relationship of nk3 receptor- immunoreactivity to subpopulations of neurons in rat spinal cord
TRANSCRIPT

Relationship of NK3 Receptor-Immunoreactivity to Subpopulations of
Neurons in Rat Spinal Cord
V.S. SEYBOLD,1* I. GRKOVIC,1 A.L. PORTBURY,1 Y.-Q. DING,2 R. SHIGEMOTO,2
N. MIZUNO,2 J.B. FURNESS,1 AND B.R. SOUTHWELL1
1Department of Anatomy and Cell Biology, University of Melbourne, 3052 Victoria, Australia2Department of Morphological Brain Science, Faculty of Medicine, Kyoto University,
Kyoto, 606-01 Japan
ABSTRACTThe distribution of immunoreactivity to the neurokinin3 receptor (NK3R) was examined
in segments C7, T11-12, L1-2, and L4-6 of the rat spinal cord. NK3R immunoreactivity wasvisualized by using two antisera generated against sequences of amino acids contained in theC-terminal region of the NK3R. NK3R-immunoreactive cells were numerous in the substantiagelatinosa of all spinal segments examined as well as the dorsal commissural nucleus of spinalsegments L1-2. Isolated, immunoreactive cells were scattered throughout other regions of thespinal cord. The relationship of NK3R-immunoreactivity with neurons was demonstrated bycolocalization with microtubule associated protein 2-immunoreactivity in individual cells.Within neurons, NK3R-immunoreactivity was associated predominately with the plasmamembrane of cell bodies and dendrites. Within the substantia gelatinosa, 86% of nitric oxidesynthase (NOS)-immunoreactive neurons were also NK3R-immunoreactive. Although NOS-immunoreactive neurons were found throughout all other regions of the spinal cord in thesegments examined, these were not NK3R-immunoreactive. When preganglionic sympatheticneurons in spinal segments T11-12 and L1-2 were visualized by intraperitoneal injection ofFluorogold, less than 1% of the Fluorogold-labeled neurons were also immunoreactive forNK3R. The large number of NK3R-immunoreactive neurons in the substantia gelatinosasuggests that some effects of tachykinins on somatosensation may be mediated by NK3R. J.Comp. Neurol. 381:439–448, 1997. r 1997 Wiley-Liss, Inc.
Indexing terms: autonomic nervous system; neurokinin B; nitric oxide; sensory nervous system;
tachykinin
Themammalian tachykinins include substance P, neuro-kinin A, and neurokinin B. Although these peptides arecontained in axons that are distributed throughout thegray matter of the spinal cord, the greatest densities ofimmunoreactive varicosities occur in the substantia gelati-nosa and the region around the central canal at all levels ofthe spinal cord, as well as regions that contain pregangli-onic sympathetic and parasympathetic preganglionic neu-rons (Barber et al., 1979; Gibson et al., 1981; Zhuo andHelke, 1993). Thus, tachykinins have been implicated insomatosensation and control of sympathetic outflow, andthe physiological effects of substance P on nociception,blood pressure, and urine retention have been described.The physiological effects of tachykinins are mediated by
neurokinin1 (NK1), NK2, and NK3 receptors, and eachreceptor is characterized by a unique order of potency ofthe tachykinins. Substance P, neurokinin A, and neuroki-nin B are the preferred endogenous agonists of NK1, NK2,
and NK3 receptors, respectively. NK1 and NK2 receptorsmediate facilitatory effects of substance P and neurokininAon nociception.Although neurokinin B occurs in varicosi-ties within the substantia gelatinosa (Zhuo and Helke,1993), its role in somatosensation has not been resolved.Intrathecal injection of neurokinin B or senktide, anagonist at NK3 receptors, causes analgesia in behavioralassays, but the effect may reflect depression of motorfunction (Papir-Kircheli et al., 1987). Neurokinin B in-creases calcium conductance in primary afferent neurons
Contract grant sponsor: NIH; Contract grant numbers: NIH/NS17702,NH&MRC963213.*Correspondence to: Dr. Virginia S. Seybold, Department of Cell Biology
and Neuroanatomy, University of Minnesota, 4-135 Jackson Hall, 321Church Street, S.E., Minneapolis, MN 55455. E-mail: [email protected] 16 September 1996; Revised 31 December 1996; Accepted 4
January 1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 381:439–448 (1997)
r 1997 WILEY-LISS, INC.

in vitro (Inoue et al., 1995), and this effect could facilitatenociception.The hypothesis that NK3 receptors have a role in
somatosensation is supported by evidence that NK3 recep-tors are expressed in the substantia gelatinosa of thespinal cord. Messenger RNA for NK3 receptor (NK3R) inthe spinal cord has been demonstrated by Northern blotanalysis (McCarson and Krause, 1994; Suzuki et al.,1993), and the level of mRNAincreases in conjunction withperipheral inflammation (McCarson and Krause, 1994).Radiohistochemical studies using receptor autoradiogra-phy (Yashpal et al., 1990; Kar and Quirion, 1995) and insitu hybridization (Ding et al., 1996) concur that NK3receptors occur in the highest density within the substan-tia gelatinosa of the spinal cord. The relationship of NK3receptors to subpopulations of neurons in the spinal cordmay provide insights into the effects of tachykinins onsomatosensation.The location of NK3R-immunoreactivity (ir) in the spi-
nal cord has recently been described (Ding et al., 1996).NK3R-immunoreactive cells and processes are concen-trated in superficial laminae of the dorsal horn of the ratspinal cord. The goals of the present study were to extendthe initial observation in three ways. First, we examinedthe co-distribution of NK3R-ir with microtubule associ-ated protein 2 (MAP2) and glial fibrillary acidic protein, aprotein associated with astrocytes, in order to directlydemonstrate a relationship of NK3R-ir with neurons.Second, we determined the relationship of NK3R-ir withnitric oxide synthase (NOS)-immunoreactive neurons inselected thoracic and lumbar segments of the spinal cordbecause NOS-immunoreactive neurons are prevalent inregions where numerous NK3R-immunoreactive neuronsoccur. Finally, we assessed whether NK3R-immunoreac-tive neurons in the dorsal commissural nucleus of spinalsegments L1-2 were preganglionic neurons because thisregion contains preganglionic sympathetic neurons thatinnervate pelvic ganglia.
