downloaded from jpet.aspetjournals.org at aspet journals...
TRANSCRIPT

JPET #104620 1
Mast cell stabilizer Ketotifen prevents mucosal mast cell hyperplasia and intestinal
dysmotility in experimental T. spiralis inflammation in the rat
Serna H, Porras M and Vergara P
Department of Cell Biology, Physiology and Immunology, Universitat Autònoma de
Barcelona, Bellaterra, Spain (HS, MP & PV)
JPET Fast Forward. Published on September 20, 2006 as DOI:10.1124/jpet.106.104620
Copyright 2006 by the American Society for Pharmacology and Experimental Therapeutics.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 2
Running title: ketotifen prevents intestinal dismotility
Corresponding author:
Patri Vergara
Unidad de Fisiologia
Facultad de Veterinaria
Universitat Autònoma de Barcelona
08193 Bellaterra
Spain
Telephone: +34935811848
Fax: +34935812006
e-mail: [email protected]
Number of text pages: 16
number of tables: 2
number of figures: 8
number of references: 46
number of words in the Abstract: 237
number of words Introduction: 432
number of words Discussion: 1,252
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 3
Non standard abbreviations:
IBS: irritable bowel syndrome
NGF : Nerve Growth Factor
RMCPII : Rat mast cell protease II
VCU: Villus-crypt unit
MPO: myeloperoxidase
iNOS: inducible nitric oxide synthase
COX-2: Cyclooxigenase-2
Section :
• Gastrointestinal, Hepatic, Pulmonary, and Renal
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 4
ABSTRACT
Trichinella spiralis infection in rats induces hypermotility and an abnormal response to
cholecystokinin (CCK) similar to motor disturbances observed in irritable bowel
syndrome (IBS). Mast cell hyperplasia is also characteristic of this experimental model.
The aim of our study was to correlate mast cell activity with the development of
dysmotility and to demonstrate if the mast cell stabilizer ketotifen could prevent the
development of intestine hypermotility. Sprague-Dawley rats were infected with
Trichinella spiralis and 5 days after infection treated with the mast-cell stabilizer
Ketotifen (10 mg . Kg -1 . día – 1). 12 days after infection, intestinal spontaneous motor
activity and response to CCK were evaluated by means of strain-gauge transducers.
Immunohistochemistry for rat mast cell protease II (RMCP II), COX-2 and iNOS was
performed in intestinal specimens. In addition, RMCPII and myeloperoxidase were
determined in serum. Infected control rats showed hypermotility, mast cell hyperplasia,
increased RMCPII levels, increased MPO and overexpression of COX-2 and iNOS. In
contrast, ketotifen treated rats showed spontaneous intestinal motility and CCK
response similar to the non-infected control rats. Mast cell hyperplasia and RMCPII
were reduced in ketotifen treated rats. Inflammatory parameters were less modified by
ketotifen but those animals that received the longest ketotifen treatment showed a slight
amelioration in these parameters. These results indicate that mast cells are implicated in
the development of hypermotility. The treatment with ketotifen prevented hypermotility
and mast cell hyperplasia and diminished mucosal mast cell activity.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 5
INTRODUCTION
Irritable bowel syndrome (IBS) is a functional, multifactorial disease characterized by
exacerbated responses to gastrointestinal motor reflexes. Both motor changes and
enhanced perception of stimuli arising from the gut are thought to be major contributors
for symptom generation (Chey et al., 2001). Causes of IBS are not well defined;
however, at least in some cases it has been associated with a previous mucosa invasive
gastrointestinal infection (Parry and Forgacs, 2005).
Mast cells are key cells in the response of the intestine to infection and inflammation
(He, 2004) as well as in food allergy and other immune responses (Galli et al., 2005).
Furthermore, mast cell activation and release of mast cell mediators have been
associated with IBS (Barbara et al., 2004).
The experimental Trichinella spiralis infection is a widely used model of experimental
intestinal inflammation and post infectious IBS (Torrents and Vergara, 2000; Bercik et
al., 2004; Weatcroft et al., 2005). Larvae of the parasite invade the duodenum and
jejunum mucosa causing a severe inflammation that spontaneously reverts to normality
in approximately 2-3 weeks. In the meantime, there is a strong motor and secretory
reaction that promotes worm expulsion and restoration of health (Palmer et al., 1984;
Wank et al., 1991). These changes are concomitant with a severe mast cell hyperplasia
that also decreases spontaneously after worm expulsion (Ruitenberg, 1979).
Motor changes due to parasite infection affect all motor patterns from fasting motility to
motor response to postprandial hormone CCK (Palmer et al., 1984; Cowles and Sarna,
1991; Torrents and Vergara, 2000; Gay et al., 2001). A close association between mast
cells and vagal afferents has been well established (Williams, 1997). Furthermore, the
exacerbated motor responses during T. spiralis infection have been associated with
afferent hypersensitivity (Torrents et al., 2002).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 6
Ketotifen is a drug extensively used to prevent mast cell activation (Eliakim et al., 1995;
Crampton et al., 2003; Poutoulakis et al., 1993). We previously demonstrated that
ketotifen stabilized intestinal mucosal mast cells in the rat and prevented mucosal mast
cell stimulation (Juanola et al., 1998).
