reductase mrna levels in maize suspension - plant physiology
TRANSCRIPT

Plant Physiol. (1989) 90, 962-9670032-0889/89/90/0962/06/$01 .00/0
Received for publication October 25, 1988and in revised form March 1, 1989
Nitrate Effects on Nitrate Reductase Activity and NitriteReductase mRNA Levels in Maize Suspension Cultures
Laura S. Privalle*, Kristi N. Lahners, Martha A. Mullins, and Steven Rothstein1
CIBA-GEIGY Biotechnology Research, P.O. Box 12257, Research Triangle Park, North Carolina 27709
ABSTRACT
Nitrate reductase (NR) activity and nitrite reductase (NiR)mRNA levels were monitored in Black Mexican Sweet maize (Zeamays L.) suspension cultures after the addition of nitrate. Maximalinduction occurred with 20 millimolar nitrate and within 2 hours.Both NR and NiR mRNA were transiently induced with levelsdecreasing after the 2 hours despite the continued presence ofnitrate in the medium. Neither ammonia nor chlorate preventedthe induction of NR. Furthermore, removal of nitrate, followed byits readdition 22 to 48 hours later, did not result in reinduction ofactivity or message. NR was synthesized de novo, since cyclo-heximide completely blocked its induction. Cycloheximide had noeffect on the induction of NiR mRNA or on the transient nature ofits induction. These results are similar to those reported previ-ously for maize seedlings.
Nitrate is the major form of nitrogen most available forassimilation by higher plants growing under normal fieldconditions (9). A putative nitrate transport protein facilitatingthe uptake of nitrate is the initial step allowing plants toassimilate nitrate. The assimilatory pathway consists of NR,2a cytoplasmic enzyme, which converts the nitrate to nitrite;NiR, localized in the plastid, which then reduces the nitriteto ammonia; and glutamine synthetase, which catalyzes theconversion of glutamate to glutamine by incorporating freeammonia (6). The assimilation of nitrate is energeticallyexpensive, and the expression of the proteins in the pathwayis highly regulated. This regulation involves a complex set ofphysiological signals with light, environmental stresses, andplant growth regulators all affecting expression ofthe pathway(7, 13, 17, 24, 25). However, the primary environmental signalregulating expression is the presence of nitrate (6).The regulation of the nitrogen assimilatory genes has been
studied to the greatest extent in fungi, where mutants in boththe structural and regulatory genes have been isolated (2). Inhigher plants, nitrate induction of nitrate uptake and of thesynthesis of both NR and NiR have been studied extensively(3, 6, 12, 13, 24). The great variability in growth conditionsand protocols used makes valid comparisons between exper-iments very difficult. Furthermore, complications resultingfrom transport, storage, and compartmentation of the nitrate
' Present address: Department of Molecular Biology and Genetics,University of Guelph, Ontario, N1G 2W1 Canada.
2 Abbreviations: NR, nitrate reductase; NiR, nitrite reductase;BMS, Black Mexican Sweet (maize).
can make the analysis of its assimilation in whole plants hardto interpret.An alternative system for studying the regulation of nitro-
gen assimilation is to utilize cell suspension cultures or algalsystems. A single nondifferentiated cell type can be analyzedwithout the complexities inherent in a whole plant system.The physiological relevance of such studies to the situation inwhole plants remains to be determined. Zielke and Filner (27)monitored the synthesis and turnover of NR in tobacco celllines following transfer to nitrate-containing medium. Bymonitoring the change in buoyant density of '5N-labeledproteins and incorporation of tritiated amino acids into newlysynthesized proteins, they showed that nitrate induced denovo synthesis and turnover of NR. Nelson et al. (22), usingsoybean suspension cultures, examined the expression ofNRactivity as a function of time after subculturing and foundNR activity only when nitrate was present in the medium.Maki et al. (18) monitored NR activity and protein that cross-reacted with anti-NR antibody as a function ofage in culturedspinach cells. They concluded that de novo synthesis of NR,and not activation/inactivation, accounted for the changes inthe enzyme levels. In morning glory suspension cells, Zink(28) found that, after subculturing into nitrate-containingmedium, three separate peaks of NR enzyme activity ap-peared as the cultures aged.
