universidade estadual de santa cruz verÔnica...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DE SANTA CRUZ
VERÔNICA CORDEIRO SILVA
INFLUÊNCIA DE FUNGOS MICORRÍZICOS ARBUSCULARES NO CRESCIMENTO E PRODUÇÃO DE ÓLEO ESSENCIAL EM MENTHA X
PIPERITA L. VAR. CITRATA (EHRH.) BRIQ.
ILHÉUS – BAHIA 2012

ii
VERÔNICA CORDEIRO SILVA
INFLUÊNCIA DE FUNGOS MICORRÍZICOS ARBUSCULARES NO CRESCIMENTO E PRODUÇÃO DE ÓLEO ESSENCIAL EM MENTHA X
PIPERITA L. VAR. CITRATA (EHRH.) BRIQ.
Dissertação apresentada para obtenção do titulo de Mestre em Produção Vegetal pela Universidade Estadual de Santa Cruz. Orientador: Prof. Dr. Eduardo Gross. Co-orientadora: Profa Dra Larissa Correa do Bomfim Costa.
Ilhéus – Bahia 2012

iii

iv
À minha mãe, o maior e melhor exemplo que tenho na vida. Dedico.

v
AGRADECIMENTOS
A Deus, por me fazer resistente diante das intempéries.
A minha mãe pelas lições de sabedoria e persistência.
Aos meus irmãos pelo encorajamento, até mesmo no silêncio.
Ao meu orientador Eduardo Gross pela paciência e valiosos ensinamentos de
humildade e profissionalismo.
A minha Co-orientadora Larissa Correa do Bomfim Costa pela confiança e
seriedade.
Aos meus colegas e amigos do Laboratório de Química e Fertilidade do Solo:
Mary, Lidi, Leo, Carol, Caique, Nairane e Gedeon pelos momentos de
descontração e em especial à minha grande parceira e conselheira Paty.
As minhas amigas de cunho acadêmico e pessoal Analine e Lívia pelos
desabafos.
Aos demais amigos e familiares.
Aos professores Rosilene e Raildo pelos valiosos auxílios.
Aos amigos do campo Roberto, Adelino e Marcelo pela simplicidade.
A Universidade Estadual de Santa Cruz e a CAPES pelo auxílio financeiro e
material.
Agradeço.

vi
“Plantas não tem raízes, elas tem micorrizas”
J. L. Harley

vii
LISTA DE FIGURAS
Revisão de literatura
Figura 1. Mentha x piperita L. var. citrata (Ehrh.) Briq. 4
1. Capítulo I
Figura 1. Imagens de raízes coradas pelo método de Phillips e
Haymann mostrando a morfologia das micorrizas encontradas
em Mentha x piperita var. citrata.
47

viii
LISTA DE TABELAS
1. Capítulo I
Tabela 1. Características químicas iniciais (antes do experimento) do
Argissolo Amarelo Distrófico típico coletado na Estação
Experimental da Almada, localizada no município de
Uruçuca, BA
27
Tabela 2. Características físicas iniciais (antes do experimento) do
Argissolo Amarelo Distrófico típico coletado na Estação
Experimental do Almada, localizada no município de
Uruçuca, BA
27
Tabela 3. Descrição dos contrastes ortogonais para a avaliação da
influencia de FMAs em Mentha x piperita var. citrata
submetida a duas doses de fósforo
31
Tabela 4. Resumo da análise de variância para as varáveis biomassa
de raiz, caule, folha e total de Mentha x piperita. var. citrata
inoculada com fungos micorrízicos arbusculares (FMA) e
submetida a doses de fósforo
32
Tabela 5. Valores médios em g referente à biomassa seca da raiz,
caule, folha e total de Mentha x piperita var. citrata
inoculada com FMA submetida a duas doses de fósforo
32
Tabela 6. Estimativas dos contrastes ortogonais das varáveis
biomassa de raiz, caule, folha e total para Mentha x piperita
var. citrata inoculada com fungos micorrízicos arbusculares
e submetida a duas doses de fósforo
33

ix
Tabela 7. Resumo da análise de variância para o teor dos macro e
micronutrientes das folhas de Mentha x piperita var. citrata
inoculada com fungos micorrízicos arbusculares e
submetida a duas doses de fósforo
37
Tabela 8. Valores médios do teor de macro (em g kg-1) e de
micronutrientes (em mg kg-1) das folhas de Mentha x
piperita var. citrata inoculada com fungos micorrízicos
arbusculares e submetida a duas doses de fósforo
37
Tabela 9. Estimativas dos contrastes ortogonais para o teor de macro
e micronutrientes nas folhas de Mentha x piperita var.
citrata inoculada com fungos micorrízicos arbusculares e
submetida a duas doses de fósforo
40
Tabela 10. Características químicas finais (depois do experimento) do
Argissolo Amarelo Distrófico típico solo coletado na
Estação Experimental da Almada, localizada no município
de Uruçuca, BA
42
Tabela 11. Resumo da Análise de Variância para o número de esporos
e porcentagem de colonização micorrízica em Mentha x
piperita var. citrata submetida a duas doses de fósforo
43
Tabela 12. Número de esporos no solo e porcentagem de colonização
micorrízica total e por estruturas fúngicas em Mentha x
piperita var. citrata inoculada com fungos micorrízicos
arbusculares e submetida a duas doses de fósforo
44

x
Tabela 13. Estimativas dos contrastes ortogonais para o número de
esporos e para a porcentagem de colonização micorrízica
Mentha x piperita var. citrata inoculada com fungos
micorrízicos arbusculares e submetida a duas doses de
fósforo
45
2. Capítulo II
Tabela 1. Características químicas iniciais (antes do experimento) do
Argissolo Amarelo Distrófico típico coletado na Estação
Experimental da Almada, localizada no município de
Uruçuca, BA
57
Tabela 2. Características físicas iniciais (antes do experimento) do
Argissolo Amarelo Distrófico típico coletado na Estação
Experimental do Almada, localizada no município de
Uruçuca, BA
57
Tabela 3. Descrição dos contrastes ortogonais para a avaliação da
influencia de FMAs em Mentha x piperita var. citrata
submetida a duas doses de fósforo
59
Tabela 4. Resumo da Análise de Variância para as variáveis
biomassa, teor e rendimento de óleo de Mentha x piperita
var. citrata inoculada com fungos micorrízicos arbusculares e
submetida a duas doses de fósforo
60

xi
Tabela 5. Valores médios da massa seca das folhas (em g), do teor
(%) e do rendimento (kg planta-1) do óleo essencial extraído
das folhas de Mentha x piperita var. citrata por
hidrodestilação
60
Tabela 6. Estimativas dos contrastes ortogonais para teor e
rendimento do óleo essencial de Mentha x piperita var.
citrata inoculada com fungos micorrízicos arbusculares e
submetida a duas doses de fósforo
62
Tabela 7. Índice de Kovats (IK) e concentração relativa em
porcentagem dos constituintes químicos do óleo essencial
da biomassa seca de folhas de Mentha x piperita var. citrata
inoculada com FMA submetida a duas doses de fósforo
65