The objective of this study was to demonstrate if the treatment with mast cell stabilizer
ketotifen could prevent the development of intestine hypermotility during T. spiralis
infection. The parameters that have been evaluated in this study are: 1) in vivo motor
activity of the intestine by measuring spontaneous activity and the response to CCK; 2)
number of mast cells in the intestinal mucosa; 3) activity of mucosal mast cells by
monitoring rat mast cell protease II in serum; and 4) evaluation of inflammation by
measurement of MPO in serum and iNOS and COX-2 immunoreactivity in the intestine.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 7
MATERIALS AND METHODS
Animals
Male SD rats (Charles River, France), 8 to 10 weeks old and weighing 300-350 g were
used in this study. Animals were kept at controlled temperature (20 21 °C), humidity
(55-60%) and photoperiod (12:12 h) room. Animals were caged in groups of three and
had free access to water and a commercial pellet food. At the initiation of ketotifen
treatment or after larvae infection, all the animals were caged individually. All
experimental protocols as well as housing and handling of animals were carried out
under the supervision and regulations of the ethical committee of the Universitat
Autonoma de Barcelona.
Trichinella spiralis infection
Rats were infected by administering 1.0 ml of 0.9 % saline solution containing 7,500
T.spiralis larvae by gavage. The larvae were obtained from female CD1 mice infected
30-90 days before as previously described (Castro and Fairbairn 1969; Torrents and
Vergara, 2000).
Drugs and Substances. Mast cell membrane stabilizer Ketotifen Fumarate Salt (4-(1-
Methyl-4-piperidylidene)-4H-benzo[4,5]cyclohepta[1,2-b]thiophen-10(9H)-one
fumarate; Sigma Chemicals, St. Louis, USA) was given to each individually caged rat
in drinking water as previously described (Juanola et al., 1998). Ketotifen was dissolved
in drinking water at a concentration of 0.1 mg . ml-1 which allowed to dose ketofiten at
10 mg . kg 1 . day 1. The amount of water drunk by each rat was controlled daily. If a
rat was found to ingest less than the required dose of ketotifen (by drinking less than 30
ml of water) it was rejected for the study. CCK-8, sulfated form (Peptide Institute,
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 8
Osaka, Japan), was diluted in 1% sodium bicarbonate to 10 4 M and in buffered saline
solution to work concentration.
Experimental groups. For this study rats were divided in the following groups: 1) Non-
infected control group, rats that did not receive either parasite larvae or treatment
(n = 6); 2) ketotifen control group, non-infected rats treated with ketotifen during 7 days
(n = 4); 3) infected control group, rats infected with T. spiralis (n = 7); 4) infected
ketotifen 5 group, rats infected with T. spiralis and treated from day 5 to 10 post
infection (PI) with ketotifen, (n = 6); and 5) infected ketotifen 7 group, rats infected
with T. spiralis and treated with ketotifen from day 5 until the day of the experiment
(day 12 PI) (n = 5). These two ketotifen groups differ in the time that ketotifen
treatment is stopped. In ketotifen 5 group, the mast cell stabilizer was removed 48 h
before conducting the experiment. In contrast, ketotifen 7 group received ketotifen until
the same day of the experiment. In addition, some infected rats were treated with
ketotifen during 48 h before the experiment (n=3) or received by garvage the daily dose
of ketotifen 1 hour before the experiment (n=2). The objective of these last two
protocols was to be able to differentiate the long-term effect from possible acute effects
of ketotifen. As it is shown in the results section, results from ketotifen 5 and 7 groups
were practically similar and the short time treatment with ketotifen did not modify
motor responses in infected rats. These results indicate that the effect observed in the
animals treated with ketotifen is due to the treatment and not to the direct effect of
ketotifen during the experiment. All motility experiments and the samples were taken
on day 12 day PI. The reason for choosing this time is that, coinciding with parasite
expulsion, there is maximum hypermotility and mastocitosis (Woodbury et al., 1984).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 9
Animal preparation for intestinal motility studies. The night before the experiment,
food was restricted to 10 g. We had previously checked that this procedure, that
amillorated stress due to food deprivation, guaranteed a fasting time of at least 6 h and
the completely emptiness of the stomach at the moment experiment was initiated.
Anesthesia was induced by inhalation of halothane to allow cannulation with a
polyethylene tubing of the right jugular vein. Level III of anesthesia was maintained
with thiopental sodium bolus infusion in the jugular as required. Body temperature was
maintained at 37°C by placing the rat on a heating pad. A tracheotomy was practiced to
facilitate spontaneous breathing. The abdomen was opened through a midline incision,
and the intestine was exposed. Three strain gauges (3 × 5 mm; Hugo Sachs Elektronik,
Hugstetten, Germany) were placed to record circular muscle activity and sutured to the
intestinal wall of the duodenum, proximal jejunum, and ileum, respectively. Strain-
gauges were connected to high-gain amplifiers (MT8P; Lectromed.Ltd, Letchworth,
Herts, UK), and amplified signals were sent to a recording unit (PowerLab/800;
ADInstruments Pty Ltd., Castle Hill, Australia) connected to a PC running PowerLab
software.
Motor activity evaluation. After an equilibration period of 20 min, spontaneous motor
activity during 50 min was recorded and the number of contractions during the time of
recording counted. Afterwards, CCK-8 (3 × 10 9 mol . kg 1 . 10 min 1) was i.v. infused
in bolus during 10 min and the area under the curve (AUC) described by the response
(phasic and tonic response) measured.
Histological study. After finishing the experimental protocol, samples of duodenum,
jejunum, and ileum, were obtained, fixed for 48 h in neutral buffered formalin,
embedded in paraffin, cut into 5-µm sections, and stained with hematoxylin-eosin. A
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 10
scoring based on the inflammatory cell infiltration was used to evaluate the
inflammatory process. At the same time, thickness of intestinal muscular layers was
measured with a scored microscope. At least four different measures were taken from
any sample, and samples from at least four animals of each group were used for the
evaluation of the muscle thickness.
