m. d. herrero-pe´rezrul Æ f. garcı´a-domı´nguez Æ...
TRANSCRIPT

M. D. Herrero-Pe rezrul á H. Reyes BonillaF. Garcõ a-Domõ nguez á C. E. Cintra-Buenrostro
Reproduction and growth of Isostichopus fuscus (Echinodermata:Holothuroidea) in the southern Gulf of California, MeÂxico
Received: 15 July 1998 /Accepted: 19 August 1999
Abstract The reproduction and growth of Isostichopusfuscus (Ludwig, 1875) at Isla Espõ ritu Santo, Baja Cali-fornia Sur, Me xico from July 1992 to September 1993and from August 1996 to July 1997 were analyzed. Thereproductive cycle was monitored by using histologicalanalysis. Five gonadal stages were described accordingto cell types present in the gonad; undetermined, ga-metogenesis, maturity, spawning and post-spawning.This holothurian reproduces annually during summer(July to September), when sea surface temperaturereaches 27 °C. The gonad index is related to the gonadalstages. The ®rst sexual maturity for I. fuscus was cal-culated at 367 g (4 to 5 years old) according to the vonBertalan�y equation, and nearly all the specimens at IslaEspõ ritu Santo were 5 to 7 years old (including someindividuals over 35 years old). Data of the length±weightrelationship showed that I. fuscus grows allometrically atthe studied site (b = 1.83). The mortality rate was rel-atively high (M = 0.51), indicating that in a typicalcohort, 40% of the recruits does not survive the ®rstyear, and less than 20% reaches maturity.
Introduction
The holothurian Isostichopus fuscus is a common speciesthat inhabits shallow waters (0 to 40 m, mostly on coraland rocky bottoms) from the northern Gulf of Califor-nia, Me xico to Ecuador (Maluf 1988). This holothurianrepresents an important economic resource and supports
artisanal ®sheries in several American countries, princi-pally Me xico, Costa Rica and Ecuador (Powell andGibbs 1995; Fajardo Leo n and Ve lez Barajas 1996;Singh Cabanillas and Ve lez Barajas 1996; Son-nenholzner 1997). In all these countries, it has beensuggested that the populations of I. fuscus and otherholothurian species have been overexploited (Camhi1995; Fajardo Leo n and Ve lez Barajas 1996). In Me xico,I. fuscus is considered an endangered species (NormaO®cial Mexicana 1994), though no studies were made tosupport this. In general, studies of the biology andpopulation dynamics of this holothurian are scarce. InMe xico, most of the early papers dealt only with tax-onomy and ecology (Caso 1967), but more recentlystudies about the reproductive biology and brief ana-lyses of the ®shery of this holothurian along the south-ern coasts of the Gulf of California have been made(Sing Cabanillas and Velez Barajas 1996; Fajardo Leo net al. 1995).
Most of the studies on reproductive biology of ho-lothurians have focused on tropical and commercialspecies (Bakus 1973; Conand 1990), which commonlybelong to the Families Stichopodidae (Cameron andFankboner 1986; Conand 1988, 1993a, b) and Ho-lothuriidae (Engstrom 1980; Harriot 1985; Ong Che andGo mez 1985). The objective of the present paper was todescribe the main patterns of the reproductive cycle ofIsostichopus fuscus and some aspects of its populationbiology. The study was done at Isla Espõ ritu Santo, inthe southern Gulf of California, Me xico.
Materials and methods
Specimens of Isostichopus fuscus (Ludwig, 1875) were collected atIsla EspõÂ ritu Santo, Gulf of California (24°30¢N; 110°15¢W), par-ticularly in San Gabriel Bay, which is characterized by shallowwaters (maximum depth 6 m) and two large coral patch reefscomposed of Pocillopora spp. with occasional appearances ofmassive colonies (Porites spp. and Pavona spp.: Reyes Bonilla1993), which are exposed during extreme low tides in summer. Atthe shore there is a mangrove forest composed of Rhizophora
Marine Biology (1999) 135: 521±532 Ó Springer-Verlag 1999
Communicated by O. Kinne, Oldendorf/Luhe
M.D. Herrero-Pe rezrul (&) á F. Garcõ a-Domõ nguezCentro Interdisciplinario de Ciencias Marinas,Calle Madero 1865-2, Centro,C.P. 23000 La Paz, Baja California Sur, Me xico
H. Reyes Bonilla á C.E. Cintra-BuenrostroUniversidad Auto noma de Baja California Sur,Depto. de Biologia Marina, Carretera al Sur km 5.5,C.P. 23080 La Paz, Baja California Sur, Me xico

mangle. Sea surface-water temperature ranges from 20 to 30 °C inthe bay.
Reproduction analysis
For the reproduction analysis, sampling was done monthly fromJuly 1992 to September 1993. A total of 259 specimens (�20ind mo)1) were collected by hand and ®xed in 10% seawater-for-malin. Each individual was kept inside a separate plastic bag tokeep the gonads from mixing in case of evisceration. Once thegonads were dissected, measured, weighed and stored in 70% al-cohol, they were histologically processed and embedded in para�n.Sections (7 lm thick) were stained with haematoxylin-eosin andexamined under a microscope (10 and 20´). The reproductive cycleof Isostichopus fuscus was monitored by using histological exami-nation of the gonadal stages and by changes in the gonad index(GI), calculated with the equation: GI = (gonad weight/drainedweight) ´ 100 (Giese and Pearse 1974). To compare the monthlyvariations of the GI in both sexes, a one-way ANOVA test wasused (a = 0.05). As GI is a percentage, the arcsin transformation(Sokal and Rohlf 1995) was used to attain normality and homo-scedasticity of data. The Tukey a posteriori test (Day and Quinn1989) was used to determine the origin of the statistical di�erences,when appropriate. Lastly, to test the hypothesis that the sex ratiowas not signi®cantly di�erent from 1:1, a v2 goodness-of-®t analysiswas carried out (Sokal and Rohlf 1995).
The size to attain ®rst sexual maturity in this species was cal-culated plotting the cumulative percentage of individuals whichwere in gametogenesis, maturity or spawning stage, against length.The point of the curve where 50% of the individuals were in thosestages was considered the weight at ®rst maturity (Conand 1990).We considered drained weight (DW) a more reliable measure forthe analysis, because length is highly variable in holothurians. Dryweight is a still better choice to estimate ®rst sexual maturity(Conand 1982, 1988, 1990), but the use of this parameter impliesthe loss of the gonad, an unacceptable condition for this study.
