inter-homologue repair in fertilized human eggs?
TRANSCRIPT

1
Inter-homologuerepairinfertilizedhumaneggs?DieterEgli1,*,MichaelV.Zuccaro2,Michal Kosicki3, GeorgeM.Church4,Allan Bradley3, andMariaJasin5,*1DepartmentofObstetricsandGynecologyandDepartmentofPediatrics,ColumbiaUniversity,NewYorkNY10032,USA;2GraduateProgram,DepartmentofPhysiologyandCellularBiophysics,ColumbiaUniversity,NewYorkNY10032,USA;3WellcomeTrustSangerInstitute,Hinxton,Cambridge,CB101SA,UnitedKingdom;4DepartmentofGenetics,HarvardMedicalSchool,77AvenueLouisPasteur,Boston,Massachusetts02115,USA,5DevelopmentalBiologyProgram,MemorialSloanKetteringCancerCenter,NewYork,NY,10065,USA*correspondence:[email protected],[email protected] Manyhumandiseaseshaveanunderlyinggeneticcomponent.Thedevelopmentandapplicationofmethods to prevent the inheritance of damaging mutations through the human germline could havesignificanthealthbenefits,andcurrentlyincludepreimplantationgeneticdiagnosisandcarrierscreening.Maetal.takethisastepfurtherbyattemptingtoremoveadiseasemutationfromthehumangermlinethrough gene editing1. They assert the following advances: (i) the correction of a pathogenic genemutationresponsible forhypertrophic cardiomyopathy in human embryos using CRISPR-Cas9 and (ii)theavoidanceofmosaicismineditedembryos.Inthecaseofcorrection,theauthorsconcludethatrepairusingthehomologouschromosomewasasormorefrequentthanmutagenicnonhomologousend-joining(NHEJ).Theirconclusionissignificant,ifvalidated,becausesucha“self-repair”mechanismwouldallowgenecorrectionwithouttheintroductionofarepairtemplate.Whiletheauthors’analysesreliedonthefailure to detect mutant alleles, here we suggest approaches to provide direct evidence for inter-homologue recombination and discuss other events consistent with the data. We also review thebiologicalconstraintsoninter-homologuerecombinationintheearlyembryo. In their first approach, Ma et al. used donor sperm from a patient heterozygous for theMYBPC3ΔGAGTmutation to fertilizewild-typeoocytes, such that half of the embryos startedout aswildtypeattheMYBPC3locusandhalfheterozygous.FertilizedzygoteswereinjectedwithCas9andansgRNAdirectedtocreateadouble-strandbreak(DSB)inthemutantpaternalallele.Theauthorsreportthat24%of the embryos at day 3 of development were mosaic, with some cells of the embryo containing themutantpaternallocus,eitherintactormodifiedbyNHEJ,togetherwithawild-typelocus.Remainingcellsof the embryo contained only a detectablewild-type allele.While some zygoteswere also co-injectedwith a wild-type, exogenous, single-stranded oligodeoxynucleotide template (ssODN) with twosynonymous mutations, no mutations consistent with ssODN-templated repair were detected.Furthermore,‘wild-typeonly’cellswerepresentatasimilarfrequencybothinthepresenceandabsenceofthessODN.Theauthorsinferthatthesecellsarosebyhomology-directedrepair(HDR)ofthemutantpaternal allele using the wild-typematernal allele as a template, i.e., inter-homologue recombination,leadingtogenecorrection.