xii
RESUMO
Os fungos micorrízicos arbusculares em solos de baixa fertilidade natural, geralmente, promovem aumento no crescimento beneficiando a produção de massa seca nas plantas hospedeiras, sendo que em plantas produtoras de óleo essencial podem ainda alterar o rendimento e a composição deste. A depender do tipo de fungo micorrízico e da espécie vegetal em questão os efeitos podem ser diferenciados e, portanto, de tal maneira o micobionte e o fotobionte deve ser testado. O estudo teve como objetivo avaliar os efeitos dos fungos micorrízicos arbusculares (FMAs), Acaulospora morrowiae Spain & Schenck, Glomus clarum Nicol. & Schenck e Scutellospora calospora (Nicol. & Gerd.) Walk. & Sand., associados com dose de P no crescimento de Mentha x piperita L. var. citrata (Ehrh.) Briq., e na produção e qualidade do óleo essencial. O experimento foi conduzido em casa de vegetação num fatorial 5x2 com cinco tratamentos micorrízicos (controle não inoculado, A. morrowiae, G. clarum, e S. calospora, e a mistura desses inóculos) e duas doses de P (60 mg e 120 mg de fósforo por dm3 de solo) com dez repetições cada, subdivida em duas partes, metade foi destinada à avaliação do crescimento e outra metade para produtividade do óleo essencial. As plantas foram colhidas após 75 dias para a avaliação da biomassa seca, porcentagem de colonização micorrízica das raízes, número de esporos no solo, conteúdo dos nutrientes nas folhas, teor, rendimento e composição do óleo essencial. Houve interação entre os fungos micorrízicos e as doses de fósforo aplicadas que influenciaram na absorção dos nutrientes e no crescimento (biomassa) das plantas. Para a colonização micorrízica G. clarum apresentou uma maior porcentagem de colonização das raízes na maior dose de P, enquanto S. calospora teve maior percentual de colonização na menor dose de P. O número de esporos presente em 50 g de solo foi significativamente maior para os tratamentos inoculados com os FMAs em comparação com o controle não inoculado, entretanto não houve influência das doses de P na quantidade de esporos encontrada. A inoculação com FMAs aumentou o teor e o rendimento de óleo essencial da planta na menor dose de fósforo. Os inóculos utilizados separadamente foram mais eficientes do que a mistura dos fungos no incremento do teor e do rendimento do óleo essencial na dose maior de fósforo sendo que G. clarum elevou significativamente o rendimento do óleo quando a dose de 120 mg dm-3 de P foi aplicada ao solo. A colonização com FMAs não alterou a composição do óleo essencial, que teve como componentes majoritários o linalol e o acetato de linalol, mas houve alterações quantitativas (percentuais) dos componentes do óleo essencial de Mentha x piperita var. citrata.
Palavras-chave: Acaulospora, Glomus, Scutellospora, Linalol, Acetato de Linalol, Adubação fosfatada.

xiii
ABSTRACT
Arbuscular mycorrhizal fungi in soils of low fertility usually increase growth and production of dry matter on host plants and on essential oil producing plants can also alter the yield and composition of oil. Depending on fungus species and plant species effects can be distinguished and thus both mycobiont as photobiont should be tested. Our study aimed to evaluate the effects of mycorrhizal fungi (AMF), Acaulospora morrowiae Spain & Schenck, Glomus clarum Nicol. & Schenck e Scutellospora calospora (Nicol. & Gerd.) Walk. & Sand., on growth of Mentha x piperita L. var. citrata (Ehrh.) Briq., and the production and quality of essential oil. The experiment was conducted in a greenhouse in a 5x2 factorial design with five mycorrhizal treatments (uninoculated control, A. morrowiae, G. clarum, and S. calospora, and the mixture of inocula) and two P levels (60 mg and 120 mg phosphorus per dm3 of soil) with ten replicates each, subdivided on two parts, half to evaluation of plant growth and half to evaluation of essential oil productivity. Plants were harvested after 75 days for evaluation of dry biomass, the percentage of mycorrhizal colonization of roots, the number of spores in soil, content of nutrients in the leaves, content, yield and essential oil composition. There was interaction between mycorrhizal fungi and phosphorus doses that influence the absorption of nutrients and growth (biomass) of plants. For the colonization G. clarum showed a higher percentage of root colonization at the highest dose of P, and S. calospora had a higher percentage of colonization in the lower dose of P. The number of spores present in 50 g of soil was significantly higher for the treatments inoculated with mycorrhizal fungi compared to uninoculated control, however there was no influence of P levels on spore number. Inoculation with AMF increased the content and oil yield at the lowest dose of phosphorus. Inoculum used alone were more effective than the mixture of fungi in increasing the content and yield of essential oil in the higher dose of phosphorus, and G. clarum significantly increased oil yield when 120 mg dm-3 of P was applied to the soil. The mycorrhizal colonization did not alter the essential oil composition, which had as its major components linalool and linalool acetate, but there were quantitative changes (percentages) of essential oil components of Mentha x piperita.
Palavras-chave: Acaulospora, Glomus, Scutellospora, Linalool, Linalool Acetate, Phosphorus fertilization.

xiv
SUMÁRIO
RESUMO xii
ABSTRACT. xiii
INTRODUÇÃO 01
REVISÃO DE LITERATURA 03
1. Capítulo I 22
1.1 Introdução 24
1.2 Material e Métodos 26
1.2.1 Determinação da biomassa 29
1.2.2 Teor de nutrientes 29
1.2.3 Porcentagem de colonização micorrízica 29
1.2.4 Número de esporos de FMAs 30
1.2.5 Análise Estatística 30
1.3 Resultados e Discussão 31
1.3.1 Biomassa 31
1.3.2 Teor dos nutrientes 36
1.3.3 Porcentagem de colonização micorrízica e número de esporos 43
1.4 Conclusões 48
1.5 Referências 48
2. Capítulo II 52
2.1 Introdução 54
2.2 Material e Métodos 55
2.2.1 Determinação da biomassa 58
2.2.2 Determinação do teor, rendimento e composição química do óleo essencial 58
2.2.3 Análise estatística 59
2.3 Resultados e Discussão 60
2.4 Conclusões 66
2.5 Referências 66
Considerações Finais 69

1
INTRODUÇÃO
Os organismos vegetais fornecem à humanidade os mais variados
subsídios para sua permanência e manutenção na terra, seja provendo matéria
prima para indústrias como a de cosméticos, construção civil, e vestuário ou
sendo diretamente consumidas na alimentação. A medicina enquadra-se como
outro segmento em que as plantas são amplamente utilizadas, necessitando
assim de um respaldo científico para corroborar as informações acerca dos
seus benefícios medicinais que na maioria das vezes são transmitidos
culturalmente.
O metabolismo secundário das plantas fornece uma gama de compostos
que são utilizados na confecção de produtos diversos, nas mais variadas
indústrias. Dentre esses compostos os óleos essenciais estão cada vez mais
valorizados devido as suas propriedades aromáticas e terapêuticas que são
conferidas por características físico-químicas tais como volatilidade e
solubilidade em solventes orgânicos apolares.
Dentre as plantas produtoras de óleo essencial destaca-se a família
Lamiacea, com espécies que geralmente sintetizam e armazenam seu
composto volátil em estruturas denominadas tricomas glandulares. Nesta
família, um dos gêneros que desperta amplo interesse é o Mentha. As espécies
pertencentes a esse gênero produzem óleo essencial rico em mentol, mentona,
mentofurano e linalol, que são considerados os principais responsáveis pelo
uso dessas espécies como condimentos e aromatizantes e pelas ações
terapêuticas nos sistemas respiratório e gastrointestinal.
Fatores genéticos, fisiológicos e condições edafoclimáticas podem influir
diretamente sobre a biossíntese, qualidade e quantidade do óleo essencial.
Dentre esses fatores a presença de microrganismos benéficos associados à
raiz pode gerar alterações no biocomposto.
Micorriza é uma associação simbiotrófica mutualista obrigatória entre
plantas e fungos, onde tanto a planta hospedeira quanto o fungo simbionte são
beneficiados. Nesta relação, o fitobionte transloca para o fungo fotoassimilados
que auxiliam na manutenção e desenvolvimento de novas formas de
propágulos infectivos e/ou estruturas morfológicas fúngicas, enquanto o

2
micobionte incrementa a produtividade do vegetal através da melhoria do
estado nutricional da planta, ampliando (com suas hifas extrarradiculares) a
superfície de contato da mesma com o solo e otimizando a absorção de
nutrientes principalmente daqueles considerados pouco móveis. Além disso, a
micorrização pode proporcionar para o vegetal tolerância ao estresse hídrico e
ao ataque de patógeno, sem mencionar a melhoria para a ecologia do solo.
Essa associação mutualista trata-se de uma co-evolução gene a gene
estabelecida por meio de sinais bioquímicos, moleculares e fisiológicos que
permitiu não somente o estabelecimento dos organismos vegetais no solo, mas
também o desenvolvimento e adaptação nos mais diversos biomas terrestres.
Diante do acima exposto que elucida a importância do estudo da
associação entre fungos micorrízicos arbusculares (FMA) e plantas medicinais,
o objetivo do trabalho foi o de avaliar o efeito desses fungos no crescimento,
composição química e produção de óleo essencial em Mentha x piperita L. var.
citrata (Ehrh.) Briq.

