lagerstätten in manitoba
TRANSCRIPT

GEOSCIENCE CANADA Volume 39 2012 201
SERIES
Great CanadianLagerstätten 3. Late Ordovician Konservat-Lagerstätten in Manitoba
Graham A. Young1, David M.Rudkin2, Edward P. Dobrzanski1,Sean P. Robson1, Michael B.Cuggy3, Matthew W. Demski4, andDeborah P. Thompson1
1The Manitoba Museum190 Rupert AvenueWinnipeg, MB, Canada, R3B 0N2 E-mail: [email protected]
2Royal Ontario Museum100 Queen’s Park CrescentToronto, ON, Canada, M5S 2C6
3Department of Geological SciencesUniversity of SaskatchewanSaskatoon, SK, Canada, S7N 5E2
4Department of Geological SciencesUniversity of ManitobaWinnipeg, MB, Canada, R3T 2N2
SUMMARYKonservat-Lagerstätten, deposits in whichsoft-bodied or lightly sclerotized fossilsare preserved, are very rare inOrdovician strata. Three significantsites are known from UpperOrdovician rocks in Manitoba: at CatHead – McBeth Point, William Lake,
and Airport Cove. These sites are intwo distinct sedimentary basins: theformer two are in the Williston Basin,while the latter is in the Hudson BayBasin. All three sites contain marinefossils, but each has a different assem-blage that contributes a distinct pieceof the diversity picture. Importantgroups represented at one or more ofthe sites include seaweeds (algae),sponges, cnidarian medusae (jellyfish),conulariids, trilobites, eurypterids,xiphosurids (horseshoe crabs), andpycnogonids (‘sea spiders’). The differ-ent biotas reflect depositional condi-tions at each site. Many of the fossilsare unknown elsewhere in theOrdovician at the family level or high-er. The province of Manitoba there-fore makes a significant contribution toknowledge of Late Ordovician biodi-versity.
SOMMAIRELes lagerstätten de conservation, ces sédi-ments contenant des fossiles d’organ-ismes à corps mou ou légèrementsclérotisés particulièrement bien con-servés, sont très rares dans les stratesordoviciennes. Trois sites d’impor-tance sont connus dans des roches del’Ordovicien supérieur à Cat Head,Manitoba, soit McBeth Point, WilliamLake et Airport Cove. Ces sites sontsitués dans deux bassins sédimentairesdistincts: les deux premiers sont situésdans le bassin de Williston, tandis quele second est situé dans le bassin de labaie d’Hudson. Les trois sites contien-nent des fossiles marins, mais chacunprésente un assemblage différent, cha-cun montrant une composante dis-tincte de la diversité biologique d’alors.Les groupes les plus importantsreprésentés, dans un ou plusieurs deces sites, sont les algues, les éponges,les cnidarian medusae (méduses), les
conularides, les trilobites, les euryp-térides, xiphosurides (limules) et pyc-nogonides. Les différents biotopesreflètent les conditions de dépôt dechaque site. Nombre de ces fossilessont inconnus ailleurs dansl’Ordovicien, au niveau de la famille oudu taxon supérieur de la classification.Ainsi, la province du Manitoba offre-t-elle une contribution importante à laconnaissance de la biodiversité del’Ordovicien supérieur.
INTRODUCTION Konservat-Lagerstätten (sing. Lagerstätte)are deposits that preserve entirely soft-bodied or lightly sclerotized organisms,or the soft anatomy of forms knownotherwise only from their mineralizedor decay-resistant anatomy. Suchdeposits are unusual in the fossilrecord, because unmineralized tissuesare typically broken down entirely bypostmortem scavenging and decompo-sition. Konservat-Lagerstätten are of sci-entific importance both in establishingthe evolutionary histories of organismsthat would be otherwise unknown asfossils, and in providing exceptionalevidence of long-vanished biologicalcommunities.
Life in the world’s oceansincreased in diversity through much ofthe Ordovician Period (488 to 444 mil-lion years ago), to the extent that wenow talk about the Great OrdovicianBiodiversification Event, or GOBE(Webby et al. 2004). This event, ofunprecedented scale, was comparablein significance to the famous CambrianExplosion of about 540 million yearsago (Webby 2004). Although the greatmajority of our knowledge of theOrdovician event is based on therecord of fossils with preservable hardparts, important evidence aboutOrdovician biodiversity may also be

202
found in Konservat-Lagerstätten. Evidence of this biodiversity
is, however, tantalizingly scarce, asOrdovician Konservat-Lagerstätten arerare globally. Soft tissue preservationfor this time period is far less well doc-umented than for the precedingCambrian or, indeed, for the subse-quent Silurian. Manitoba is fortunate tobe endowed with three distinct sites orareas known to contain significantOrdovician soft tissue preservation.These are at Cat Head (or the CatHead – McBeth Point area), AirportCove, and William Lake (Fig. 1).Manitoba is still little-explored geologi-cally in comparison with many otherparts of the world, and there are prob-ably additional sites waiting to befound. This province may thus make aglobally significant contribution to ourunderstanding of Late Ordovician bio-diversity. The three known sites havenot yet been thoroughly documented,and much still remains to be learned;studies are ongoing.
Ordovician rocks in Manitobawere deposited in two distinct sedi-mentary basins: the Williston Basin tothe southwest and the Hudson BayBasin to the northeast (Elias 1991;Young et al. 2008; Fig. 1). Of the threeLagerstätten, the Cat Head and WilliamLake biotas occur in the WillistonBasin, while the Airport Cove biota isin the Hudson Bay Basin. The CatHead biota, of mid Katian (‘earlyMaysvillian’) age, is significantly olderthan the other two, which are consid-ered to be of latest Katian (‘lateRichmondian’) age (Fig. 1).
Ordovician rocks in Manitobarecord several cycles of sea level riseand fall within the interior of the paleo-continent called Laurentia, which
makes up most of modern NorthAmerica (Elias 1991). Each of thethree Lagerstätten represents conditionsin a particular part of a recognized sealevel cycle. Cat Head strata (Fig. 2a) aregenerally considered to represent deep-er water, quiet conditions near thetransgressive maximum of the RedRiver Cycle. Strata at William Lake(Fig. 2b, c) record very restricted shal-low marginal marine conditions nearthe regressive extreme at the top ofthe Stony Mountain Cycle, while theAirport Cove rocks (Fig. 2d-f) werealso deposited under restricted margin-
al marine conditions, possibly duringan interval of sea level fluctuation nearthe extreme of a regression. As a resultof the distinctions in age and paleoen-vironment, the Cat Head biota, domi-nated by groups such as trilobites andnon-calcareous algae, and distinguishedby an extraordinarily diverse conulariidfauna (six species), is quite differentfrom the horseshoe crabs andeurypterids that characterize the othertwo Lagerstätten.
On a global level, Ordoviciansoft-bodied fossils are very rare.Perhaps the two most remarkable
MANITOBA
Cat Head -McBeth Pt.
Airport Cove
Ordovicianoutcrop belts
N
HUDSONBAY
BASIN
WILLISTONBASIN
200 km
MB
5 km
MP
II
CHLB
KB
William Lake
AC
PC–
Churchill Area
Severn RiverFormation
ProterozoicChurchillQuartzite
Chu
rchi
ll R
iver
Gro
up?
OR
DO
VIC
IAN
Kat
ian
?SI
LUR
IAN
Ll
ando
v.