Inasecondapproach,earlier,MII-phaseoocyteswerecoinjectedwithCas9complexesanddonorsperm.Inthiscase,mosaicismwasnotdetected,exceptinasingleembryo,whichcontainedboth‘wild-type only’ cells and ones heterozygous forwild-type and ssODN-templated alleles. Althoughwild-typeembryos were expected at 50% frequency, they appeared to comprise 72% of embryos. The authors
.CC-BY-NC-ND 4.0 International licensepeer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/181255doi: bioRxiv preprint first posted online Aug. 28, 2017;

2
arguethattheexcess(22%)ofapparentlywild-typeembryosaroseintheMII-injectedoocytesbyHDRusing thematernal allele to correct thepaternal allele, as in the zygote injections, and rarelyused thedonortemplate.Thus,amajorinferenceofthisarticleisthataDSBgeneratedbyCas9inhumangametesandzygotes isefficientlyrepairedby inter-homologuerecombination(Fig.1a).Theconclusionthat thepathogenic allele can be efficiently correctedwithoutmosaicism has far-reaching implications for theauthors’statedgoalofusingsuchmethodstoaddressthepublichealthburdenofmonogenicdisease. Given the far-reaching implications, providing direct evidence for correction of the pathogenicallele,ratherthantheinabilitytodetectthemutantallele,cannotbeoveremphasized.Anotherstudyinmice similarly relied on the absence of a mutant allele to conclude repair by inter-homologuerecombination2. Considering the data presented in Ma et al., alternatives to recombination betweenhomologuesarepossibleandwouldseemmorelikely,asthecellbiologyoffertilizedeggswouldappearto preclude the direct interaction between the maternal and paternal genomes required for inter-homologue HDR. Therefore, clear evidence for a novel linkage of maternal and paternal alleles is animperativeforanyembryothatwouldbeconsideredforfutureimplantation. Novel DNA linkages can be detected directly by sequencing, but the authors do not indicatewhethertheirgenomesequencingdatawasphasedtodetecttheincorporationofthewild-typesequencefromoneofthematernalhomologuesattheDSBintothepaternalchromosome(Fig.1A).Newparentallinkages can also be directly assayed by long-range PCR using allele-specific primers; the onlyrequirement is that SNPs/indels exist to distinguish the maternal and paternal chromosomes in thevicinityofthemutation3.Ofrelevance,thistypeofanalysiscanbeperformedonindividualoocytes4. Without direct verification of inter-homologue recombination at the mutant allele, attemptsshouldbemadetoruleoutothertypesofevents.Duringgeneediting,NHEJisusuallyconsideredtoleadto small indels atDSB sites.However,with appropriate experimental design, longdeletions andotherevents canbedetected in culturedcells and inbothmouseandpig zygotes5-7. InMaetal., genotypinginvolved amplification of a ~534 bp fragment inwhich theMYBPC3ΔGAGTmutation is ~200 bp from aprimer-bindingsite.Deletions>200bpwouldbesufficienttoremovethisprimer-bindingsiteandleadtoamplification only of thematernal allele (Fig. 1A, B), giving themisleading appearance of a correctedpaternalallele.Todetectlongerdeletions,amatrixofprimerpairsneedtobetiledatincreasingdistanceson both sides of themutation. In a study designed to systematically score these events, Cas9-inducedDSBs in mouse embryonic stem cells were found to resolve into large deletions (250-9500 bp) inapproximately20%ofeditedcells(M.K.andA.B.,unpublishedresults).Thisapproachremainsimperfecttodetectallevents,though,becauseverylargedeletionsorothereventssuchastranslocationspreventamplification and thus escape characterization. Given the ramifications, more studies of this type arerequiredtoquantifytheseeventsatother loci,particularly inembryos.BecausefertilizationbymutantspermintheMaetal.studycanbeconfidentlyinferredonlyformosaicembryos,thistypeofanalysisisnotsuitableforembryosderivedfromMII-phaseoocyteinjections.Thus,linkageanalysisisnecessaryinthesecases. Arethereotherpossibleoutcomesthatcanresultinawild-typegenotypeinaPCRassaybutnot
.CC-BY-NC-ND 4.0 International licensepeer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/181255doi: bioRxiv preprint first posted online Aug. 28, 2017;

3
involve interhomolog recombination? Zygotes with a single pronucleus are not uncommon afterintracytoplasmic sperm injection, occurring in ~10% of fertilization attempts, and are mostly ofparthenogenetic origin, containing only the maternal genome8 (Fig. 1C). These zygotes are normallydiscarded,andMaetal.showthepresenceoftwopronuclei,althoughtheydonotprovideinformationonthenumberandtypesofabnormalfertilizations.Furthermore,parthenogenesiscanalsoresultinzygoteswith two maternal genomes when extrusion of the second polar body fails (Fig. 1C)9,10. A paternalcontributionwasverifiedbycytogeneticanalysisinsomeofthestemcelllinesgeneratedfromembryosbyMaetal. (2/6); reportingon thepresenceofuniquepaternalpolymorphisms inall embryoswouldaddress the frequency of parthenogenesis. It also remains possible that a fraction of embryos derivedfromsuccessfulfertilizationwithmutantspermareatmoreriskofpaternalchromosomelossduetotheoccurrenceoftheCas9-inducedDSB.