Mucosal mast cell identification. Immunodetection of RMCP II was carried out on
paraformaldehyde-fixed sections using a monoclonal antibody (1:500; Moredun Animal
Health, Edinburgh, UK). Detection was performed with avidin/peroxidase (Vectastain
ABC kit; Vector Laboratories, Burlingame, CA, USA). Sections were counterstained
with haematoxylin and counted at 400 magnification. Positively stained mast cells
were counted in three to five sections per animal. Seven to 10 well-oriented villus-crypt
units (VCU) were examined per section. Analysis of all morphological data was
performed blinded to prevent observer bias.
Immunohistochemistry of iNOS and COX-2. Immunohistochemistry of iNOS and
COX-2 was carried out on paraformaldehyde-fixed sections using either an anti-iNOS
antibody (1:100; Neo-markers, Fremont, CA, USA) or anti-COX-2 antibody (1:100;
Santa Cruz Biotechnology, Santa Cruz, CA, USA). Detection was performed with
avidin/peroxidase (Vectastain ABC kit; Vector Laboratories, Burlingame, CA, USA),
and sections were counterstained with haematoxylin.
Measurement of RMCP II and mieloperoxidase (MPO). Serum samples were taken
in all rats: at time 0, before infection; at 5 days PI, just before the beginning of ketotifen
treatment and at 12 days PI at the time of motor activity evaluation. RMCPII and MPO
concentration in the serum was measured by enzyme-linked immunosorbent assay
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 11
(ELISA) using commercial kits (RMCPII; Moredun Animal Health, Edinburgh, UK,
and MPO; HyCult biotechnology, Uden, The Netherlands).
Data Analysis One-way analysis of variance (ANOVA) followed by a post hoc
Bonferroni test was used to compare motor parameters as well as mast cell number and
intestinal wall thickness. RMCPII and MPO concentration results were compared by
repeated measures ANOVA analysis. Differences between groups were considered
statistically significant when P < 0.05. All data are expressed as mean ± SEM.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 12
RESULTS
Intestinal motor activity
In the non-infected control group spontaneous activity of the intestine consisted of
single contractions at a variable frequency of 1 to 2 contractions every 10 minutes. In
contrast, this pattern of isolated contractions was replaced in infected rats by an
irregular pattern with strong groups of contractions (clusters) that alternated with
periods of inactivity with a variable frequency of 3-4 every 10 minutes or by a
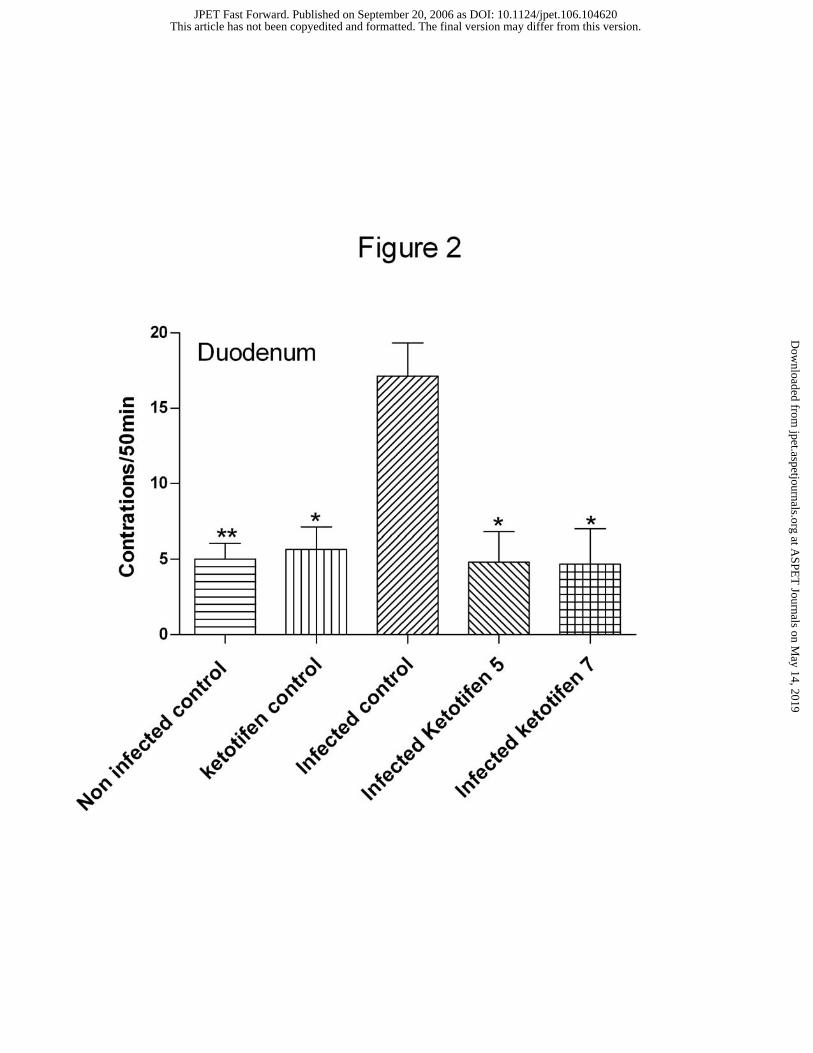
continous irregular activity as shown in Fig 1B. In consequence, the total number of
contractions in duodenum and jejunum was significantly higher in the parasite infected
rats compared to the non-infected control group (Fig. 2). Treatment with ketotifen either
during 5 or 7 days in infected rats prevented cluster contractions or hypermotility to
appear (Fig. 1), being spontaneous motility in form and frequency of contractions in
these animals similar to those of the non-infected control group (Fig. 2). Ketotifen in
non-infected animals did not induce any motor change.
Similar changes were observed in the response to CCK-8 infusion (3x10-9 mols .Kg-1 .