MATERIALS AND METHODS
Six adult male Sprague-Dawley rats (200–250 g) wereused in this study. Fluorogold injections and perfusionfixation were conducted under anesthesia induced bysodium pentobarbitone (60 mg/kg, ip). All procedures wereapproved by the University of Melbourne Animal Experi-mentation Ethics Committee.
Injection of Fluorogold
Preganglionic neurons were labeled in two animals withFluorogold by injection of Fluorogold (Fluorochrome, Inc.,Englewood, IL; 200 µl of a 1% solution) into the peritonealcavity through a small abdominal incision made undersurgical anesthesia (Anderson and Edwards, 1994). Ani-mals were perfused with fixative 4 days later.
Immunohistochemistry
Rats were perfused via the ascending aorta with 400 mlof Zamboni fixative (Stefanini et al., 1967). Followingperfusion, spinal segments C7, T11-12, and L1-6 and theircorresponding dorsal root ganglia were removed and im-mersed in fixative for an additional 2 hours. Tissue sampleswere then transferred to 5% sucrose in 0.1 M sodiumphosphate buffer, pH 7.2, for storage at 4°C. Cryostatsections (10 µm) through spinal cord and dorsal root
ganglia were cut in the coronal or horizontal plane of thespinal cord and mounted on slides coated with a 2%solution of 3-aminopropyltriethoxy-saline (Sigma Aldrich,Castle Hill, NSW).NK3 receptor-immunoreactivity was determined by im-
munofluorescence using antisera generated in rabbitsagainst sequences of amino acids contained in the C-terminal region of the NK3 receptor (R). One antiserumwas generated against a fusion protein containingNK3R388-452 (Ding et al., 1996, code DNK3R) and was used at a 1/50dilution. The second antiserum was generated againstNK3R410-417 coupled to keyhole limpet hemocyanin (Gradyet al., 1996, code AP951) and was used at a 1/500 dilution.Primary antisera were applied to tissue sections followingincubation of the tissue for 2 hours with 10% normal horseserum, 1% Triton X100 in phosphate-buffered saline (0.9%NaCl in 0.01 M sodium phosphate buffer, pH 7.2; PBS).Tissue sections were incubated with primary antiseraovernight at room temperature in a humid atmosphere.The rabbit anti-NK3R antisera were visualized by usingCy3-labeled donkey-anti rabbit IgG (1/500 dilution; Jack-son Immunoresearch, West Grove, PA). All antisera werediluted in PBS.Specificity of the antisera generated against the NK3R
was determined by incubation of the diluted antiseraovernight at 4°C with NK3R410-417 coupled to keyholelimpet hemocyanin (1 µg of conjugate/ml of diluted serum).The same concentration of conjugate was also incubatedwith a rabbit anti-substance P serum (1/1,000 dilution,Morris et al., 1986) in order to assess nonspecific interac-tion of the conjugate with the immunohistochemical proce-dure.In some experiments, the co-distribution of NK3R-ir
with other antigens was determined by the simultaneousvisualization of two antigens in one tissue section. Nitricoxide synthase (NOS)-ir was visualized by using a sheepanti-NOS serum (1/1,000 dilution; a gift of Dr. P.C. Emson)and fluorescein isothiocyanate (FITC)-labeled donkey anti-sheep IgG (1/25 dilution; Jackson Immunoresearch). Glialfibrillary acid protein (GFAP)-ir was detected by usingmouse anti-GFAP (1/200 dilution; Boehringer Mannheim,Indianapolis, IN), and microtubule associated protein-2(MAP2)-ir was detected by using mouse anti-MAP2 (1/100dilution, Sigma Chemical Co., St. Louis, MO). Antibodiesgenerated in mice were visualized using biotinylated horseanti-mouse IgG (1/200 dilution, Vector Lab, Burlingame,CA) and FITC-labeled streptavidin (1/50 dilution, Amer-sham, Melbourne, VIC). The distribution of fluorophoresin tissue sections was visualized with a Zeiss Axioskopfluorescence microscope or a BioRad MRC1000 confocalscanning laser microscope with a krypton/argon laserinstalled on an Olympus IMT2 microscope. Both micro-scopes are fitted with filter sets that allow the independentvisualization of Cy3 and FITC. Fluorogold could be distin-guished only on the Zeiss microscope. Immunofluorescencevisualized by using the Zeiss microscope was recorded byusing an ImagePoint cooled CCD camera (Photometrics,Ltd., Tucson, AZ) and V for Windows imaging software(Digital Optics, Ltd., Auckland, New Zealand). Imageswere processed using a contrast adjustment. All plateshave been made using Adobe Photoshop for Macintosh(ver. 3.0, Adobe Systems, Inc., Mountain View, CA).The frequencies of coexistence of NK3R-ir with Fluoro-
gold or NOS-ir were determined by counting neurons thatexhibited one or both markers within specified regions.
440 V.S. SEYBOLD ET AL.

Neurons within the substantia gelatinosa were counted intransverse sections through the spinal cord; neuronswithinthe dorsal commissural nucleus and intermediolateral cellcolumn were counted in horizontal sections. To avoiddouble counts, only neurons with nuclei were counted, andsections selected for counting were separated by 50 µm.