Algal systems such as Chlorella differ from the higher plantsand cell cultures in that: (a) activation of an inactive NRprecursor has been reported (5); (b) ammonia represses NRinduction (5) whereas it enhances induction in rose suspen-sion cultures (19); and (c) nitrogen limitation (in the absenceof nitrate) results in NR induction (5). A single higher plantprecedent for this last point has been reported in tobaccosuspension cells, where nitrogen limitation was reported toresult in the induction ofNR activity (1 1).
In the work presented here, the regulation by nitrate ofNRenzyme activity and NiR mRNA has been analyzed in BlackMexican Sweet maize (Zea mays L., BMS) suspension cul-tures. The two showed a similar regulatory pattern. We foundthat they were both transiently induced by the addition ofnitrate, whereas neither chlorate nor ammonia preventedinduction. When nitrate was added to the growth mediumand then removed after 2 h, readdition of nitrate 24 h laterfailed to induce either NR activity or NiR mRNA synthesis.NR was synthesized de novo since cycloheximide completelyblocked its induction. However, cycloheximide had no effecton the induction of NiR mRNA. These results are generallysimilar to those seen for NiR protein and mRNA levels inmaize seedlings (14). Accordingly, Black Mexican Sweet sus-
962
Dow
nloaded from https://academ
ic.oup.com/plphys/article/90/3/962/6084322 by guest on 19 January 2022

NITRATE EFFECTS IN MAIZE SUSPENSION CULTURES
pension cultures appear to be a reasonable model system tostudy aspects of the regulation of the genes involved in nitrateassimilation using complicated manipulations that are diffi-cult or impossible with maize seedlings.
MATERIALS AND METHODS
Tissue Source
Black Mexican Sweet maize (Zea mays L.) suspensioncultures were grown on modified Murashige and Skoog me-dium (20): 1.2 g/L succinic acid, 1.08 g/L NH4Cl, 30 g/Lsucrose, 2 mg/L 2,4-D, 0.44 g/L CaCl2 .2 H20, 0.37 g/LMgSO4-7 H20, 0.17 g/L KH2PO4, 0.95 g/L KCl, 6.2 mg/LH3BO3, 15.1 mg/L MnSO4, 10.6 mg/L ZnS04-7 H20, 0.83mg/L KI, 0.25 mg/L Na2MoO4 2 H20, 0.025 mg/L CuSO4a5 H20, 0.025 mg/L CoCl2 .6 H20, 13.9 mg/L FeSO4.7 H20,18.65 mg/L K2EDTA, 0.2 mg/L glycine, 0.1 mg/L thiamine.HCI, 0.05 mg/L pyridoxine- HCI, 0.05 mg/L nicotinic acid.The sole nitrogen source in this medium is ammonia (20mM). The medium was adjusted to pH 5.8 with NH40H.Cultures were grown in 250-ml DeLong flasks with continu-ous shaking (135 rpm) at 26°C. Cells were subcultured weeklyby a 1: 10 dilution into fresh medium and were harvested forexperimental purposes when in logarithmic phase.
Induction Procedure
BMS cultures, approximately 1 week after subculturing,were treated with 0 to 50 mM KNO3, with 20 mM being thestandard concentration used. The cells were induced in theactual medium in which they had been grown. Preliminaryexperiments indicated no differences were found if the cellswere pelleted, washed, repelleted, and then resuspended intofresh medium with or without ammonia prior to the additionof nitrate. The age ofthe culture was found to be an importantparameter in obtaining maximal induction. Cultures less than6 days old or greater than 11 days old were less responsive tonitrate, with the density of the cells also having an effect.Therefore, care was taken to assure that the cultures weregrowing logarithmically and contained approximately 0.2 mLof packed cell volume/mL when utilized. After the period ofinduction, 10-mL aliquots were removed in duplicate one forNR enzyme assays and the other for Northern analysis ofNiR mRNA. In the reinduction experiment, the nitrate wasremoved at 2 h by pelleting the cells, washing two times infresh medium, and then resuspending in fresh medium. Ni-trate was not added back until 22 h later, i.e. 24 h after theinitial addition.