10 min). CCK induced a contractile response at the duodenum and an inhibitory
response in the jejunum of non-infected control rats. In infected control rats, CCK
induced contraction in the duodenum was of a greater magnitude (1368±53.9 AUC
compared to 598.4±53.8 AUC in controls), simultaneous with a contractile response of
the jejunum that completely overlaped the inhibition observed in non-infected rats.
Ketotifen, given either during 5 or 7 days in infected rats, reduced contractile response
in the duodenum (777.5±83.4 and 575.5±201.2 in ketotifen 5 and ketotifen 7 group
respectively). However, the most significant result was that ketotifen in both groups
abolished the contractile response in the jejunum and restored the inhibitory response
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 13
observed in non-infected control rats (Fig. 3). A summary of CCK response in the
jejunum in all groups is given in Fig. 4. Ketotifen in non-infected animals did not
modify CCK response.
RMCPII in serum
RMCPII was measured in serum as an indication of mast cell activity. In non-infected
control rats RMCPII concentration was low at day 0 (246±33.6 ng . ml-1) and remained
low during the whole period of study. In contrast, RMCPII concentration significantly
increased in all T. spiralis infected rats. This increase was about 6 fold by day 5 PI
(814.1±126.3 ng . ml-1) and reached 7,438±2,136 ng . ml-1 at day 12 PI. Both ketotifen
treatments reduced RMCPII concentration increase in infected animals and RMCPII
values were in ketotifen treated animals no significantly different from those in serum
of non-infected control rats (Fig. 5).
RMCP II immunohistochemistry and mucosal mast cell count
Mast cells were stained by the RMCPII antibody in the intestinal mucosa. Number of
mast cells in non-infected control rats was of 5-7 cells per VCU in the duodenum and
jejunum. Infection with T. spiralis larvae induced a clear mast cell hyperplasia (Fig. 6)
(50-60 mast cells per villus-crypt unit in the duodenum and 40-45 cells per VCU in the
jejunum). Ketotifen treatment in infected animals significantly reduced the number of
mast cells stained by RMCPII compared to infected control group. These results
indicate that ketotifen reduced mast cell hyperplasia. Mast cell number in duodenum
and jejunum of each experimental group is shown in Fig. 7. A significant mast cell
increase was also observed at the ileum of infected animals (16.20±0.66 per VCU,
compared to 3.95±0.25 cells in non-infected rats). Ketotifen treatment also reduced mast
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 14
cell number in the ileum, although this reduction was more moderate that in the
inflamed areas (9.40±1.10 cells in ketotifen 5 group and 12.69±0.45 cells in ketotifen 7
group).
Evaluation of inflammation
Histological evaluation. Infected control group showed signs of inflammation at the
mucosa and submucosa of duodenum and jejunum but not in the ileum. A mixed
inflammatory infiltrate with neutrophil and eosinophil cells was observed. Ketotifen did
not prevent inflammation but the number of infiltrate cells was smaller (data not
shown). Infected rats showed a significant hypertrophia of both muscular layers. This
hypertrophia affected all the areas of the small intestine, including those with no sign of
mucosa inflammation as the ileum. Muscular hypertrophia was still observed in
ketotifen treated groups but muscle layer thickness was of significant less magnitude in
the duodenum of infected rats treated with ketotifen (Table 1).
MPO. In non-infected control rats MPO concentration in serum was low at day 0 (517.9
± 68.62 ng . ml-1) and remained low during the whole period of study. In contrast, MPO
concentration significantly increased in all T. spiralis infected rats. This increase was
about 2 fold by day 5 PI (1029.3 ± 97.56 ng . ml-1, P<0.001) and reached 1545.8 ±
157.75 ng . ml-1 at day 12 PI (P<0.001). MPO values at day 12 in both ketotifen treated
groups were significantly increased when compared with values of non-infected control
rats. However, both ketotifen treatments reduced MPO concentration when compared
with infected-control animals, especially in those animals that received the longest
ketotifen treatment. Ketotifen in non-infected animals did not induce any change in
MPO concentration (Table 2).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 15
iNOS and COX-2 immunohistochemistry. In non-infected control animals weak iNOS
and COX-2 immunoreactivity was only detectable in the cytoplasm of some enterocytes
located in the apical side of intestinal villi (Fig. 8A). Infected control animals presented
a marked iNOS and COX-2 immunoreactivity throughout the gut wall. This positive
immunostaining was particularly noticeable in the epithelial cells, in the cytoplasm of
inflammatory cells located in the lamina propria and submucosa, and in both smooth
muscle layers (Fig. 8B). No differences were observed in samples from infected animals
receiving ketotifen treatment during 5 days (data not shown). In contrast, a general
reduction of iNOS and COX-2 immunoreactivity in all intestinal layers was observed in
those animals that received the longest ketotifen treatment (Fig. 8C).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 16
DISCUSSION
This study demonstrates that mast cell stabilization prevents both mucosal mast cell
hyperplasia and exacerbated motor responses in reaction to intestine inflammation.
Results show the strong correlation between mast cell activity and the development of
intestinal dysmotility, suggesting a potential use of mast cell stabilizers in the treatment
of motor disorders observed in post infectious IBS.