RESULTS
Distribution of NK3R-immunoreactivityin the rat spinal cord
NK3R-immunoreactivity was distributed throughout thegray matter of the spinal cord at all levels examined (C7,T11-12, L1-2, and L4-6; Fig. 1). Results were similar withbothNK3R-antisera, but immunofluorescencewas brighter
with antiserum DNK3R (Fig. 2). Immunoreactive pro-cesses were most numerous in the substantia gelatinosa ofthe dorsal horn at all levels examined, and the frequency ofoccurrence of the processes was particularly evident inhorizontal sections where the processes were parallel tothe plane of the section. In cross-section, some processesappeared as annular profiles, approximately 1–2 µm indiameter (Fig. 3C). The number of immunoreactive pro-cesses in areas outside of the substantia gelatinosa wasgreatest at lumbar levels of the spinal cord. Immunoreac-tive processes were also evident in the white matter,especially in the lateral funiculus of lumbar segments(Figs. 1, 4). No immunoreactivity was observed in thedorsal root ganglia that were sampled (C7, T11-12, L1-2,and L4-6).
Fig. 1. NK3 receptor immunoreactivity (NK3R-ir) in a coronalsection through spinal segment L4 (montage of four confocal images,total optical thickness of 2 µm). The tissue section was stained withantiserum DNK3R. The greatest density of NK3R-ir occurred insubstantia gelatinosa (SG). The single arrow indicates an NK3R-
immunoreactive cell body in the marginal zone; the double arrowindicates a cell body of the lateral spinal nucleus. The arrowheadsindicate immunoreactive processes in the lateral funiculus. CC, centralcanal.
NK3 RECEPTORS IN RAT SPINAL CORD 441

NK3R immunoreactive perikarya exhibited the bright-est staining at the level of the cell membrane (Figs. 2, 3). Incoronal sections, immunoreactive perikarya were mostnumerous in the substantia gelatinosa at all levels of thespinal cord (an average of 10 cells/side/section), and themajority of immunoreactive perikarya occurred in theinner half of the region. The perikarya were approximately8–10 µm in diameter, and they appeared evenly distrib-uted in the medial to lateral dimension. Isolated peri-karya, approximately 10–20 µm in diameter, were sparselydistributed throughout all regions of the gray matter at alllevels examined, including the marginal zone (Figs. 1, 3A),the lateral spinal nucleus (Figs. 1, 3B) and the intermedio-lateral cell column (Fig. 4). The most intensely stainedperikarya and processes occurred in the dorsal commis-sural nucleus of spinal segments L1-2 (Fig. 2A,B).Specificity of the NK3R-antisera was tested by absorp-
tion of each antiserum with the antigen used to generatethe antiserum AP951. When either NK3R-antiserum waspreincubated with this antigen, immunoreactivity waseliminated (Fig. 2). It is not likely that the change inimmunoreactivity was the result of a nonspecific effect ofthe antigen since incubation of an anti-substance P serum
with the same concentration of antigen did not alter thedetection of substance P-ir in rat spinal cord.
Distribution NK3R-immunoreactivityrelative to GFAP- and
MAP2-immunoreactivity
In order to determine whether the NK3R-ir occurred onneurons or glia, tissue sections were incubated for thesimultaneous visualization of two antigens in one tissuesection. There was no co-localization of GFAP-ir andNK3R-ir, but there was extensive overlap in the occur-rence of NK3R-ir and MAP2-ir in individual cells. Allcellular profiles that were NK3R-immunoreactive exhib-ited MAP2-immunoreactive cytoplasm (Fig. 3). Further-more, NK3R-immunoreactive processes in the lateral fu-niculus were also MAP2-ir (Fig. 4) suggesting that theseprocesses were dendrites rather than axons.
Distribution NK3R-immunoreactivityrelative to NOS-immunoreactive neurons
Within the rat spinal cord, NOS-containing neurons areprominent in the substantia gelatinosa and regions that
Fig. 2. Specificity of antisera generated against amino acids withinthe C-terminal region of the NK3R. A–C: Images of the dorsalcommissural nucleus in adjacent, horizontal sections through spinalsegment L2. Similar cellular profiles were visualized by using antise-rum DNK3R (A) and AP951 (B), although immunofluorescence wasstronger with antiserum DNK3R. Immunoreactivity with antiserumAP951 was absorbed by preincubation of the antiserum with the
immunogen against which it was raised (C). D, E: Images of thesubstantia gelatinosa in adjacent tissue sections. NK3R-ir was visual-ized by using DNK3R (D), and preincubation of this antiserum withNK3R410-418 conjugated to keyhole limpet hemocyanin absorbed thestaining (E).All images were obtained by using the CCD camera on theZeiss microscope. Scale bar in A applies to A–C; bar in D applies toD, E.