3
REVISÃO DE LITERATURA
Mentha x piperita L. var. citrata (Ehrh.) Briq.
A família Lamiaceae é uma das mais diversificadas e apresentam
distribuição cosmopolita. Nessa família estão incluídos cerca de 300 gêneros e
7500 espécies (SOUZA; LORENZI, 2005), algumas com grande importância
econômica devido a produção de óleos essenciais (COSTA, 2008).
Um dos principais gêneros dessa família é o Mentha. Comumente são
espécies herbáceas e perenes com altura variável entre 20 e 70 cm de acordo
com o material genético e condições de manejo. Apresentam folhas opostas e
ovaladas com margens serrilhadas, flores hermafroditas e caules subterrâneos
(rizomas). As espécies mais cultivadas mundialmente são Mentha arvensis L. e
Mentha x piperita L. M. arvensis é originária de clima temperado e é cultivada
principalmente no Paraná e em São Paulo. M. x piperita é originária do
cruzamento entre M. aquática e M. viridis, sendo que normalmente apresenta
desenvolvimento rasteiro e grande produção de rizomas (BIASI; DESCHAMPS,
2009).
O uso de plantas do gênero Mentha é amplamente difundido, tendo em
vista sua adaptabilidade a diferentes condições edafo-climáticas, ao ciclo
vegetativo anual e a quantidade de informações já existentes sobre suas
características, desde anatômicas, bioquímicas, taxonômicas e agronômica
(MONTEIRO, 2009).
M. arvensis L. é conhecida popularmente como hortelã-japonesa, vique
ou hortelã, tem como principal componente de seu óleo o mentol, é muito
utilizada como matéria prima em indústrias farmacêutica e de confeitaria, seu
óleo pode conferir sabor e odor a remédios e balas, na indústria de cosméticos
serve para atribuir sensação refrescante a loções e cremes. De acordo com
informações etnobotânicas possui propriedades antidispéptica, antivomitiva,
descongestionante nasal e antigripal, age contra dor de cabeça e coceira na
pele (LORENZI; MATOS, 2002).
A espécie M. x piperita L. apresenta propriedades espamolíticas,
antivomitivas, carminativas, estomáquicas e anti-helmínticas, por via oral,

4
antibacterianas, antifúngicas e antiprurido em uso tópico (LORENZI; MATOS,
2002). A composição do óleo essencial de M. x piperita varia em função da
variedade, do quimiotipo e dos fatores abióticos, não existindo uma definição
exata de sua composição (OLIVEIRA, 2011a). Outras espécies de destaque
são M. pulegium, conhecida como poejo, registra ações contra desordens
digestivas, amenorreia, gota, resfriados e pode aumentar micção e quando em
excesso tem ação abortiva e hepatotóxica e M. x villosa mais utilizada como
condimento de carnes e massas, além de ações espasmolítica, antivomitiva,
carminativa, estomática, anti-helmíntica, antisséptica e antiprurido registrado
pela literatura etnobotânica (LORENZI; MATOS, 2002).
M. x piperita L. var. citrata (Ehrh.) Briq. (Figura 1) é conhecida como
hortelã-limão, possui óleo essencial rico em linalol e acetato de linalila, em
contraste com outras espécies do gênero Mentha (MURRAY; LINCOLN, 1970).
Portanto a necessidade de estudos fitotécnicos sobre a espécie aumenta para
otimizar a produção dos compostos de interesse para as indústrias.
Figura 1. Mentha x piperita L. var. citrata (Ehrh.) Briq.

5
Óleo essencial
Metabólitos secundários são compostos orgânicos produzidos pelos
vegetais que parecem não ter função direta no seu crescimento e
desenvolvimento. Muitas vezes são distribuídos de maneira diferente entre
limitados grupos taxonômico dentro do reino vegetal, ou seja, metabólitos
secundários específicos são restritos a uma espécie vegetal ou a um grupo de
espécies relacionadas, sendo assim, esses compostos podem ser utilizados
em estudos taxonômicos (quimiossistemática). Apesar do metabolismo
secundário nem sempre ser necessário para que uma planta complete seu
ciclo de vida, ele desempenha um papel importante na interação das plantas
com o meio ambiente, apresentando funções ecológicas importantes tais como:
proteção contra herbivoria e infecção por organismos patogênicos, atrativos
para animais polinizadores e dispersores de sementes, bem como agentes na
competição planta-planta (PERES, 2004; TAIZ; ZEIGUER, 2004; CROTEAU et
al., 2000).
Os metabólitos secundários podem ser dividos em três grupos principais:
compostos fenólicos, alcalóides e terpenos, sendo, os terpenos os que ocorrem
com maior frequência e abundância (BIASI; DESCHAMPS, 2009; TAIZ;
ZEIGUER, 2004; SANGWAN et al., 2001).
Os compostos fenólicos são produtos secundários que possuem um
grupo fenol, um grupo hidroxila funcional e um anel aromático, são derivados
do ácido chiquímico ou ácido mevalônico. Constituem um grupo quimicamente
heterogêneo, alguns são solúveis apenas em solventes orgânicos, outros são
ácidos carboxílicos e glicosídeos solúveis em água e há, ainda, aqueles que
são grandes polímeros insolúveis. Devido a diversidade química, os compostos
fenólicos apresentam uma variedade de funções, muitos agem como composto
de defesa contra herbívoros e patógenos, outros tem função de suporte
mecânico, como atrativo de polinizadores e dispersores, na proteção da
radiação ultravioleta e ação alelopática (TAIZ; ZEIGUER, 2004).
Os compostos fenólicos são bastante utilizados cotidianamente
conferindo sabor, odor e coloração a diversos produtos, alguns desses
compostos, como o aldeído cinâmico da canela (Cinnamomum zeyllanicum) e a