Growth analysis
For growth determination, length measurements of a monthlymean of 120 individuals were obtained in six locations around IslaEspõÂ ritu Santo, from August 1996 to July 1997 (total = 1446 in-dividuals). Specimens were measured underwater from mouth toanus with ¯exible tapes (�0.5 cm precision) trying to avoid bodycontraction. They were then put into individual plastic bags andtaken to a boat, where the weight of each was determined withspring scales. The search for specimens was haphazard, looking atopen sites on the sandy bottom and near rocky and coral reef areas,as well as under large coral heads (Pocillopora spp.), boulders andcoral rubble. Once length and weight were measured, all sampledholothurians were returned to the bottom in areas delimited bynatural markings, with the purpose of collecting in approximatelythe same locations every month and thus measuring as many of thesame organisms as possible each time. As in the case of gonadindex, di�erences of weight and length among months were de-termined with ANOVA (a = 0.05) and Tukey tests.
Size-frequency data (grouped in 2-cm intervals) and its monthlymodal composition were analyzed to calculate the parameters ofthe von Bertalan�y growth model (Ricker 1975) using the com-puter package FiSAT, programmed by the FAO and ICLARM(Gayanillo et al. 1996). The following routines were used: Powell±Wetherall (for calculation of mean asymptotic length, Linf), Shep-herd (to estimate K, individual growth coe�cient), and Pauly'sempirical equation (to obtain t0, the hypothetical age when growthis zero). Further details of all the analytical techniques used andtheir respective assumptions were presented by Pauly and Morgan(1987), Sparre and Venema (1995) and Gayanillo et al. (1996). The®nal model for growth in length was arranged to also estimategrowth in weight, on the basis of von Bertalan�y's length±weightequation calculated by a non-linear regression of all data (Ricker1975; Sparre and Venema 1995).
The constants, Linf and K, and mean sea surface temperature inthe studied area (averaged from monthly measurements in 1992 to1993, and data from 1960 to 1990 included in the CD-ROM CO-ADS, produced by the NOAA) were used to estimate the naturalmortality rate (M) of the population with Pauly's empirical model(Sparre and Venema 1995). In turn, this rate was used in the generalequation for determination of population size in a non-exploitedstock: (Nt = N0 � e()M*t)), where t is the number of years (Ricker1975) to calculate yearly percentage of survival in a ``typical'' cohort.
Finally, to more precisely determine the von Bertalan�y modelconstants and mortality rates, the con®dence intervals of Linf, Winf
(asymptotic weight), K and M were calculated with the jacknifemethod, in which each parameter was recomputed 11 times, ig-noring one observation in turn each time (Sokal and Rohlf 1995).
Results
Isostichopus fuscus is a dioecious species. There is noevidence of external sexual dimorphism, and the gonadscan only be sexed by microscopic examination. Whenripe, the gonads consist of a single tuft of numerousunbranched tubules that lie on the anterior end of thecoelom. After spawning, the gonad decreases in size by>90%. Of the 259 specimens dissected for reproductionanalysis, only 173 (66.7%) had visible gonads: 64 weremales, 78 females, 29 undetermined and 2 hermaphro-dites. The annual sex ratio was not signi®cantly di�erentfrom 1:1 (v2 = 20.27, df = 12, P > 0.05).
On the basis of morphological and histological ob-servations, ®ve gonadal stages were described for Iso-stichopus fuscus: undetermined, gametogenesis(oogenesis and spermatogenesis), mature, spawning andpost-spawning. These stages showed an annual period-icity and were related to the GI variations.
Undetermined (Fig. 1a)
In this stage, no gametes could be distinguished. Allundetermined gonads were similar. We found no evi-dence of gametes. The presence of abundant connectivetissue was observed in the lumen and between the folli-cles, which were collapsed. It was common to observesmall phagocytes (about 3 lm diameter). The gonadwall was thick. The GI values were <2% at this stage(see Fig. 4). Undetermined gonads were seen from Oc-tober to January. A total of 86 specimens found duringthis period had no gonads, but it is possible that theywere undetermined gonads.
Developmental stages of the male
Spermatogenesis (Fig. 1b)
In the gonad wall, longitudinal folds were observed,extending towards the lumen. At the periphery of thefollicles, rounded spermatocytes proliferated. Sperm-atocytes were ovate (<1 lm diameter) and stronglystained with haematoxylin. Some mature spermatozoa
522

were found in the lumen. The connective tissue wasthinner and the follicles increased their size because ofaccumulation of sperm. Spermatogenesis was seen fromFebruary to June (Fig. 3a).
Maturity (Fig. 1c)
All the follicles were completely ®lled with dense layersof mature spermatozoa, which had rounded heads
Fig. 1 Isostichopus fuscus. Photomicrographs of gonadal stages ofthe males at Isla EspõÂ ritu Santo (July 1992 to September 1993). aUndetermined, b spermatogenesis, c maturity, d spawning, e post-spawning (c connective tissue; f follicles; g gonad wall; phphagocytes; sp spermatocytes; s mature spermatozoa). Scale bars100 lm
523

(about 1 lm diameter). The gonad wall was thin anddistended, and at the periphery, a thin layer of sperm-atocytes was present. In this stage, some spherule cellswere observed. Male gonads appeared in July to August(Fig. 3a). This stage showed the highest GI values 8 to12% (Fig. 4).
Spawning (Fig. 1d)
Spawned gonads showed empty spaces inside the folli-cles because spermatozoa had already been released.Some developing spermatocytes were still observed atthe periphery. On the gonad wall, the presence of a layerof connective tissue was seen. Spawning occurred duringsummer, and the GI values decreased by almost one-halfin this stage (Figs. 3a, 4).
Post-spawning (Fig. 1e)
The post-spawning gonads were reabsorbed by phago-cytes. In the gonad wall of the follicles, abundant con-nective tissue was observed. Some follicles still presentedmature spermatozoa. Numerous groups of phagocyteswere observed inside and outside the follicles. Post-spawning gonads were observed in November, with lowGI values (Figs. 3a, 4).
Developmental stages of the female
Oogenesis (Fig. 2a)
The oogonia were embedded in the germinal epithelium,and this made them di�cult to identify. The oogonialcells began to proliferate towards the lumen. Developingoocytes were arranged in a single layer. Numerous pre-vitellogenic oocytes (20 lm average diameter) wereobserved in this stage, each having a large nucleus, withtwo or three peripheric nucleoli, and being surroundedby small follicular cells. The presence of spherule cellswas seen during this stage. Gametogenesis was observedfrom January to June (Fig. 3b). The GI values for thisstage were low, 2 to 4% (Fig. 4).
