nonhistone proteins of the oocyte nucleus …jcs.biologists.org/content/joces/15/1/145.full.pdf ·...
TRANSCRIPT

J. Cell Sci. i s , 145-161 (1974) 145Printed in Great Britain
NONHISTONE PROTEINS OF THE OOCYTE
NUCLEUS OF THE NEWT
R. J. HILL,* K. MAUNDRELL AND H. G. CALLAN
Department of Zoology, The University, St Andrezvs, Fife, Scotland
SUMMARY
Evidence has been obtained which indicates that disulphide bond crosslinks contribute to themorphological integrity of isolated lampbrush chromosomes (both chromomeres and lateralloops) and nucleoli. It is suggested that the progressive formation of these bonds in vitro byaerial oxidation may provide the basis for the previously recognized time-dependent hardeningor 'denaturing' of these structures.
Manually isolated germinal vesicle nuclei have been massed and fractionated by low-speedcentrifugation into nucleoplasm and chromatin. Phase-contrast microscopy demonstrates thechromatin to consist of nucleoli, lampbrush chromosomes and nuclear membranes.
Urea gel electrophoresis has been employed to resclve the reduced and 5-carboxymethylatedproteins of whole nuclei into some 12 components, negatively charged at pH 8. The nucleoplasmalone gives an essentially similar pattern, but with the distinct depletion of one component andslight depletion of another. Both of these components are much enriched in the chromatin pelletwhere they predominate over all other proteins.
The total chromatin has been subfractionated by microdissection, taking advantage of thedifferential attachment of nucleoli to the nuclear membrane at different stages of oogenesis. It isconcluded that the nuclear membrane per se does not contribute to the major chromatin proteins.The two major polypeptides are components of the nucleoli. Preparations of isolated lampbrushchromosomes have not, to date, provided sufficient material to give a distinctive electrophero-gram; only one faint band, a major component of whole nuclei, was apparent.
Sodium dodecyl sulphate gel electrophoresis has resolved some 25 components in wholenuclei, and again demonstrates the enrichment of the two major species in the total chromatinfraction. The apparent molecular weights of these two species are 43 kilodaltons andn o kilodaltons. Approximately 20 minor species are also present in the chromatin and areobviously good candidates as components of the nucleolar and chromosomal structures. Histones,at most, make only a minor contribution to the overall chromatin protein population.
INTRODUCTION
The problem of assignment of intranuclear location and function to the nonhistoneproteins of the eukaryotic cell nucleus is very much in a state of flux at present (see e.g.MacGillivray, Carroll & Paul, 1971; Teng, Teng & Allfrey, 1971; Hill, Poccia & Doty,1971; Elgin & Bonner, 1972). These proteins probably include structural componentsof the chromosomes and nuclear membrane, enzymes, perhaps' informatin' in nascentRNA-protein combination (Georgiev & Samarina, 1972) and, as suggested by theexperiments of Paul & Gilmour (1968), specific gene activators and/or repressors. Inview of this diversity of function, it is not surprising that nonhistone protein populationshave, in general, been found to display a high degree of complexity.
* Present address: CSIRO Division of Animal Genetics, P.O. Box 90, Epping, N.S.W. 2121,Australia.
10 C E L 15

146 R. J. Hill, K. Maundrell and H. G. Callan
Nonhistone proteins of the newt germinal vesicle offer a potentially interesting focusof inquiry. This giant nucleus may be isolated by microdissection in an operationmonitored directly under the microscope and performed much more rapidly thanclassical bulk biochemical separations. Furthermore, there is much cytologicalinformation available; within the isolated nucleus peripheral nucleoli and a centralmass of lampbrush chromosomes are demonstrable by phase microscopy (Gall, 1954;Callan & Lloyd, i960). Both the lampbrush chromosomes and nucleoli have beenshown to consist predominantly of protein (Macgregor & Callan, 1962; Isawa, Allfrey& Mirsky, 1963; Macgregor, 1972).
There has long been circumstantial evidence suggesting that proteins of the germinalvesicle play an important role in embryogenesis (Brachet, 1938). Recent reports haveprovided more direct evidence that this nucleus contains relatively high concentrationsof morphogenetic determinants, and that these, at least in part, consist of protein(Briggs & Justus, 1968; Malacinski, 1972).
Further interest in the proteins of the germinal vesicle has been engendered by theremarkable visualization of transcription complexes in newt oocyte nucleoli andlampbrush chromosomes by Miller and co-workers (reviewed by Miller, Beatty &Hamkalo, 1972). These pictures are obtained by phosphotungstic acid staining of theprotein matrix investing the nucleic acids, and thus draw attention to the role playedby this protein.
In this communication we describe some observations on the properties of theproteins in newt germinal vesicles isolated by microdissection and fractionated by acombination of biochemical, biological and microdissection techniques.
A preliminary account of some of this work has been published elsewhere (Hill,Maundrell & Callan, 1973).
MATERIALS AND METHODS
MaterialsFemales of the great crested newt, Triturus cristatus carnifex (Laurenti), were purchased
from the dealers L. Haig and Son, Newdigate, Surrey. They were maintained in the laboratoryat 18 °C and used in the period October to July. Ovaries were removed as described by Callan& Lloyd (i960) and stored dry in embryo cups on ice for use later the same day.
Preparation of lampbrush chromosomes and nucleoli for microscopic observation of theeffect of dissociating agents
Lampbrush chromosomes and nucleoli from oocytes of 08 + CC05 mm diameter were pre-pared for microscopic observation along the lines of Callan & Lloyd (i960), with the followingmodifications.
The oocytes were opened, and their germinal vesicles cleaned and transferred in 0075 M KC1,0-025 M NaCl. Removal of the nuclear membrane was performed in preparation chambersmounted on disk-shaped microscope slides (ca. 1 in. (25-4 mm) diameter, cut from 3 x 1 in.bored slides) containing 0075 M KC1, 0025 M NaCl, 001 M Tris, o-ooi M MgCl2, 000005 M
CaCl2, pH 7-2. After 20-30 min at room temperature to allow dispersal of the nuclear contents,each upper circular coverslip was lowered flush with the surface of the microscope slide byblotting away excess fluid, and the chambers were placed on adaptors near the base of 50-mlSorvall polyethylene centrifuge tubes for 10 min centrifugation at 5000 g in a Sorvall HB-4