6
vanilina da baunilha (Vanilla planifolia), são empregados na indústria de
alimentos (PERES, 2004).
Os alcalóides são metabólitos secundários que possuem nitrogênio na
sua estrutura, considerados de interesse devido ao efeito tóxico para humanos
e sua propriedades medicinais. Eles são derivados de aminoácidos aromáticos
(triptofano, tirosina), os quais são derivados do ácido chiquímico, e também de
aminoácidos alifáticos (ornitina, lisina) (PERES, 2004; TAIZ; ZEIGER, 2004).
Os terpenóides são originados a partir do ácido mevalônico (no
citoplasma) ou do piruvato e 3-fosfoglicerato (no cloroplasto) (PERES, 2004;
TAIZ; ZEIGER, 2004). Eles são sintetizados a partir de cinco unidades de
carbono os isoprenos, dessa união formam-se os demais: monoterpenos (10
carbonos), sesquiterpenos (15 carbonos), diterpenos (20 carbonos) e
triterpenos (30 carbonos) (BIASI; DESCHAMPS, 2009).
Os monoterpenos possuem maior diversidade química e devido ao seu
baixo peso molecular, costumam ser substâncias voláteis, sendo, portanto
denominados óleos essenciais ou essências. Contudo os óleos essenciais
podem ser derivados de qualquer uma das quatro rotas precursoras dos
metabólitos secundários: do ácido chiquímico, do mevalonato, do acetato e dos
aminoácidos, havendo ainda compostos de origem mista, ou seja, provenientes
de mais de uma rotas (CASTRO et al., 2004).
Óleos essenciais são frações líquidas e voláteis que contem as
substâncias responsáveis pelo aroma das plantas, também podem ser
chamados de óleos voláteis, óleos etéreos ou essências, essas denominações
derivam de algumas características físico-químicas tais como: aparência oleosa
à temperatura ambiente; volatilidade; aroma agradável e intenso; solubilidade
em solventes orgânicos apolares; incolores ou amarelados; instáveis em
presença de luz, ar calor umidade e metais (CASTRO et al., 2004).
A biossíntese dos monoterpenos pode ocorrer em células
parenquimáticas diferenciadas (Lauraceae, Piperaceae, Poaceae); canais
oleíferos (Apiaceae) ou em bolsas lisígenas ou esquizolisígenas (Pinaceae,
Rutaceae). Podem estar estocadas em flores (laranjeira), folhas (capim-limão,
eucalipto, louro) ou nas cascas dos caules (canelas), madeiras (sândalo, pau-
rosa) e frutos (erva-doce) (PERES, 2004). Nas mentas, assim como nas

7
demais lamiáceas os monoterpenos são produzidos e acumulados em
estruturas secretoras especializadas conhecidas como tricomas glandulares
peltados, localizados na parte aérea da planta, principalmente nas folhas e
cálices florais (TURNER et al., 2000).
Diferentes métodos para a extração de óleos essenciais podem ser
usados de acordo principalmente com o valor comercial do produto e órgão da
planta aonde se concentra o óleo. Os métodos mais utilizados incluem a
hidrodestilação e arraste a vapor (BIASI; DESCHAMPS, 2009).
Do ponto de vista biológico a função dos óleos essenciais nas plantas
pode ser tanto para atrair polinizadores (principalmente os noturnos) quanto
para repelir insetos (pragas) (PERES, 2004). No entanto plantas produtoras de
óleo essencial adquiriram importância econômica e tiveram uma ascensão em
seu cultivo, pois são usadas como medicamentos fitoterápicos, alguns
fabricados por conceituados laboratórios e suprem as indústrias de cosméticos,
perfumarias e alimentícias, no preparo dos mais variados produtos
(RUSSOMANO, 2006).
Apesar do alto valor econômico dos óleos das mentas, o papel biológico
ainda é pouco conhecido acredita-se que seus constituintes atuam na defesa
da planta contra perda de água, efeitos alelopáticos, ataques biótico e de
herbívoros (KAROUSOU et al., 1998).
Fatores que afetam a produção de óleo
A produção do óleo essencial em plantas aromáticas é regulada por
diversos fatores, que podem ser genéticos, bioquímicos, fisiológicos e de
origem metabólica. Os processos bioquímicos e metabólicos incluem
mecanismos moleculares regulados por rotas biossintéticas de carbono,
terpenóides e fenilpropanóides, e a regulação fisiológica além de ser exercida
por um padrão de desenvolvimento específico é influenciado por fatores
ambientais (SANGWAN et al., 2001).
Dentre estes fatores, podem-se ressaltar as interações
planta/microrganismos, planta/insetos e planta/planta; idade e estádio de
desenvolvimento, fatores abióticos como luminosidade, temperatura,

8
pluviosidade, nutrição, época e horário de coleta, bem como técnicas de
colheita e pós–colheita (MORAIS, 2009). Alguns desses fatores não atuam
isoladamente, podendo influir em conjunto no metabolismo secundário, como
por exemplo: desenvolvimento e sazonalidade; temperatura e altitude, entre
outros (GOBBO-NETO; LOPES, 2007).
Segundo Morais (2009) a alteração dos compostos majoritários nos
óleos essenciais, seja por fatores genéticos, técnicos (coleta, estabilização e
armazenamento), bióticos ou abióticos, pode influenciar diretamente na
qualidade e, consequentemente, nos resultados de tratamentos e de testes
biológicos sobre patógenos humanos ou fitopatógenos.
Fungos Micorrízicos Arbusculares
Os fungos micorrízicos arbusculares são simbiotróficos mutualistas
obrigatórios, isto é, só se desenvolvem no interior de células radiculares em
hospedeiros vivos, sem prejudicá-los, muito pelo contrário, auxiliando-os.
Raízes fossilizadas evidenciam o surgimento de micorrizas há cerca de 400
milhões de anos, acredita-se que durante a evolução conjunta fungos e plantas
desenvolveram a capacidade de se comunicarem molecularmente, através de
mecanismos de reconhecimento, tropismo e tactismo que permitiram a
interação célula a célula e a integração morfológica e funcional (MOREIRA;
SIQUEIRA, 2006).
Atualmente os fungos micorrízicos estão incluídos em um filo próprio o
Glomeromycota, essa organização foi baseada na convergência filogenética de
características morfológicas, bioquímicas e moleculares (MOREIRA;
SIQUEIRA, 2006), essa classificação é a mais reconhecida e aceita. Outras
classificações também são adotadas, como a do INVAM que apresenta uma
lista completa das espécies de FMAs e divide o filo em uma ordem, duas
subordens, cinco famílias e sete gêneros (MORTON; BENNY, 1990; MORTON;
REDECKER, 2001). Outra classificação é a proposta pela Universidade
Federal de Lavras que subdivide o filo em quatro ordens, nove famílias e doze
gêneros (SCHüBLER et al. 2001; http://www.lrz.de/~schuessler/amphylo/).

9
A colonização tem início com a formação de uma hifa infectiva, a partir
de um esporo germinado, segmento de raiz infectado ou de hifas no solo. As
hifas crescem na rizosfera e, ao entrar em contato com as raízes, formam uma
estrutura de penetração do tipo apressório que por meio de mecanismos
mecânicos e enzimáticos degrada a parede celular. Após penetração, o fungo
forma hifas que de uma maneira geral são asseptadas e que podem ser
classificadas em inter ou intracelulares e intra ou extrarradicular. No apoplasto
as hifas se diferenciam formando arbúsculos para ter acesso ao suprimento de
carbono da planta, em algumas espécies pode haver desenvolvimento de
vesículas que aparentemente possuem função de reserva. O estádio final da
simbiose culmina com a formação de novos esporos que acontece na maioria
das vezes na rizosfera, mas também pode ocorrer no interior das raízes
(MOREIRA; SIQUEIRA, 2006).
Efeito dos fungos micorrízicos arbusculares no crescimento das
plantas e na absorção de nutrientes
A micorrização é considerada como sendo mutualista nutricional, onde o
fotobionte supre o fungo com fotoassimilados para crescimento e reprodução,
enquanto que o micobionte provê a planta nutrientes e água. Assim, esta
simbiose amplia a capacidade de absorção de nutrientes por parte do
simbionte autotrófico e, conseqüentemente, a sua competitividade inter-
específica e produtividade (BERBARA et al., 2006).
Várias espécies de plantas respondem positivamente à inoculação com
FMAs, dentre elas café, soja, milho, batata-doce, mandioca, cana-de-açúcar,
além de várias espécies florestais e frutíferas. Algumas são consideradas
obrigatórias, isto é, não crescem na ausência de FMAs em níveis normais de
disponibilidade de nutrientes. Esta característica é encontrada com frequência
em espécies nativas de solos de baixa fertilidade natural (SIQUEIRA; SAGGIN-
JUNIOR, 2001). Nestes solos, inúmeras espécies vegetais são incapazes de
absorver fósforo na ausência de micorrização, como mandioca e batata-doce
(BERBARA et al., 2006).