Maturity (Fig. 2b)
Mature oocytes reached their maximum size (104 lmdiameter) and occupied the entire lumen of all tubules.The oocyte membranes were separated from the cyto-plasm because of tissue contraction caused by the ®xa-tion process. At the periphery, there were somedeveloping oocytes. The nucleus and nucleoli are wellde®ned in the cells. In this stage, it was common to ob-serve groups of phagocytes forming spherule cells insidethe tubules. Male gonads appeared from June to Augustand in females from July to August (Fig. 3b). The GIvalues for this stage were high, 8 to 12% (Fig. 4).
Spawning (Fig. 2c)
There was a noticeable decrease in the abundance ofoocytes inside the tubules. Almost all were ripe, thougha few developing oocytes were still present near the go-nad wall. Large, empty spaces were observed in the lu-men because of the absence of gametes. A fewphagocytes were observed. Spawning occurred duringsummer (July to September). The GI values decreasedby one-half in this stage (Fig. 4).
Post-spawning (Fig. 2d)
The few oocytes that were observed in this stage haddecreased in volume. The presence of many phagocytesinside and outside the tubules was detected, and theiractivity was related to a signi®cant decrease in the size ofthe gonads. The phagocytes had a granular appearance.Large amounts of connective tissue were seen in thegonad walls. Post-spawning female gonads were ob-served in October and November, with very low GIvalues (Figs. 3b, 4).
Reproductive cycle
The reproductive cycle of Isostichopus fuscus showed aclear annual pattern for both sexes. Spawning occurredfrom July to September in 1992 and in 1993. Post-spawning gonads were found in October of both years.Undetermined gonads were found from October toJanuary, but because their frequency of occurrence waslow, we did not include this stage in the graph (Fig. 3a,b). Gametogenesis was observed from January to June1993, and females started gametogenesis a month earlierthan males (Fig. 3a, b).
The female and male GI evolution had a signi®cantrelation with sea surface temperature variations andwith the gonadal stages. There was no signi®cantdi�erence between GI values of males and females(ANOVA: F2,142 = 2.036, P > 0.05). The highest GIvalues coincided with mature gonads which were foundduring summer, the warmest months of the year. The GIvalues for this stage were <12%. During spawning, theGI values diminished when sea surface temperature wasup to 27 °C (Fig. 4). The lowest GI values correspondedto post-spawning, undetermined and early gametogenicgonads, which were found during the winter months(October to January).
Monthly di�erences in the GI values were detected(ANOVA: F12,171 = 53.01, P < 0.05). A Tukey testrecorded three groups; the ®rst included the coldermonths, from October 1992 to January 1993, whichshowed GI values <1% (undetermined and post-spawning stages). The second group had the highest GIvalues, corresponding to July and August of both years,when gonads were ripe. The third group was represented
524

by August and September 1992, when most specimenshad spawned and had post-spawning gonads; in thesemonths, GI values were 1% to 5% (Fig. 4).
The weight at ®rst maturity (when 50% of the indi-viduals in the population showed developing gonads)was calculated considering only specimens that could besexed (n = 142) and that were in gametogenic, matureor spawning stage. The smallest reproductive specimenweighed 167 g, but the calculated ®rst-maturity weightwas 367 g (Fig. 5).
Growth
The population of Isostichopus fuscus at Isla EspõÂ rituSanto had an average (�SE) length and weight of
23.25 � 0.26 cm and 385.97 � 7.81 g, with modes of24 cm and 350 g (Fig. 6; Table 1). Because ®rst matu-rity was attained at 367 g, size-frequencies indicatedthat most of the sampled specimens were actively inreproduction. The smallest individuals (<5 cm and<50 g) were observed from August to November 1996,and again from May to July 1997. Mean size of thespecies di�ered signi®cantly among months (F11, 1434 =8.85, P < 0.001 for length; F11,1434 = 8.87, P < 0.001for weight); individuals were especially heavier andlonger in January, February and April, 1997 (Table 1).The length±weight regression was signi®cant(r2 = 0.714, F1,1144 = 3607.5, P < 0.001), and showedthat I. fuscus grows allometrically at the studied site(Fig. 7).
The estimated von Bertalan�y growth equations forlength and weight were:
Lt � 36:118�1ÿ �e�0:180�tÿ0:024����;Wt � 815:485�1ÿ �e�0:180�tÿ0:024����1:832 ;
where Lt is length in time t (years), Wt is weight in timet (years), 36.118 = Linf (asymptotic length in cm;