442 V.S. SEYBOLD ET AL.

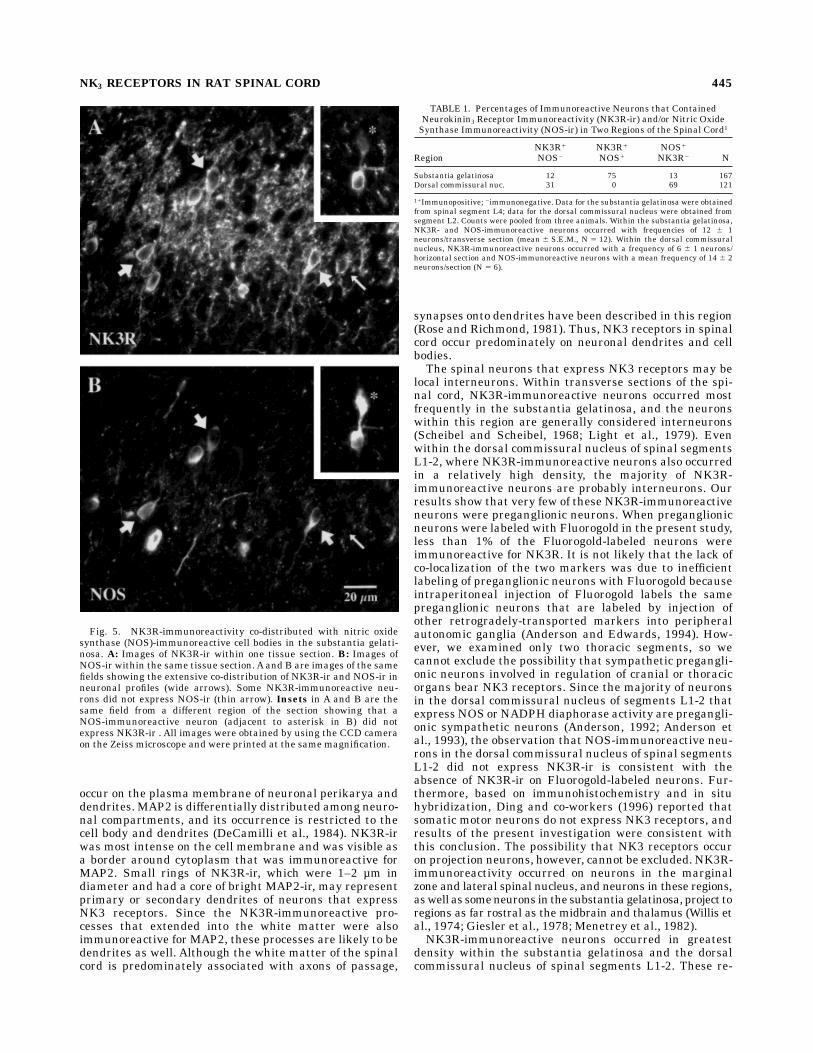
give rise to preganglionic sympathetic neurons, includingthe dorsal commissural nucleus of spinal segments L1-2(Anderson, 1992; Dun et al., 1992). NK3R-ir and NOS-irwere detected simultaneously in tissue sections in order todetermine whether NOS-immunoreactive neurons alsoexpress NK3 receptors. Within the substantia gelatinosa,NK3R- and NOS-immunoreactive perikarya occurred witha similar frequency within all spinal segments examined(Fig. 5). Within spinal segment L4 86% of NK3R-immuno-reactive neurons of the substantia gelatinosa were alsoNOS-immunoreactive, and a comparable proportion ofNOS-immunoreactive neurons expressed NK3R-ir (Table1). Thus, there was a high degree of overlap of NOS andNK3 receptors in neurons within the substantia gelati-nosa. In contrast, NOS-immunoreactive neurons were alsonumerous within the intermediolateral cell column ofspinal segments T11-12, but NOS-immunoreactive neu-rons within this region rarely expressed NK3R-ir. How-
ever, the occasional NK3R-immunoreactive neuron thatoccurred in the intermediolateral cell column exhibited amoderate level of NOS-ir. In the dorsal commissuralnucleus of spinal segments L1-2, where NK3R- and NOS-immunoreactive neurons were both numerous, there wasno overlap in the occurrence of the two antigens (Fig. 6A,B,Table 1). NK3R-immunoreactive neurons that were scat-tered throughout other regions of the gray matter did notcontain NOS-ir within the spinal segments examined (C7,T11-12, and L4-6).
Relationship of NK3R-immunoreactivityin the dorsal commissural nucleus
to preganglionic neurons
Because NK3R-immunoreactive neurons were numer-ous in the dorsal commissural nucleus of spinal segmentsL1-2, we examined whether NK3R-immunoreactive neu-
Fig. 3. NK3R-immunoreactivity was associated with neurons. A,C, E: Confocal images of NK3R-ir (stained with DNK3R). B, D, F:Images of microtubule associated protein-2 (MAP2)-ir within the sametissue sections. A/B, C/D, E/F are pairs of optical sections (A–D were0.5 µm thick; E and F were 1 µm thick) viewed under conditions fordifferent fluorophores. A and B illustrate a neuron in the marginalzone, C and D neurons in the substantia gelatinosa, and E and F a
neuron in the lateral spinal nucleus. NK3R-immunofluorescence isbrightest on the plasmamembrane of the cells. MAP2-immunofluores-cence is moderate in the cytoplasm of cells that also expressedNK3R-ir (asterisks). The arrow in C indicates a smaller, annularNK3R-immunoreactive profile, likely to be a dendrite, that is associ-ated with bright MAP2-ir in D (arrow). All images were printed at thesame magnification.
NK3 RECEPTORS IN RAT SPINAL CORD 443

rons were preganglionic neurons. Preganglionic sympa-thetic neurons were labeled with Fluorogold followingintraperitoneal injection of the dye, and tissue sectionsthrough T11-12 and L1-2 were stained for NK3R-ir. Gener-ally, there was little overlap in the distribution Fluorogold-labeled and NK3R-immunoreactive neurons, although0.5–1% of Fluorogold-labeled neurons within each regioncontained NK3R-ir (Fig. 6C–F, Table 2).
DISCUSSION
The present work extends the initial description of thedistribution of NK3R-ir in rat spinal cord (Ding et al.,1996) by demonstrating that the immunoreactivity occurson neuronal cell bodies and dendrites and by the discoverythat there is a high degree of co-localization of NOS- andNK3R-ir in neurons within the substantia gelatinosa. TheNK3R-ir is likely to be specific to NK3R because twoantisera produced similar results, and immunoreactivity
was absorbed by a peptide contained in the C-terminalregion of the NK3R. Furthermore, both antisera have beencharacterized using receptor transfected cells and shownto reveal NK3 receptors but not NK2 or NK1 receptors byimmunocytochemistry and Western blot analysis (Ding etal., 1996; Grady et al., 1996).In the initial report, the relationship of NK3R-ir with
neurons was based on morphology, and the local synthesisof NK3R in the spinal cord was confirmed using in situhybridization (Ding et al., 1996). In the present report, therelationship of NK3R-ir with neurons was directly demon-strated by the colocalization of NK3R-ir and MAP2-irwithin individual cells. MAP2 is a microtubule associatedprotein that occurs only in neurons (DeCamilli et al.,1984). The lack of co-distribution of NK3R-ir and GFAP-irindicates that NK3 receptors are not detectable on astro-cytes in the spinal cord.Simultaneous visualization of NK3R-ir and MAP2-ir
using confocal microscopy indicated that NK3 receptors
Fig. 4. NK3R-immunoreactive processes in the lateral funiculuswere associated with MAP2-ir. A, B: Confocal images of MAP2-ir. C:Aconfocal image of NK3R-ir. The area shown in B is indicated by the boxin A. B and C are the same optical section (0.5 µm thickness) viewed
under conditions for different fluorophores. Short and long arrows in Band C indicate processes that were immunoreactive for MAP2 andNK3R visualized with DNK3R.