Several studies correlate mast cell activity with the development of functional disorders
such as alteration of intestinal permeability (Santos et al., 2001), motor disorders (Gay
et al., 2000a) and visceral hypersensitivity (Kreis et al., 1998). We previously
demonstrated that activation of mast cells could modify intestinal motility even in the
absence of apparent intestinal inflammation (Saavedra and Vergara, 2005). Moreover,
there are also several experimental and clinical studies that report an increase of mast
cell activity in both IBS and IBD patients (Raithel et al., 2001; Barbara et al., 2004; He,
2004). In spite of these evidences, there are only a few studies that have tested the use
of mast cell stabilizers to ameliorate symptoms derived from IBS and IBD (Stefanini et
al., 1992; Stefanini et al., 1995). The use of ketotifen has been reported in a couple of
case reports with 1 to 3 patients, but we could only find a pilot study using ketotifen for
the treatment of colitis in children (Jones et al., 1998). However, our results demonstrate
that ketotifen stabilizes mast cells and prevents all motor alterations induced by
inflammation in response to parasite infection.
Trichinella spiralis infection is a well accepted model of post infectious IBS (Bercik et
al., 2004; Wheatcroft et al., 2005). It has been widely used to study mechanisms
underlying the motor changes induced by inflammation (Vallance et al, 1999; Torrents
et al., 2002; Torrents et al., 2003; Khan et al., 2005), and constitutes a good model to
study the adaptation of the intestine to expel a clear cause of disease. Motor activity of
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 17
the intestine is due to the activation of stereotyped motor behavior patterns accompanied
by secretion and blood flow changes. One of these patterns is the defense program
characterized by power propulsive motility (Wood, 2004). However, this motor
reaction, if prolonged longer than infection resolution, can be the cause of IBS
symptoms (Barbara et al., 2004).
Several studies have demonstrated that power propulsive motility requires remodeling
of the enteric nervous system to increase muscle excitation necessary to expel the cause
of the disease, the parasite in this case (Palmer et al., 1984; Swain et al., 1992; Torrents
and Vergara, 2000; Tanovic et al., 2002). Mast cells are strategically placed to signal the
ENS to program the protective response (Wood, 2004) and therefore, our hypothesis
was that mast cells were implicated in the development of motor changes. Our results
show that the treatment with ketotifen, applied after the full development of
inflammation, reverted dysmotility. The ketotifen effect indicates that mast cell activity
is determinant for the development of the necessary changes at the enteric nervous
system that drive the development of hypermotility.
Two main mechanisms have been suggested for the remodelation of intestinal motor
patterns during inflammation: 1) increasing sensitivity of the GI reflexes increasing
activity of vagal afferents (Stead, 1992; Gay et al., 2000b; Di Giorgio et al., 2001;
Torrents et al., 2002) and 2) modifying sensitivity of the smooth muscle (Tanovic et al.,
2002). Mast cells have been located in close connection to vagal afferents. Furthermore,
a parallel increase of vagal afferents together with mucosal mast cell hyperplasia and a
greater involvement of vagus nerves on CCK response have been reported in another
similar model of nematode infection (Stead, 1992; Gay et al., 2001). In our model, the
response to CCK is mediated through vagal afferents since it is completely blocked by
capsaicin (Torrents et al., 2002). Parasite infection modifies CCK action exacerbating
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 18
excitatory response through a mechanism that implies NGF (Torrents et al., 2002). Our
present study shows that ketotifen treatment impaired the development of the
exacerbation of vagal response to CCK, most probably because of the reduction of mast
cell mediator(s) responsible for the development of vagal afferent hypersensitivity. We
believe this is the main site of action for the development of hypermotility. However,
we cannot rule out the effect of ketotifen reducing muscular thickness as a mechanism
of reducing motor response. Our study also shows that mast cell response and muscle
hypertrophy occur even at non-inflamed areas (ileum) in agreement with other authors’
findings (Tanovic et al., 2002).
Mast cells are bone marrow cells that migrate and differentiate in different tissues in the
body. Several factors released locally contribute to both mast cell migration and
proliferation including stem cell factor and interleukins (Galli et al., 2005). Our study
indicates that mast cell stabilization and therefore, the decrease of released mediators
diminish the “chemotactic call” as demonstrated by the smaller number of mast cells
found in the mucosa of treated animals, further contributing to diminish the
consequences of parasite infection.
In addition, there are a few studies that indicate that mast cell stabilizers could also
ameliorate inflammation (Hogaboam et al., 1993; Pothoulakis et al., 1993), and there is
an in vitro study that demonstrated that ketotifen is able to reduce nitric oxide
generation in human inflamed intestine specimens (Rachmilewitz et al., 1995). We
measured MPO in serum and the expression of iNOS and COX-2 in the intestine. These
enzymes have been shown to increase in the T. spiralis model (Hogaboam et al. 1996,
Torrents et al, 2003; Akiho et al. 2005). COX-2 and iNOS expression was present at all
layers of the inflamed intestine. In contrast to hypermotility and mast cell activity, MPO
did not vary significantly in ketotifen treated animals except for those receiving the
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 19
longest ketotifen treatment. A reduction of both iNOS and COX-2 was also observed in
ketotifen 7 group but not in the ketotifen 5 group. Although ketotifen has a limited
effect on inflammation a more prolonged treatment might induce a more significant
reduction of inflammation. In addition, it is necessary to remark that prevention of mast
cell hyperplasia and of hypermotility did not result in a worsening of parasite induced
inflammation.
Although the mechanism of action of ketotifen has not yet been well established and it
could be acting in other cell types such as blocking M-currents in neurons (Sato et al.,
2005) or inducing necrosis of human eosinophils (Hasala et al., 2005), we think that the
main action of ketotifen has been in stabilizing mucosal mast cells. Ketotifen is widely
accepted as a mast cell stabilizer (Eliakim et al., 1993; Hogaboam et al, 1993; Abe et
al., 2000). A direct effect on mucosal mast cells has been described (Abe et al., 2000;
Schoch, 2003) and we previously demonstrated, by measuring the RMCPII released in
the intestine, that ketotifen stabilizes intestinal mucosal mast cells in the rat (Juanola et
al., 1998). In this paper, we demonstrate that ketotifen significantly diminishes RMCPII
concentration in Trichinela spiralis infected rats, corroborating the effect of ketotifen as
an intestinal mucosal mast cell stabilizer.