Fig. 2 Isostichopus fuscus. Photomicrographs of gonadal stages offemales at Isla EspõÂ ritu Santo (July 1992 to September 1993). aOogenesis, b maturity, c spawning, d post-spawning (c connectivetissue; do developing oocytes; f follicles; fc follicular cells; g gonadwall; n nucleus; o ripe oocytes; ph phagocytes; po previtellogenicoocytes). Scale bars 100 lm
525
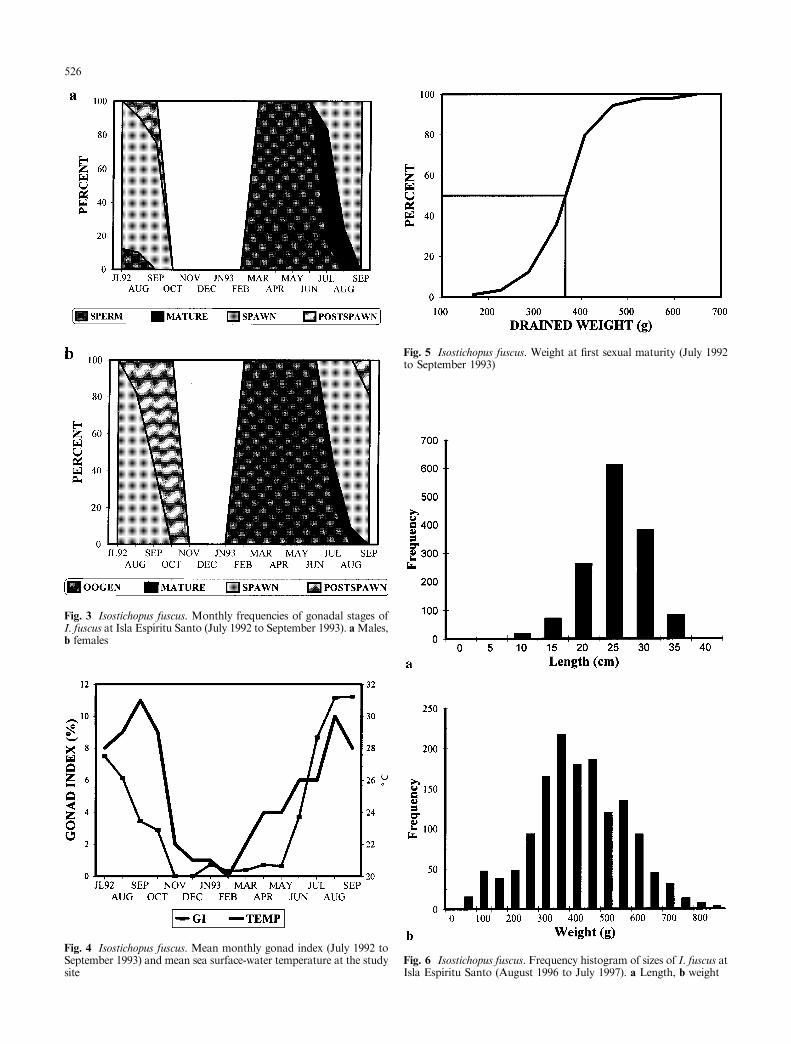
Fig. 3 Isostichopus fuscus. Monthly frequencies of gonadal stages ofI. fuscus at Isla EspõÂ ritu Santo (July 1992 to September 1993). aMales,b females
Fig. 4 Isostichopus fuscus. Mean monthly gonad index (July 1992 toSeptember 1993) and mean sea surface-water temperature at the studysite
Fig. 5 Isostichopus fuscus. Weight at ®rst sexual maturity (July 1992to September 1993)
Fig. 6 Isostichopus fuscus. Frequency histogram of sizes of I. fuscus atIsla EspõÂ ritu Santo (August 1996 to July 1997). a Length, b weight
526

con®dence interval: �0.244), 815.485 = Winf (asymp-totic weight in g; con®dence interval: �5.509),0.180 = K (individual annual growth rate; con®denceinterval: �0.044), 0.024 = t0 (estimated age whengrowth is zero) and 1.832 = b (allometry coe�cientcalculated with the length±weight relation; con®denceinterval: �0.003).
Graphic representations of the models (Fig. 8)showed that in this population, individuals with sizes of<6 cm or 40 g were younger than a year, whereas thelargest specimens (>35 cm and 700 g) were very scarce,and were about 15 years old. The population mean and
modal values included individuals 5 to 6 years old, but 7-year-old animals were also very abundant in the area(Fig. 9). An age of 5 years also signals the time for ®rstmaturity in Isostichopus fuscus.
The estimated natural mortality of the population was0.51 � 0.03. Survival percentage abruptly decreasedduring the ®rst year (about 40% of original recruits diewithin that time), and during the following 2 years(Fig. 10). By age ®ve, <10% of the cohort remains.
Discussion
Reproduction
The holothurian Isostichopus fuscus is a dioecious spe-cies. Specimens exhibit no sexual dimorphism, exceptwhen the gonad is ripe. Female gonads are pale orange,whilst male gonads are whitish. The population sex ratiowas approximately 1:1, as is the case for most aspido-chirotes (Cameron and Fankboner 1986; Conand 1993a;Foster and Hodgson 1995). The gonochoric condition isa common feature of holothurians, although completeor casual hermaphroditism has been reported (Harriot
Table 1 Isostichopus fuscus. Length (L, in cm) and weight (W, in g)data of the population from Isla Espõ ritu Santo, Me xico
Month n Mean Mode SE Min. Max.
Aug 1996 W 97 362.1 480 11.4 15 525L 97 23.3 23 0.4 7 33
Sep W 105 383.3 410 12.5 45 660L 105 24.3 26 0.5 9 35
Oct W 113 348.7 410 11.6 20 650L 113 23.1 22 0.4 5 33
Nov W 103 366.9 320 13.4 20 790L 103 21.9 24 0.4 5 30
Dec W 99 389.9 340 11.7 105 700L 99 23.9 26 0.4 16 33
Jan 1997 W 107 444.9 310 13.7 65 800L 107 24.6 24 0.5 12 40
Feb W 131 470.2 420 13.0 70 810L 131 25.9 26 0.4 11 36
Mar W 152 391.0 320 10.6 120 690L 152 23.4 24 0.4 11 34
Apr W 108 411.3 380 14.9 70 760L 108 23.3 28 0.4 11 34
May W 134 381.1 350 15.3 45 830L 134 22.5 26 0.4 9 33
Jun W 164 340.1 330 14.9 20 810L 164 21.4 25 0.5 7 35
Jul 1997 W 133 353.9 290 13.1 45 760L 133 22.1 22 0.4 8 33
Overall W 1446 385.97 350 3.99 15 830L 1446 23.25 24 0.13 5 40
Fig. 7 Isostichopus fuscus. Length±weight relationship of I. fuscus atIsla EspõÂ ritu Santo (August 1996 to July 1997). Equation:Weight = 1.142 (Length)1.832; con®dence intervals: a =1.142 � 1.100; b = 1.832 � 0.003
Fig. 8 Isostichopus fuscus. Individual growth in a length and b weightof I. fuscus at Isla EspõÂ ritu Santo (August 1996 to July 1997)
527

1982; Tyler et al. 1985; Tyler and Billet 1987; HerreroPe rezrul et al. 1998).
The histological analysis of the gonad of Isostichopusfuscus showed that its characteristics resembled thosedescribed previously for other aspidochirotes (Harriot1982, 1985; Ong Che and Go mez 1985; Smiley 1988;Tuwo and Conand 1992; Fajardo Leo n et al. 1996). Inthe undetermined stage, there was no evidence ofgametogenic activity (Fig. 1a). The gonad was so smallthat it became very di�cult to see, a situation that hasbeen previously reported in studies of other holothurians(Hyman 1955; Bakus 1973; Conand 1981). Although theabsence of gonads due to evisceration processes ismentioned in the literature (Bakus 1973), evidence ofsuch a situation was not found in our study; except forthe gonads, all organs were present. At Isla Espõ rituSanto, we found 83 specimens of I. fuscus without go-nads. Most of these individuals were found in the colder
months (October to January), a time when the popula-tion was characterized by the presence of individuals inpost-spawning and undetermined gonadal stages(Fig. 3a, b).