10 m
Lake Winnipeg Area
CM
Dog HeadMember
Cat Head Mbr.
SelkirkMember
Fort GarryMember
PenitentiaryMember
GuntonMember
Williams Mbr.
StonewallFormation
t-marker
Fisher Br. Fm.
OR
DO
VIC
IAN
Kat
ian
SIL.
Ll
.H.
?
Red
Riv
er F
orm
atio
nS
tony
Mtn
. Fm
.
10 m
WL
Figure 1. Geographic locations and stratigraphic positions of the Cat Head –McBeth Point (CM), William Lake (WL), and Airport Cove (AC) sites, Manitoba(MB). Lake Winnipeg area stratigraphy is after Elias (1981, 1991), Bezys (1996), andYoung et al. (2008). Churchill area stratigraphy is after Elias et al. (1999) and Younget al. (2003). CH = Cat Head; Fisher Br. = Fisher Branch; H.? = Hirnantian?; II =Inmost Island; KB = Kinwow Bay; LB = Lynx Bay; Ll. & Llandov. = Llandovery;MP = McBeth Point; SIL. = Silurian; Stony Mtn. = Stony Mountain.
Figure 2 (page 203). Fossil localities and examples of fossils and lithotypes. (a) The Cat Head Member of the Red RiverFormation is exposed in cliffs along the North Basin of Lake Winnipeg at Cat Head, looking southeastward toward McBeth Point;inset is an example of the probable rhodophytic alga Manitobia patula, collected from Inmost Island in 1884 by T.C. Weston (col-lection of the Geological Survey of Canada, Ottawa, GSC 136962). (b) At the William Lake site, thin-bedded dolomudstones ofthe Williams Member, Stony Mountain Formation, show broad ripples and channel features. (c) On the upper surface of a dolo-stone bed at William Lake, a fossil medusa (jellyfish) rests in a homogeneous mud body, while laterally a pavement of mouldic lin-gulid brachiopod shells fills a depressed area. (d) Field collecting at the Airport Cove site in July, 2011; note proximity of ridge ofProterozoic Churchill quartzite in the background, which represents the flank of a paleo-island; note also the water pail for wet-ting fossil slabs for examination, and shotgun in the foreground for protection against polar bears. (e) A polished vertical sectionthrough the dolomudstone from Airport Cove shows dark pyritic laminae and wavy bedding. (f) Airport Cove represents a broad,arcuate indentation on the north side of a paleo-island of Churchill quartzite. Ordovician rocks tentatively assigned to theChurchill River Group outcrop in various places in the eastern part of the cove; the Lagerstätte biota occurred in loose slabs nearthe base of the peninsula to right of centre.

GEOSCIENCE CANADA Volume 39 2012 203

204
assemblages are in Gondwanan Africa:in the Lower Ordovician Fezouata for-mations of Morocco, and the UpperOrdovician Soom Shale of SouthAfrica. The Fezouata biota, whichseems to exhibit ‘Burgess Shale-type’preservation, constitutes an extraordi-nary array of sponges, annelid worms,and a great variety of superbly-pre-served arthropods, along with grapto-lites, conulariids, hyolithids and othermolluscs, brachiopods, bryozoans, andechinoderms (Van Roy 2006; Vintheret al. 2008; Van Roy et al. 2010).These fossils apparently represent crea-tures that lived on an open marineshelf. The Soom Shale assemblage,deposited in cold temperate shelf con-ditions, comprises a more modestassortment of fossils, including rarenaraoiid arthropods, eurypterids,straight-shelled ‘nautiloid’ cephalopods,conodont animals, and anaspid-like‘fishes’ (Aldridge et al. 2001; Seldenand Nudds 2004; Aldridge, pers. comm.2004). More recently, work on Middleand Late Ordovician sequences inAvalonian Wales has revealed sites pre-serving mixed shelly and soft-bodiedbiotas in a variety of preservationalmodes (Botting et al. 2011; Botting andMuir 2012).
In other parts of Laurentia,many of the known OrdovicianLagerstätten represent marginal marineenvironments. The Middle OrdovicianWinneshiek Lagerstätte of Iowa (Liu etal. 2006, 2011; Witzke et al. 2011) isquite diverse, including conodonts,primitive fishes, lingulid brachiopods,phyllocarid crustaceans, and euryp-terids. The Middle Ordovician rocks atDouglas Dam, Tennessee (Caster andBrooks 1956; Dunlop et al. 2004) andthe uppermost Ordovician GeorgianBay Formation of Manitoulin Island,Ontario (Stott et al. 2005) hold low-diversity assemblages of mostly che-licerate arthropods. Body fossils in theUpper Ordovician Harding Sandstoneof Colorado are diverse but poorlypreserved (Allulee and Holland 2005),and additional undescribed marginalmarine fossils are coming to light inplaces such as Wisconsin. In a muchdeeper water deposit, spectacularly pre-served trilobites occur in the famousUpper Ordovician Beecher’s TrilobiteBed of New York State (Briggs et al.1991; Etter 2002; Selden and Nudds
2004).The Manitoba sites are provid-
ing data that are not duplicated by orsubsidiary to that from any other place.Rather, every soft tissue site discoveredto date is contributing a separate, com-plementary piece of the puzzle that isOrdovician biodiversity.
CAT HEAD – McBETH POINT In the Cat Head – McBeth Point areaof Lake Winnipeg’s North Basin, theCat Head Member of the Red RiverFormation contains a considerablevariety of fossils, combining soft-tissuepreservation of certain groups withother forms representing a normalmarine mineralized biota. It is mostfamous for its remarkable assemblagesof marine algae (seaweeds), but thereare also conulariids, trilobites, lingulidbrachiopods, and many other groups.The fossils are found along the westshore of Lake Winnipeg betweenMcBeth Point, Cat Head, and LynxPoint, north and west of Kinwow Bay(Fig. 1). There, about 12 m of strataare exposed in cliffs along thelakeshore, over a horizontal distance ofabout five km (Fig. 2a). In addition,similar fossils are found at InmostIsland in the middle of Kinwow Bay(formerly known as Birch Island orKinwow Bay Island); these are proba-bly derived from slightly older beds. Allof the fossiliferous units are consideredto belong to the lower part of the CatHead Member of the Red RiverFormation, of mid Katian age (Fig. 1).
Cat Head and McBeth Pointare prominent topographic features onthe North Basin, and the latter formsan important anchorage. Thus, it ishardly surprising that these spectacularfossils were known from the early daysof collecting in this region. Fossils inthe Cat Head area were apparently firstsystematically collected by members ofGeological Survey of Canada (GSC)field parties, including T.C. Weston inthe 1880s and D.B. Dowling and L.M.Lambe in 1890s (Fig. 2a inset). Furthercollecting for the GSC was done byG.W. Sinclair and others in the 1950s.These sites were visited in 1997 bystaff from The Manitoba Museum andassociates from the Manitoba GeologicalSurvey and Royal Ontario Museum,and again by The Manitoba Museum in2006. Many other specimens now in
collections have been picked up byfishermen and pleasure boaters overthe years. The first description ofsome of the fossils from this area wasby Whiteaves (1897); shortly thereafter,Dowling (1900) produced a very gooddescription of the basic geology andincluded a substantial listing of thefossils. Sporadic paleontological studiessince that time have most notablyincluded the description of palyno-morphs, cephalopods, and sponges in acollaborative GSC Bulletin (McGregorand Cramer 1971; Flower 1971; Rigby1971) and a study of the remarkablenon-calcareous algae (Fry 1983). Majorcollections of Cat Head fossils arelocated at The Manitoba Museum, theUniversity of Manitoba, and theGeological Survey of Canada (Ottawa).