10
A simbiose micorrízica arbuscular tem um importante potencial
biotecnológico e ecológico. Ela causa impactos que vão desde suas relações
com plantas (processos de absorção de nutrientes, principalmente dos
considerados pouco móveis no solo), com comunidades vegetais (influenciando
em sua diversidade e abundância) e com processos relacionados à
estabilidade de ecossistemas, ao participarem de forma ativa e significante na
agregação do solo, incrementos à resistência de plantas frente ao ataque
patogênico (HWANG et al., 1992), à tolerância ao estresse hídrico, à eficiência
fotossintética (BROWN; BETHLENFALVAY, 1987), ao intemperismo de
minerais (VAN BREEMEN et al., 2000).
O efeito dos fungos micorrízicos arbusculares sobre o crescimento das
plantas é especialmente significativo com relação aos nutrientes de baixa
mobilidade no solo,os quais praticamente não se movem por fluxo de massa,
porém chegam à raiz por meio de mecanismos de difusão (MARSCHNER,
1995). Neste grupo situam-se primordialmente o macronutriente fósforo e os
micronutrientes zinco e cobre (CARDOSO et al., 2010). Já o nitrogênio quando
se encontra na forma de amônio (menos móvel que o nitrato) é comum
observar-se o aumento de sua absorção em plantas micorrizadas (GEORGE et
al., 1992).
A melhora da nutrição fosfatada das plantas tem sido reconhecida como
um dos maiores benefícios das micorrizas, as respostas, entretanto, variam
com o nível de fósforo, pH e teor de alumínio, com a espécie de FMA presente
e com a cultivar ou espécie de planta. O mecanismo de atuação da micorriza
mais aceito para explicar o benefício em relação ao fósforo, é que as micorrizas
exploram uma área maior de solo, em relação às raízes, portanto um processo
físico (CARDOSO et al., 2010). Todavia as micorrizas podem ser mais do que
simplesmente uma extensão do sistema radicular, contribuindo na absorção de
nutrientes pela planta em até 80% de fósforo, 60% de cobre, 25% de
nitrogênio, 25% de zinco e 10% de K (MARSCHNER; DELL, 1994).
Além do benefício das micorrizas arbusculares na nutrição de fósforo
para as plantas, a absorção de outros macronutrientes também é influenciada
pela associação simbiótica. A absorção de nitrogênio por plantas micorrizadas
ocorre preferencialmente na forma amoniacal (NH4+) (GEORGE et al., 1992),

11
que pode está relacionado com a mobilidade da forma amoniacal do nitrogênio,
considerada menor qua a do nitrato (CANTARELLA, 2007). Além disso, devido
ao pequeno diâmetro, as hifas penetram mais facilmente em material orgânico
em decomposição, competindo melhor pelo nitrogênio recentemente
mineralizado (HODGE, 2003). Esse padrão é detectado também na absorção
de potássio, as hifas são as mediadoras desse processo (GEORGE et al.,
1992).
Para os micronutrientes a micorrização não só aumentam a
concentração dos elementos, em especial zinco, cobre e ferro, mas também
auxiliam as plantas a suportar condições de excesso, amenizando a toxidez,
podendo reduzir a absorção ou a translocação (CARDOSO et al., 2003;
SOARES; SIQUEIRA, 2008).
Plantas respondem de maneira diferente à micorrização, não somente
com nível de colonização, absorção de nutrientes e crescimento, mas também
ao nível de expressão gênica. Em alguns casos a resposta molecular e
fisiológica da planta ao fungo pode ser explicada pela abilidade do fungo em
suprir o hospedeiro com fosfato inorgânico (BURLEIGH, et al., 2002;
BURLEIGH; BECHMANN, 2002).
Freitas et al. (2006) avaliaram os efeitos de fungos micorrízicos
arbusculares no crescimento e composição mineral de Mentha arvensis L.,
cultivada com diferentes doses de fósforo, seus resultados demonstraram que
houve interações entre as espécies de fungos e doses de fósforo na produção
de matéria seca, nos conteúdos de nitrogênio, fósforo, potássio e de
manganês. Para todos esses parâmetros os resultados nos tratamentos
micorrizados foram superior aos tratamentos sem inoculação. Quando não se
utilizou adubação fosfatada, as plantas inoculadas com FMAs, apresentaram
conteúdos de N, P e K superiores ao tratamento controle, já os conteúdos de
cálcio, magnésio e enxofre foram influenciados pela aplicação de fósforo, mas
não foram afetados pelos tratamentos fúngicos.
Gupta et al. (2002) avaliaram o efeito de um fungo específico o Glomus
fasciculatum em diferentes cultivares de menta e demonstraram que a
inoculação incrementou a absorção de nitrogênio, fósforo e potássio, no
entanto de maneira mais pronunciada na absorção de fósforo.

12
Efeito do fósforo na colonização micorrízica
Micorrizas são sistemas biológicos compartimentalizados e sofrem
enorme influência do ambiente e de inúmeros fatores edáficos que influenciam
de modo direto ou indireto a formação, o funcionamento e a ocorrência dos
FMAs. Os componentes e os fatores controlados apresentam interação
constante e intensa de modo que a alteração em qualquer deles exercerá
influência sobre as micorrizas e os propágulos dos FMAs (MOREIRA;
SIQUEIRA, 2006).
As micorrizas são geralmente inibidas em condições de elevada
fertilidade tendo a taxa de colonização micorrízica intrarradicial uma diminuição
drástica com níveis elevados de fósforo no solo, enquanto adições moderadas
de fósforo podem até favorecer o efeito do fungo micorrízico sobre a nutrição e
crescimento da planta (CARDOSO et al., 2010).
Moreira e Siqueira (2006) propõem que o efeito de fósforo na
colonização não torna as plantas imunes à colonização, apenas reduz a
intensidade da micorrização e que geralmente, em concentrações próximas do
ótimo para o crescimento da planta hospedeira, já ocorre inibição da
colonização micorrízica. Nessas concentrações de fósforo, não há efeito
inibitório sobre os propágulos de fungo na rizosfera, a colonização é reduzida
por mecanismos de auto-regulação da simbiose. Os exsudatos de plantas
deficientes em fósforo estimulam o crescimento assimbiótico do fungo e, assim,
em condições de suprimento ótimo de fósforo, a colonização não será
estimulada.
Freitas et al. (2006) avaliaram os efeitos de fungos micorrízicos
arbusculares no crescimento e composição mineral de Mentha arvensis L.,
cultivada com diferentes doses de fósforo, eles verificaram que houve
interações entre as espécies de fungos e as doses de fósforo para a
porcentagem de colonização micorrízica. Quando não se utilizou adubação
fosfatada, as maiores porcentagens de colonização micorrízica foram com
Glomus clarum (69%) e Gigaspora margarita (81%), que também
proporcionaram maiores produções de matéria seca. Cada fungo micorrízico
apresentou um comportamento diferenciado, G. clarum e G. margarita