The presence of spherule cells in the early stages ofdevelopment (gametogenesis) in females has also beennoted in other holothurians; the phagocyted material isused to nurture other developing gametes (Hyman 1955;Boolotian 1966; Costelloe 1985, 1988; Smiley and Clo-ney 1985; Tyler and Billet 1987; Sewell and Bergquist1990; Sewell 1992; Hammel et al. 1993). After spawning,the gonads of I. fuscus decreased in size because ofphagocytosis, and by January the gonad was practicallyreabsorbed. Also during gametogenesis, the germinallayer of the gonads of I. fuscus became thinner, andlongitudinal folds extended to the lumen. This process iscommon in the Class Holothuroidea and produces asigni®cant increase in the surface area for developmentof gametes (Costelloe 1985; Tyler et al. 1985; Cameronand Fankboner 1986).
It was common to observe the presence of gametes inboth female and male gonads during the post-spawningstage. This may show that partial events of sheddingtake place. Mosher (1982) showed that some ho-lothurians are capable of spawning more than onceduring the reproductive period, as a way of maximizingreproductive success. This strategy might be very usefulfor Isostichopus fuscus, because its oocytes are very small(100 lm) and size is a factor that decreases the proba-bility of fertilization (Levitan 1993). Repeated events ofgamete expulsion, possibly triggered by proximity toconspeci®cs, would help to avoid this problem.
The spawning of invertebrates is under the in¯uenceof endogenous and exogenous factors. Temperatureseems to be one of the most important variables thatdetermines the timing of reproduction (Boolotian 1966;Giese and Pearse 1974). In the tropical eastern Paci®c,the reproductive seasonality of other echinodermscoincides with high temperatures and upwellings(Strathmann 1985; Lessios 1990). Isostichopus fuscusshowed a clear annual reproductive pattern at IslaEspõ ritu Santo, in which this holothurian spawned dur-ing summer. In this season, sea surface temperature was27 to 30 °C. Although other aspidochirotes also repro-duce in months with high water temperatures or insummer (Table 2), this does not seem to pervadethroughout the family, as reproduction can also occurduring cold months (Conand 1993b), twice a year(Krishnaswamy and Krishnan 1967; Ong Che andGo mez 1985), or during the whole year (Hyman 1955).It is probable that temperature only synchronizes thetiming of spawning in I. fuscus, as occurs in other easternPaci®c holothurians (Cameron and Fankboner 1986).
Growth
Mean length and weight of the Isostichopus fuscuspopulation from Isla EspõÂ ritu Santo (23.3 cm and
Fig. 9 Isostichopus fuscus. Age structure of the population from IslaEspõÂ ritu Santo (August 1996 to July 1997)
Fig. 10 Isostichopus fuscus. Survival percentage of a model cohort(August 1996 to July 1997)
528

385.9 g; Table 1) were lower than values reported forthe species in the central and northern Gulf of Cali-fornia (27° to 29°N; 25.1 to 32.0 cm and 458 to 562 g:Fajardo Leo n et al. 1995; Fajardo Leo n and Ve lezBarajas 1996), but higher than in Oaxaca, Me xico(16°N; 351 g: Fajardo Leo n et al. 1995) and Gala pagos(0°N; 20 cm and 271 g: Sonnenholzner 1997), indicat-ing a north±south size gradient for this holothurian. Itis doubtful that this tendency is directly caused by suchdi�erences in environmental conditions as productivityor mean temperature, since these oceanographic factorsdo not follow latitudinal trends in the cited locations ofthe eastern Paci®c (Fiedler 1992). The appearance oflarge specimens in the north can be considered justanother example of a marine invertebrate which in-creases in size with latitude (Atkinson and Sibly 1997).Conand (1990) presented mean length and weight datafor some of the most important commercial ho-lothurians of the South Paci®c; values ranged from 22to >40 cm and 270 to >400 g. The size of I. fuscus isnoticeably smaller than that of most ``®rst rate'' taxa,however, the American species has been well receivedin Asiatic markets, in part because its price is relativelylow (Conand 1997).
In recent decades, several approaches to estimateholothurian growth have been tried, e.g. tagging andenclosure of specimens (Conand 1990; Chao et al. 1994;Wiedemeyer 1994). Unfortunately, these techniqueshave some methodological constraints that confoundtheir use. For example, animals usually expel tags, aprocess which can cause physical damage and tissuenecrosis (Ebert 1978; Conand 1990). Keeping specimensin restricted spaces may a�ect growth-rate determina-tions, because food can be a limiting factor when indi-viduals grow, or food may be more scarce or abundantat the experimental sites than in the natural environ-ment. Because of these limitations, the use of vonBertalan�y±like numerical models to study growthof holothurians and other echinoderms has been fa-
vored (Zann et al. 1987; Ebert and Russell 1993).These models are based on three assumptions: that thespecies grow isometrically, recruit annually and underliea size limit (Ricker 1975; Sparre and Venema 1995). Ingeneral, commercial holothurians reproduce and recruitonce every year, and seem to reach determinate size(Conand 1990; Lawrence 1987), but they show allo-metric growth (Shelley 1985; Conand 1988; Bulteel et al.1992). Populations of Isostichopus fuscus from the Gulfof California are no exception to the general pattern,and because of this lack of strict adherence to the as-sumptions of the models, traditional ®shing-stock as-sessment methods can only be considered a heuristicalternative for the estimation of age and growth for thisspecies, and for holothurians in general. Consequently,the results of such studies (and the derived conclusions)have to be regarded with caution.
Several studies on the growth of tropical commercialholothurians have been published (Table 3). As ob-served, maximum length and growth rates of Isostich-opus fuscus are similar and even greater than those ofseveral important taxa, an indication of its adequacy asa commercial species. The speci®c values of the vonBertalan�y parameters for each holothurian are deter-mined by the life history and local selective pressures ofeach species and population, and therefore they are noteasily comparable. However, considering that smallerspecies like Holothuria scabra or Actynopyga echinitesare currently under cultivation (Conand 1990; Anony-mous 1997), it is feasible to suggest aquaculture as analternative for ®shermen interested in commercializingI. fuscus. In this way, the strong di�erences in opinionbetween the government and the ®shermen of Me xico,Ecuador and other American countries concerning theexploitation of this resource can be worked out.