NR Extraction and Assay
Extraction of nitrate reductase was done using aliquots (10mL) of cells that had been treated as indicated for eachexperiment. The cells were pelleted at 500g, washed withextraction buffer (100 mM KPi, pH 7.5, 1 mM EDTA, 1 mMdithiothreitol), repelleted, and quick-frozen in dry ice/ethanol. Samples were stored at -2O°C until extracted. Ex-traction for NR activity was by physically grinding the frozentissue using Kontes Eppendorf pestles followed by centrifu-
gation for 10 min at 14,000g in a microcentrifuge. Aliquotsof the supernatant fraction were then assayed for NR by theprocedure of Nakagawa et al. (21), in which each 1-mLreaction mix contained (final concentration) 5 mM KNO3,100 mm KPi, 100 lM NADH. Assays were initiated by addi-tion of NADH and were carried out for 30 min at 30°C.Assays were terminated by addition of 0.5 mM K3Fe(CN)6.After 5 min, nitrite concentration was determined colorimet-rically by comparison to a nitrite standard curve followingthe addition of 1 ml each of 1% sulfanilamide in 3 M HCIand 0.2% N-( l-naphthyl)-ethylenediamine hydrochloride.
Protein was determined using the Bio-Rad Coomassie blueprocedure of Bradford (4).
mRNA Extraction and Northern Analysis
Aliquots (10-20 mL) ofcells were removed, pelleted, quick-frozen in dry ice/ethanol, and stored at -20°C. The tissue (1g) was suspended in 3 mL of extraction buffer (50 mM Tris-HCI, pH 8.0, 4% sodium para-aminosalicylate, 1% sodium1,5-napthalenedisulfonate) and 3 mL of buffer-saturatedphenol. The mixture was homogenized with a BrinkmannPolytron for 1 min and then shaken for 20 min at 300 rpm.After the addition of 2 mL of chloroform, the mixture wasshaken for an additional 10 min prior to centrifugation at7000 rpm in an SS34 rotor (Sorvall). The aqueous phase wasreextracted with 2 mL ofchloroform and made 2 M with LiCl.After overnight precipitation at 4°C, the RNA was sedimentedin a SW 50.1 rotor (Beckman) at 25 K for 2 h. The RNA wasresuspended in 1% SDS, 5 mM EDTA, 20 mM NaOAc, 40mM Tris-HCl, pH 7.5, and precipitated with EtOH. Theresulting pellet was resuspended in distilled H20, reprecipi-tated with ethanol, finally redissolved in distilled H20, andstored at -80°C.For RNA blot hybridization, 10 ,g of the LiCl precipitate
(from above) was subjected to electrophoresis through a 1.2%agarose, 2.2 M formaldehyde gel, and the RNA was blottedonto nitrocellulose. The probe used for the hybridization wasan NiR cDNA insert from the plasmid pCIB801 (15). Equalamounts of RNA were loaded on each lane as confirmed byreprobing the blot with a portion of a maize alcohol dehydro-genase cDNA clone (a gift ofJeffBennetzen, data not shown).Alcohol dehydrogenase is expressed constitutively in BMScells (26). Densitometric scanning (Zeineh soft laser scanningdensitometer model SL-504-XL) of autoradiographs fromseveral different exposures of the same blot allowed quantifi-cation of the levels ofmRNA induced by nitrate.
Cycloheximide and NADH were obtained from Sigma.
RESULTS
Induction of NR by Nitrate
The nitrate concentration dependence of the induction ofnitrate reductase in BMS is shown in Figure 1. A logarithmi-cally growing BMS culture, maintained on 20 mM ammonia,was divided into aliquots to which varying amounts of 0.5 MKNO3 were added to give the indicated concentrations. After2 h, the cells were pelleted, washed, and quick-frozen. Theywere then extracted and assayed for NR activity as described
963
Dow
nloaded from https://academ
ic.oup.com/plphys/article/90/3/962/6084322 by guest on 19 January 2022

Plant Physiol. Vol. 90,1989
0a-0.