Although inter-homologue recombination in fertilized oocytes and zygotes cannot currently beexcluded,physicalseparationofmaternalandpaternalgenomeswouldbeexpectedtobeasubstantialimpediment.Uponfertilization,distinctmaternalandpaternalnucleiform(pronuclei),suchthatthetwogenomesareseparateinacellthat ismorethan100µmindiameter(Fig.1D-G).ThismaypreventtheincorporationofpaternalchromosomesintotheoocyteMIIspindle(Fig.1E).Duringthefirstinterphase,maternal and paternal pronuclei migrate from the site of their formation towards the center ofthezygote,buttheseparationpersiststhroughoutinterphase(Fig.1F,G),atwhichtimeindividualnucleican be manipulated11. In both human and mouse zygotes, maternal and paternal genomesundergo DNAreplicationinseparatenuclei,andenterthefirstmitosisasseparateentities,atwhichtimetheycanstillbe manipulated separately (Fig. 1F,G). Merging of maternal and paternal chromosomesdoes not occuruntilmicrotubuleactionassemblesbothgenomesonacommonmetaphaseplateatthefirstmitosis12,13.Therefore, direct interactions between maternal and paternal genomes required forinter-homologue repair do not seemingly occur until embryos enter the 2-cell stage when the twogenomesarepackagedwithinthesamenucleus.
Although the study of DSB repair in human embryos is in its infancy, inter-homologuerecombination inmitotic cells appears tobe significantly less frequent than inter-sister recombination(orNHEJ),whichmaybe due, at least in part, to themuch larger nuclear volumehomologues occupycompared to sister chromatids14. By contrast, inter-homologue recombination in meiosis, which isessentialforthereductionaldivisiontoformgametes,isefficient,likelyduetothelargenumberofDSBsthatareprogrammedto formoneachchromosometopromotehomologuepairing15. It is important tonote,however,thatmeioticinter-homologuerecombinationoccursduringfetaldevelopmentinfemales16andsoit istemporallyremovedfromtheeventsdescribedinMaetal.WhethermeioticrecombinationfactorsarestillexpressedandactiveinMII-phaseoocytesdecadeslaterhasnotbeenexaminedasfarasweareaware.
Insummary,theconclusionofgenecorrectioninhumanembryosrequiresfurtherinvestigation,includingdirectverification.Efficientinter-homologuerecombinationinembryosinwhichthematernaland paternal genomes are undergoing distinct biological programs and in distinct nuclei would be astunningbiologicalfinding.Butitwouldalsoputcellsatriskforunmaskingdeleteriousrecessivealleles
.CC-BY-NC-ND 4.0 International licensepeer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/181255doi: bioRxiv preprint first posted online Aug. 28, 2017;

4
throughlossofheterozygosity(notshowninFig.1A).Theclinicalimplicationsofgeneeditinginhumanembryosaresubstantial.Whilegeneeditingcouldreducedisease-causingalleles,inadvertentchangestothehumangermline like longdeletionsand lossofheterozygosityhavenotbeenruledout.Thus,eachembryo needs to be carefully evaluated to confirm (or not) gene correction and lack of mosaicism.Despitetheinherentlimitationsimposedonsuchresearch,itisessentialthatconclusionsregardingtheability to correct amutation in human embryos be fully supported. Absent such data, the biomedicalcommunityand,critically,patientswithdisease-causingmutations interested insuchresearchmustbemadeawarethatnumerouschallengesingenecorrectionremain.