The unusual fossils in the CatHead – McBeth Point area are derivedfrom bedded fine-grained dolostonesthat lack storm deposits and extensiveburrow mottling. Chert nodules ofvariable colour and form are abundantin some beds. The lithology, and theabundance of fossils interpreted asalgae, are consistent with deposition inthe photic zone, but probably belowstorm wave base. This interpretationfits with inferred transgressive-regres-sive cycles for the Williston Basin,which place the Cat Head biota in thedeepest water conditions of the RedRiver Cycle (Elias 1991). It has, how-ever, also been suggested that the con-ditions were those of a restrictedmarine environment having little con-tact with the open sea (McGregor andCramer 1971).
Fossils, generally rare butabundant at some levels, includemacroscopic non-calcareous algae(Figs. 2a inset, 3a, 3b), possible hydro-zoans (Fig. 3c), palynomorphs, sponges(Fig. 3d), , conulariids (Fig. 3e, 3f),‘nautiloid’ cephalopods, gastropods,articulated trilobites (notably Isotelussp.; Fig. 3g), brachiopods (obolids, lin-gulids, orthids, and strophomenids; Fig.3h), , crinoids, dendroid graptolites (Fig.3i), and the problematic Sphenothallus(Whiteaves 1897; McGregor andCramer 1971; Flower 1971; Rigby1971; Fry 1983; Westrop andLudvigsen 1983; Jin and Zhan 2001;Robson and Young in press).
The soft algae (seaweeds) areamong the best to be found anywhere

GEOSCIENCE CANADA Volume 39 2012 205
in Early Paleozoic rocks, and havebeen interpreted as rhodophytes, het-erokontophytes, and chlorophytes (Fry1983). The conulariids and trilobitesare remarkable for their articulation,indicating that they were buried underlow-energy conditions. The beds with-in which fossils are abundant typicallycontain one or very few species, sug-gesting that briefly favourable condi-tions resulted in ‘blooms’ of life (Fig.3h).
Preservation of the Cat Headfossils has not been studied in detail.
Many of those with mineralized cal-careous skeletons, such as trilobitesand strophomenid brachiopods, appearto be preserved through replacementby fine-grained dolomite and chert.Phosphatic or chitinophosphatic skele-tons, such as those of conulariids andlingulid brachiopods, are wonderfullypreserved and have the appearance ofunmodified material. The seaweedsoccur as films that include carbon andiron oxide (which may have originallybeen pyrite). The preservation of theseaweeds is such that they are sensitive
to light and weather. If left exposedoutside, they fade and become more‘rusty’ as carbon is lost. Hydroids anddendroid graptolites are preserved ascarbon, apparently with some pyrite. Afew rare fossils, such as sponges, areoccasionally found as pyritic replace-ments of soft tissue within chert nod-ules.
The overall composition ofthe Cat Head biota is similar to thoseof certain younger, Silurian Lagerstättenfrom elsewhere in Laurentia. These lat-ter, termed the DG biotas by
Figure 3 (continues on page 206). Representative fossils from the Cat Head – McBeth Point area. (a) Examples of the probableheterokontophytic alga (seaweed) Winnipegia cuneata, spread across the surface of a slab (The Manitoba Museum, MM B-241). (b)The feather-like alga Dowlingia (MM B-224). (c) Part of a possible hydrozoan colony (MM I-4052). (d) The sponge Aulocopella sp.cf. A. winnipegensis (MM I-3614).( e, f) Examples of the conulariid Conularia new species (Robson and Young, in press); note finepreservation of the apex in (f) (MM I-4234 and I-4254). (g) The trilobite Isotelus sp. (MM I-229).
206
Kluessendorf (1994), are characterizedby dendroid graptolites associated withorganisms such as algae, trilobites, andsponges. The DG biotas were pre-served under a variety of conditions,but appear to be generally linked byrapid burial in places where anoxic bot-tom waters were associated withtopography. Like the Cat Head assem-blages, many of the constituents repre-sent normal-marine organisms pre-served under unusual circumstances.
WILLIAM LAKE AND AIRPORTCOVE: SHARED CHARACTERS ANDSHARED DISCOVERYThe William Lake and Airport Covesites represent distinct depositionalenvironments, but both were formedin restricted and marginal marine set-tings, in very shallow water and shore-line to nearshore conditions. Both areof latest Katian age, and they share anumber of biotic elements, though
with differing relative abundances:most notable are xiphosurids (horse-shoe crabs) and eurypterids, but thereare also non-calcareous algae, con-odonts, lingulid brachiopods, gas-tropods, and large phosphatic orchitinophosphatic tubes of uncertainaffinity (Young et al. 2007). Theseassemblages are strikingly differentfrom normal marine biotas of nearbyunits. They completely lack corals,trilobites, and echinoderms, whilerhynchonelliformean brachiopods and‘nautiloid’ cephalopods are rare.
Fossil sites such as those atWilliam Lake (Fig. 4) and Airport Cove(Fig. 5) are poorly represented in thegeological record because they form inshoreline environments that migrate,are limited in area, and have lowpreservation potential for body fossils.Both biotas occur in what initiallyappeared to be monotonous andunfossiliferous dolomudstones of gen-
erally similar character, and these unitswere largely ignored until the unusualfossils were discovered. It is, in fact,the similarities between the biotas andlithotypes that resulted in the discoveryof both sites within the past dozenyears.
Intensive study of these sitesarose from two fortuitous discoveriesof arthropod fossils. In the 1990s, J.-D.Fournier, a geologist working in theGrand Rapids Uplands, spent his sparetime searching for flat slabs of rock tobe used as substrates for artwork. Hediscovered an area where suitable slabshad been piled up, and selected appro-priate pieces on which to paint. Later,noticing fossils on the reverse sides ofslabs, he brought these to the attentionof Manitoba Survey geologist RuthBezys, who passed the information onto one of us (G.Y.; see Fig. 4e).Manitoba Museum staff first visitedthe area briefly in 2001, finding an
Figure 4 (page 207). Representative fossils from the William Lake site. (a) Large cnidarian medusa (jellyfish), probably a hydro-zoan; note quadrate form and wrinkled bell (The Manitoba Museum, MM I-4261). (b) Cnidarian medusa with traces of tentaclesto top and right of wrinkled bell (MM I-4579). (c) Three comb rows of a fossil ctenophore (comb jelly) preserved as organic mate-rial (MM I-4580). (d) A large phosphatic or chitinophosphatic tube of uncertain affinity shows a very slender base and an expand-ed aperture; the arrow points to the brachiopod Holtedahlina sp. adjacent to the side of the tube (MM I-4581). (e) This partialeurypterid abdomen is from one of the first fossiliferous slabs found near this site by J.-D. Fournier (MM I-4582). (f) This artic-ulated eurypterid moult possesses superbly-preserved appendages (MM I-4308). (g) Spines on the tip of a eurypterid prosomalappendage are preserved as organic material (MM I-4059A). (h) This fossil pycnogonid (‘sea spider’) includes a ‘head’ and articu-lated trunk segments (MM I-4282A). (i) A large specimen of the xiphosurid (horseshoe crab) Lunataspis aurora (MM I-4583). (j) Atiny juvenile of L. aurora; note the very long and slender telson (tail)(MM I-4584).