E
L:
a-
z1-
0 0 20 30 40 50
Nitrate Concentration (mM)
Figure 1. Nitrate concentration-dependent induction of NR. A loga-rithmically growing BMS culture was divided into aliquots to which0.5 M KNO3 was added to the indicated final concentration, withoutchlorate (0), or in the presence of 25 mm chlorate (A). Cells werepelleted after 2 h, frozen, extracted, and assayed for NR as describedin Materials and Methods.
in Materials and Methods. Maximal induction was achievedby 20 mM KNO3 (Fig. 1). This experiment was carried out inthe medium in which the cells had been growing, since itsreplacement with fresh medium with or without 20 mM
ammonia had no effect on the nitrate-mediated induction ofNR.
Chlorate, at low concentrations, appears to be transportedby the same carrier protein as nitrate and can be reduced bynitrate reductase (10). In BMS suspension cultures, chloratedid not induce NR activity (data not shown). It did, however,decrease the effectiveness of nitrate induction of NR at lowlevels of nitrate (Fig. 1).The time course of NR induction by 20 mm nitrate was
monitored in logarithmically growing BMS cultures. Aliquots(10 mL) were removed at intervals and analyzed for NRactivity. The results shown in Figure 2 indicate a 200-foldinduction of NR peaking at 2 h after addition of nitrate,followed by a sharp decline. The activity returned to itsoriginal level within 24 h, whereas nitrate in the mediumremained essentially unchanged (as measured by a nitrateelectrode). The decrease in NR activity after 2 h indicates thatadditional modes of regulation other than induction in thepresence of nitrate are important. In cells from the same
cultures using standard procedures (16), we were not able tomeasure NiR activity. The presence or absence of ammoniahad no effect on the transient nature ofNR induction.To test the transient nature of the NR induction, nitrate
was removed at the peak of activity as described in Materialsand Methods. As before, this experiment involved the addi-tion of nitrate at time zero to a logarithmically growing BMSculture. The nitrate was removed at 2 h, and the cells werewashed and resuspended in fresh medium. Aliquots were
removed at various times, and NR activity was determined.Nitrate removal at 2 h did not prevent the loss ofNR activity(Figs. 2A and 3A). Furthermore, when 20 mm nitrate is addedat 24 h, there is little reinduction ofNR activity (Figure 3A).These results have two possible explanations: (a) whatever is
A
t
0 0.5 & -
Figure 2. Time course of induction of NR activity and NiR mRNA by20 mm nitrate. A BMS culture was brought to 20 mm nitrate by theaddition of 0.5 M KNO3 at time zero. Aliquots were removed andextracted for NR assay and for NiR mRNA Northern blot analysis atthe indicated times. Panel A, NR specific activity (0); relative intensityof NiR mRNA as measured densitometrically using the autoradi-ograph shown in panel B (+).
limiting the NR activity is not replenished 24 h after the initialnitrate induction, or (b) whatever is inhibiting NR inductionis not destroyed within 24 h after the initial nitrate induction.
Protein Synthesis Inhibitor Effects
Protein synthesis inhibitors were utilized to examine furtherthe transient nature of the induction of NR activity. Cyclo-heximide, at 10 jig/mL, added simultaneously with 20 mMnitrate, essentially blocked the induction ofNR (Fig. 4). Whencycloheximide (5 ,ug/mL) was added 1 h after the addition ofnitrate, NR activity began to decline immediately and neverreached the 2-h peak value of the minus cycloheximide con-trol (Fig. 5). These results indicate that the increase in activitywas due to de novo synthesis rather than activation of aninactive precursor.
Induction of NiR mRNA by Nitrate
We lacked an effective probe for the NR message but hadavailable a maize NiR cDNA clone (15). Furthermore, similarexperiments had been done using maize seedlings monitoringNiR mRNA, and transient induction was observed (14).Therefore, to monitor the effects of nitrate at the transcrip-tional level in BMS cell suspensions, the kinetics of inductionof the message for NiR was monitored concurrently with NRactivity even though these two may be unrelated. During thestudy ofthe time course of induction ofNR activity (Fig. 2A),aliquots were removed for NiR mRNA analysis by Northernblots, using a maize NiR cDNA clone as the probe (15). The
964 PRIVALLE ET AL.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/90/3/962/6084322 by guest on 19 January 2022
NITRATE EFFECTS IN MAIZE SUSPENSION CULTURES
-2
E
E ,.-
a:1
16
A1N3
t~NO3-
Timne (hr)
+ ,N0-+2524 26hr
Figure 3. Reinduction of NR activity and NiR mRNA by nitrate. ABMS culture was brought to 20 mm KNO3 by the addition of 0.5 MKNO3 at time zero. Nitrate was removed after 2 h and replaced at 24h. Aliquots of cells were removed, extracted and assayed for NRactivity (panel A) or analyzed by Northern blots (panel B) for NiRmRNA at the times indicated.