References:1 Ma,H.etal.Correctionofapathogenicgenemutationinhumanembryos.Nature,
doi:10.1038/nature23305(2017).2 Wu,Y.etal.CorrectionofageneticdiseaseinmouseviauseofCRISPR-Cas9.Cellstemcell13,659-
662,doi:10.1016/j.stem.2013.10.016(2013).3 Jeffreys,A.J.&May,C.A.Intenseandhighlylocalizedgeneconversionactivityinhumanmeiotic
crossoverhotspots.NatGenet36,151-156,doi:10.1038/ng1287(2004).4 Cole,F.etal.Mousetetradanalysisprovidesinsightsintorecombinationmechanismsandhotspot
evolutionarydynamics.NatGenet46,1072-1080,doi:10.1038/ng.3068(2014).5 Shin,H.Y.etal.CRISPR/Cas9targetingeventscausecomplexdeletionsandinsertionsat17sites
inthemousegenome.Naturecommunications8,15464,doi:10.1038/ncomms15464(2017).6 Whitworth,K.M.etal.UseoftheCRISPR/Cas9systemtoproducegeneticallyengineeredpigs
frominvitro-derivedoocytesandembryos.Biologyofreproduction91,78,doi:10.1095/biolreprod.114.121723(2014).
7 Parikh,B.A.,Beckman,D.L.,Patel,S.J.,White,J.M.&Yokoyama,W.M.DetailedphenotypicandmolecularanalysesofgeneticallymodifiedmicegeneratedbyCRISPR-Cas9-mediatedediting.PloSone10,e0116484,doi:10.1371/journal.pone.0116484(2015).
8 Sultan,K.M.,Munne,S.,Palermo,G.D.,Alikani,M.&Cohen,J.Chromosomalstatusofuni-pronuclearhumanzygotesfollowingin-vitrofertilizationandintracytoplasmicsperminjection.Humanreproduction(Oxford,England)10,132-136(1995).
9 Kim,K.etal.Recombinationsignaturesdistinguishembryonicstemcellsderivedbyparthenogenesisandsomaticcellnucleartransfer.Cellstemcell1,346-352(2007).
10 Paull,D.etal.NucleargenometransferinhumanoocyteseliminatesmitochondrialDNAvariants.Nature493,632-637,doi:nature11800[pii]
10.1038/nature11800[doi](2013).11 Kattera,S.&Chen,C.Normalbirthaftermicrosurgicalenucleationoftripronuclearhuman
zygotes:casereport.Humanreproduction(Oxford,England)18,1319-1322(2003).12 Egli,D.etal.Reprogrammingwithinhoursfollowingnucleartransferintomousebutnothuman
zygotes.Naturecommunications2,488,doi:ncomms1503[pii] 10.1038/ncomms1503[doi](2011).13 Egli,D.,Rosains,J.,Birkhoff,G.&Eggan,K.Developmentalreprogrammingafterchromosome
transferintomitoticmousezygotes.Nature447,679-685(2007).14 Stark,J.M.&Jasin,M.Extensivelossofheterozygosityissuppressedduringhomologousrepairof
chromosomalbreaks.MolCellBiol23,733-743(2003).15 Kauppi,L.etal.Numericalconstraintsandfeedbackcontrolofdouble-strandbreaksinmouse
meiosis.GenesDev27,873-886,doi:10.1101/gad.213652.113(2013).16 Baker,T.G.AQUANTITATIVEANDCYTOLOGICALSTUDYOFGERMCELLSINHUMANOVARIES.
ProceedingsoftheRoyalSocietyofLondon.SeriesB,Biologicalsciences158,417-433(1963).