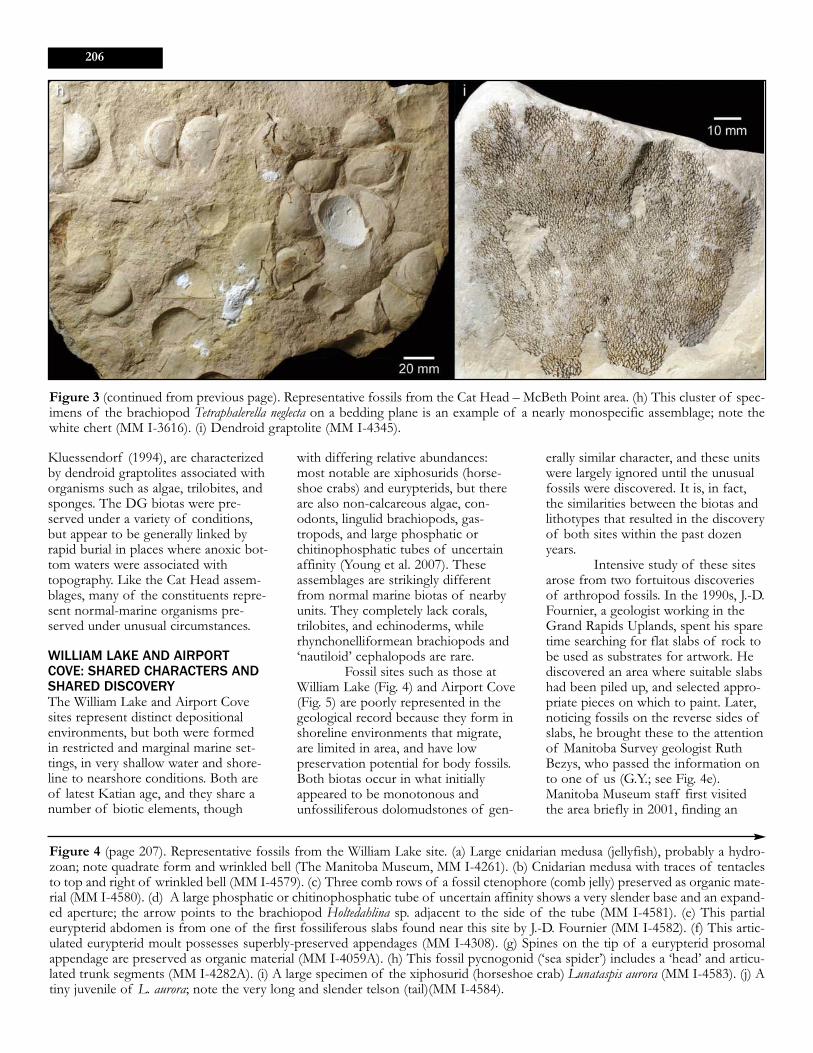
Figure 3 (continued from previous page). Representative fossils from the Cat Head – McBeth Point area. (h) This cluster of spec-imens of the brachiopod Tetraphalerella neglecta on a bedding plane is an example of a nearly monospecific assemblage; note thewhite chert (MM I-3616). (i) Dendroid graptolite (MM I-4345).

GEOSCIENCE CANADA Volume 39 2012 207

208
articulated eurypterid and other fossils,but the bedrock source for this materi-al could not be located until a muchlonger visit in the autumn of 2004.This locality, now called the WilliamLake site, has subsequently been visitedmany times, and since 2007 a systemat-ic, bed-by-bed collection has been car-ried out.
In 2000, then-student SashaHerwig was assisting Robert Elias andGraham Young with paleoecologicalfield research approximately 650 kmnortheast of William Lake, at the covenorth of Churchill Airport. Examiningloose surface material, she discoveredthe beautiful anterior shield of a smalleurypterid-like arthropod and associat-ed fragments (Fig. 5g). However, as atWilliam Lake, the bedrock sourcecould not initially be located.
At William Lake in June, 2005,three of us (E.D., S.R. and G.Y.) col-lected several unusual fossils, includingthe first articulated specimen of a crea-ture that was to be later described asthe early horseshoe crab Lunataspisaurora (Rudkin et al. 2008). Examiningthe rocks in detail, we realized thatsimilar bedrock could be found insome areas of Airport Cove. At thelatter site a few weeks later, a substan-tial area of loose angular blocks ofdolostone having the appropriateappearance was located; splitting theseblocks, within 30 minutes additionalspecimens of Lunataspis were discov-ered, along with eurypterids and a vari-ety of other fossils! The Airport Covesite was revisited in July, 2011; it nowappears certain that it consists ofbedrock that had been largely frost-wedged in situ (i.e., very little transporthad occurred), but there is no exposedoutcrop. All visible fossils at that sitehave now been collected.
All xiphosurids from bothsites have, to date, been assigned toLunataspis aurora, which is among theoldest known horseshoe crabs, repre-
senting a lineage still found in marginalmarine environments today (Rudkin etal. 2008; Rudkin and Young 2009; Figs.4i, j; 5h, i). Currently, it is difficult todetermine how many eurypterid taxaare present among the two biotasbecause the superb material shows dif-ferent modes of preservation and anumber of unusual features. Theseimportant fossils, particularly thosefrom William Lake, form a significantaddition to the sparse Ordovicianrecord of the Eurypterida (Cuggy et al.2010; Figs. 4e-g, 5e, f).
WILLIAM LAKE SITE: PALEOENVI-RONMENT AND PALEOBIOLOGYThe William Lake site is notable bothfor the remarkable fossil groups itshares with Airport Cove, and forthose that are unique to it: jellyfish andother gelatinous zooplankton, andbizarre arthropods such as pycnogo-nids (‘sea spiders’). This site is locatedon the Grand Rapids Uplands betweenGrand Rapids and Ponton, in an arearich in fossiliferous Ordovician andSilurian bedrock outcrops. The WilliamLake fossils occur in the WilliamsMember of the Stony MountainFormation (Fig. 1); the stratigraphicposition and associated conodonts(Rhipidognathus symmetricus,Drepanoistodus? sp., and Aphelognathussp.) are consistent with a LateOrdovician (late Katian) age (Young etal. 2007).
Thin-bedded dolomudstonesof the Williams Member representrestricted, shallow, and very low-energyconditions in a marginal marine envi-ronment (Fig. 2b). The conodontassemblage in this interval(Rhipidognathus biofacies) indicates theshallowest conditions for the LateOrdovician of the Williston Basin(Nowlan and Haidl 2001). The fossilif-erous beds occur within a shallowing-upward succession, above burrow-mot-tled rocks bearing normal-marine fos-
sils, and below laminated, virtuallyunfossiliferous dolomudstones inwhich some bedding planes are cov-ered with salt crystal lattices.
Over the past several years,detailed collecting has sampled aninterval about 2.5 m thick; mostLagerstätte-type fossils occur within themiddle 1.5 m of this interval. Somebed surfaces are broadly rippled orhummocky. Cut and polished verticalsections reveal parallel lamination andtrough cross-lamination; channel-likefeatures cut across the laminae inplaces. Some beds consist of monoto-nous mudstones, while others arereplete with mud rip-up clasts. Featuressuch as apparent dewatering structuresand microbial mat textures are occa-sionally visible. The fossils are general-ly rare, although narrow intervals local-ly contain common to abundant lin-gulid valves, sometimes mixed witharthropod fragments and other shellymaterial (Fig. 2c). These horizons com-monly coincide with bed partings,whereas beds between these fossil-richhorizons show less distinct internalstructure, and local lenses or layers ofmonotonous dolomudstone.