13
apresentaram colonização decrescente linearmente com o aumento da dose de
fósforo, já Glomus etunicatum e Acaulospora scrobiculata apresentaram uma
resposta quadrática, com ponto de máximo nas doses de fósforo estimadas em
99,1 e 95,5 mg kg-1 de solo.
Resultados semelhantes foram encontrado por Freitas et al. (2008) para
tanchagem (Plantago major L.), nesse trabalho cada espécie de FMAs também
apresentou comportamento diferente de acordo com a disponibilidade de P no
solo. A maior porcentagem de colonização micorrízica foi observada nas
espécies G. clarum (72,6%) e G. margarita (70,5%), quando não se utilizou
adubação fosfatada sendo que de uma maneira geral a porcentagem de
colonização diminuiu de acordo com o aumento da dose de fósforo.
Os efeitos negativos de alta disponibilidade de fósforo são muito
consistentes, representando quase uma regra na ecologia das micorrizas, mas
o mecanismo exato dessa inibição é ainda assunto não resolvido. Esses
mecanismos são complexos e podem diferir para os tipos de micorrizas e até
combinação fungo-planta, dificultando as generalizações (MOREIRA;
SIQUEIRA, 2006).
Fungos Micorrízicos Arbusculares e Plantas medicinais
Os FMAs têm um papel vital para sustentabilidade da agricultura em
regiões tropicais e apresentam grande potencial biotecnológico, que estão
ligados, justamente aos efeitos benéficos das micorrizas sobre a nutrição e
crescimento das plantas (BERBARA et al., 2006).
De uma forma geral, a maioria dos estudos demonstra a eficiência da
inoculação de fungos micorrízicos arbusculares no aumento do
desenvolvimento das espécies e na concentração dos princípios ativos, o que
incrementa o teor e o rendimento dos mesmos, como foi observado com,
Rosmarinus officinalis L. e Ocimum basilicum L. (RUSSOMANNO et al., 2008),
Mentha arvensis L. (GUPTA et al., 2002), Coriandrum sativum L. (KAPOOR et
al., 2002), Baccharis trimera (Less.) DC. (FREITAS et al., 2004b), Ocimum
basilicum L. var. Genovense (COPETTA et al., 2006), Artemisia umbelliformis

14
Lam (BINET et al., 2011) Inula ensifolia L. (ZUBEK et al., 2010) e Origanum sp.
(KHAOSAAD et al., 2006).
Algumas pesquisas relatam não somente a influência dos FMAs no teor
e no rendimento do óleo essencial, mas também na composição. Copetta et al.
(2007) demonstraram que Ocimum basilicum var. Genovense obteve uma
porcentagem maior de metileugenol e δcadineno quando inoculada com
Gigaspora margarita e Gigaspora rosea. Algo semelhante também foi
observado por Portugal et al. (2006) com Estevia rebaudiana (Bert.). Neste
trabalho os autores constataram que a quantidade de esteviosídeos foi
significativamente superior para Glomus intraradices (51%) em relação ao
tratamento com Acaulospora sp. e ao controle não inoculado. Com Artemisia
annua L., CHAUDHARY et al. (2008) verificaram que a concentração do
composto majoritário e de interesse industrial aumentou em tratamentos
micorrizados.
Determinados experimentos associam a eficiência micorrízica e
adubação fosfatada em busca de resultados que comprovem a eficiência dos
FMAs na absorção do fósforo e até que ponto este nutriente interfere no
desenvolvimento do fitobionte e do micobionte. Nesta linha de pesquisa
destaca-se trabalhos como o desenvolvido por Freitas et al. (2004a e 2006),
eles constataram que, sem adubação fosfatada o teor do óleo essencial de
Mentha arvensis L. e do seu composto majoritário o mentol foram menores em
plantas sem inoculação, e com a presença da adubação não houve diferença
nesses parâmetros entre os tratamentos. Outro trabalho de destaque foi o
publicado por Nell et al. (2009) comprovando que em Salvia officinalis L. o
conteúdo de fósforo foi maior em plantas fertilizadas e inoculadas, e o teor de
ácido rosmarínico foi afetado de maneira diferente na folha e na raiz sendo
incrementado em tratamentos submetidos à fertilização. No entanto, a
composição e concentração deste óleo não foram afetados. Kapoor et al.
(2004) realizaram trabalho semelhante para Foeniculum vulgare Mill,
verificando incremento na absorção de fósforo e no teor de óleo em
tratamentos micorrizados.
Na tentativa de comprovar a eficiência do FMA na absorção de fósforo
alguns estudos analisam o conteúdo de fósforo na planta, mas não realizam

15
adição de adubação fosfatada, como o que foi desenvolvido por Sailo e
Bagyaraj (2005). Nesse trabalho a biomassa, conteúdo de forskolin (diterpeno
presente na raiz da planta) e absorção de fósforo foram superiores em plantas
micorrizadas.
Outros trabalhos vão além e analisam até mesmo o potencial
bioprotetor. Toussaint et al. (2008) perceberam que a concentração de ácidos e
óleo essencial em Ocimum basilicum L. incrementam quando estão
micorrizadas e inoculadas com o fungo fitopatogênico Fusarium oxysporum. No
entanto o efeito bioprotetor não está relacionado a nutrição suplementar de
fósforo, pois a concentração deste foi similar em todos os tratamentos, mas a
simbiose com FMA diminuiu a concentração de metileugenol em plantas
tratadas com patógeno.
Os resultados apresentados em pesquisas que relacionam plantas
medicinais, FMAs e adição suplementar de fósforo podem em alguns
momentos alterar a composição, mas frequentemente afetam o teor e o
rendimento do óleo, provando que nem sempre os metabólitos secundários são
influenciados pelo fósforo, mas depende da interação planta-FMA (KHAOSAAD
et al., 2006).

16
REFERÊNCIAS BERBARA, R. L. L.; SOUZA, F. A.; FONSECA, H. M. A. C. Fungos Micorrízicos Arbusculares: muito além da nutrição 2006. In: FERNANDES, M. S. Nutrição Mineral de Plantas. Viçosa, MG. Sociedade Brasileira de Ciências do Solo. 2006. 432 p. BIASI, L. A.; DESCHAMPS, C. Plantas Aromáticas do Cultivo à produção de óleo essencial. 1ª Edição. Curitiba: Layer Studio Gráfico e Editora Ltda, 2009. 160 p. BINET, M. N.; VAN TUINEN, D.; DEPRÊTRE, N.; KOSZELA, N.; CHAMBON, C.; GIANINAZZI, S. Arbuscular mycorrhizal fungi associated with Artemisia umbelliformis Lam, an endangered aromatic species in Southern French Alps, influence plant P and essential oil contents. Mycorrhiza, v. 21, p. 523–535, 2011. BROWN M. S.; BETHLENFALVAY, G. J. Glycine-Glomus-Rhizobium symbiosis. VI. Photosynthesis in nodulated, mycorrhizal, or N- and P-fertilized soybean plants. Plant Physiology, v. 85, p. 120-123, 1987. BURLEIGH, S. H.; BECHMANN, I. E. Plant nutrient transporter regulation in arbuscular mycorrhizas. Plant and Soil, v. 244, p. 247-251, 2002. BURLEIGH, S. H.; CAVAGNARO, T.; JAKOBSEN, I. Functional diversity of arbuscular mycorrhizas extends to the expression of plant genes involved in P nutrition. Journal of Experimental Botany, v. 53, n. 374, p. 1593-1601, july 2002. CANTARELLA, H. Nitrogênio. In: NOVAIS, R. F.; ALVAREZ, V. H. V.; BARROS, N. F.; FONTES, R. L. F.; CANTARUTTI, R. B.; NEVES, J. C. L. Fertilidade do Solo. Viçosa, Sociedade Brasileira de Ciência do Solo, 2007. p. 375-470. CARDOSO, E. J. B. N.; CARDOSO, I. M.; NOGUEIRA, M. A.; BARRETA, C. R. D. M.; de PAULA, A. M. Micorrizas Arbusculares na aquisição de nutrientes pelas plantas. In: SIQUEIRA, J. O.; de SOUZA, F. A.; CARDOSO, E. J. B. N.; TSAI, S. M. Micorrizas: 30 anos de pesquisas no Brasil. Lavras: Ufla, 2010. 716 p. CARDOSO, I. M.; BODDINGTON, C.; JANSSEN, B. H.; OENEMA, O.; KUYPER, T. W. Distribuition of micorrhizal fungal spore in soils under agroforestry and monocultural coffee systems in Brazil. Agroforest. Syst., v. 58, p. 33-43, 2003. CASTRO, H. G.; FERREIRA, F. A.; SILVA, D. J. H.; MOSQUIM, P. R. Contribuição ao estudo das plantas medicinais: metabólitos secundários. 2. ed. Visconde do Rio Branco: Editora Suprema, 2004. 113 p.

