cells. (A)
Dose- and time-dependent toxicity exerted by mevastatin in C2C12 myoblasts. Mevastatin (MEV;
0.1-100 μM) was added to GM of proliferating C2C12 cells, and cell toxicity was assessed by the
MTT reduction assay 24 (filled squares), 48 (filled circles), or 72 hours (filled inverted triangles)
later. Data are expressed as OD at 595 nm, normalized to controls. In this and following panels,
each point is the Mean ± S.E.M. of 3 experiments each performed in quadruplicate. The solid lines
represent the fits of the experimental data to the following binding isotherm : y=max/(1+X/IC50),
where X is the MEV concentration. Fitted values for IC50 were ~20 μM after 24 hrs of exposure, ~6
μM after 48 hours, and ~2 μM after 72 hours of MEV incubation. (B) Reversal of MEV-induced
toxicity by retigabine. C2C12 myoblasts were exposed to 3 μM MEV for 48 hours in the presence of
1-100 μM RT. The solid line represents the fit of the experimental data to the described binding
isotherm. (C) XE-991 prevents RT-induced protection from MEV toxicity. Proliferating C2C12 cells
were exposed to GM or GM added with MEV (3 μM), RT (10 μM), XE-991 (10 μM), MEV (3
μM)+RT (10 μM), or MEV (3 μM)+RT (10 μM)+XE-991 (10 μM), as indicated, for 24, 48, and 72
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
27
hours. At the end of these periods, mitochondrial function was evaluated by the MTT reduction
assay. *=p<0.05 vs respective control (GM with no drug added).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #162800
28
Cav1.1 -
Kv1.5 - +
Cav1.2 - -
Kv1.7 - -
Cav1.3 - -
Kv2.1 - -
Cav3.1 - +
Kv2.2 - -
Cav3.2 - -
Kv3.1 - +
Cav3.3 - -
Kv3.4 - +
Nav1.4 - +
Kv4.1 - -
Nav1.5 + +
KCa
1.1 + +
Nav1.9 - +
KCa
3.1 + -
Kir2.1 + +
Kir2.2 + +
KCa
2.2 + +
KCa
2.3 + +
Kv1.1 - -
Kv1.4 - -
Gene Myoblasts Myotubes
+
Kv11.1 - -K
v11.1 - -
Kv10.1 +
Kv10.2 - -
Kv7.1 +
Kv7.2 + +
Kv7.3 +
Kv7.4 +
Kv7.5 + +
Table 1. Ion channel genes expressed in C2C12 myoblasts and myotubes.
- = not detected+ = detected- down-regulated in myotubes
up-regulated in myotubes
Myoblasts MyotubesGene
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

Fig. 1
mRNA expression (2-
Δct
)
B
A
Kv7.1 K
v7.3 K
v7.4
0
0,01
0,02
0,03
0,04
0,05
0,06 myoblasts
myotubes
*
*
*
S16
(bp)
100
200300
Kv7.1 K
v7.2
500
400
Kv7.3 K
v7.4 K
v7.5
1
2
GM 24 48 72 mRNA expression
(OD Kv7.1/OD S16)
* *
GM 24 48 72
time (h) in DM
0,5
1,0
1,5
mRNA expression
(normalized OD)
1
2
3
GM 24 48 72 mRNA expression
(OD Kv7.2/OD S16)
1
2
GM 24 48 72
* *
mRNA expression
(OD Kv7.3/OD S16)
1
2
GM 24 48 72
* *
mRNA expression
(OD Kv7.4/OD S16)
1
2
GM 24 48 72 mRNA expression
(OD Kv7.5/OD S16)
100
200300400 700
600
100
200300400
100
200300400
100
200300400
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