None of the ketotifen effects were reproduced when ketotifen was applied shortly before
the experiment was conducted, indicating that our results are not a consequence of the
immediate stabilization of mast cells while evaluating motor action but of long term
action on mast cells.
In summary, our study demonstrates that mast cell activity is directly related to the
development of motor disorders caused by infection and inflammation. Our results
suggest that mast cell stabilizers could be a tool for the treatment of motor disorders in
IBD and IBS.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 20
ACKNOWLEDGEMENTS
Authors are thankful to A. Marco for his assistance in the histological evaluation of
intestine specimens, to A. Acosta for the care with the rats and to A.C. Hudson for
editorial revision of the manuscript.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 21
REFERENCES
Abe M, Kurosawa M, Igarashi Y, Ishikawa O and Miyachi Y (2000) Influence of IgE-
mediated activation of cultured human mast cells on proliferation and type I collagen
production by human dermal fibroblasts. J Allergy Clin Immunol 106:S72-S777.
Akiho H, Deng Y, Blennerhassett P, Kanbayashi H, Collins SM (2005) Mechanisms
underlying the maintenance of muscle hypercontractility in a model of postinfective gut
dysfunction. Gastroenterology 129:131-141.
Barbara G, DeGiorgio R, Stanghellini V, Cremon C, Salvioli B and Corinaldesi R
(2004) New pathophysiological mechanisms in irritable bowel syndrome. Aliment
Pharmacol Ther 20 Suppl 2: 1-9.
Bercik P, Wang L, Verdu FF, Mao YK, Blennerhasset P, Khan WI, Kean I, Tougas G
and Collins SM (2004) Visceral hyperalgesia and intestinal dysmotility in a mouse
model of postinfective gut dysfunction. Gastroenterology 127:179-187.
Castro GA and Fairbairn D (1969) Carbohydrates and lipids in Trichinella spiralis
larvae and their utilization in vitro. J Parasitol 55:51-58.
Chey WY, Jin HO, Lee MH, Sun SW and Lee KY (2001) Colonic motility abnormality
in patients with irritable bowel syndrome exhibiting abdominal pain and diarrhea. Am J
Gastroenterol 96:1499-1506.
Cowles VE and Sarna SK (1991) Trichinella spiralis infection alters small bowel motor
activity in the fed state. Gastroenterology 101:664-669.
Crampton HJ (2003) Comparison of ketotifen fumarate ophthalmic solution alone,
desloratadine alone, and their combination for inhibition of the signs and symptoms of
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 22
seasonal allergic rhinoconjunctivitis in the conjunctival allergen challenge model: a
double-masked, placebo- and active-controlled trial. Clin Ther 25:1975-1987.
De Giorgio R, Barbara G, Blennerhassett P, Wang L, Stanghellini V, Corinaldesi R,
Collins SM and Tougas G (2001) Intestinal inflammation and activation of sensory
nerve pathways: a functional and morphological study in the nematode infected rat. Gut
49:822-827.
Eliakim R, Karmeli F, Okon E and Rachmilewitz D (1995) Ketotifen ameliorates
capsaicin-augmented acetic acid-induced colitis. Dig Dis Sci 40:503-509.
Galli SJ, Nakae S and Tsai M (2005) Mast cells in the development of adaptive immune
responses. Nat Immunol 6:135-142.
Gay J, Fioramonti J, Garcia-Villar R and Bueno L (2000 a). Alterations of intestinal
motor responses to various stimuli after Nippostrongylus brasiliensis infection in rats:
role of mast cells. Neurogastroenterol Motil 12:207-214.
Gay J, Fioramonti J, Garcia-Villar R and Bueno L (2000b) Development and sequels of
intestinal inflammation in nematode-infected rats: role of mast cells and capsaicin-
sensitive afferents. Neuroimmunomodulation 8:171-178.
Gay J, Fioramonti J, Garcia-Villar R and Bueno L (2001) Enhanced intestinal motor
response to cholecystokinin in post-Nippostrongylus brasiliensis-infected rats:
modulation by CCK receptors and the vagus nerve. Neurogastroenterol Motil 13:155-
162.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 23
Hasala H, Malm-Erjefalt M, Erjefalt J, Giembycz MA, Zhang X, Moilanen E and
Kankaanranta H (2005) Ketotifen induces primary necrosis of human eosinophils. J
Ocul Pharmacol Ther 21:318-327.
He SH (2004) Key role of mast cells and their major secretory productos in
inflammatory bowel disease. World J gastroenterol 10:309-318.
Hogaboam CM, Bissonnette EY, Chin BC, Befus AD and Wallace JL (1993)
Prostaglandins inhibit inflammatory mediator release from rat mast
cells.Gastroenterology 104:122-129.
Hogaboam CM, Collins SM and Blennerhassett MG (1996) Effects of oral L-NAME
during Trichinella spiralis infection in rats. Am J Physiol 271:G338-G346.
Jones NL, Roifman CM, Griffiths AM and Sherman P. (1998) Ketotifen therapy for
acute ulcerative colitis in children: a pilot study.Dig Dis Sci. 43:609-615.
Juanola C, Giralt M, Jimenez M, Mourelle M and Vergara P (1998) Mucosal mast cells
are involved in CCK disruption of MMC in the rat intestine. Am J Physiol 275:G63-
G67.
Khan WI, Motomura Y, Blennerhassett PA, Kanbayashi H, Varghese AK, El-Sharkawy
RT, Gauldie J and Collins SM (2005) Disruption of CD40-CD40 ligand pathway
inhibits the development of intestinal muscle hypercontractility and protective immunity
in nematode infection. Am J Physiol Gastrointest Liver Physiol 288:G15-G22.