17
COSTA, L. C. B. Condições culturais, anatomia foliar, processamento e armazenamento de Ocimum selloi em relação ao óleo essencial. 2008. 161f. Dissertacão (Doutorado em Agronomia), Universidade Federal de Lavras, Lavras – MG, 2008. CHAUDHARY, V.; KAPOOR, R.; BHATNAGAR, A. K. Effectiveness of two arbuscular mycorrhizal fungi on concentrations of essential oil and artemisinin in three accessions of Artemisia annua L. Applied Soil Ecology, v.40, p. 174-181, 2008. COPETTA, A.; LINGUA, G.; BARDI, L.; MASOERO, G.;BERTA, G. Influence of arbuscular mycorrhizal fungi on growth and essential oil composition in Ocimum basilicum var. Genovese. CARYOLOGIA, v. 60, n. 1-2, p. 106-110, 2007. COPETTA, A.; LINGUA, G.; BERTA, G. Effects of three AM fungi on growth, distribution of glandular hairs, and essential oil production in Ocimum basilicum L. var. Genovese. Mycorrhiza, v. 16, p. 485-494, 2006. CROTEAU, R.; KUTCHAN, T. M.; LEWIS, N. G. Natural Products (Secondary Metabolites) In: Buchanan, B.; Gruissem, W.; Jones, R. (Eds). Biochemistry & Molecular Biology of Plants. Rockville: Courier Companies, 2000, p. 1250-1318. FREITAS, M. S. M.; MARTINS, M. A.; CARVALHO, A. J. C. Produção de biomassa e teores de macronutrientes da tanchagem (Plantago major L.) em resposta a adubação fosfatada e micorrizas arbusculares. Revista Brasileira Plantas Medicinais, Botucatu, v.10, n.3, p.31-37, 2008. FREITAS M. S. M; MARTINS M. A.; CARVALHO A. J. C. Crescimento e composição da menta em resposta à inoculação com fungos micorrízicos arbusculares e adubação fosfatada. Horticultura Brasileira, v. 24, p.11-16, 2006. FREITAS, M. S. M.; MARTINS, M. A.; VIEIRA, I. J. C. Produção e qualidade de óleos essenciais de Mentha arvensis em resposta à inoculação de fungos micorrízicos arbusculares. Pesquisa Agropecuária Brasileira, v. 39, p.887-894, 2004a. FREITAS, M. S. M.; MARTINS, M. A.; CARVALHO, A. J. C.; CARNEIRO, R. F. V. Crescimento e Produção de fenóis totais em carqueja [Baccharis trimera (Less.) DC.] em resposta à inoculação com fungos micorrízicos arbusculares, na presença e na ausência de adubação mineral. Revista Brasileira de Plantas Medicinais, Botucatu, v.6, n. 3, p.30-34, 2004b. GEORGE, E. K.; HAUSSIER, G.; VETTERLEIN, E. G.; MARSCHNER, H. Water and nutrient translocation by hiphae of Glomus mossae. Can. J. Bot. v. 70, p. 2130-2137, 1992.

18
GOBBO-NETO, L.; LOPES, N. P. Plantas medicinais: fatores de influência no conteúdo de metabólitos secundários. Química Nova, v. 30, n. 2, p. 374-381, 2007. GUPTA, M. L.; PRASAD, A.; RAM, M.; KUMAR, S. Effect of tha vesicular-arbuscular mycorrhizal (VAM) fungus Glomus fasciculatum on the essential oil yield related characters and nutrient acquisition in the crops of different cultivars of menthol mint (Mentha arvensis) under fild conditions. Bioresource Technology, v. 81, p. 77-79, 2002. HODGE, A. Plant nitrogen capture from organic matter as affected by spatial dispersion, interspecific competition and mycorrhizal colonization. New Phytol., v. 157, p. 303-314, 2003. HWANG S. F.; CHANG K. F.; CHAKRAVARTY, P. Effects of vesicular-arbuscular mycorrhizal fungi on the development of Verticillium and Fusarium wilts of alfalfa. Plant Disease, v. 76, p. 239-243, 1992. KAPOOR, R.; GIRI, B.; MUKERJI, K. G. Improved growth and essential oil yield and quality in Foeniculum vulgare mill on mycorrhizal inoculation supplemented with P-fertilizer. Bioresource Technology, v. 93, p. 307–311, 2004. KAPOOR, R.; GIRI, B.; MUKERJI, K.G. Mycorrhization of coriander (Coriandrum sativum L.) to enhance the concentration and quality of essential oil. Journal of the Science of Food and Agriculture, v. 82, p. 339-342, 2002. KAROUSOU, R.; GRAMMATIKOPOULOS, G.; LANARAS, T.; MANETAS, Y.; KOKKINI, S. Effects Of Enhanced Uv-B Radiation On Mentha spicata Essential Oils. Phytochemistry, v. 49, n. 8, p. 2273±2277, 1998. KHADE, S. W.; RODRIGUES, B. F. Applications of arbuscular mycorrhizal fungi in agroecosystems. Tropical and Subtropical Agroecosystems, v. 10, p. 337-354, 2009. KHAOSAAD, T.; VIERHEILIG, H.; NELL, M.; ZITTERL-EGLSEER, K.; NOVAK, J. Arbuscular mycorrhiza alter the concentration of essential oils in oregano (Origanum sp., Lamiaceae). Mycorrhiza, v. 16, p. 443 – 446, 2006. LORENZI, H.; MATOS, F. J. De A. Plantas Medicinais no Brasil: nativas e exóticas. Nova Odessa,SP: Instituto Plantarum, 2002. 544 p. MARSCHNER, H. Mineral nutrition of higher plants. San Diego, Academic Press, 1995. 889p. MARSCHNER, H.; DELL, B. Nutrient uptake in mycorrhizal symbiosis. Plant Soil, v. 159, p. 89-102, 1994.

19
MONTEIRO, R. Desenvolvimento de menta e produção de óleo essencial sob diferentes condições de manejo. 81f. Dissertação (Mestrado em Agronomia) Ciências Agrárias, Universidade Federal do Paraná. Curitiba, 2009. MORAIS L. A. S. Influência dos fatores abióticos na composição química dos óleos essenciais. Horticultura Brasileira v. 27, p. S4050 - S4063, 2009. MOREIRA, F. M. S.; SIQUEIRA, J. O. Microbiologia e bioquímica do solo. Lavras, MG: Editora UFLA, 2006. 729p. MORTON, J. B.; BENNY, G. L. Revised classification of arbuscular mycorrhizal fungi (Zygomycetes): a new order Glomales and Gigasporineae and two new families Acaulosporaceae and Gigasporaceae with an emendation of Glomaceae. Mycotaxon. v. 37, p. 471–491, 1990 MORTON, J. B.; REDECKER, D. Two new families of Glomales, Archaeosporaceae and Paraglomaceae, with two new genera Archaeospora and Paraglomus, based on concordant molecular and morphological characters. Mycologia. v. 93, p. 181–195, 2001. MURRAY, M.J.; LINCOLN, D. E. The genetic basis of acyclic oil constituents in Mentha citrate Ehrh. Genetics, v. 65, p. 457-471, 1970. NELL, M.; VÖTSCH, M.; VIERHEILIG, H.; STEINKELLNER, S.; ZITTERL-EGLSEER, K.; FRANZA, C.; NOVAK, J. Effect of phosphorus uptake on growth and secondary metabolites of garden sage (Salvia officinalis L.) Journal Science Food Agriculturae, v. 89, p. 1090-1096, 2009. OLIVEIRA de, A. R. M. F. Produção de óleo essencial de Mentha x piperita var. citrata sob diferentes condições de manejo. 2011, 83f. Dissertação (Mestrado em Produção Vegetal) Universidade Estadual de Santa Cruz, 2011a. OLIVEIRA de, A. R. M. F.; JEZLER, C. N.; OLIVEIRA, R. A.; COSTA, L. C. do B. Potencial alelopático, produção de biomassa e óleo essencial de alevante (Mentha x piperita var. citrata (Ehrh.) Briq.) em cultivo solteiro e consorciado com cebolinha (Allium schoenoprasum L.) e chicória (Chicorium endivia L.). R. Bras. Bioci., v. 9, n. 4, p. 497-501, out./dez. 2011b. PORTUGAL, E. P.; QUITÉRIO, G. C. M.; HONÓRIO, S. L. Seleção de fungos micorrízicos arbusculares para estévia, Stevia Rebaudiana (Bert.) Bertoni. Construindo a História dos Produtos Naturais. Multi Ciência, Outubro 2006. PERES, L. E. P. Metabolismo Secundário. Piracicaba – São Paulo: Escola Superior de Agricultura Luiz de Queiroz. ESALQ/USP, 2004. p. 1-10. RICHARDSON, A. E.;BAREA, J. M.; McMEILL, A. M.; PRIGENT-COMBARET, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant and Soil, v. 321, p. 305-339, 2009.