Proteins of newt oocyte nucleus 147
swinging bucket rotor. This centrifugation served to attach the lampbrush chromosomes andnucleoli to the floor of the chamber, thereby simplifying their optical observation and preventingtheir dislodgement during addition of reagents to the chamber.
Semibulk collection of nuclei and centrifugal fractionation
Nuclei from 0 8 ± 0-05 mm diameter oocytes were cleaned in 0075 M KC1, 0025 M NaCl,and collected either in i-ml glass centrifuge tubes or on dimple dishes and subsequently trans-ferred to tubes with a ioo-/tl ' Micropet' attached to a Hamilton syringe. Glass surfaces employedin transfer and storage of the cleaned nuclei were siliconized with Repelcote (Hopkins andWilliams).
For analysis of proteins from whole nuclei, 25 nuclei were collected at room temperature in50 fil of 4/ig per ml pancreatic ribonuclease, o-i M NaCl, 4 M urea to give a final volume of100 /i\. Dissociation of the cytological structures was then continued as described below in thesection headed Dissolution of cytological structures and stabilization of the dispersed proteins.
Nucleoplasm and chromatin were obtained by centrifugal fractionation of nuclei collected in0-075 M KC1, 0-025 M NaCl, 0-00005 M CaCl, at 1-2 °C. For the preparation of nucleoplasm 25nuclei were collected in a final volume of ca. 50 fi\. The material was briefly mixed by gentlevortexing and then centrifuged at 1500 g for 7 min at 3 °C. The supernatant nucleoplasm wasremoved under a dissecting microscope, using a polythene tube connected to a Hamilton syringe.Chromatin was separately prepared from 50 nuclei, collected in 150-200/41. Following gentlemixing and centrifugation, as much supernatant as possible was removed without disturbing thepellet. The chromatin was then washed by adding another 200 /tl of 0075 M KC1, 0025 M NaCland repeating the mixing and centrifugation. Withdrawal of the second supernatant left thechromatin pellet covered by 5-10 fi\ of saline.
Fractionation of the chromatin by biological and microdissection techniques
Chromosomes alone were obtained from germinal vesicles of 0-6—0-7 mm diameter oocytes.The nuclear membranes with attached nucleoli (vide infra) were dissected away in 0075 M KC1,0-025 M NaCl. Before dispersal of the nuclear contents had occurred they were transferred witha fine-tipped pipette to a o-5-ml conical tube filled with 0075 M KC1, 0-025 M NaCl, 000005 M
CaCl2 and maintained at 1-2 °C. (The CaCl2 is added to aid dispersal of the nucleoplasm and isknown to have no significant effect on the matrix of the chromosomes as judged by phase micro-scopy.) After the contents of 65 nuclei had been allowed to settle and disperse in the tube, someof the supernatant fluid was carefully withdrawn leaving the chromosomes covered by ca. 200 ju\.The solution was then mixed by stirring with a fine glass tube connected to a Hamilton syringeor by gentle vortexing. Following 7 min centrifugation at 1500 g, the supernatant was withdrawnand the pellet washed by resuspension in 200 fi\ of 0075 M KC1, 0-025 M NaCl. The final pelletwas left covered by 5-10 fi\ of saline ready for dissolution.
Membranes with attached nucleoli were collected from oocytes of diameter 0-6-0-7 mm.Washed germinal vesicles were allowed to settle individually in 0-075 M KC1, 0025 M NaClcontained in an embryo cup. They were then slashed open with a tungsten needle and theircontents pumped away with a fine-tipped pipette. The membranes and still attached nucleoliwere washed by pipette in saline in a second embryo cup; 65 of these membranes were collectedin a conical tube and washed twice more by suspension and centrifugation (7 min, 1500 g) in200 fi\ of saline. The final supernatant was removed leaving the membranes covered by 5-10 jAof saline and ready for dissolution.
Membranes without nucleoli were obtained by the same procedure as described above exceptfor the modification that they were dissected from 25 oocytes of diameter 1-5-1-7 mm; at thisstage most nucleoli have detached from the nuclear membrane. Twenty-five membranes fromoocytes at the later stage have slightly more surface area than 65 membranes at the earlier stage.
Dissolution of cytological structures and stabilization of the dispersed proteins
Total nuclei and the fractions described above were treated for 30 tc 60 min at room tempera-ture with 2 fig per ml of pancreatic ribonuclease in o-i M NaCl, 2 M urea. This step was intro-duced as a precaution to degrade RNA and minimize any RNA-protein interactions that might