444 V.S. SEYBOLD ET AL.
occur on the plasma membrane of neuronal perikarya anddendrites. MAP2 is differentially distributed among neuro-nal compartments, and its occurrence is restricted to thecell body and dendrites (DeCamilli et al., 1984). NK3R-irwas most intense on the cell membrane and was visible asa border around cytoplasm that was immunoreactive forMAP2. Small rings of NK3R-ir, which were 1–2 µm indiameter and had a core of bright MAP2-ir, may representprimary or secondary dendrites of neurons that expressNK3 receptors. Since the NK3R-immunoreactive pro-cesses that extended into the white matter were alsoimmunoreactive for MAP2, these processes are likely to bedendrites as well. Although the white matter of the spinalcord is predominately associated with axons of passage,
synapses onto dendrites have been described in this region(Rose and Richmond, 1981). Thus, NK3 receptors in spinalcord occur predominately on neuronal dendrites and cellbodies.The spinal neurons that express NK3 receptors may be
local interneurons. Within transverse sections of the spi-nal cord, NK3R-immunoreactive neurons occurred mostfrequently in the substantia gelatinosa, and the neuronswithin this region are generally considered interneurons(Scheibel and Scheibel, 1968; Light et al., 1979). Evenwithin the dorsal commissural nucleus of spinal segmentsL1-2, where NK3R-immunoreactive neurons also occurredin a relatively high density, the majority of NK3R-immunoreactive neurons are probably interneurons. Ourresults show that very few of these NK3R-immunoreactiveneurons were preganglionic neurons. When preganglionicneurons were labeled with Fluorogold in the present study,less than 1% of the Fluorogold-labeled neurons wereimmunoreactive for NK3R. It is not likely that the lack ofco-localization of the two markers was due to inefficientlabeling of preganglionic neurons with Fluorogold becauseintraperitoneal injection of Fluorogold labels the samepreganglionic neurons that are labeled by injection ofother retrogradely-transported markers into peripheralautonomic ganglia (Anderson and Edwards, 1994). How-ever, we examined only two thoracic segments, so wecannot exclude the possibility that sympathetic pregangli-onic neurons involved in regulation of cranial or thoracicorgans bear NK3 receptors. Since the majority of neuronsin the dorsal commissural nucleus of segments L1-2 thatexpress NOS or NADPH diaphorase activity are pregangli-onic sympathetic neurons (Anderson, 1992; Anderson etal., 1993), the observation that NOS-immunoreactive neu-rons in the dorsal commissural nucleus of spinal segmentsL1-2 did not express NK3R-ir is consistent with theabsence of NK3R-ir on Fluorogold-labeled neurons. Fur-thermore, based on immunohistochemistry and in situhybridization, Ding and co-workers (1996) reported thatsomatic motor neurons do not express NK3 receptors, andresults of the present investigation were consistent withthis conclusion. The possibility that NK3 receptors occuron projection neurons, however, cannot be excluded. NK3R-immunoreactivity occurred on neurons in the marginalzone and lateral spinal nucleus, and neurons in these regions,aswell as someneurons in the substantia gelatinosa, project toregions as far rostral as the midbrain and thalamus (Willis etal., 1974; Giesler et al., 1978; Menetrey et al., 1982).NK3R-immunoreactive neurons occurred in greatest
density within the substantia gelatinosa and the dorsalcommissural nucleus of spinal segments L1-2. These re-
Fig. 5. NK3R-immunoreactivity co-distributed with nitric oxidesynthase (NOS)-immunoreactive cell bodies in the substantia gelati-nosa. A: Images of NK3R-ir within one tissue section. B: Images ofNOS-ir within the same tissue section. A and B are images of the samefields showing the extensive co-distribution of NK3R-ir and NOS-ir inneuronal profiles (wide arrows). Some NK3R-immunoreactive neu-rons did not express NOS-ir (thin arrow). Insets in A and B are thesame field from a different region of the section showing that aNOS-immunoreactive neuron (adjacent to asterisk in B) did notexpress NK3R-ir . All images were obtained by using the CCD cameraon the Zeiss microscope and were printed at the same magnification.
TABLE 1. Percentages of Immunoreactive Neurons that ContainedNeurokinin3 Receptor Immunoreactivity (NK3R-ir) and/or Nitric OxideSynthase Immunoreactivity (NOS-ir) in Two Regions of the Spinal Cord1
RegionNK3R1
NOS2
NK3R1
NOS1
NOS1
NK3R2 N
Substantia gelatinosa 12 75 13 167Dorsal commissural nuc. 31 0 69 121
11Immunopositive; 2immunonegative. Data for the substantia gelatinosa were obtainedfrom spinal segment L4; data for the dorsal commissural nucleus were obtained fromsegment L2. Counts were pooled from three animals. Within the substantia gelatinosa,NK3R- and NOS-immunoreactive neurons occurred with frequencies of 12 6 1neurons/transverse section (mean 6 S.E.M., N 5 12). Within the dorsal commissuralnucleus, NK3R-immunoreactive neurons occurred with a frequency of 6 6 1 neurons/horizontal section and NOS-immunoreactive neurons with a mean frequency of 14 6 2neurons/section (N 5 6).