200
E
0)E
10
2 3 4 5
TIME (hr)
Figure 4. Effects of protein synthesis inhibitors on NR induction bynitrate. Nitrate (20 mm) and cycloheximide (10 ug/ml, A), or nitrate(20 mm) only (0) were added at time zero. Aliquots of cells wereremoved, extracted, and assayed for NR activity at the indicatedtimes.
pattern of induction ofNiR mRNA is quite similar to the NRactivity induction profile. The message peaks in intensity at2 h (Fig. 2B). Densitometric scanning of the film indicatedan approximate 100-fold increase in NiR message level (Fig.2A). The message decreased after this 2-h time point andreturned to the level seen at zero time by 24 h.
Samples from the reinduction experiment (Fig. 3A) werealso analyzed for NiR rnRNA induction. As was seen for theNR activity, there was no reinduction of the NiR mRNA
123
- 100
0)E80)E 80
) 6C0
C
4
>
< 2C
z
5
TIME (hr)
Figure 5. Effect of cycloheximide on the transient nature of NRinduction. Nitrate (20 mM) was added at time zero. After 1 h, 5 ig/mL cycloheximide was added to the indicated culture (A). No cyclo-heximide was added to the control culture (0). Aliquots of cells wereremoved, extracted, and assayed for NR activity at the indicatedtimes.
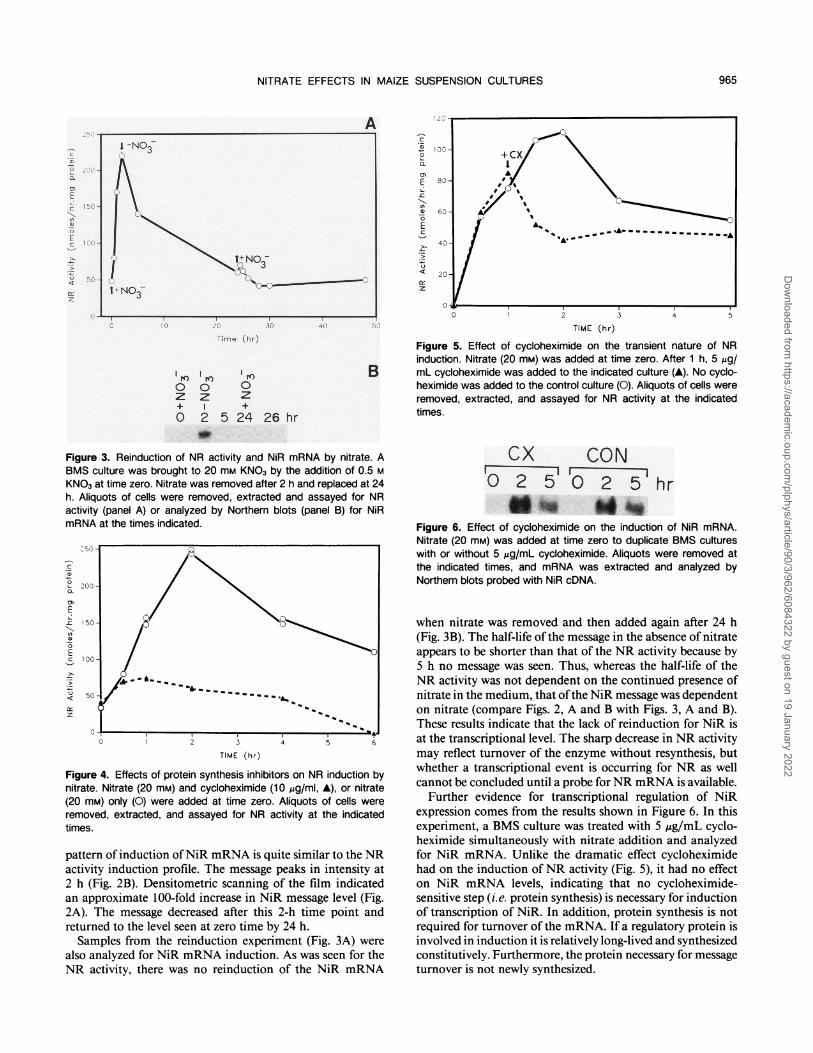
cxI0 2 5
CONI0Y 5hr
Figure 6. Effect of cycloheximide on the induction of NiR mRNA.Nitrate (20 mM) was added at time zero to duplicate BMS cultureswith or without 5 Ag/mL cycloheximide. Aliquots were removed atthe indicated times, and mRNA was extracted and analyzed byNorthern blots probed with NiR cDNA.