148 R. J. Hill, K. Maundrell and H. G. Callan
occur during gel electrophoresis. Concentrated guanidine hydrochloride/sodium pyrophos-phate, pH 8 1 , was added to give final concentrations of 4 M and 001 M respectively. Thesolution was made 011 M in mercaptoethanol and allowed to stand overnight under nitrogen atroom temperature. The next morning Tns buffer, EDTA and iodoacetate were added to thefinal concentrations specified by Crestfield, Moore & Stein (1963) to alkylate protein sulphydrylgroups. After allowing 15 min for reaction, excess reagents were removed by dialysis against7 M urea, 0005 M Tris, pH 8-6. The 5-carboxymethylated proteins were finally dialysedagainst 7 M urea/Tris stacking buffer or 01 % sodium dodecyl sulphate, 001 M sodium phos-phate, pH 7-1 depending on the method of electrophoresis.
Gel electrophoresis
pH 9, 7 M urea gel electrophoresis. This was performed in 7 % gels by the procedure of Davis &Ornstein (Davis, 1964) with the following modifications. 7 M urea was included throughout thestacking and separating gels and 1 M urea in the upper electrode vessel buffer. Electrophoresiswas carried out on a semimicro scale in 02-cm internal diameter tubes: stacking gel length 1 cm,separating gel length 4-5 cm. Stacking was performed at 100 V (constant voltage) until all thesample had entered the stacking gel, and then at 200 V. Upon completion of stacking, separationwas carried out at 360 V.
Proteins were visualized by staining with Coomassie Brilliant Blue R.pH 7 1 , o-i % sodium dodecyl sulphate. The 5 % gel system was essentially that of Shapiro,
Vifiuela & Maizel (1967). Samples were prepared in 001 M sodium phosphate pH 7 1 , 0-005 %bromphenol blue, 5 % glycerol, 5 % mercaptoethanol, o- i% sodium dodecyl sulphate exceptin the case of bovine serum albumin dimer where mercaptoethanol was omitted. They were runon 5 x 0 2 cm gels at a constant current of 25 mA per gel. A plot of log molecular weight versusmobility was established with the following standard proteins: myoglobin, carbonic anhydrase,ovalbumin and phosphorylase a, all from Worthington, and bovine serum albumin from Pentex.
RESULTS
Conditions for the dissolution of structures within the germinal vesicle
The macromolecular interactions stabilizing chromosome structure must surelyinvolve hydrogen, hydrophobic and ionic bonding. Aqueous guanidine hydrochloride,thought to break all of these classes of non-covalent bonds (Tanford, 1964), might wellbe expected to be the dissociating agent of choice for chromosomes. In fact this reagenthas been used for the dispersal of interphase chromatin by several workers (see, e.g.Huang & Huang, 1969; Hill et al. 1971). The well defined chromosomal structures ofthe germinal vesicle provide a valuable opportunity for direct microscopic monitoringof the effect of this solvent on the genetic apparatus of an actively transcribing nucleus.
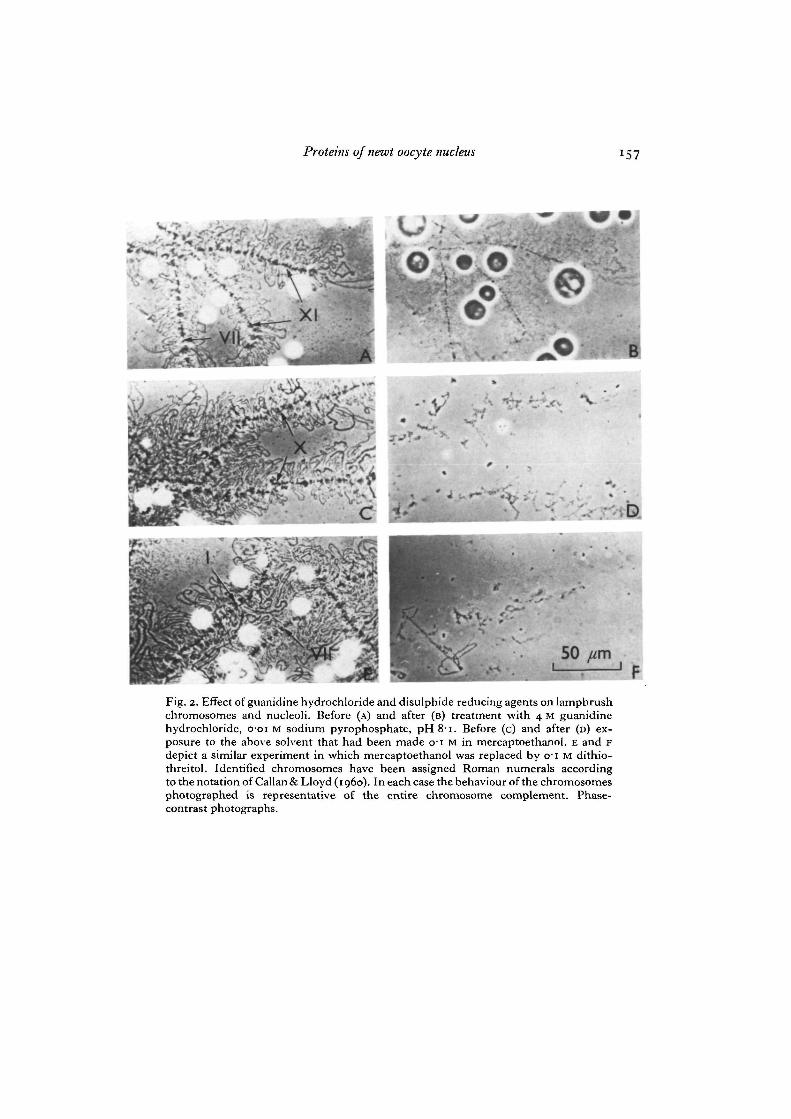
Removal of the nuclear membrane from the germinal vesicle by microdissectionreleases the lampbrush chromosomes, nucleoplasmic granules and a variable propor-tion of the nucleoli. Centrifugation attaches these structures to the floor of the chamberand presents lateral loops and chromomeres of the lampbrush chromosomes, nucleo-plasmic granules and nucleoli all in the one plane. Portions of typical preparations areshown under phase contrast in Figs. 2 A, C and E. When guanidine hydrochloride,buffered at pH 8-i, was added under the top coverslip to a final concentration of 4 M,the optical contrast of the preparation decreased considerably (Fig. 2 B), due in partto the increased refractive index of the solvent, and probably in part to dissolution ofmaterial. The evident swelling of the nucleoli may be another reflexion of partial dis-solution of protein. However, all cytological structures of the original preparation arestill apparent.