Kreis ME, Haupt W, Kirkup AJ and Grundy D (1998) Histamine sensitivity of
mesenteric afferent nerves in the rat jejunum. Am J Physiol. 275:G675-G680.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 24
Parry S and Forgacs I (2005) Intestinal infection and irritable bowel syndrome. Eur J
Gastroenterol Hepatol 17:5-9.
Palmer JM, Weisbrodt NW and Castro GA (1984) Trichinella spiralis: intestinal
myolectric activity during enteric infection in the rat. Exp Parasitol 57:132-141.
Pothoulakis C, Karmeli F, Kelly CP, Eliakim R, Joshi MA, O'Keane CJ, Castagliuolo I,
LaMont JT and Rachmilewitz D (1993) Ketotifen inhibits Clostridium difficile toxin A-
induced enteritis in rat ileum. Gastroenterology 105:701-707.
Rachmilewitz D, Stamler JS, Bachwich D, Karmeli F, Ackerman Z and Podolsky DK
(1995) Enhanced colonic nitric oxide generation and nitric oxide synthase activity in
ulcerative colitis and Crohn’s disease. Gut 36:718-723.
Raithel M, Winterkamp S, Pacurar A, Ulrich P, Hochberger J and Hahn EG 2001
Release of mast cell tryptase from human colorectal mucosa in inflammatory bowel
disease.Scand J Gastroenterol 36:174-179.
Ruitenberg EJ, Elgersma A and Kruizinga W. (1979) Intestinal mast cells and globule
leucocytes: role of the thymus on their presence and proliferation during a Trichinella
spiralis infection in the rat. Int Arch Allergy Appl Immunol 60:302-309.
Saavedra Y and Vergara P 2005) Hypersensitivity to ovalbumin induces chronic
intestinal dysmotility and increases the number of intestinal mast cells.
Neurogastroenterol Motil 17:112-122.
Santos J, Yang PC, Soderholm JD, Benjamin M and Perdue MH (2001) Role of mast
cells in chronic stress induced colonic epithelial barrier dysfunction in the rat.Gut.
48:630-636.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 25
Sato I, Munakata M and Iinuma K (2005) Histamine H1 antagonists block M-currents
in dissociated rat cortical neurons.Brain Res 1057:81-87.
Schoch C. (2003) In vitro inhibition of human conjunctival mast-cell degranulation by
ketotifen. J Ocul Pharmacol Ther. 19:75-81.
Stead RH (1992) Nerve remodelling during intestinal inflammation. Ann N Y Acad Sci
664:443-455.
Stefanini GF, Prati E, Albini MC, Piccinini G, Capelli S, Castelli E, Mazzeti M and
Gasbarrini G (1992) Oral disodium cromoglycate treatment on irritable bowel
syndrome: an open study on 101 subjects with diarrheic type. Am J Gastroenterol
87:55-57.
Stefanini GF, Saggioro A, Alvisi V, Angelini G, Capurso L, di Lorenzo G, Dobrilla G,
Dodero M, Galimberti M and Gasnarrini G (1995) Oral cromolyn sodium in comparison
with elimination diet in the irritable bowel syndrome, diarrheic type. Multicenter study
of 428 patients. Scand J Gastroenterol 30:535-541.
Swain MG, Agro A, Blennerhassett P, Stanisz A and Collins SM (1992) Increased
levels of substance P in the myenteric plexus of Trichinella-infected rats.
Gastroenterology 102:1913-1919.
Tanovic A, Jimenez M and Fernandez E (2002) Changes in the inhibitory responses to
electrical field stimulation of intestinal smooth muscle from Trichinella spiralis infected
rats. Life Sci 71:3121-3136.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 26
Torrents D, Prats N and Vergara P (2003) Inducible nitric oxide synthase inhibitors
ameliorate hypermotility observed after T. spiralis infection in the rat. Dig Dis Sci
48:1035-1049.
Torrents D, Torres R, De Mora F and Vergara P (2002) Antinerve growth factor
treatment prevents intestinal dysmotility in Trichinella spiralis-infected rats. J
Pharmacol Exp Ther 302:659-665.
Torrents D and Vergara P (2000) In vivo changes in the intestinal reflexes and the
response to CCK in the inflamed small intestine of the rat. Am J Physiol Gastrointest
Liver Physiol 279:G543-G551.
Vallance BA, Blennerhassett PA, Deng Y, Matthaei KI, Young IG and Collins SM
(1999) IL-5 contributes to worm expulsion and muscle hypercontractility in a primary
T. spiralis infection.Am J Physiol 277:G400-G408.
Wang YZ, Palmer JM and Cooke HJ (1991) Neuroimmune regulation of colonic
secretion in guinea pigs. Am J Physiol 260:G307-G314.
Wheatcroft J, Wakelin D, Smith A, Mahoney CR, Mawe G and Spiller R (2005)
Enterochromaffin cell hyperplasia and decreased serotonin transporter in a mouse model
of postinfectious bowel dysfunction. Neurogastroenterol Motil 17:863-870.
Williams RM, Bertoud HR and Stead RH (1997) Vagal afferent nerve fibres contact
mast cells in the rat small intestine mucosa. Neuroimmunomodulation 4:266-270.
Wood JD (2004) Enteric neuroimmunophysiology and pathophysiology.
Gastroenterology 127:635-657.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 27
Woodbury RG, Miller HR, Huntley JF, Newlands GF, Palliser AC and Wakelin D.