In the Gulf of California and most of Me xico,Isostichopus fuscus was under strong ®shing pressurefrom 1989 to 1994, a situation that depleted populations,a�ecting above all animals larger than 20 to 25 cm
Table 2 Reproductive characters of holothurian species of the Family Stichopodidae (± no data)
Species Location Reproductiveseason
Temperature forreproduction
GI (%) Sex ratio Oocytediameter (lm)
Reference
Parastichopusparvimensis
SouthernCalifornia
May±Jun ± ± ± ± Kastendiek andMuscat (1982)
Stichopuscalifornicusa
Washington ± ± ± ± 200 Smiley andCloney (1985)
P. californicus 49°21¢N Jul±Aug 16±18 32 1:1 180 Cameron andFankboner (1986)
S. mollis 36°19¢S Nov±Jan ± ± ± 180 Sewell (1992)S. variegatus 20°S Dec±Apr 24±26 5 1:1 180 Conand (1993a)S. variegatus 20°S Dec±Apr 24±26 2.8 (males)
2.7 (females)1:1 180 Conand (1993b)
Neostichopusgrammatus
33°58¢S Oct±Jan 22±23 6.9 1:1 350 Foster andHodgson (1995)
P. parvimensis 31°40¢N Apr±May ± 3.9 1:1 ± Pe rez Plascencia(1995)
Isostichopusfuscus
24°35¢N Jul±Sep 28±31 11.2 1:1 104 Present study
aStichopus californicus is synonymous with Parastichopus californicus
529

(Fajardo Leo n and Ve lez Barajas 1996) which, accord-ing to the present model, were older than 4 years(Fig. 9). In 1996 and 1997, the most abundant cohorts ofthe species at Isla Espõ ritu Santo were 5 to 7 years old(Fig. 9), meaning they were recruited between 1989 and1993, years when the ®shery was active. It is probablethat individuals were too small to be captured then andconsequently they survived. This ®nding suggests thatthe current age and size structure of the local populationof I. fuscus re¯ects the e�ects of human activities.
Another important consideration related to age isthat individuals attain sexual maturity at age ®ve(Fig. 9). Taking into account that the number of adultholothurians was much reduced in the southern gulf inthe early 1990s because of ®shing mortality, it is possiblethat a sizeable portion of the local population is nowproducing gametes for the ®rst time in the decade. If thisconclusion is correct, an increase in local population sizemay become apparent at Isla EspõÂ ritu Santo and prob-ably elsewhere in America in the coming years.
In the southern gulf, a subtropical area, Isostichopusfuscus attains ®rst reproduction at small weights (367 g,Fig. 5). In the tropical western Paci®c, other commercialaspidochirotes reach their ®rst sexual maturity at sizes>500 g dry weight (Conand 1981, 1982, 1990) andalong the northern Paci®c coast of Baja California,Me xico (31° N), Parastichopus parvimensis reaches ma-turity at 160 g (Pe rez Plascencia 1995). These data, al-though still minimal, point to an inverse gradientbetween ®rst-maturity weight and latitude (and possiblysea surface temperature), a tendency which provides aninteresting line of future investigation regarding man-agement of resources as important as holothurians.
In marine species mortality rate is related to asymp-totic length and to age at maturity, which, in turn,represent a group of parameters dependent on size(Jensen 1997; Sparre and Venema 1995). The estimatednatural mortality of Isostichopus fuscus at Isla EspõÂ rituSanto (M = 0.51, Fig. 10) is among the lowest ratesknown for commercial taxa (Table 3), which indicatesthat the species may be more resistant to natural factorsthat cause death, like predation. There are just a fewspecies known to feed on holothurians (Birkeland 1989;Wiedemeyer 1994), but none live in the eastern Paci®c, a
situation that may explain the lack of chemically activeagents for defense in I. fuscus (Encarnacio n-Dimayugaand Keer-Garcõ a 1992) and its high survival probability.The life history characteristics of this sea cucumber arethose of a K-strategist; I. fuscus has low mortality andgrowth rates, a late start of sexual maturity, large sizeand great longevity (Conand 1988). The common ap-pearance of these traits in a species is only the conse-quence of several active agents working together (Jensen1997), needing no particular explanation for their com-mon occurrence.
Acknowledgements The Direccio n de Estudios de Posgrado eInvestigacio n del Instituto Polite cnico Nacional, and Comisio nNacional para el Conocimiento y Uso de la Biodiversidad (ProjectFB315/H216/96) granted funds for this work . The Comisio n deOperacio n y Fomento de Actividades Acade micas fellowship wasawarded to F. Garcõ a-Domõ nguez, and the Programa Nacionalpara la Formacio n de Investigadores (PIFI-IPN) and the ConsejoNacional de Ciencia y Tecnologõ a (Number 82643) scholarships toM.D. Herrero-Pe rezrul.
References
Anonymous (1997) Successful production of juvenile sand®shHolothuria scabra by ICLARM in the Solomon Islands. Beà che-de-mer Inf Bull 9: 3±4
Atkinson D, Sibly RM (1997) Why are organisms usually bigger incolder environments? Making sense of a life history puzzle.Trends Ecol Evolut 12: 235±239
Bakus JG (1973) The biology and ecology of tropical holothurians.In: Jones OA, Endean R (eds) Biology and geology of coralreefs. Vol. 2. Academic Press, New York, pp 325±367
Birkeland C (1989) The in¯uence of echinoderms on coral-reefcommunities. In: Jangoux M, Lawrence JM (eds) Echinodermstudies. Vol. 3. Balkema, Rotterdam, pp 1±79
Boolootian RA (1966) Reproductive physiology. In: BoolootianRA (ed) Physiology of Echinodermata. John Wiley and SonsInterscience, New York, pp 561±614
Bulteel P, Jangoux M, Coulon P (1992) Biometry, bathymetricdistribution and reproductive cycle of the holothuridHolothuriatubulosa (Echinodermata) from Mediterranean seagrass beds.Publ Staz zool Napoli (I: Mar Ecol) 13: 53±62