Proteins of newt oocyte nucleus 149
On repeating this experiment, but with the incorporation of o-1 M mercaptoethanolin the final aqueous guanidine solvent, a far more dramatic effect is observed (Fig.2 c, D). Dissolution now has gone almost to completion and the original cytologicalstructures have collapsed. The chromosome axes can be traced as just recognizablestrings of remnants, presumably held together by the DNA backbone. Chromomeresand loops are no longer distinguishable. Traces of cores are all that remain to be seenof the nucleoli. Replacing mercaptoethanol with dithiothreitol leads, if anything, tomore complete dissolution of the structures (Fig. 2E, F). Thioglycollate produced asimilar effect (not shown).
These observations establish mild conditions for dispersion of the macromolecularcomponents of the cytological structures within the germinal vesicle. Furthermore,mercaptoethanol, dithiothreitol and thioglycollate are well established as specificreagents for the reductive cleavage of disulphide bonds (White, i960; Cleland, 1964;Smithies, 1964). They might also act as chelating agents to sequester divalent cations.However, since the strong chelating agent ethylenediaminetetra-acetate at pH 8,acting in concert with guanidine hydrochloride, produced marginal, if any, increaseddissociation over guanidine hydrochloride alone, it seems unlikely that the primarybasis of the effect of the sulphydryl reagents could be the breakage of divalent-cationbridges. Our findings thus provide good circumstantial evidence that disulphide bondcrosslinks contribute to the morphological integrity of isolated lampbrush chromosomes(both chromomeres and lateral loops) and nucleoli.
The above observations were made within 2 h of removal of the nucleus from theoocyte. It is known that the germinal vesicle is rich in protein sulphydryl groups(Brachet, 1938, 1950). As sulphydryl groups, including those of proteins, are subject toatmospheric oxidation (Anfinsen & Haber, 1961) it is possible that the disulphide bondcrosslinks demonstrated are progressively formed during isolation of the germinalvesicle and subsequent manipulations. Indeed, such a reaction would explain ourobservations that isolated lampbrush chromosomes become more refractory to dis-solution as time passes, and a similar phenomenon exhibited by amphibian oocytenucleoli (O. L. Miller, personal communication of 1971, and Miller & Beatty, 1969).
Proteins of the germinal vesicle, its nucleoplasm and chromatin
Having established conditions for solubilization which overcome the time-dependentannealing of the structures within the germinal vesicle, it became possible to accumulatehand-isolated nuclei and to obtain a semi-bulk preparation of material for the analysisof proteins. Nuclei were individually isolated from o-8-mm diameter oocytes andrapidly cleaned by pumping with a fine-drawn glass pipette until microscopic observa-tion indicated that they were completely freed from cytoplasm and yolk granules.Following this operation, which was normally completed in 30 s or less, the isolatednuclei were bulked.
On standing in 0-075 M KG, 0-025 M NaCl for 30 min, the nuclei ruptured spon-taneously with the partial release of their chromosome complement and any nucleolinot attached to the nuclear envelope. This mixture could be fractionated by low-speedcentrifugation to give a nucleoplasm supernatant over a pellet of chromatin. Fig. 3

150 R. J. Hill, K. Maundrell and H. G. Callan
shows the microscopic appearance of the centrifugal product when these operationswere performed in a bored slide. The resulting chromatin consists of lampbrushchromosomes spilling out through the ruptured nuclear membrane. In this particularinstance effectively all of the nucleoli have remained attached to the nuclear membrane.The release of nucleoli from the membrane actually depends upon the stage of oogenesis{vide infra).
Nuclear proteins were prepared for gel electrophoretic analysis from massed totalnuclei, nucleoplasm and washed chromatin by solubilization under reducing conditionsand stabilization of sulphydryl groups by alkylation as described in Materials andMethods. Gel electropherograms at pH 9 in 7 M urea are shown in Fig. 4. Totalnuclear material is resolved into about 12 bands. Of these, one (1-30) is much depletedin the nucleoplasmic fraction. A second (2-65) appears to be present in slightly decreasedproportions in the nucleoplasm. Both of these components are much enriched in thechromatin fraction. The increase in the proportion of band 2-65 relative to band 1-30 inthe centrifugable material may result from a higher association constant for binding theformer. The presence of traces of the major nucleoplasmic species i-oo and 2-25 in thechromatin may be a reflexion of their distribution between the nucleoplasm and chro-matin, or may represent limitations of the washing procedure; the present data areequivocal on this point. However, if it is assumed that they are not bound, their presencegives an upper limit for entrainment of nucleoplasmic species through the washingprocedure. It would then appear that the minor component 1-75 does preferentiallybelong to the chromatin fraction.
Fractionation of the chromatin: an approach to the further localization of protein species
Shortly after the onset of vitellogenesis, in oocytes of 0-6-0-7 m m diameter in whichthe lampbrush lateral loops are very extended, the nucleolar population may generallybe found at the periphery of the nucleus and firmly bound to the nuclear membrane.Much later in oogenesis the inverse generally holds and animals may be obtained inwhich oocytes of 1-5-1-7 mm diameter display some 90% of their nucleoli detachedfrom the membrane and free in the nucleoplasm. Exploitation of this stage-dependentdifferential attachment of nucleoli to the membrane, and the fact that the membranemay be isolated from the free contents of the germinal vesicle by microdissection,allows fractionation of the components of the chromatin.
The total chromatin pellet obtained by low-speed centrifugation of ruptured wholenuclei consists of nuclear membranes, nucleoli and lampbrush chromosomes (Fig. 5 A).If, prior to collection of nuclear material from oocytes at the early stages of oogenesis,the nuclear membranes are removed, nucleoli are removed along with them so that theresultant washed chromatin pellet contains lampbrush chromosome material and littleelse (Fig. 5B). The membranes and attached nucleoli may also be massed and washedthoroughly to give material corresponding to that illustrated in Fig. 5 c. Finally,isolated membranes with very few attached nucleoli may be obtained from oocytesat the later stages of oogenesis (Fig. 5D).
The protein pattern given by whole nuclei (nucleoplasm plus chromatin) is depictedin Fig 6A. Total chromatin, displaying the two enriched components (1-30 and 2-65),