when nitrate was removed and then added again after 24 h(Fig. 3B). The half-life of the message in the absence of nitrateappears to be shorter than that of the NR activity because by5 h no message was seen. Thus, whereas the half-life of theNR activity was not dependent on the continued presence ofnitrate in the medium, that ofthe NiR message was dependenton nitrate (compare Figs. 2, A and B with Figs. 3, A and B).These results indicate that the lack of reinduction for NiR isat the transcriptional level. The sharp decrease in NR activitymay reflect turnover of the enzyme without resynthesis, butwhether a transcriptional event is occurring for NR as wellcannot be concluded until a probe forNR mRNA is available.
Further evidence for transcriptional regulation of NiRexpression comes from the results shown in Figure 6. In thisexperiment, a BMS culture was treated with 5 ,ug/mL cyclo-heximide simultaneously with nitrate addition and analyzedfor NiR mRNA. Unlike the dramatic effect cycloheximidehad on the induction ofNR activity (Fig. 5), it had no effecton NiR mRNA levels, indicating that no cycloheximide-sensitive step (i.e. protein synthesis) is necessary for inductionof transcription of NiR. In addition, protein synthesis is notrequired for turnover of the mRNA. If a regulatory protein isinvolved in induction it is relatively long-lived and synthesizedconstitutively. Furthermore, the protein necessary for messageturnover is not newly synthesized.
965
Dow
nloaded from https://academ
ic.oup.com/plphys/article/90/3/962/6084322 by guest on 19 January 2022

Plant Physiol. Vol. 90, 1989
DISCUSSION
Nitrate reductase activity in logarithmically growing BMSsuspension cultures was induced 100-fold to 200-fold afterthe addition of 20 mm nitrate. The mRNA for NiR was
induced approximately 100-fold by the same treatment andfollowed the same pattern in that both levels peaked 2 h afterthe addition of nitrate and decreased thereafter. The transientnature of this induction, although quite similar to that re-
ported for roots of maize seedlings treated with 20 mm nitrate(14), is in contrast to that found in soybean (8), spinach (18),or morning glory (28) cell suspensions. The age of our BMScultures affects the degree of induction but not the transientnature of the phenomenon (data not shown).The removal of nitrate from the culture medium after 2 h
of incubation followed by its readdition 22 h later did notresult in the reinduction of NR activity or NiR mRNA. Inmaize seedlings, reinduction was found to occur after 48 h(14), whereas in the BMS cells, reinduction did not occur
even 72 h after the initial addition of the nitrate. There are
several possible explanations for the rapid decrease in NRactivity followed by the insensitivity to the reintroduction ofnitrate: (a) specific proteases and/or RNases could be nitrate-induced that are longer lived than either the mRNA or theNR protein; (b) a long-lived, constitutive induction-mediatingprotein might be decreased in concentration through degra-dation or modification (perhaps by binding nitrate); (c) an
additional regulatory protein may be repressing the synthesisor translation ofadditional mRNA; or (d) nitrate that remainswithin the cells continues to exert some sort of substrateinhibition.To explore the first three possibilities, we studied the effects
ofthe protein synthesis inhibitor cycloheximide on the induc-tion of both NR activity and NiR mRNA. Cycloheximidewas effective at preventing the nitrate-mediated induction ofNR activity when added simultaneously with or prior to thenitrate. The apparent de novo synthesis of NR is in goodagreement with the results ofZielke and Filner (27) in tobaccocell lines as well as Maki et al. ( 18) in cultured spinach cells.However, the results are in contrast to findings in the Chlorellasystem, where activation of a previously synthesized but in-active NR has been reported (5). Cycloheximide did not affectthe decrease in activity, indicating that this decrease is prob-ably not due to the synthesis of a protease. This rules out one