The distribution of fossilsindicates variable environmental condi-tions that were sometimesunfavourable for all fossil groups, butoccasionally favourable for particulargroups. Some groups occur throughmuch of the studied interval, althoughthey are rare in most beds, e.g. lingulidbrachiopods, eurypterids (Fig. 4e-g),gastropods, and the horseshoe crabLunataspis aurora (Fig. 4i, j). Other fos-sils are narrowly concentrated in spe-cific beds. Jellyfish, which includehydrozoan and possibly scyphozoanmedusae, are common or abundant atsome horizons in the lower to middlepart of the interval (Fig. 4a, b) butnearly absent from the upper part. Thelarge tubes of undetermined affinityalso occur only in the lower part (Fig.
Figure 5 (page 209). Representative fossils from the Airport Cove site. (a, b) Spectacular examples of dasycladalean green algaeon bedding plane surfaces; note the fine organic filaments (The Manitoba Museum, MM B-339, B-340). (c) Articulated scole-codont apparatus (jaws of a polychaete worm)(MM I-4062). (d) Large phosphatic or chitinophosphatic tubes of uncertain affin-ity (MM I-4061). (e) Finely preserved organic eurypterid cuticle (MM I-4063). (f) Partial eurypterid abdomen (MM I-4586). g.‘Head’ region of the eurypterid-like arthropod discovered by Sasha Herwig (MM I-4064A). (h) The holotype specimen of thexiphosurid (horseshoe crab) Lunataspis aurora; note the preserved eye and dark-stained areas (MM I-4000). (i) This degraded exam-ple of L. aurora is surrounded by a halo of dark staining, possibly the result of postmortem decomposition of soft tissue and leak-age of body fluids (MM I-4585).

GEOSCIENCE CANADA Volume 39 2012 209

210
4d). Probable algal fronds are commononly in a single upper bed, a thinlylaminated unit that is unusual for theoccurrence of ostracodes and tiny juve-nile specimens of Lunataspis (Fig. 4j).
Some of the strangest fossilsat William Lake are represented by ahandful of examples. Several pycnogo-nid specimens are the geologically ear-liest known adult sea spiders, and thefirst report of this group in the NorthAmerican fossil record (Rudkin et al. inpress; Fig. 4h). Remarkable fossils thatprobably represent the comb rows ofnumerous ctenophores (comb jellies)occur in a cluster within a single slab(Fig. 4c).
The fossils at William Lake arevariably preserved depending on taxo-nomic group, and on the type of stra-tum in which they occur. Those show-ing the finest preservation, themedusae (jellyfish) and articulatedarthropods, are found in largely homo-geneous layers or lenses of rapidlydeposited mudstone (Fig. 2c). Themedusae are preserved as sparry dolo-mite that is slightly enriched in ironand silica, and they are surrounded byhalos of degraded pyrite.
The original quality of preser-vation of some of the arthropod fos-sils may have been much higher, but itis likely that they lost easily weatheredorganic components as the host bedswere exposed at or near the surface forat least the past several thousand years(their upper surfaces are glacially striat-ed). The best-preserved eurypterids arelargely mouldic but include someorganic material and pyrite (Fig. 4g).Xiphosurids and pycnogonids at thissite, on the other hand, show no evi-dence of organic skeletal material;these differences are probably relatedto differing original exoskeletal struc-tures and compositions.
Lingulid brachiopods, gas-tropods, and disarticulated arthropodsare most common in thin, distincthorizons that also show evidence ofhorizontal burrows (Fig. 2c). These lay-ers may include lags that accumulatedduring times of reduced sedimentation,as channel fills, and as concentrationsin the troughs of broad ripples. Thefossils within them are generally pre-served as moulds, although patches oforganic material are occasionallyfound. Almost all lingulids occur on
these sorts of bedding plane surfaces,suggesting that they were not buried inlife position.
All evidence suggests that theWilliam Lake fossils were preserved inan intermittently harsh, unpleasantplace, possibly a ponded intertidalmudflat, or a restricted subtidal lagoon(cf. Pratt 2010; Rankey and Berkeley2012). The climate was hot, there waslittle wave or current motion, and thewater was very shallow and at timeshypersaline. The sediment was ananoxic or dysoxic fine soupy mud. It ispossible that this was not generally aplace where creatures permanentlylived; rather, it was where they lan-guished and died. This seems to be thecase for the cnidarian medusae.Horizons containing abundantmedusae may record jellyfish bloomsthat had drifted into unfavourable peri-tidal environments. The ‘halos’ aroundmany of the jellyfish may have resultedfrom anaerobic conditions associatedwith decomposition. Small horizontalburrows in the substrate very rarelyintersect the jellyfish; rather, they tendto stop outside a zone of visible dis-seminated iron oxide that surroundseach medusa. Modern blooms can besufficiently profuse that decomposingjellyfish cause dramatic temporarydeclines in oxygen content of lagoonalsediments (West et al. 2009); somebeds at William Lake may provide pos-sible evidence of this phenomenon inthe distant past.
Most of the more ‘normalmarine’ forms at William Lake, such as‘nautiloid’ cephalopods and rhyncho-nelliformean brachiopods, possessshelly hard parts that would have beenreadily shifted or floated by very slightwave or current action. It is likely thatsome of them were transported fromenvironments that had more favour-able conditions, while others may havehad specific adaptations that permittedthem to live in this unusual environ-ment (Fig. 4d).
Many of the eurypterids andhorseshoe crabs at this site are smalland may represent juveniles (Fig. 4j).These might have blundered or beenwashed into conditions where theycould not survive, or it is possible thatthe very shallow water served as a‘nursery’ from which they would havemigrated into more open and unpro-
tected waters as they grew. A lagoonalnursery situation has been suggestedfor eurypterids elsewhere (see Braddy2001). Interestingly, the majority ofarticulated eurypterid remains exhibitorientations suggesting they representmoulted individuals, rather than intactcarcasses. It is also notable that we arebeginning to see significantly largereurypterids as we collect lower in thesection, into rocks that show evidenceof having been deposited under lessharsh conditions.
The composition of theWilliam Lake biota is similar to thoseof the younger ‘EP biotas’ that occurat Upper Silurian sites in centralLaurentia (Kluessendorf 1994). EPbiotas are characterized by eurypteridsand other arthropods (sometimesincluding xiphosurans), often associat-ed with hypersaline conditions. Likethe William Lake biota, the EP organ-isms were preserved in anoxic condi-tions in peritidal paleoenvironments.
BIOTA AND LITHOLOGY AT THEAIRPORT COVE SITE The fossil assemblage at Airport Covediffers from that at William Lake in therelative abundance of beautiful sea-weeds, particularly non-calcifying dasy-cladacean green algae, the occurrenceof polychaete worms in the form ofarticulated scolecodont assemblages,and the presence of organically-pre-served arthropod cuticle (Young et al.2007). Among the shared constituents,eurypterids are notably rarer here thanat William Lake, and they are more fre-quently disarticulated. As is the case atWilliam Lake, the fossils at AirportCove are generally sparsely distributedand difficult to find, but locally con-centrated.