Proteins of newt oocyte nucleus 151
along with other minor components, is represented by Fig. 6B. Both of these enrichedspecies are again present in the pelleted membranes with attached nucleoli (Fig. 6 c).Approximate calculations, based on the assumption that the nuclear membrane maybe considered to consist of the equivalent of 2 unit membranes and annuli (Stevens &Andre, 1969), and the consideration that membrane proteins have generally been foundto be heterogeneous (Lenard, 1970; Fairbanks, Steck & Wallach, 1971), suggest thatmembranes, per se, are not contributing the 2 major protein species. This is demon-strated directly by Fig. 6D which records the electropherogram produced by isolatedmembranes. We must therefore conclude that the 2 major species are stemming, atleast in considerable part, from the nucleoli.
The final gel records the pattern given by the chromatin pellet, consisting of lamp-brush chromosomes alone. Only one very faint band in the position i-oo could bediscerned on the original gel. Since this corresponds to the mobility of a major speciesin the whole nuclei it remains an open question whether this is a genuine chromosomalcomponent or simply a reflexion of entrainment of nucleoplasm into the chromosomalpellet. The relatively small amount of protein in the chromosomal pellet may at firstsight seem surprising in the light of previous estimations of the composition oflampbrush chromosome preparations (Isawa et al. 1963; Edstrom & Gall, 1963).However in some of these estimations formaldehyde fixation was performed onmicroscope slides in the presence of nuclear sap before the area corresponding to thechromosomes was collected off the slide. In such operations performed in the presenceof a great excess of nucleoplasm, the elimination of contaminating nucleoplasmiccomponents from the chromosomes might prove difficult. On the other hand it shouldbe pointed out that 'shedding' of matrix material from the chromosomes may beoccurring during our washing and centrifugation procedures, though no significantdifferences in the whole chromatin patterns were apparent when resuspension ofmaterial was performed by procedures as diverse as vortexing and gentle mixing with afine glass Micropet capillary tube. The questions raised here obviously demandfurther study.
The clear conclusion emerging at present is that the 2 major polypeptides of oocytechromatin are components of the nucleoli. Order of magnitude calculations, based onthe fact that within the germinal vesicle there are some thousand nucleoli (Buongiorno-Nardelli, Amaldi & Lava-Sanchez, 1972) consisting very largely of protein and esti-mated by interference microscopy to have a dry mass of the order of 100 pg each(Macgregor & Moon, 1971), are consistent with these structures providing a majorsource of the 2 proteins.
Sodium dodecyl sulphate electrophoresis of nuclear proteins and molecular weight estima-tions
Sodium dodecyl sulphate is one of the most powerful agents available for themolecular dispersion of proteins; moreover, in its presence, both basic and acidicproteins adopt a high net negative charge. A total display of nuclear proteins is thusobtained by ionic electrophoresis in the presence of this detergent (Fig. 7). The wholenuclear pattern reveals some 25 components migrating in the molecular weight range

152 R. J. Hill, K. Maundrell and H. G. Callan
10-150 kilodaltons and above. Again the chromatin pattern differs markedly from thewhole nuclear pattern and once more demonstrates the presence of 2 distinct majorcomponents. However, these gels also reproducibly demonstrate a considerable number(ca. 20) of minor species present in the chromatin pellet. Some or all of these are likelyto be components of the chromosomal and nucleolar structures.
15
ox 6
* 4
"o 3
O Serum albumin dimer
O \ Phosphorylase a
Serum albumin
Ovalbumin
CarbonicI \ anhydrase
Myoglobin O
0-2 0-4 0 6 0-8Relative mobility
10
Fit;. 1. Plot of log molecular weight versus mobility for 6 standard proteins showingthe interpolation of apparent molecular weights for the 2 major chromatin species.
The molecular weight dependence of mobility for the semimicro-electrophoresissystem employed here was determined empirically, and is documented in Fig. 1. Theapparent molecular weights of the 2 major chromatin proteins, interpolated from thisrelationship, are n o and 43 kilodaltons. The approximate region where the 5 majorhistone classes would be expected to appear was determined by running rat liverhistones and is indicated by the bracket in Fig. 7. These proteins thus, at most, makeonly a minor contribution to the overall chromatin protein population. The 2 majornonhistones therefore predominate over all other chromatin proteins.
DISCUSSION
There are now numerous examples of polymerization of chromosomal proteins viadisulphide bonding. Zittle & O'Dell (1941), and Lung (1968) have adduced evidence