Cameron JL, Fankboner PV (1986) Reproductive biology of thesea cucumber Parastichopus californicus (Stimpson) (Echi-nodermata: Holothuroidea). I. Reproductive periodicity andspawning behavior. Can J Zool 64: 168±175
Camhi M (1995) Industrial ®sheries threaten ecological integrity ofthe Gala pagos Islands. Conserv Biol 9: 715±724
Table 3 Comparison of published parameters of the von Bertalan�y growth model calculated for holothurians (ND no data)
Species Latitude Linf (cm) K (annual) M (annual) Reference
Holothuria atra 11°N 32.4 0.11 1.02 Ebert (1978)Actynopyga echinites 20°S 23.1 0.78 ND Shelley (1985)A. echinites 20°S 23.0±29.5 0.09±0.30 0.64 Conand (1988)A. mauritania 20°S 34.0 0.12 1.45 Conand (1988)Stichopus japonicus 34°N 36.7 0.33 ND Hamano et al. (1989)Thelenota ananas 20°S 66.3 0.20 0.50±0.63 Conand (1990)S. chloronotus 20°S 34.2 0.45 1.79 Conand (1990)H. scabra 5°S 29.0 0.52 ND Pauly et al. (1993)H. pulla 5°S 30.7 0.24 ND Pauly et al. (1993)Opheodesma spectabilis 5°S 32.6 0.18 ND Pauly et al. (1993)Parastichopus parvimensis 31°N 52.5 0.30 ND Pe rez Plascencia (1995)Isostichopus fuscus 24°N 36.1 � 0.2 0.18 � 0.04 0.51 � 0.03 Present study
530

Caso ME (1967) Morfologõ a interna y ecologõ a de Stichopus fu-scus Ludwig. An Inst Biol Univ Me x (Ser Cienc Mar) 37: 175±182
Chao SM, Chien CP, Alexander PS (1994) Reproduction andgrowth of Holothuria atra (Echinodermata: Holothuroidea) attwo contrasting sites in southern Taiwan. Mar Biol 119: 565±570
Conand C (1981) Sexual cycle of three commercially importantholothurian species (Echinodermata) from the lagoon of NewCaledonia. Bull mar Sci 31: 523±543
Conand C (1982) Reproductive cycle and biometric relations in apopulation of Actinopyga echinites (Echinodermata: Ho-lothuroidea) from the lagoon of New Caledonia, westerntropical Paci®c. In: Lawrence JM (ed) Proc. Int. EchinodermConf. Tampa Bay. Vol. 1. Balkema, Rotterdam, pp 437±442
Conand C (1988) Comparison between estimations of growth andmortality of two stichopodid holothurians: Thelenota ananasand Stichopus chloronotus (Echinodermata: Holothuroidea). In:Choat JH et al. (eds) Proc. 6th Int. Coral Reef Symp. Vol. 2.Symposium Executive Committee, Townsville, pp 661±665
Conand C (1990) The ®sheries resources of Paci®c island countries.Part 2. Holothurians. FAO Fish Tech Pap 272.2: 1±143
Conand C (1993a) Ecology and the reproductive biology ofStichopus variegatus, an Indo Paci®c coral reef sea cucumber(Echinodermata: Holothuroidea). Bull mar Sci 52: 970±981
Conand C (1993b) Reproductive biology of the holothurians fromthe major communities of the New Caledonian Lagoon. MarBiol 116: 439±450
Conand C (1997) Are holothurian ®sheries for export sustainable?In: Lessios HA, MacIntyre IG (eds) Proc. 8th Int. Coral ReefSymp. Vol. 2. Smithsonian Tropical Research Institute, Balboa,Panama, pp 2021±2026
Costelloe J (1985) The annual reproductive cycle of the holothurianAslia lefevrei (Dendrochirota: Echinodermata). Mar Biol 88:155±165
Costelloe J (1988) Reproductive cycle, development and recruit-ment of two geographically separated populations of the den-drochirote holothurian Aslia lefevrei. Mar Biol 99: 535±545
Day RW, Quinn GP (1989) Comparisons of treatments after ananalysis of variance in ecology. Ecol Monogr 59: 433±463
Ebert TA (1978) Growth and size of the tropical sea cucumberHolothuria (Halodeima) atra Jaeger, at Eniwetok Atoll, Mar-shall Islands. Pacif Sci 32: 183±191
Ebert TA, Russell MP (1993) Growth and mortality of subtidal redsea urchins (Strongylocentrotus franciscanus) at San NicolasIsland, California, USA: problems with models. Mar Biol 117:79±89
Encarnacio n-Dimayuga R, Keer-Garcõ a S (1992) Compuestos conactividad antimicrobiana de organismos marinos. Revta Socmex Cienc farmac 22: 33±41
Engstrom AN (1980) Reproductive cycles of Halodeima ¯oridanaHolothuria mexicana and their hybrids (Echinodermata:Holothuroidea) in southern Florida, USA. Int J Invert Reprod2: 237±244
Fajardo Leo n MC, Michel Guerrero E, Singh Cabanillas J, Ve lezBarajas JA, Masso Rojas A (1995) Estructura poblacional yciclo reproductor del pepino de mar Isostichopus fuscus enSanta Rosalõ a, B.C.S., Me xico. Cienc Pesq 11: 45±53
Fajardo Leo n MC, Ve lez Barajas A (1996) Pesquerõ a del pepino demar. In: Casas-Valdez MC, Ponce-Dõ az G (eds) Estudio delpotencial pesquero y acuõ cola de Baja California Sur. Vol. 1.SEMARNAP/ CICIMAR, LaPaz, Me xico, pp 151±165
Fiedler PC (1992) Seasonal climatologies and variability of easterntropical Paci®c surface waters. NOAA tech Rep NMFS 109: 1±34
Foster GG, Hodgson AN (1995) Annual reproductive cycles ofthree sympatric species of intertidal holothurians (Echinoder-mata) from the coast of the eastern Cape Province of SouthAfrica. Invert Reprod Dev 27: 49±59
Gayanillo FC, Sparre P, Pauly D (1996) FAO-ICLARM stockassessment tools (FiSAT). FAO Computerized InformationSeries (Fisheries), No. 8, FAO, Rome
Giese CA, Pearse JS (1974) Reproduction of marine invertebrates.Vol. 1. Academic Press, New York
Hamano T, Amio M, Hayashi KI (1989) Population dynamics ofStichopus japonicus Selenka (Holothuroidea: Echinodermata) inan intertidal zone and on the adjacent subtidal bottom witharti®cial reefs for Sargassum. Suisanzoshoku 37: 179±186
Hammel JF, Himmelman JH, Dufresne L (1993) Gametogenesisand spawning of the sea cucumber Psolus fabricii (Duben andKoren). Biol Bull mar biol Lab, Woods Hole 184: 125±143
Harriot VJ (1982) Sexual and asexual reproduction of Holothuriaatra Jaeger at Heron Island Reef, Great Barrier Reef. Aust MusSyd Mem 16: 53±66
Harriot VJ (1985) Reproductive biology of three congeneric seacucumber species, Holothuria atra, H. impatiens and H. edulis,at Heron Reef, Great Barrier Reef. Aust J mar Freshwat Res36: 51±57