NK3 RECEPTORS IN RAT SPINAL CORD 445

gions are involved in somatosensory and autonomic sys-tems, respectively, but the role of NK3 receptors in somato-sensation and autonomic functions remains to be resolved.Both areas are densely innervated by tachykinin-contain-ing terminals (Barber et al., 1979; Gibson et al., 1981; ZhuoandHelke, 1993), andneurokininB-containing terminalsmayarise from neurons intrinsic to the spinal cord (Ogawa et al.,1985;WardenandYoung, 1988) or thediencephalon (ZhuoandHelke, 1993). Therefore, it is likely that endogenousligands are released in these areas.Within the dorsal horn of the spinal cord, electrophysi-
ological studies have reported varied results concerningeffects of NK3R agonists on neuronal responses. Applica-
tion of NK3R agonists to neurons in the substantia gelati-nosa (Bleazard et al., 1994) or the ventral horn (Ireland etal., 1992; Fisher et al., 1994) of the neonatal rat spinal cordin vitro results in depolarization of neurons. The reportthat tetrodotoxin blocks the effect of an NK3R agonist onmotor neurons (Fisher et al., 1994) is consistent with theabsence of NK3R-ir on these neurons. However, a compa-rable study conducted in vitro on the adult rat spinal cordfound no effect of an NK3R agonist on neurons in thesubstantia gelatinosa (Bentley and Gent, 1995). The differ-ence in results could reflect a greater expression of NK3receptors in neonatal rats (Kar and Quirion, 1995). Alter-natively, activation of NK3 receptors in adult spinal cordmay modulate the properties of receptors for other neuro-transmitters.Although senktide, a selective agonist for theNK3R, did not alter background firing of spinal neurons orneuronal responses to noxious heat, senktide significantlyenhanced the firing of dorsal horn neurons in response tokainate (Cumberbatch et al., 1995). The absence of NK3R-irassociated with dorsal root ganglia of adult rats (Ding etal., 1996; present results) indicates that effects of tachyki-nins on primary afferent neurons in vitro (Bowie et al.,1994; Inoue et al., 1995) may be due to activity of thetachykinins at another neurokinin receptor, greater expres-sion of NK3R in neonatal compared to adult rats, orexpression of a variant of the NK3R in dorsal root ganglia.
TABLE 2. Percentages of Labeled Neurons that Contained Fluorogold(FG) and/or NK3R-Immunoreactivity in TwoAutonomic Regions of the
Spinal Cord1
RegionNK3R1
FG2
NK3R1
FG1
FG1
NK3R2 N
Intermediolateral cell column (T11/12) 7 1 92 206Dorsal commissural nuc. (L1–2) 32 1 66 200
11Immuno- or FG-positive; 2immuno- or FG-negative. Data were pooled from twoanimals. Within the intermediolateral cell column, NK3R-immunoreactive neuronsoccurred with a frequency of 3 6 1 neurons/horizontal section (mean 6 S.E.M.) andFG-labeled neurons with a frequency of 32 6 6 neurons/section (N 5 6). Within thedorsal commissural nucleus, NK3R-immunoreactive neurons occurred with a frequencyof 9 6 2 neurons/horizontal section and FG-labeled neurons with a frequency of 19 6 5neurons/section (N 5 7).
Fig. 6. There was little overlap of NK3R-ir with NOS- and Fluoro-gold-labeled neurons in regions of preganglionic sympathetic neurons.NK3R-ir was visualized with antiserum AP951 in horizontal sectionsthrough the dorsal commissural nucleus of spinal segment L2 (A,E)and the intermediolateral cell column of segment T12 (C). B: NOS-irin the same field shown inA; there is no co-localization of NK3R-ir andNOS-ir.D, F: Preganglionic sympathetic neurons labeled with Fluoro-gold in the same fields shown in C and E, respectively. Comparison of
C and D illustrates the general absence of NK3R-ir (C) in Fluorogold-labeled neurons (D; asterisk marks the position of the NK3R-immunoreactive profile seen in C). Comparison of E and F illustratesthe occasional NK3R-immunoreactive neuron (asterisk) that waslabeled with Fluorogold (F). All images were obtained by using theCCD camera on the Zeiss microscope. Scale bar in B applies to A, B;bar in F applies to C-F.
446 V.S. SEYBOLD ET AL.

Activation of NK3R on neurons in the substantia gelati-nosa may release NO, GABA and glycine from the NK3R/NOS neurons and unknown substances from the NK3R-immunoreactive neurons that were not NOS-immunoreactive. The release of GABA and glycine ispostulated because NOS neurons within the substantiagelatinosa also contain these substances (Spike et al.,1993). Whereas nitric oxide promotes hyperalgesia (Haleyet al., 1992; Meller et al., 1992), GABA and glycineattenuate hyperalgesia (Yaksh, 1989; Hao et al., 1992;Sivilotti andWoolf, 1994). Thus, effects of NK3R activationare likely to be complex.Antinociceptive effects of intrathe-cally injected NK3R agonists have been described inbehavioral studies (Laneuville et al., 1988; Couture et al.,1993), but these results may be confounded by the motordeficits caused by the agonists (Papir-Kircheli et al., 1987).The availability of a new, selective NK3R antagonist(Oury-Donat et al., 1995) may help resolve the role of NK3receptors in somatosensory pathways.The dorsal commissural nucleus of spinal segments L1-2
contains preganglionic sympathetic neurons that projectto the pelvic and inferior mesenteric ganglia (Strack et al.,1988; Hosoya et al., 1994, Grkovic and Anderson, 1996).Because NK3R-immunoreactive neurons near the centralcanal were especially numerous within segments L1-2relative to the other segments examined (C7, T11-12, andL4-6), it is likely that NK3 receptors participate in path-ways involved in the central regulation of pelvic viscera.However, preganglionic neurons within the dorsal commis-sural nucleus of spinal segments L1-2 were not NK3R-immunoreactive. The observations in the present reportare consistent with the reported absence of NK3R-ir insuperior cervical or pelvic ganglia (Ding et al., 1996). Sincea high proportion of preganglionic neurons within thedorsal commissural nucleus of spinal segments L1-2 ex-press NK1 receptors (Grkovic and Anderson, 1996), theeffects of tachykinins on the function of pelvic viscera arelikely to be mediated directly by NK1 receptors, andindirectly by NK3 receptors.