Airport Cove is located a fewkilometres east of the Town ofChurchill, and immediately north ofChurchill Airport (Fig. 1). Ordovicianand Silurian sedimentary rocks in thisarea surround a remarkable paleo-archipelago composed of ProterozoicChurchill quartzite (Nelson andJohnson 2002; Young et al. 2003).Elsewhere in the Hudson Bay Basin,several distinctive Upper Ordovicianlithostratigraphic units are recognized(Nelson 1963, 1964; Norford 1971;Norris 1993). In the Churchill area,however, the distinctions between units

GEOSCIENCE CANADA Volume 39 2012 211
are complicated by the archipelago’sinfluence on local sedimentation (Eliaset al. 1999).
The fossils occur in angular,relatively fresh slabs found loose withina narrowly-defined area in AirportCove (Fig. 2d). These slabs of distinc-tive greenish dolostone have yieldedconodonts consistent with a LateOrdovician (Late Katian) age:Rhipidognathus symmetricus, ?Aphelognathusdivergens, and Drepanoistodus suberectus(Young et al. 2007). Nearly identicalrocks occur near the top of a drillcoreextracted nearby within the cove, in aunit that was previously tentativelyassigned to the Churchill River Group(Young et al. 2003; Young et al. 2007).Recent analysis of the upper portionof the 100 m of section observed indrillcore from Airport Cove suggeststhat these dolostone beds occur withinan interval that may instead belong tothe Port Nelson Formation (Wheadon2011).
The lithotype in which theAirport Cove fossils are found is adolomudstone with distinctive grey-green to buff laminae 2-4 mm thick,alternating with thinner dark grey-green pyrite-rich bands (Fig. 2e). Inplaces, polished vertical surfaces showrepeated small-scale lenticular to wavybedding characteristic of tidal environ-ments (Davis 2012). Sedimentary fea-tures include apparent microbial mats,soft sediment deformation, and evap-orite crystal moulds. These features,together with the unusual macrofossilsand associated conodonts, all indicatevery shallow and restricted conditions.Since the site is almost adjacent to anangular unconformity against theProterozoic quartzites that define thecove, this was clearly a nearshore toshoreline environment, beside aquartzite paleoisland (Fig. 2d, f).
Well-preserved macrofossilstend to occur within the darker bands,while scolecodonts and small calcare-ous fossils are locally common in thepaler intervals. Macrofossils are pre-served as pyrite, as carbonate, and asorganic matter (cf. Briggs 2003). Thebest preserved fossils at Airport Coveinclude arthropods and algae, both ofwhich are largely preserved as organicmaterial. Arthropod sclerites are oftendisarticulated and broken, but cuticulardetail is remarkable (Fig. 5e). Speci-
mens of the horseshoe crab Lunataspisaurora exhibit a combination of organicmaterial, localized pyrite, and mouldicpreservation (Fig. 5h, i). Dasycladaleanalgae show very finely-detailed car-bonaceous compression preservation(Fig. 5a, b). Scolecodonts are commonon bedding planes, occasionally asarticulated apparatuses (Fig. 5c).
As is the case for WilliamLake, the Airport Cove biota is some-what similar to the younger ‘EP biotas’that occur at Upper Silurian sites incentral Laurentia (Kluessendorf 1994),though it appears that water circulationwas at times more open than that atWilliam Lake (i.e. it may have beenphysically less removed from openmarine conditions). The mixture ofarticulated and disarticulated fossils atAirport Cove may indicate variationthrough time in the amount of waveand current activity. The alternation oflaminae in the host dolostone is consis-tent with this: the darker laminae mayrepresent intervals of stagnation, orpossibly record the development ofmicrobial mats or crusts.
SIGNIFICANCEMany of the fossils recovered at thethree exceptional Manitoba sites arequite different from those found inother Late Ordovician settings, withinLaurentia and globally. They record thepresence of rare or otherwiseunknown taxonomic groups and thuscontribute key data points to ourunderstanding of rapidly expandingmarine biodiversity associated with theGOBE. They also provide criticalinsights into the evolutionary historiesof groups of organisms with lowpreservation potential. The authors aremost actively studying the WilliamLake biota, so the most detailed under-standing pertains primarily to that site,which represents deposition inextremely restricted and shallow-waterconditions. The other sites, though lesswell understood, supply complementa-ry information on different settings.
Together, William Lake andAirport Cove provide unique dataabout tropical shoreline communitiesprior to the Late Ordovician extinc-tions. Almost half the taxa (ordinallevel or higher) are rare or unknownelsewhere in the Ordovician of NorthAmerica. Considering taxonomic
groups, 33% of fossil groups atWilliam Lake are calcareous, in contrastwith normal marine carbonates lower inthe Stony Mountain Formation, where95% are calcareous (Young et al. 2010).In total numbers, fossils with calcare-ous skeletons are remarkably rare atWilliam Lake and Airport Cove, mak-ing up only four to five percent of thetotal numbers of macrofossils at each.
Many of the fossils found atWilliam Lake and Airport Cove belongto still-extant groups such as horseshoecrabs and lingulid brachiopods. As weconsider these biotas, it is easy to fallinto the trap of thinking of them asthough they represent environmentsvery similar to those found in modernshoreline conditions. We must not,however, lose sight of the fact that theworld in which these organisms livedwas very different, with a higher globalmean surface temperature, loweratmospheric oxygen, extremely highatmospheric CO2 (possibly up to 15xmodern pre-industrial levels), consider-able volcanic activity, and global sealevels up to 220 m above modern lev-els (Trotter et al. 2008; Servais et al.2009)! The occurrence of these verylong-lived groups is not evidence ofunchanging conditions. Rather, itdemonstrates the considerable adapt-ability and resilience of these organ-isms.
All three sites include manyforms that are not preserved in stan-dard normal marine Late Ordovicianassemblages, and each has the potentialto improve our understanding of bio-diversity during this critical interval inEarth’s history.
ACKNOWLEDGEMENTSFunding for this research was providedby NSERC, The Manitoba MuseumFoundation, and the Royal OntarioMuseum Governors. We are grateful toNorman Aime, Peter Fenton, MarionFoster, Brian Iwama, ChristineKaszycki, Lori Stewart, MarjorieTurton, and David Wright, for theirparticipation in fieldwork at one ormore of these fossil sites. We wouldlike to thank Ruth Bezys, J.-D.Fournier, and Sasha Herwig for theirrole in the initial discovery of theWilliam Lake and Airport Cove sites.We have appreciated discussions ofthese rocks and fossils with many indi-

212
viduals, including Nicholas Butterfield,Robert Elias, and Whitey Hagadorn.David Greenwood is thanked for pro-viding kind access to microscope facili-ties at Brandon University, and for dis-cussions. Our research has benefitedfrom Godfrey Nowlan’s conodontanalyses, and from collaboration withthe Manitoba Geological Survey and itsdrill crew (Doug Berk, Gerry Benger,and Vio Varga), both of which con-tributed to the stratigraphic and pale-oenvironmental understanding of theWilliam Lake and Airport Cove sites.This manuscript was improved consid-erably thanks to thorough reviews byJakob Vinther and Joe Botting.
REFERENCESAldridge, R.J., Gabbott, S.E., and Theron,
J.N., 2001, The Soom Shale, in Briggs,D.E.G., and Crowther, P.R., eds.,Palaeobiology II: Blackwell Science,Oxford, UK, p. 340-342,http://dx.doi.org/10.1002/9780470999295.ch79.