Proteins of newt oocyte nucleus 153
suggesting that disulphide bonding plays a role in the maintenance of the tightly con-densed state of chromatin in human and bull sperm. Gruzdev & Belaja (1968) havesuggested that disulphide bridges are involved in the maintenance of the structure ofpolytene chromosomes. The finding that the sulphydryl groups of histone III arereduced in interphase HeLa nuclei but oxidized in metaphase chromosomes ledSadgopal & Bonner (1970) to suggest that polymerization of proteins via disulphidebridges may contribute to the condensed state of metaphase chromosomes.
Our observations suggesting that disulphide-bond crosslinks contribute to themorphological stability of newt germinal vesicle lampbrush chromosomes and nucleoliare thus not without precedent in chromatin biochemistry. The possibility that thesecrosslinks arise, at least in part, by aerial oxidation subsequent to removal from thesulphydryl-rich germinal vesicle, and that they thereby provide the chemical basis ofthe well known hardening or 'denaturing' of these structures, deserves further study.In any event, recognition that disulphide bonds must be broken in order to obtainmolecular dispersion of the components of lampbrush chromosomes and nucleoliremoves a barrier to further biochemical studies.
It has been suggested that the reality of nonhistone chromosomal proteins should bequestioned because of the binding of cytoplasmic proteins to DNA during isolation(Johns & Forrester, 1969) and because of an apparent artifactual conversion of histonesto nonhistones by acid treatment (Sonnenbichler & Nobis, 1970). The mobility of the12 or so whole germinal vesicle proteins towards the anode in urea at pH 9 eliminatesthem from the histone class (see, e.g. Tidwell, Allfrey & Mirsky, 1968). In the particularcase of the 2 major chromatin proteins this conclusion is reinforced by the sodiumdodecyl sulphate electropherograms. The rapid nuclear isolation by microdissection, inwhich the germinal vesicle is separated from the bulk of the cytoplasm in a few secondsand completely cleaned in thirty seconds or less, leaves little doubt that its proteincontents are indeed nuclear in origin. Macgregor (1962) has shown, by interferencemicroscopy, that the dry mass of the germinal vesicle remains constant in 0-25 M KC1,0-05 M NaCl during the interval from one to three minutes following isolation; thisincreases our confidence that protein exchanges do not occur across the nuclearmembrane during isolation from the cytoplasm. Finally, the nonhistones of the germinalvesicles have not been exposed to pH below 6. These oocyte nonhistone proteins musttherefore be accepted as well established entities.
The suggestion has been made that, at least in some systems, nonhistone proteinsmay originate predominantly from the nuclear membrane (Harlow, Tolstoshev &Wells, 1972). In the case of oocyte nonhistone nuclear proteins our direct observationson manually isolated nuclear membranes demonstrate unequivocally that this is notthe case. By far the bulk of the acidic protein within the germinal vesicle is present inthe nucleoplasm. The nucleoplasm gives a very similar electropherogram to wholenuclei, but with some depletion of 2 protein species. These species have been shown tobe major components of the chromatin.
The oocyte chromatin nonhistone proteins have been resolved, in the presence ofsodium dodecyl sulphate, into approximately 20 species. These are all good candidatesas components of the nucleoli and chromosomes. Perhaps the most outstanding feature

154 R. J. Hill, K. Maundrell and H. G. Callan
of this nonhistone population is its distribution into 2 distinct major species and manyminor species. We have presented evidence indicating that the 2 major componentsoriginate, at least in large part, from the nucleoli. It may be of interest that Vincent(i960, 1964) presented data, albeit at a lower level of resolution, indicating that thereis a single predominant protein species in starfish oocyte nucleoli. An unusual featureof oocyte chromatin (both nucleoli and lampbrush chromosomes) is the high contentof RNA relative to DNA (Macgregor, 1972; Edstrom & Gall, 1963). This appears to bea reflexion of the high rate of transcription (Miller et al. 1972). Thus much of theprotein of germinal vesicle chromatin is likely to be associated with nascent RNA. Atthis point it may be germane to recall the very simple pattern given by fully dissociated'informatin' protein in the hands of Krichevskaya & Georgiev (1969). This protein,which is believed to associate with nascent nuclear RNA and be involved in its process-ing, gives a single band on electrophoresis. It is suggestive that the molecular weightof one of the two major oocyte chromatin proteins should be in the same range as that ofinformatin, viz. 40-45000. It may be relevant that informatin has been shown to poly-merize via disulphide bond formation.
The striking photographs of oocyte nucleolar genes in flagrante transcripto byMiller and co-workers (see e.g. Miller et al. 1972) are actually, as pointed out by theseinvestigators, visualized via phosphotungstic acid staining of the protein matrixcoating the RNA and DNA. It is an intriguing possibility that the nucleolar proteinswe are detecting may well be components of this matrix.
Finally, there remains the question of the protein components of the lampbrushchromosomes themselves. Direct isolation of manually dissected chromosomes has,to date, provided insufficient material to give a distinctive electropherogram. The mostobvious conclusion from our present data would be that these structures contain lessprotein than previous estimates would suggest, and/or that the chromosomal proteinis distributed between many species; this may well prove to be the case. (In fact,evidence from an independent approach, consistent with the latter possibility, hasrecently been presented by Sommerville & Hill, 1973.) However, the chromosomesthemselves are exceedingly fragile structures which show a marked predilection forshedding components on the application of mild shear forces. The semibulk isolationof lampbrush chromosomes by a combination of microdissection and biochemicalprocedures remains an important challenge for the future.
We are indebted to Mrs L. Lloyd for skilled technical assistance, and to Mr David Roche forhis contribution to the photography. This project has been supported by a Science ResearchCouncil Grant.
REFERENCESANFINSEN, C. B. & HABER, E. (1961). Studies on the reduction and reformation of protein
disulphide bonds. J. biol. Client. 236, 1361-1363.BRACHET, J. (1938). La localisation des proteines sulphydritees pendant le developpement des
amphibiens. Bull. Acad. r. Belg. Cl. Sci. 24, 499-510.BRACHET, J. (1950). In Chemical Embryology, pp. 55-56. New York and London: Interscience
Publishers.