.CC-BY-NC-ND 4.0 International licensepeer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/181255doi: bioRxiv preprint first posted online Aug. 28, 2017;

5
FigureLegend
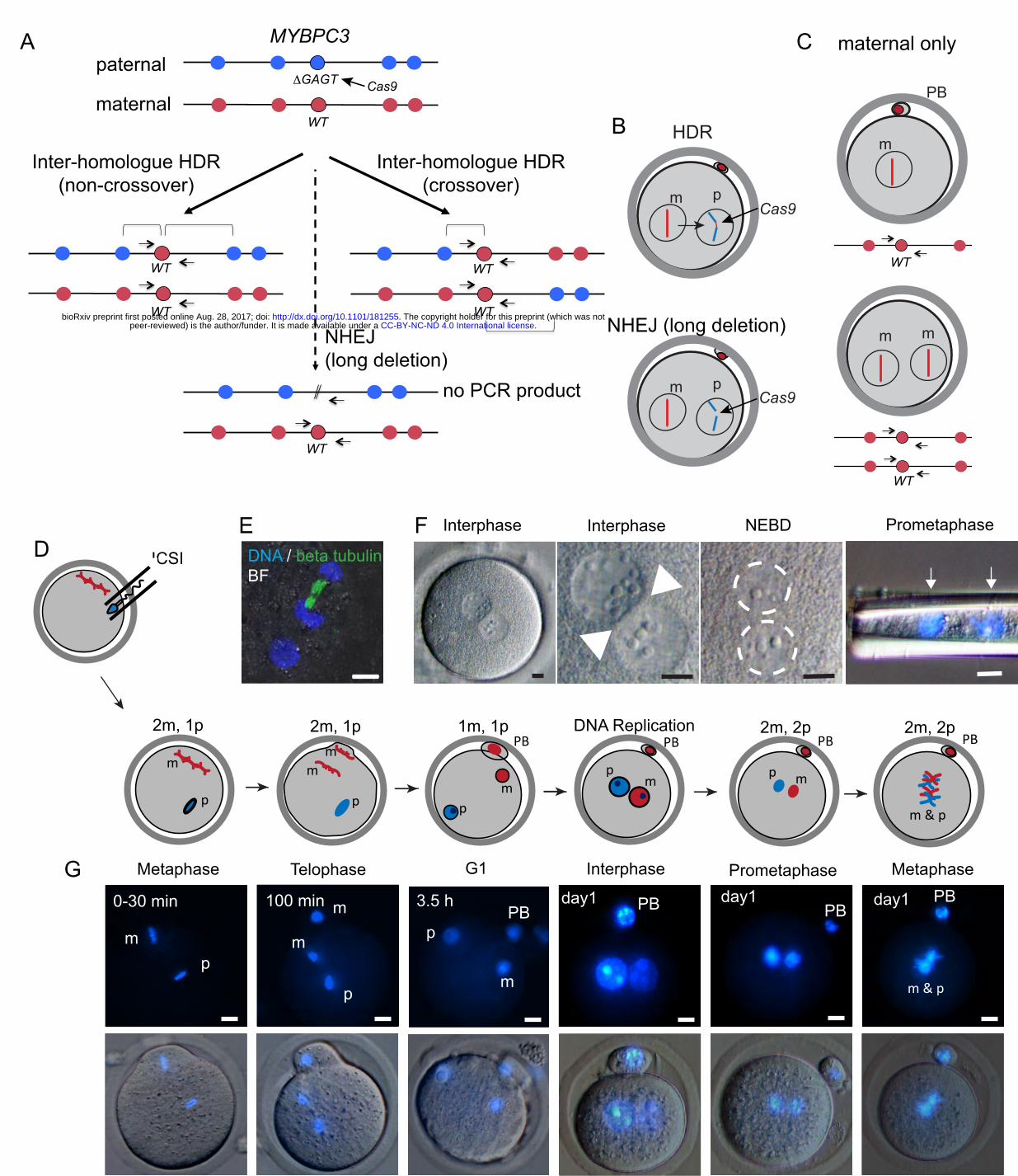
Figure 1 |Constraints on gene editing by inter-homologue recombination in the early humanembryo.A. PossiblerepairoutcomesafteraCas9-inducedDSBatthepaternalMYBPC3∆GAGTlocus.Redandblue circlesindicateuniquematernalandpaternalgeneticvariants.Inter-homologueHDRresultsingene conversionofthepaternalallelebythewild-type(WT)maternalallele.Therepairoutcomecanbeanon-crossoveroracrossover.Onlyoneoutcomeofcrossing-overisshowninwhichtherecombined chromosomesareinthesamenucleus.Thealternativeisthattherecombinedchromosomessegregateto differentdaughtercells,suchthatlossofheterozygositywouldoccuronthechromosomefromthepoint oftheHDReventtothetelomereinbothdaughtercells,onewithhomozygosityforthematernal chromosomeandtheotherforthepaternalchromosome.Thisoutcomewouldbeexpectedinhalfofthe crossing-overeventsthatunderwentinter-homologueHDRinG2phase.NHEJeventsarealsopossible thatresultinthedeletionofaprimer-bindingsiteusedforgenotyping.B. Schematic of possible repair outcomes after Cas9 cleavage in the human zygote from panel A. m, maternalchromosome,p,paternalchromosome.C. Parthenogenesis after fertilization failurewith (top) andwithout (bottom) second polar body (PB) extrusion.OutcomesofA-CareindistinguishableingenotypingassaysusingflankingPCRprimersalone.D. Schematic of intracytoplasmic sperm injection (ICSI) followed by progression through the first cell cycleduringday1ofdevelopment.Thenumberofmaternalandpaternalgenomesareindicatedateach phaseofthecellcycle.E. Immunofluorescenceofamousezygoteattelophaseofthesecondmaternalmeioticdivision.Notethat