Allulee, J.L., and Holland, S.M., 2005, Thesequence stratigraphic and environ-mental context of primitive verte-brates: Harding Sandstone, UpperOrdovician, Colorado, USA: Palaios, v. 20, p. 518-533,http://dx.doi.org/10.2110/palo.2003.p03-120.
Bezys, R.K., 1996, Sub-Paleozoic structurein Manitoba’s northern Interlake alongthe Churchill Superior BoundaryZone: A detailed investigation of theFalconbridge William Lake study area:Manitoba Energy and Mines OpenFile OF94-3, 32 p.
Botting, J.P., Muir, L.A., Sutton, M.D., andBarnie, T., 2011, Welsh gold: A newexceptionally preserved pyritizedOrdovician biota: Geology, v. 39, p. 879-882,http://dx.doi.org/10.1130/G32143.1.
Botting, J.P., and Muir, L.A., 2012, Faunaand ecology of the holothurian bed,Llandrindod, Wales, UK (Darriwilian,Middle Ordovician), and the oldestarticulated holothurian:Palaeontographic Electronica, v. 15(1): 9A, 28 p.
Braddy, S.J., 2001, Eurypterid palaeoecolo-gy: palaeobiological, ichnological andcomparative evidence for a‘mass–moult–mate’ hypothesis:Palaeogeography, Palaeoclimatology,Palaeoecology, v. 172, p. 115-132,http://dx.doi.org/10.1016/S0031-0182(01)00274-7.
Briggs, D.E.G., Bottrell, S.H., and Raiswell,R., 1991, Pyritization of soft-bodied
fossils: Beecher’s trilobite bed, UpperOrdovician, New York State: Geology,v. 19, p. 1221-1224,http://dx.doi.org/10.1130/0091-7613(1991)019<1221:POSBFB>2.3.CO;2.
Briggs, D.E.G., 2003, The role of decayand mineralization in the preservationof soft-bodied fossils: Annual Reviewof Earth and Planetary Sciences, v. 31,p. 275-301,http://dx.doi.org/10.1146/annurev.earth.31.100901.144746.
Caster, K.E., and Brooks, H.K., 1956, Newfossils from the Canadian – Chazyan(Ordovician) hiatus in Tennessee:Bulletins of American Paleontology, v. 36, p. 157-199.
Cuggy, M.B., Rudkin, D.M., and Young,G.A., 2010, A new Late Ordovicianeurypterid from the William LakeLagerstatte, Manitoba, Canada: ThirdInternational PalaeontologicalCongress, London, Programme &Abstracts, p. 134.
Davis, R.A., Jr., 2012, Tidal signatures andtheir preservation potential in strati-graphic sequences, in Davis, R.A., Jr.and Dalrymple, R.W., eds., Principlesof Tidal Sedimentology: Dordrecht,Springer, p. 35-55,http://dx.doi.org/10.1007/978-94-007-0123-6_3.
Dowling, D.B., 1900, Report on the geolo-gy of the west shore and islands ofLake Winnipeg: Geological Survey ofCanada, Annual Report (New Series),1898, Part F, p. 1F-100F.
Dunlop, J.A., Anderson, L.I., and Braddy,S.J., 2004, A redescription ofChasmataspis laurencii Caster & Brooks,1956 (Chelicerata: Chasmataspidida)from the Middle Ordovician ofTennessee, USA, with remarks onchasmataspid phylogeny: Royal Societyof Edinburgh Transactions, EarthSciences, v. 94, p. 207-225.
Elias, R.J., 1981, Solitary rugose corals ofthe Selkirk Member, Red RiverFormation (late Middle or UpperOrdovician), southern Manitoba:Geological Survey of Canada, Bulletin344, 53 p.
Elias, R.J., 1991, Environmental cycles andbioevents in the Upper OrdovicianRed River – Stony Mountain solitaryrugose coral province of NorthAmerica: Geological Survey ofCanada Paper 90-9, p. 205-211.
Elias, R.J., Young, G.A., Nowlan, G.S.,Dobrzanski, E.P., and Rudkin, D.M.,1999, Ordovician – Silurian boundarysection discovered near Churchill,Manitoba: Preliminary report: ActaUniversitatis Carolinae – Geologica
1999, v. 43(1/2), p. 229-232.Etter, W., 2002, Beecher’s Trilobite Bed:
Ordovician pyritization for the otherhalf of the trilobite, in Bottjer, D. J.,Etter, W., Hagadorn, J.W. and Tang,C.M., eds., Exceptional FossilPreservation: A Unique View on theEvolution of Marine Life: ColumbiaUniversity Press, New York, p. 131-141.
Flower, R.H., 1971, Cephalopods of theOrdovician Cat Head Member, LakeWinnipeg, Manitoba: GeologicalSurvey of Canada Bulletin, 202, p. 13-33.
Fry, W.L., 1983, An algal flora from theUpper Ordovician of the LakeWinnipeg region, Manitoba, Canada:Review of Palaeobotany andPalynology, 39, p. 313-341,http://dx.doi.org/10.1016/0034-6667(83)90018-0.
Jin, J., and Zhan, R., 2001, Late Ordovicianarticulate brachiopods from the RedRiver and Stony Mountain formations,southern Manitoba: National ResearchCouncil Research Press, Ottawa, 117 p.
Kluessendorf, J., 1994, Predictability ofSilurian Fossil-Konservat-Lagerstätten inNorth America: Lethaia, v. 27, p. 337-344, http://dx.doi.org/10.1111/j.1502-3931.1994.tb01584.x.
Liu, H.P., McKay, R.M., Young, J.N.,Witzke, B.J., McVey, K.J., and Liu, X.,2006, A new Lagerstätte from theMiddle Ordovician St. PeterFormation in northeast Iowa, USA:Geology, v. 34, p. 969-972,http://dx.doi.org/10.1130/G22911A.1.
Liu, H.P., Witzke, B.J., Briggs, D.E.G., andMcKay, R., 2011, Primitive fish fromthe Middle Ordovician WinneshiekLagerstätte of northeast Iowa, USA:Geological Society of AmericaAbstracts with Programs, v. 43, no. 5,p. 545.
McGregor, D.C., and Cramer, F.H., 1971,Palynomorphs of the Ordovician CatHead Member, Lake Winnipeg,Manitoba, in Contributions to CanadianPaleontology: Fossils of the OrdovicianRed River Formation (Cat HeadMember), Manitoba: Geological Surveyof Canada, Bulletin 202, p. 1-11.
Nelson, S.J., 1963, Ordovician paleontologyof the northern Hudson BayLowland: Geological Society ofAmerica, Memoir 90, 152 p.
Nelson, S.J., 1964, Ordovician stratigraphyof northern Hudson Bay Lowland,Manitoba: Geological Survey ofCanada, Bulletin 108, 36 p.
Nelson, S.J., and Johnson, M.E., 2002, JensMunk Archipelago: Ordovician-Silurian islands in the Churchill area ofthe Hudson Bay Lowlands, northern

GEOSCIENCE CANADA Volume 39 2012 213
Manitoba: The Journal of Geology, v. 110, p. 577-589, http://dx.doi.org/10.1086/341757.
Norford, B.S., 1971, Silurian stratigraphy ofnorthern Manitoba, in GeoscienceStudies in Manitoba: GeologicalAssociation of Canada Special PaperNo. 9, p. 199-207.
Norris, A.W., 1993, Hudson Platform –Geology, in Stott, D.F. and Aitken,J.D., eds., Sedimentary Cover of theCraton in Canada: Geological Surveyof Canada, Geology of Canada(Seventh Edition), no. 5, p. 653-700.