Proteins of newt oocyte nucleus 155
BRIGGS, R. & JUSTUS, J. T . (1968). Partial characterisation of the component from normaleggs which corrects the maternal effect of gene O in the Mexican Axolotl {Ambystoma mexi-canum).J. exp.Zool. 167, 105-116.
BUONGIORNO-NARDELLI, M., AMALDI, F. & LAVA-SANCHEZ, P. (1972). Amplification as arectification mechanism for the redundant rRNA genes. Nature, New Biol. 238, 134-137.
CALLAN, H. G. & LLOYD, L. (i960). Lampbrush chromosomes of crested newts Trituruscristatus (Laurenti). Phil. Trans. R. Soc. Ser. B 243, 135-219.
CLELAND, W. W. (1964). Dithiothreitol, a new protective reagent for SH groups. Biochemistry,N. Y. 3, 480-482.
CRESTFIELD, A. M., MOORE, S. & STEIN, W. H. (1963). The preparation and enzymatic hydrolysisof reduced and S-carboxymethylated proteins..?. biol. Chem. 238, 622-627.
DAVIS, B. J. (1964). Disc electrophoresis II . Method and application to human serum proteins.Ann. N. Y. Acad. Sci. 121, 404-427.
EDSTROM, J. E. & GALL, J. G. (1963). The base composition of ribonucleic acid in lampbrushchromosomes, nucleoli, nuclear sap, and cytoplasm of Triturus oocytes. J. Cell Biol. 19,279-284.
ELGIN, S. C. R. & BONNER, J. (1972). Partial fractionation and chemical characterisation of themajor nonhistone chromosomal proteins. Biochemistry, N.Y. n , 772-781.
FAIRBANKS, G., STECK, T. L. & WALLACH, D. F. H. (1971). Electrophoretic analysis of themajor polypeptides of the human erythrocyte membrane. Biochemistry, N.Y. 10, 2606-2617.
GALL, J. G. (1954). Lampbrush chromosomes from oocyte nuclei of the newt. J. Morph. 94,283-351-
GEORGIEV, G. P. & SAMARINA, O. P. (1972). D-RNA containing ribonucleoprotein particles.In Advances in Cell Biology, vol. 2 (ed. D. M. Prescott, L. Goldstein & E. McConkey),pp. 47-110. Amsterdam: North Holland Publishing.
GRUZDEV, A. D. & BELAJA, A. N. (1968). On the linkage in polytene chromosomes. Tsitologiya10, 955-1001.
HARLOW, R.,TOLSTOSHEV, P. & WELLS, J. R. E. (1972). Membrane-a major source of chromatin-associated RNA and nonhistone proteins in avian erythroid cells. Cell Differentiation 2,341-349-
HILL, R. J., MAUNDRELL, K. G. & CALLAN, H. G. (1973). Proteins of the newt germinal vesiclenucleus. Nature, Neiv Biol. 242, 20-22.
HILL, R. J., POCCIA, D. L. & DOTY, P. (1971). Towards a total macromolecular analysis of seaurchin embryo chromatin. J. molec. Biol. 61, 445-462.
HUANG, R. C. & HUANG, P. C. (1969). Effect of protein-bound RNA associated with chickembryo chromatin on template specificity of the chromatin. J. molec. Biol. 39, 365-378.
ISAWA, M., ALLFREY, V. G. & MIRSKY, A. E. (1963). Composition of the nucleus and chromo-somes in the lampbrush stage of the newt oocyte. Proc. natn. Acad. Sci. U.S.A. 50, 811-817.
JOHNS, E. W. & FORRESTER, S. (1969). Studies on nuclear proteins. The binding of extra acidicproteins to deoxyribonuclear-protein during the preparation of nuclear proteins. Eur. J.Biochem. 8, 547-551.
KRICHEVSKAYA, A. A. & GEORGIEV, G. P. (1969). Further studies on the protein moiety innuclear DNA-like RNA containing complexes. Biochim. biophys. Ada 164, 619-621.
LENARD, J. (1970). Protein and glycolipid components of human erythrocyte membranes.Biochemistry, N.Y. 9, 1129-1132.
LUNG, B. (1968). Whole-mount electron microscopy of chromatin and membranes in bull andhuman sperm heads. J'. Ultrastruct. Res. 22, 485-493.
MACGILLIVRAY, A. J., CARROLL, D. & PAUL, J. (1971). The heterogeneity of the nonhistonechromatin proteins from mouse tissues. FEBS Letters, Amsterdam 13, 204-208.
MACGREGOR, H. C. (1962). The behaviour of isolated nuclei. Expl Cell Res. 26, 520-525.MACGREGOR, H. C. (1972). The nucleolus and its genes in amphibian oogenesis. Biol. Rev. 47,
177-210.MACGREGOR, H. C. & CALLAN, H. G. (1962). The actions of enzymes on lampbrush chromo-
somes. Q.Jl microsc. Sci. 103, 173-203.MACGREGOR, H. C. & MOON, S. J. (1971). Some measurements on amphibian oocyte nucleoli.
Z. Zellforsch. mikrosk. Anat. 122, 273-282.MALACINSKI, G. M. (1972). Identification of a presumptive morphogenetic determinant from
the amphibian oocyte germinal vesicle nucleus. Cell Differentiation 1, 253-264.