Fig. 2
Kv7.2
Kv7.3
Kv7.4
α-tubulin
MW(kDa)
77
97
95
55
CHO/Kv7.2
A
B
myoblasts
myotubes
Kv7.x protein expression
(normalized to myoblasts)
0
1
2
3
Kv7.2 K
v7.4K
v7.3
*
*
CHO
CHO/
Kv7.3
CHO/
Kv7.4
CHO
CHO
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from
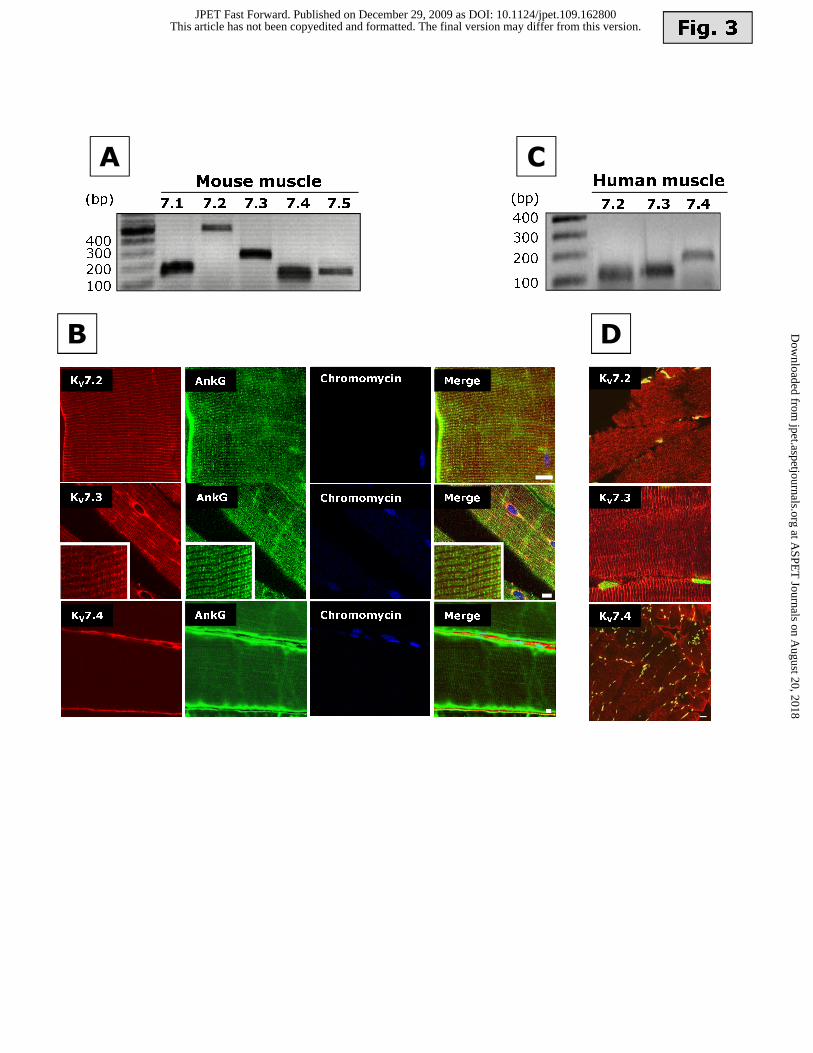
Fig. 3
A C
(bp)
Mouse muscle
7.57.1 7.2 7.3 7.4
100
200300400
KV7.3
KV7.2
KV7.4
DB
AnkG MergeChromomycin
MergeAnkG Chromomycin
AnkG Chromomycin
KV7.3
KV7.2
MergeKV7.4
Human muscle
100
200
300
400
(bp)7.2 7.3 7.4
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

Fig. 4
A
B
XE-991 RT RT+
XE-991
0.1% FBS (DM)
*
*
10% FBS (GM)
Myogenin mRNA expression
(OD myogenin/OD S16)
Control (DM)
RT
RT + XE-991
XE-991
Time (h)
0
0,5
1
1,5
24 48 720
*
*
50
100
Proliferating
C2
C1
2
myoblasts
(%)
Control
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from

Fig. 5
A B
Reversal of MEV-
induced toxicity (%)
C
Cell viability
(OD at 595 nm)
0
0,2
24 h 48 h
GM
GM+MEV+RT+XE-991
GM+MEV
GM+RT GM+MEV+RT
GM+XE-991
72 h
*
Mevastatin (μM)
Cell viability
(normalized OD at 595 nm)
0,1 1 10 100
0,0
0,5
1,0
24h
48h
72h
1 10 100
Retigabine (μM)
0
50
100
**
*
*
*
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on December 29, 2009 as DOI: 10.1124/jpet.109.162800
at ASPE
T Journals on A
ugust 20, 2018jpet.aspetjournals.org
Dow
nloaded from