onlythematernalgenomesareattachedtomicrotubules,whilethepaternalgenomebeginstoforman interphasenuclearmembranetoreplacethespermmembrane.BF=brightfield.F. Progression of human zygotes through the first cell cycle from the two-pronuclear stage to prometaphase,whenthetwogenomescanberemovedfromtheeggbyaneedle.Notetheseparationof thetwogenomes(arrowsanddashedcircles).NEBD,pronuclearenvelopebreakdown.G. Cellcycleprogressionduringday1infertilizedmousezygotes.Of23mouseeggs,noneshoweddirect contactbetweenthematernalandpaternalgenomes.Sizebars(independentofcoloror length ineach panel)=10µm.PanelsoneandfourinFandpanelsfourtosixinGareaspublishedinreference12andinreference13.
AuthorcontributionstatementD.E.andM.J.wrotethepaperwithcontributionsfromM.K.,A.B,M.Z.andG.M.C.M.Z.andD.E.performedICSIandimagingofeggs.M.K.andA.B.providedunpublishedinformation.
.CC-BY-NC-ND 4.0 International licensepeer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/181255doi: bioRxiv preprint first posted online Aug. 28, 2017;
paternal
maternal
MYBPC3
WT
ΔGAGT Cas9
Inter-homologue HDR(non-crossover)
Inter-homologue HDR(crossover)
NHEJ(long deletion)
no PCR product
WT
WT
WT
WT
WT
ICSI
A
D
1m, 1pPB
mp
DNA ReplicationPB
mp
2m, 2p
mp
PB2m, 2p
m & p
PB
Interphase Prometaphase Metaphase
day1 day1 day13.5 hPB
m
p
G1
PB
2m, 1pm
m
p
2m, 1p
m
p
100 min
TelophaseMetaphase
0-30 min
m m
p
m
G
p
maternal only
B
m pCas9
NHEJ (long deletion)
m p
HDR
Cas9
C
m m
m
PB
DNA / beta tubulin BF
NEBDInterphaseInterphaseF
DNA
PrometaphaseE
PBPB
m & p
WT
WT
.CC-BY-NC-ND 4.0 International licensepeer-reviewed) is the author/funder. It is made available under aThe copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/181255doi: bioRxiv preprint first posted online Aug. 28, 2017;