156 R. J. Hill, K. Maundrell and H. G. Callan
MILLER, O. L. & BEATTY, B. R. (1969). Visualisation of nucleolar genes. Science, N.Y. 164,955-957-
MILLER, O. L., BEATTY, B. R. & HAMKALO, B. A. (1972). Nuclear structure and function duringamphibian oogenesis. In Oogenesis (ed. J. D . Biggers & A. E. Schuetz), pp. 119-128. Balti-more : University Park Press.
PAUL, J. & GILMOUR, R. S. (1968). Organ-specific restriction of transcription in mammalianchromatin.X molec. Biol. 34, 305-316.
SADGOPAL, A. & BONNER, J. (1970). Proteins of interphase and metaphase chromosomes com-pared. Biochim. biophys. Ada 207, 227-239.
SHAPIRO, A. L., VINUELA, E. &MAIZEL, J. V. (1967). Molecular weight estimation of polypeptidechains by electrophoresis in SDS-polyacrylamide gels. Biochem. biophys. Res. Commun. 28,815-820.
SMITHIES, O. (1964). Starch-gel electrophoresis. Metabolism 13, 974-984.SOMMERVILLE, J. & HILL, R. J. (1973). Proteins associated with heterogeneous nuclear RNA
of newt oocytes. Nature (in Press).SONNENBICHLER, J. & NOBIS, P. (1970). The so-called nonhistones from acid-treated calf thymus
chromatin. Eur.J. Biochem. 16, 60-65.STEVENS, B. J. & ANDRE, J. (1969). The nuclear envelope. In Handbook of Molecular Cytology
(ed. A. Lima-de-Faria), pp. 837-871. Amsterdam: North-Holland Publishing.TANFORD, C. (1964). Cohesive forces and disruptive reagents. In Sub-Unit Structure of Pro-
teins, Brookhaven National Laboratory Symp. pp. 154-173.TENG, C. S., TENG, C. T. & ALLFREY, V. G. (1971). Studies on nuclear acidic proteins. J. biol.
Chem. 246, 3597-3609.TIDWELL, T., ALLFREY, V. G. & MIRSKY, A. E. (1968). The methylation of histones during
regeneration of the liver. J. biol. Client. 243, 707-715.VINCENT, W. S. (i960). A biochemical approach to cell morphology. In New Approaches in Cell
Biology (ed. P. M. B. Walker), pp. 89-94. New York and London: Academic Press.VINCENT, W. S. (1964). The nucleolus. In Genetics Today, Proc. Xlth int. Congr. Genetics,
vol. 2 (ed. S. J. Geerts), pp. 343-358. London: Pergamon.WHITE, F. H. (i960). Regeneration of enzymatic activity by air-oxidation of reduced ribo-
nuclease with observations on thiolation during reduction with thioglycollate. J. biol. Chem.235, 383-389-
ZITTLE, C. A. & O'DELL, R. A. (1941). Chemical studies of bull spermatozoa. J. biol. Chem.140, 899-907.
{Received 1 October 1973)
Proteins of newt oocyte nucleus
Fig. 2. Effect of guanidine hydrochloride and disulphide reducing agents on lampbrushchromosomes and nucleoli. Before (A) and after (B) treatment with 4 M guanidinehydrochloride, o-oi M sodium pyrophosphate, pH 81. Before (c) and after (D) ex-posure to the above solvent that had been made 01 M in mercaptoethanol. E and Fdepict a similar experiment in which mercaptoethanol was replaced by 01 M dithio-threitol. Identified chromosomes have been assigned Roman numerals accordingto the notation of Callan & Lloyd (1960). In each case the behaviour of the chromosomesphotographed is representative of the entire chromosome complement. Phase-contrast photographs.

158 RJ. Hill, K. Maundrell and H. G. Callan
Fig. 3. Oocyte 'chromatin' prepared from a manually isolated germinal vesicle whichwas allowed to rupture spontaneously in a solution of 0075 M KC1, 0-025 M NaCl.A mass of lampbrush chromosomes can be seen to have emerged from the rupturednucleus lying to the left of the photograph. The bright objects associated with thenuclear membrane are nucleoli.Fig. 4. pH 9, 7M urea gel electropherograms given by solubilized, reduced and S-car-boxymethylated proteins, A, nucleoplasmic proteins; B, whole nuclear proteins; c,chromatin proteins; all from the same newt. D, a higher concentration of chromatinproteins from another newt.

Proteins of newt oocyte nucleus
i
100
» I
I1-30^ M-75 •65

i6o RJ. Hill, K. Maundrell and H. G. Callan
Fig. 5. Chromatin and its isolated subfractions. A, total chromatin, oocyte diameter08 mm; B, bulk lampbrush chromosomes, oocyte diameter 0-6-0-7 mm; c, nuclearmembrane with attached nucleoli, oocyte diameter 0-6-0-7 mm; D, membrane isolatedfrom oocyte of diameter i-6 mm showing very few attached nucleoli. The bars corre-spond to 50 /tm.

Proteins of newt oocyte nucleus
6 A J
CT
l J
7A
B
Fig. 6. pH 9, 7M urea gel electropherograms of S-carboxymethylated proteins preparedfrom: A, whole nuclei; B, total chromatin; c, membranes with attached nucleoli; D,membranes alone; and E, isolated chromosomes.Fig. 7. Sodium dodecyl sulphate gel electropherograms of S-carboxymethylatedproteins from: A, whole germinal vesicles; and B, total chromatin. The bracket delineatesthe region where the 5 common histones would be expected to run.II CEt I j