CONCLUSIONS
This work has shown that NK3 receptors are locatedpost-synaptically on spinal neurons. There was a strongrelationship between NK3 receptors and NOS neurons inthe substantia gelatinosa, a region rich in tachykinin-containing nerve terminals. Thus, it is likely that some ofthe transmitter actions of tachykinins in sensory path-ways are mediated via NK3 receptors.
ACKNOWLEDGMENTS
The authors are grateful to Dr. Colin Anderson fordiscussions of the organization of spinal autonomic nuclei.V.S.S. was a visiting research fellow in the Department ofAnatomy and Cell Biology, University of Melbourne.
LITERATURE CITED
Anderson, C.R. (1992) NADPH diaphorase-positive neurons in the ratspinal cord include a subpopulation of autonomic preganglionic neu-rons. Neurosci. Lett. 139:280–284.
Anderson, C.R., S.L. Edwards, J.B. Furness, D.S. Bredt, and S.H. Snyder(1993) The distribution of nitric oxide synthase-containing autonomicpreganglionic terminals in the rat. Brain Res. 614:78–85.
Anderson, C.R., and S.L. Edwards (1994) Intraperitoneal injection offluorogold reliably labels all sympathetic preganglionic neurons in therat. J. Neurosci. Methods 53:137–141.
Barber, R.P., J.E. Vaughn, J.R. Slemmon, P.M. Salvaterra, E. Roberts, andS.E. Leeman (1979) The origin, distribution and synaptic relationshipsof substance P axons in rat spinal cord. J. Comp. Neurol. 184:331–352.
Bentley, G.N., and J.P. Gent (1995) Neurokinin actions on substantiagelatinosa neurones in an adult longitudinal spinal cord preparation.Brain Res. 673:101–111.
Bleazard, L., R.G. Hill, and R. Morris (1994) The correlation between thedistribution of the NK1 receptor and the actions of tachykinin agonistsin the dorsal horn of the rat indicates that substance P does not have afunctional role on substantia gelatinosa (lamina II) neurons. J. Neuro-sci. 14:7655–7664.
Bowie, D., P. Feltz, and R. Schlichter (1994) Subpopulations of neonatal ratsensory neurons express functional neurotransmitter receptors whichelevate intracellular calcium. Neuroscience 58:141–149.
Cumberbatch, M.J., B.A. Chizh, and P.M. Headley (1995) Modulation ofexcitatory amino acid responses by tachykinins and selective tachyki-nin receptor agonists in the rat spinal cord. Br. J. Pharmacol. 115:1005–1012.
Couture, R., S. Boucher, P. Picard, and D. Regoli (1993) Receptor character-ization of the spinal action of neurokinins on nociception: a threereceptor hypothesis. Regul. Peptides 46:426–429.
DeCamilli, P., P.E. Miller, F. Navone, W.E. Theurkauf, and R.B. Vallee(1984) Distribution of microtubule-associated protein 2 in the nervoussystem of the rat studied by immunofluorescence. Neuroscience 11:819–846.
Ding, Y.-Q., R. Shigemoto, M. Takada, H. Ohishi, S. Nakanishi, and N.Mizuno (1996) Localization of the neuromedin K receptor (NK3) in thecentral nervous system of the rat. J. Comp. Neurol. 364:290–310.
Dun, N.J., S.L. Dun, U. Forstermann, and L.F. Tseng (1992) Nitric oxidesynthase immunoreactivity in rat spinal cord. Neurosci. Lett. 147:217–220.
Fisher, N.D., G. Baranauskas, and A. Nistri (1994) Multiple types oftachykinin receptor mediate a slow excitation of rat spinal motoneu-rones in vitro. Neurosci. Lett. 165:84–88.
Gibson, S.J., J.M. Polak, S.R. Bloom, and P.D. Wall (1981) The distributionof nine peptides in the rat spinal cord with special emphasis on thesubstantia gelatinosa and the area around the central canal (lamina X).J. Comp. Neurol. 201:65–79.
Giesler, G.J., J.T. Cannon, G. Urca, and J.C. Liebeskind (1978) Longascending projections from substantia gelatinosa Rolandi and thesubjacent dorsal horn in the rat. Science 202:984–986.
Grady, E.F., P. Baluk, S. Bohm, P.D. Gamp, H. Wong, D.G. Payan, J. Ansel,A.L. Portbury, J.B. Furness, D.M. McDonald, and N.W. Bunnett (1996)Characterization of antisera specific to NK1, NK2 and NK3 neurokininreceptors and their utilization to localize receptors in the rat gastroin-testinal tract. J. Neurosci. 16:6975–6986.
Grkovic, I., and C.R. Anderson (1996) Distribution of immunoreactivity forthe NK1-receptor on different subpopulations of sympathetic pregangli-onic neurons in the rat. J. Comp. Neurol. 37:1–11.
Haley, J.E., A.H. Dickenson, and M. Schachter (1992) Electrophysiologicalevidence for a role of nitric oxide in prolonged chemical nociception inthe rat. Neuropharmacol. 31:251–258.
Hao, J.X., X.J. Xu, Y.X. Yu, A. Seiger, and Z. Wiesenfeld-Hallin (1992)Baclofen reverses the hypersensitivity of dorsal horn wide dynamicrange neurons to mechanical stimulation after transient spinal cordischemia: Implications for a tonic GABAergic inhibitory control ofmyelinated fiber input. J. Neurophysiol. 68:392–396.
Hosoya, Y., I. Nadelhaft, D. Wang, and K. Kohno (1994) Thoracolumbarsympathetic preganglionic neurons in the dorsal commissural nucleusof the male rat: An immunohistochemical study using retrogradelabeling of cholera toxin subunit B. Exp. Brain Res. 98:21–30.