(1984) Mucosal mast cells are functionally active during spontaneous expulsion of
intestinal nematode infections in rat. Nature 1984 312:450-452.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 28
Footnotes:
Supported by GRANT SAF2002-03463 by DGI Ministerio Educación y Ciencia and
Grant 2001SGR 00214 and SGR2005 00255 by DURSI, Generalitat Catalunya. H.Serna
personal grant by HarlanIbérica SL
Reprint request: P. Vergara, Unidad de Fisiologia, Facultad de Veterinaria, Universitat
Autonoma de Barcelona, 08193 Bellaterra, Spain e-mail: [email protected]
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 29
LEGENDS FOR FIGURES
Figure 1: Representative mechanical recording of the spontaneous motor activity in the
small intestine from A, control group; B, infected control group and C, ketotifen 5
group. Similar recordings were obtained in all animals of the same group. D,
duodenum; J, jejunum; I, ileum.
Figure 2. Total number of spontaneous contractions recorded at duodenum in all
experimental groups. Similar results were found in the jejunum. *, **, P<0.05 and
P<0.01 respectively compared to infected control group.
Figure 3: Representative tracings showing the response to CCK-8 in duodenum (D) and
jejunum (J). CCK-8 (3 × 10 9 mol . kg 1 . 10 min 1) was i.v. infused. A: a control
group rat; B: a control infected rat; C: ketotifen 5 group rat. Horizontal line represents
10 min CCK infusion.
Figure 4: Quantification of CCK-8 (3 × 10 9 mol . kg 1 . 10 min 1) response in the
jejunum. The graph shows control inhibitory response to CCK-8 in jejunum of control
and ketotifen treated groups. In contrast, infected control group shows a significant
excitatory response. *, **, P<0.05 and P<0.01 respectively compared to infected control
group.
Figure 5: RMCPII concentration is serum at time 0 (before infection), day 5 (before
ketotifen treatment) and at day 12 PI in all experimental groups. **, P<0.01 compared
to infected control group.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 30
Figure 6: Microphotographs showing RMCPII immunopositive cells (mucosal mast
cells) in the intestinal mucosa of duodenum. A: control group rat; B: control infected
rat; C: ketotifen 5 group rat; D: ketotifen 7 group rat
Figure 7: Mast cell number in the mucosa of duodenum and jejunum of all experimental
groups. *, **, ***P<0.05, P<0.01 and P<0.001 respectively compared to infected
control group.
Figure 8: Immunohistochemical localization of iNOS and COX-2 proteins in
duodenum. A: non-infected control rat; B: control infected rat; C) ketotifen 7 rat.
Original magnification 400x.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

JPET #104620 31
Table 1. Thickness of circular and longitudinal muscle layers of duodenum and jejunum
Duodenum Jejunum Ileum
GROUP Circular Longitudinal Circular Longitudinal Circular Longitudinal
Non infected control 43.4 ± 1.43*** 32.5 ± 1.31*** 45.4 ± 8.16*** 22.7 ± 1.61*** 40.9 ± 1.66*** 25.6 ± 1.07
Infected control 163.1 ± 6.19 66.2 ± 2.76 116.6 ± 4.67 46.1 ± 2.29 74.5 ± 3.71 30.2 ± 1.98
Infected Ketotifen 5 104.3 ± 4.26*** 45.8 ± 1.59*** 94.5 ± 5.07 39.8 ± 2.37 82.7 ± 5.61 32.4 ± 2.83
Infected Ketotifen 7 116.6 ± 4.92*** 50.8 ± 2.62*** 104.6 ± 4.78 44.1 ± 2.59 85.2 ± 5.80 31.9 ± 1.91
Data in µm (means ± SEM)
*** P < 0.001 compared to the infected control group
This article has not been copyedited and form
atted. The final version m
ay differ from this version.
JPET
Fast Forward. Published on Septem
ber 20, 2006 as DO
I: 10.1124/jpet.106.104620 at ASPET Journals on May 14, 2019 jpet.aspetjournals.org Downloaded from

JPET #104620 32
Table 2. Concentration of serum myeloperoxidase (MPO) in basal situation and at days 5 and 12 PI
GROUP Basal Day 5 Day 12
Non infected control 517.9 ± 68.62 502.9 ± 60.02*** 572.3 ± 52.98***
Ketotifen control 475.1 ± 48.49 518.1 ± 51.11*** 493.4 ± 44.83***
Infected control 474.7 ± 61.30 1029.3 ± 97.56 1545.8 ± 157.75
Infected Ketotifen 5 514.4 ± 36.56 1018.5 ± 88.24 1260.6 ± 60.88
Infected Ketotifen 7 394.5 ± 47.22 1138.7 ± 85.33 838.4 ± 112.70**
Data in ng/ml (means ± SEM)
**,*** P < 0.01 and P< 0.001 respectively compared to the infected control group
This article has not been copyedited and form
atted. The final version m
ay differ from this version.
JPET
Fast Forward. Published on Septem
ber 20, 2006 as DO
I: 10.1124/jpet.106.104620 at ASPET Journals on May 14, 2019 jpet.aspetjournals.org Downloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and form
atted. The final version m
ay differ from this version.
JPET
Fast Forward. Published on Septem
ber 20, 2006 as DO
I: 10.1124/jpet.106.104620 at ASPET Journals on May 14, 2019 jpet.aspetjournals.org Downloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on September 20, 2006 as DOI: 10.1124/jpet.106.104620
at ASPE
T Journals on M
ay 14, 2019jpet.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and form
atted. The final version m
ay differ from this version.
JPET
Fast Forward. Published on Septem
ber 20, 2006 as DO
I: 10.1124/jpet.106.104620 at ASPET Journals on May 14, 2019 jpet.aspetjournals.org Downloaded from