Nowlan, G.S., and Haidl, F.M., 2001,Biostratigraphy and paleoecology ofLate Ordovician conodonts from acomposite section in the subsurface ofSaskatchewan, in Christopher, J.E.,Gilboy, C.F., Haidl, F.M., and Kreis,L.K., eds., Summary of Investigations2001, Volume 1: SaskatchewanGeological Survey, SaskatchewanEnergy and Mines, MiscellaneousReport 2001-4.1, p. 14-31.
Pratt, B.R., 2010, Peritidal carbonates, inJames, N.P. and Dalrymple, R.W., eds.,Facies Models 4: GeologicalAssociation of Canada, Geotext 6, p. 401-420.
Rankey, E.C., and Berkeley, A., 2012,Holocene carbonate tidal flats, in Davis,R.A., Jr. and Dalrymple, R.W., eds.,Principles of Tidal Sedimentology:Dordrecht, Springer, p. 507-535,http://dx.doi.org/10.1007/978-94-007-0123-6_19.
Rigby, J.K., 1971, Sponges of theOrdovician Cat Head Member, LakeWinnipeg, Manitoba, in Contributionsto Canadian Paleontology: Fossils ofthe Ordovician Red River Formation(Cat Head Member), Manitoba:Geological Survey of Canada, Bulletin202, p. 35-79.
Robson, S.P., and Young, G.A., in press,Late Ordovician conulariids fromManitoba, Canada. Journal ofPaleontology.
Rudkin, D.M., and Young, G.A., 2009,Horseshoe crabs – an ancient ancestryrevealed, in Tanacredi, J.T., Botton,M.L., and Smith, D., eds., Biology andConservation of Horseshoe Crabs:Springer-Verlag, p. 25-44,http://dx.doi.org/10.1007/978-0-387-89959-6_2.
Rudkin, D.M., Young, G.A., and Nowlan,G.S., 2008, The oldest horseshoe crab:A new xiphosurid from LateOrdovician Konservat-Lagerstättendeposits, Manitoba, Canada.
Palaeontology, v. 51, p. 1-9,http://dx.doi.org/10.1111/j.1475-4983.2007.00746.x.
Rudkin, D.M., Cuggy, M.B., Young, G.A.,and Thompson, D.P., in press, AnOrdovician pycnogonid (sea spider)with serially subdivided ‘head’ region:Journal of Paleontology.
Selden, P., and Nudds, J., 2004, Evolutionof Fossil Ecosystems: The Universityof Chicago Press, Chicago, 160 p.
Servais, T., Harper, D.A.T., Li, J., Munnecke,A., Owen, A.W., and Sheehan, P.M.,2009, Understanding the GreatOrdovician Biodiversification Event(GOBE): Influences of paleogeogra-phy, paleoclimate, or paleoecology?:GSA Today, v. 19, p. 4-10, http://dx.doi.org/10.1130/GSATG37A.1.
Stott, C.A., Tetlie, O.E., Braddy, S.J.,Nowlan, G.S., Glasser, P.M., andDevereux, M.G., 2005, A neweurypterid (Chelicerata) from theUpper Ordovician of ManitoulinIsland, Ontario, Canada: Journal ofPaleontology, v. 79, p. 1166-1174,http://dx.doi.org/10.1666/0022-3360(2005)079[1166:ANECFT]2.0.CO;2.
Trotter, J.A., Williams, I.S., Barnes, C.R.,Lécuyer, C., and Nicoll, R.S., 2008,Did cooling oceans trigger Ordovicianbiodiversification? Evidence fromconodont thermometry: Science, v. 321, p. 550-554, http://dx.doi.org/10.1126/science.1155814.
Van Roy, P., 2006, Non-trilobite arthropodsfrom the Ordovician of Morocco:unpublished Ph.D. dissertation, GhentUniversity, 230 p.
Van Roy, P., Orr, P.J., Botting, J.P., Muir,L.A., Vinther, J., Lefebvre, B., elHariri, K., and Briggs, D.E.G., 2010,Ordovician faunas of Burgess Shaletype: Nature, v. 465, p. 215-218,http://dx.doi.org/10.1038/nature09038.
Vinther, J., Van Roy, P., and Briggs, D.E.G.,2008, Machaeridians are Palaeozoicarmoured annelids: Nature, v. 451, p.185-188, http://dx.doi.org/10.1038/nature06474.
Webby, B.D., 2004, Introduction, in Webby,B.D., Paris, F., Droser, M.L., andPercival, I.G., eds., The GreatOrdovician Biodiversification Event:Columbia University Press, New York,p. 1-37.
Webby, B.D., Paris, F., Droser, M.L., andPercival, I.G., eds., 2004, The GreatOrdovician Biodiversification Event:Columbia University Press, New York,484 p.
West, E.J., Welsh, D.T., and Pitt, K.A.,
2009, Influence of decomposing jelly-fish on the sediment oxygen demandand nutrient dynamics: Hydrobiologia,v. 616, p. 151-160, http://dx.doi.org/10.1007/s10750-008-9586-7.
Westrop, S.R., and Ludvigsen, R., 1983,Systematics and paleoecology ofUpper Ordovician trilobites from theSelkirk Member of the Red RiverFormation, southern Manitoba:Manitoba Department of Energy andMines, Mineral Resources Division,Geological Report GR 82-2, 51 p.
Wheadon, B., 2011, Ordovician-Silurianboundary interval near Churchill,Manitoba: Lithostratigraphy,chemostratigraphy and biostratigraphyof Airport Cove west core and quarrysamples: Unpublished B.Sc. Honoursthesis, Department. of GeologicalSciences, University of Manitoba, 159 p.
Whiteaves, J.F., 1897, The fossils of theGalena – Trenton and Black River for-mations of Lake Winnipeg and itsvicinity: Geological Survey of Canada,Palaeozoic Fossils, v. 3, no. 3, p. 129-242, pls. 16-22.
Witzke, B.J., McKay, R.M., Liu, H.P., andBriggs, D.E.G., 2011, The MiddleOrdovician Winneshiek Shale ofnortheast Iowa – correlation and pale-ogeographic implications: GeologicalSociety of America Abstracts withPrograms, v. 43, no. 5, p. 315.
Young, G.A., Elias, R.J., and Dobrzanski,E.P., 2003, GS-23 Paleozoic drillcorefrom the Churchill area, northernManitoba: Preliminary results (NTS54K and 54L): Manitoba GeologicalSurvey, Report of Activities 2003, p. 171-177.
Young, G.A., Rudkin, D.M., Dobrzanski,E.P., Robson, S.P., and Nowlan, G.S.,2007, Exceptionally preserved LateOrdovician biotas from Manitoba,Canada: Geology, v. 35, p. 883-886,http://dx.doi.org/10.1130/G23947A.1.
Young, G.A., Elias, R.J., Wong, S., andDobrzanski, E.P., 2008, UpperOrdovician rocks and fossils in south-ern Manitoba: Winnipeg, CanadianPaleontology Conference Field TripGuidebook No. 13, 97 p.
Young, G.A., Rudkin, D.M., Dobrzanski,E.P., Robson, S.P., Cuggy, M., andThompson, D.P., 2010, LateOrdovician Lagerstätten in Manitoba,Canada: Glimpses of soft-bodieddiversity: The Third InternationalPalaeontological Congress, London,Abstracts.
Received November 2012Accepted as revised November 2012