Inoue, K., K. Nakazawa, K. Inoue, and K. Fujimori (1995) Nonselectivecation channels coupled with tachykinin receptors in rat sensoryneurons. J. Neurophysiol. 73:736–742.
Ireland, S.J., I.K. Wright, and C.C. Jordan (1992) Characterization oftachykinin-induced ventral root depolarization in the neonatal ratisolated spinal cord. Neuroscience 46:217–223.
Kar, S., and R. Quirion (1995) Neuropeptide receptors in developing andadult rat spinal cord: An in vitro quantitative autoradiography study ofcalcitonin gene-related peptide, neurokinins, µ-opioid, galanin, somato-statin, neurotensin and vasoactive intestinal polypeptide receptors. J.Comp. Neurol. 354:253–281.
NK3 RECEPTORS IN RAT SPINAL CORD 447

Laneuville, O., J. Dorais, and R. Couture (1988) Characterization of theeffects produced by neurokinins and three agonists selective for neuro-kinin receptor subtypes in spinal nociceptive reflex in the rat. Life Sci.42:1295–1305.
Light, A.R., D.L. Trevino, and E.R. Perl (1979) Morphological features offunctionally defined neurons in the marginal zone and substantiagelatinosa of the spinal dorsal horn. J. Comp. Neurol. 186:151–172.
McCarson, K.E., and J.E. Krause (1994) NK-1 and NK-3 type tachykininreceptor mRNA expression in the rat spinal cord dorsal horn isincreased during adjuvant or formalin-induced nociception. J. Neuro-sci. 14:712–720.
Meller, S.T., C. Dykstra, and G.F. Gebhart (1992) Production of endogenousnitric oxide and activation of soluble guanylate cyclase are required forN-methyl-D-aspartate-produced facilitation of the nociceptive tail-flickreflex. Eur. J. Pharmacol. 214:93–96.
Menetrey, D., A. Chaouch, D. Binder, and J.M. Besson (1982) The origin ofthe spinomesencephalic tract in the rat: An anatomical study using theretrograde transport of horseradish peroxidase. J. Comp. Neurol.206:193–207.
Morris, J.L., I.L. Gibbins, G. Campbell, R. Murphy, J.B. Furness, and M.Costa (1986) Innervation of the large arteries and heart of the toad(Bufo Marinus) by adrenergic and peptide-containing neurons. CellTiss. Res. 243:171–184.
Ogawa, T., I. Kanazawa, and S. Kimura (1985) Regional distribution ofsubstance P, neurokinin a and neurokinin ¬ in rat spinal cord, nerveroots and dorsal root ganglia, and the effects of dorsal root section orspinal transection. Brain Res. 359:152–157.
Oury-Donat, F., P. Carayon, O. Thurneyssen, V. Pailhon, X. Edmonds-Alt, P.Soubrie, and G. LeFur (1995) Functional characterization of thenonpeptide neurokinin3 (NK3) receptor antagonist, SR142801 on thehuman NK3 receptor expressed in Chinese hamster ovary cells. J.Pharmacol. Exp. Ther. 274:148–154.
Papir-Kricheli, D., J. Frey, R. Laufer, C. Gilon, M. Chorev, Z. Selinger, andM. Devor (1987) Behavioural effects of receptor-specific substance Pagonists. Pain 31:263–276.
Rose, P.K., and F.J.R. Richmond (1981) White-matter dendrites in the
upper cervical spinal cord of the adult cat: A light and electronmicroscopic study. J. Comp. Neurol. 199:191–203.
Scheibel, M.F., and A.B. Scheibel (1968) Terminal axonal patterns in catspinal cord. II. The dorsal horn. Brain Res. 9:32–58.
Sivilotti, L., and C.J. Woolf (1994) The contribution of GABAA and glycinereceptors to central sensitization: Disinhibition and touch-evoked allo-dynia in the spinal cord. J. Neurophysiol. 72:169–179.
Spike, R.C., A.J. Todd, and H.M. Johnston (1993) Coexistence of NADPHdiaphorase with GABA, glycine and acetylcholine in rat spinal cord. J.Comp. Neurol. 335:320–333.
Stefanini, M., C. deMartino, and L. Zamboni (1967) Fixation of ejaculatedspermatozoa for electron microscopy. Nature 215:173–174.
Strack, A.M., W.B. Sawyer, L.M. Marubio, and A.D. Loewy (1988) Spinalorigin of sympathetic preganglionic neurons in the rat. Brain Res.455:187–191.
Suzuki, H., K. Yoshioka, T. Maehara, R.M. Hagan, S. Nakanishi, and M.Otsuka (1993) Pharmacological characteristics of trachykinin receptorsmediating acetylcholine release from neonatal rat spinal cord. Eur. J.Pharmacol. 241:105–110.
Warden, M.K., and W.S. Young III (1988) Distribution of cells containingmRNAs encoding substance P and neurokinin B in the rat centralnervous system. J. Comp. Neurol. 272:90–113.
Willis, W.D., D.L. Trevino, J.D. Coulter, and R.A. Maunz (1974) Responsesof primate spinothalamic tract neurons to natural stimulation ofhindlimb. J. Neurophysiol. 37:358–372.
Yaksh, T.L. (1989) Behavioral and autonomic correlates of the tactileevoked allodynia produced by spinal glycine inhibition: Effects ofmodulatory receptor systems and excitatory amino acid antagonist.Pain 37:111–123.
Yashpal, K., T.-V. Dam, and R. Quirion (1990) Quantitative autoradio-graphic distribution of multiple neurokinin binding sites in rat spinalcord. Brain Res. 506:259–266.
Zhuo, H., and C.J. Helke (1993) Neurokinin B peptide-2 neurons projectfrom the hypothalamus to the thoracolumbar spinal cord of the rat.Neuroscience 52:1019–1028.
448 V.S. SEYBOLD ET AL.