part of the first model.Cycloheximide had no effect on induction or loss of NiR
mRNA. In mammalian cells, transiently induced genes havebeen shown to have superinduced levels of mRNA in thepresence of protein synthesis inhibitors (23). This is presum-ably due to the loss ofa labile protein causing either decreaseddegradation of the mRNA or to increased mRNA synthesis.No effect of cycloheximide was seen on the level of NiRmRNA even when it was added to the cells 24 h prior to theaddition of nitrate (data not shown). This is evidence againstthe third model. The proteins involved in induction of NiRmRNA synthesis thus appear to be present constitutively as
stated in possibility two. Furthermore, there does not appearto be a labile protein whose absence would cause superinduc-
tion in the NiR mRNA level. The fourth model has not beenruled out.The end product of the nitrate assimilation pathway is
ammonia. The induction of NR activity or NiR mRNA bynitrate in these BMS suspension cultures was not preventedby the presence of ammonia, whereas Zink and Veliky (29)reported repression-like effects by ammonium in morningglory suspension cells. Ammonia has been reported not toaffect induction in maize seedlings (14), but it has beenreported to be inhibitory in Chlorella (5) and fungi (1).
Chlorate, which is an analog of nitrate useful in uptakeanalyses as well as a substrate for NR (10), did not induceNR activity under our conditions. Furthermore, chlorate didnot prevent the induction by nitrate, although it appeared tocompete at low nitrate concentrations. Thus chlorate is notan analog of nitrate at the regulatory or "inducer" level.
In summary, nitrate induces the expression ofNR activityand NiR mRNA synthesis with similar kinetics. Both aretransiently induced with maximal levels reached after 2 h.After maximal levels are reached, there is a rapid decrease inboth NR activity and NiR mRNA levels. When the cells aretransferred to nitrate-free medium 2 h after the addition ofnitrate and grown for an additional 22 h, reinduction bynitrate does not occur. These results are similar to what wehave found for hydroponically grown maize seedlings (14).Thus, Black Mexican Sweet maize suspension cultures appearto be a good model system to study the regulation of nitrateassimilation.
LITERATURE CITED
1. Arst HN, Cove DJ (1973) Nitrogen metabolite repression inAspergillus nidulans. Mol Gen Genet 126: 111-141
2. Arst HN, Scazzochio C (1985) Formal genetics and molecularbiology of the control of gene expression in Aspergillus nidu-lans. In JW Bennett, L Lasure, eds, Gene Manipulations inFungi. Academic Press, Inc., Orlando, pp 309-343
3. Back E, Burkhart W, Moyer M, Privalle LS, Rothstein S (1988)Isolation ofcDNA clones coding for spinach nitrite reductase:complete sequence and nitrate induction. Mol Gen Genet 212:20-26
4. Bradford MM (1976) A rapid and sensitive method for thequantitation of proteins utilizing the principle of protein dyebinding. Anal Biochem 72: 248-254
5. Funkhouser EA, Shen T-C, Ackermann R (1980) Synthesis ofnitrate reductase in Chlorella. I. Evidence for an inactiveprotein precursor. Plant Physiol 65: 939-943
6. Guerrero MG, Vega JM, Losada M (1981) The assimilatorynitrate-reducing system and its regulation. Annu Rev PlantPhysiol 32: 169-204
7. Gupta SC, Beevers L (1987) Regulation of nitrite reductase. Cell-free translation and processing. Plant Physiol 83: 750-754
8. Hahlbrock K (1974) Correlation between nitrate uptake, growthand changes in metabolic activities of cultured plant cells. InHE Street, ed, Proceedings of the International Congress onPlant Tissue Cell Culture. Academic Press, London, Vol. 3. pp363-378