Herrero Pe rezrul MD, Reyes Bonilla H, Garcõ a Domõ nguez F(1998) Casual hermaphroditism in gonochoric Isostichopus fu-scus (Ludwig, 1875) (Echinodermata: Holothuroidea) in thesouthern Gulf of California, Mexico. Bull mar Sci 63(3): 611±615
Hyman LH (1955) The invertebrates: Echinodermata, the coelo-mate bilateria. McGraw Hill, New York
Jensen AL (1997) Origin of the relationship between K and Linf andsynthesis of relationships among life history parameters. Can JFish aquat Sciences 54: 987±990
Kastendiek J, Muscat A (1982) Aspects of the biology of the seacucumber Parastichopus parvimensis: a developing commercial®shery. Univ South Calif Sea Grant Inst Prog A Rep 1: 63±66
Krishnaswamy S, Krishnan S (1967) A report on the reproductivecycle of the holothurian Holothuria scabra Jaeger. Curr Sci 6:155±156
Lawrence JM (1987) A functional biology of echinoderms. CroomHelm, London
Lessios HA (1990) Adaptation and phylogeny as determinants ofegg size in echinoderms from the two sides of the Isthmus ofPanama. Am Nat 135: 1±73
Levitan RD (1993) The importance of sperm limitation to theevolution of egg size in marine invertebrates. Am Nat 141: 517±536
Maluf LY (1988) Composition and distribution of the centraleastern Paci®c echinoderms. Nat Hist Mus Los Angel Cty TechRep 2: p 242
Mosher C (1982) Spawning behavior of the aspidochirote ho-lothurian Holothuria mexicana Ludwig. In: Lawrence JM (ed)Proc. Int. Echinoderm Conf. Tampa Bay. Balkema, Rotterdam,pp 467±468
Norma O®cial Mexicana (1994) Determinacio n de las especies ysubespecies de ¯ora y fauna silvestre terrestres y acua ticas enpeligro de extincio n, amenazadas, raras y las sujetas a pro-teccio n especial, y que establece especi®caciones para su pro-teccio n. NOM-059-ECOL-1994, Diario O®cial de laFederacio n, Me xico, pp 2±59
Ong Che RG, Go mez ED (1985) Reproductive periodicity of Ho-lothuria scabra Jaeger at Catalan, Batangas, Philippines. Asianmar Biol 2: 21±30
Pauly D, Morgan GR (eds) (1987) Length-based methods in ®sh-eries research. ICLARM-Kuwait Institute for Scienti®c Re-search, Sicily
Pauly D, Sambilay V, Opitz S (1993) Estimates of relative foodconsumption by ®sh and invertebrate populations required formodeling the Bolinao reef ecosystem, Philippines. In: Chris-tensen V, Pauly D (eds) Trophic models of aquatic ecosystems.ICLARM Conf. Proc. 26. ICLARM, Manila, pp 236±251
Pe rez Plascencia G (1995) Crecimiento y reproduccio n del pepinode mar Parastichopus parvimensis en la Bahõ a de Todos Santos,Baja California, Me xico. M.Sc. thesis, Universidad Auto nomade Baja California, Me xico
Powell JR, Gibbs JP (1995) A report from Gala pagos. Trends EcolEvolut 10(9): 351±354
Reyes Bonilla H (1993) Biogeogra®a y ecologia de los coraleshermatipicos (Anthozoa: Scleractinia) del Paci®co de Me xico.
531

In: Salazar Vallejo SI, Gonza lez NE (eds) Biodiversidad marinay costera de Me xico. Commisio n Nacional de la Biodiversidad(CONABIO)/Centro de Investigaciones de Quintana Roo(CIQRO), Cheturnal, pp 207±222
Ricker WC (1975) Computation and interpretation of biologicalstatistics of ®sh populations. Bull Fish Res Bd Can 191: 1±382
Sewell MA (1992) Reproduction of the temperate aspidochiroteStichopus mollis (Echinodermata: Holothuroidea) in New Zea-land. Ophelia 2: 103±121
Sewell MA, Bergquist PR (1990) Variability in the reproductivecycle of Stichopus mollis (Echinodermata: Holothuroidea).Invert Reprod Dev 17: 1±7
Shelley C (1985) Growth of Actynopyga echinites and Holothuriascabra (Holothuroidea: Echinodermata) and their ®shery po-tential (as Beà che-de-mer) in Papua, New Guinea. In: Gabrie Cet al. (eds) Proc. 5th Int. Coral Reef Congr. Vol. 5. AntenneMuseum ± EPHE, Moorea, French Polynesia, pp 297±302
Singh Cabanillas J, Ve lez Barajas A (1996) La pesquerõ a del pepinode mar Isostichopus fuscus en la costa oriental de Baja Cali-fornia Sur y propuestas de regulacio n. Cienc Pesq 12: 13±18
Smiley S (1988) The dynamics of oogenesis and the annual ovariancycle of Stichopus californicus (Echinodermata: Holothuroidea).Biol Bull mar biol Lab, Woods Hole 175: 79±93
Smiley S, Cloney RA (1985) Ovulation and a ®ne structure of theStichopus californicus (Echinodermata: Holothuroidea) fecundovarian tubules. Biol Bull mar biol Lab, Woods Hole 169: 342±364
Sokal RR, Rohlf FJ (1995) Biometry, 3rd edn. W.H. Freeman andCompany, New York
Sonnenholzner J (1997) A brief survey of the commercial sea cu-cumber Isostichopus fuscus (Ludwig, 1875) in the Gala pagosIslands. Beà che-de-mer Inf Bull 9: 11±15
Sparre P, Venema SC (1995) Introduccio n a la evaluacio n de re-cursos pesqueros tropicales. FAO Doc. Tec. Pesca 306/1.DANIDA-FAO, Rome
Strathmann RR (1985) Feeding and nonfeeding larval developmentand life-history evolution in marine invertebrates. A Rev EcolSyst 16: 339±361
Tuwo A, Conand C (1992) Reproductive biology of the ho-lothurian Holothuria forskali (Echinodermata). J mar biol AssUK 72: 745±758
Tyler PA, Billet DSM (1987) The reproductive ecology of ela-sipodid holothurians from the North East Atlantic. BiolOceanogr 4: 273±296
Tyler PA, Muirhead A, Gage JD, Billet DSM (1985) Reproductivebiology of the deep-sea holothurians Laetmogone violacea andBenthogone rosea (Elasipoda: Holothuroidea). Mar Ecol ProgSer 23: 269±277
Wiedemeyer WL (1994) Biology of small juveniles of the tropicalholothurian Actynopyga echinites: growth, mortality and habi-tat preferences. Mar Biol 120: 81±93
Zann L, Brodie J, Berryman C, Nagasima M (1987) Recruitment,ecology, growth and behavior of juvenile Acanthaster planci (L.)(Echinodermata: Asteroidea). Bull mar Sci 41: 561±575
532