9. Hewitt EJ, Hucklesby DP, Notton DA (1976) Nitrate metabo-lism. In J Bonner, JE Varner, eds, Plant Biochemistry. Aca-demic Press, New York, pp 663-672
10. Hofstra JJ (1977) Chlorate toxicity and nitrate reductase activityin tomato plants. Physiol Plant 41: 65-69
11. Heimer YM, Riklis E (1979) On the mechanism ofdevelopmentof nitrate reductase activity in tobacco cells. Plant Sci Lett 16:135-138
12. Ingemarsson B (1987) Nitrogen utilization in Lemna I. Relations
PRIVALLE ET AL.966
Dow
nloaded from https://academ
ic.oup.com/plphys/article/90/3/962/6084322 by guest on 19 January 2022

NITRATE EFFECTS IN MAIZE SUSPENSION CULTURES
between net nitrate flux, nitrate reduction, and in vitro activityand stability of nitrate reductase. Plant Physiol 85: 856-859
13. Jackson WA, Flesher D, Hageman RH (1973) Nitrate uptake bydark-grown corn seedlings. Some characteristics of apparentinduction. Plant Physiol 51: 120-127
14. Kramer V, Lahners K, Privalle LS, Rothstein S (1989) Transientaccumulation of nitrate reductase mRNA in maize followingthe addition of nitrate. Plant Physiol (in press)
15. Lahners K, Kramer V, Back E, Privalle LS, Rothstein S (1988)Molecular cloning of complementary DNA encoding maizenitrite reductase: molecular analysis and nitrate induction.Plant Physiol 88: 741-746
16. Lancaster JR, Batie CJ, Kamin H, Knaff DB (1982) Purificationof spinach nitrite reductase. In M Edelman, RB Hallick, NHChua, eds, Methods in Chloroplast Molecular Biology.Biomedical Press, New York, pp 723-734
17. MacKown CT (1987) Nitrate uptake and assimilation followingnitrate deprivation. J Exp Bot 38: 1079-1090
18. Maki H, Yamagishi K, Sato T, Ogura N, Nakagawa H (1986)Regulation of nitrate reductase activity in cultured spinachcells as studied by an enzyme-linked immunosorbent assay.Plant Physiol 82: 739-741
19. Mohanty B, Fletcher JS (1976) Ammonium influence on thegrowth and nitrate reductase activity of Paul's Scarlet rosesuspension cultures. Plant Physiol 58: 152-155
20. Murashige T, Skoog F (1962) A revised medium for rapid growthand bioassays with tobacco tissue cultures. Physiol Plant 15:473-497
21. Nakagawa H, Yonemura Y, Yamamoto H, Sato T, Ogura N,Sato R (1985) Spinach nitrate reductase, purification, molec-ular weight, and subunit composition. Plant Physiol 77: 124-128
22. Nelson RS, Horn ME, Harper JE, Widholm J (1984) Nitratereductase activity and nitrogenous gas evolution from hetero-trophic photomixotrophic and photoautotrophic soybean sus-pension cultures. Plant Sci Lett 34: 145-152
23. Pachter JS, Yen TJ, Cleveland DW (1987) Autoregulation oftubulin expression is achieved through specific degradation ofpolysomal tubulin mRNAs. Cell 51: 283-292
24. Remmler JL, Campbell WH (1986) Regulation of corn leafnitrate reductase II. Synthesis and turnover of the enzyme'sactivity and protein. Plant Physiol 80: 442-447
25. Roa LVM, Datta N, Mahadeva M, Guha-Mukherjee S, SoporySK (1984) Influence of cytokinins and phytochrome on nitratereductase activity in etiolated leaves of maize. Phytochemistry23: 1875-1879
26. Shimamoto K, King PJ (1983) Adhl-0:a selectable marker inZea mays cell culture. Mol Gen Genet 191: 271-275
27. Zielke HR, Filner P (1971) Synthesis and turnover of nitratereductase induced by nitrate in cultured tobacco cells. J BiolChem 246: 1772-1779
28. Zink MW (1982) Regulation of nitrate reductase by variousnitrogen sources in cultured Ipomoea sp. Can J Bot 60: 386-396
29. Zink MW, Veliky IA (1977) Nitrogen assimilation and regulationof nitrate and nitrite reductases in cultured Ipomoea cells. CanJ Bot 55: 1557-1568
967
Dow
nloaded from https://academ
ic.oup.com/plphys/article/90/3/962/6084322 by guest on 19 January 2022