20
RUSSOMANNO, O. M. R. Influência de fungos micorrízicos arbusculares associados ou não a Fusarium oxysporum Schecht. sobre plantas de alecrim (Rosmarinus officinalis l.) e manjericão (Ocimum basilicum L.). 2006. Tese. 88 p. Faculdade de Ciências Agronômicas da UNESP - Câmpus de Botucatu. Maio 2006 RUSSOMANO, O. M. R.; KRUPTA, P. C.; MINHONI, M. T. A. Influencia de Fungos Micorrízicos Arbusculares no Desenvolvimento de Plantas de Alecrim e Manjericão. Arquivos do Instituto de Biologia, v. 75, p. 37-43, 2008. SAILO, G. L., BAGYARAJ, D. J. Influence of different AM-fungi on the growth, nutrition and forskolin content of Coleus forskohlii. Mycological Research, v. 109, p. 795-798, 2005. SANGWAN, N. S.; FAROOQI, A.H.A.; SHABIH, F.; SANGWAN, R. S. Regulation of essential oil production in plants. Plant Growth Regulation, v. 34, p. 3–21, 2001. SIQUEIRA J. O., SAGGIN-JUNIOR O. J. Dependency on arbuscular mycorrhizal fungi and responsiveness of some Brazilian native woody species. Mycorrhiza, v. 11, p. 245-255, 2001. SOARES, C. R. F. S.; SIQUEIRA, J. O. Mycorrhizal and phosphate protection of tropical Grass species against heavy metal toxicity in muilti-contaminad soil. Biol. Fert. Soils, v. 44, p. 833-841, 2008. SOUZA, V. C.; LORENZI, H. Botânica Sistemática: guia ilustrado para identificação das famílias de angiospermas da flora brasileira, baseado em APG II. Nova Odessa,SP: Instituto Plantarum, 2005. 523 p. SCHÜBLER, A. Glomeromycota PHYLOGENY. Disponível em http://www.lrz.de/~schuessler/amphylo/ Acesso em: 20 de setembro de 2012. SCHÜBLER, A.; SCHWARZOTT, D.; WALKER, C. A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycological Research. v. 105, p. 1413-1421, 2001. TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 3. ed. Porto Alegre: Artmed, 2004. 719 p. TOUSSAINT, J. P.; KRAML, M.; NELL, M.; SMITH, S. E.; SMITH,F. A.; STEINKELLNER, S.; SCHMIDERER, C.; VIERHEILIG, H.; NOVAK, J. Effect of Glomus mosseae on concentrations of rosmarínico and caffeic acids and essential oil compounds in basil inoculated with Fusarium oxysporum f.sp. basilica. Plant Pathology, v. 57, p. 1109–1116, 2008. TURNES, G. W.; GERSHENZON, J.; CROTEAU, R.B. Distribution of Peltate Glandular Trichomes on Developing Leaves of Peppermint. Plant Physiology, v. 124, p. 655–663, October 2000.

21
VAN BREEMEN, N.; LUNDSTROM, U. S.; JONGMANS, A. G.; VAN BREEMEN, N.; LUNDSTROM, U. S.; VAN BREEMEN, N.; BAIN, D. C. Do plants drive podzolization via rock-eating mycorrhizal fungi? Geoderma, v. 94, p. 163-171, 2000. VOLPIN, H.; ELKIND, Y.; OKON, Y.; KAPULNIK, Y. A vesicular arbuscular mycorrhizal fungus (Glomus intraradix) induces a defense response in alfafa roots. Plant Physiology, v. 104, p. 683-689, 1994. ZUBEK, S.; STOJAKOWSKA, A.;ANIELSKA, T.; TURNAU, K. Arbuscular mycorrhizal fungi alter thymol derivative contents of Inula ensifolia L. Mycorrhiza, v. 20, p. 497–504, 2010.

22
1. Capítulo I - CRESCIMENTO E COMPOSIÇÃO MINERAL DE MENTHA X PIPERITA L. VAR. CITRATA (EHRH.) BRIQ. INOCULADA COM FUNGOS
MICORRÍZICOS ARBUSCULARES SUBMETIDA A DUAS DOSES DE FÓSFORO
RESUMO
A associação entre fungos micorrízicos arbusculares e raízes é benéfica para ambos os simbiontes, para a planta hospedeira ela amplia a capacidade de absorção dos nutrientes melhorando o estado nutricional do vegetal e aumentando a sua produtividade. Essa associação recebe influência de fatores edáficos como, por exemplo, a disponibilidade de fósforo no solo. O presente trabalho teve como objetivo avaliar a influência de fungos micorrízicos arbusculares no crescimento e na composição mineral de Mentha x piperita L. var. citrata (Ehrh.) Briq. submetida a duas doses de fósforo no solo. O experimento foi conduzido em casa de vegetação na Universidade Estadual de Santa Cruz num fatorial 5x2, sendo cinco tratamentos micorrízicos (controle não inoculado, inoculação com Acaulospora morrowiae Spain & Schenck, Glomus clarum Nicol. & Schenck, e Scutellospora calospora (Nicol. & Gerd.) Walk. & Sand., e a mistura desse inóculos) e duas doses de fósforo (60 mg e 120 mg de fósforo por dm3) com cinco repetições cada. Após 75 dias as plantas foram coletadas e avaliadas quanto à produção de biomassa seca, porcentagem de colonização micorrízica, número de esporos no solo e composição de macro e micronutrientes das folhas. Os FMAs incrementaram a produção de biomassa seca da raiz, caule, folha e total. Para a colonização micorrízica G. clarum apresentou uma maior porcentagem na maior dose de P, enquanto S. calospora obteve maior colonização das raízes na menor dose de P. O número de esporos presente em 50 g de solo foi significativamente maior para os tratamentos inoculados com os FMAs em comparação com o controle não inoculado, entretanto não houve influência das doses de P na quantidade de esporos encontrada. Os tratamentos micorrízicos e as doses de fósforo influenciaram a absorção dos nutrientes e o crescimento (biomassa) das plantas. Palavras-chave: Acaulospora, Glomus, Scutellospora, Conteúdo de nutrientes, Planta Medicinal.

23
ABSTRACT
The association between mycorrhizal fungi and roots is beneficial to both symbionts, to the host plant it increases the capacity for absorption of nutrients by improving the nutritional status of the plant and increasing its productivity. This association is influenced by the soil factors such as the availability of phosphorus. This study aimed to evaluate the influence of mycorrhizal fungi on growth and mineral composition of Mentha x piperita L. var. citrata (Ehrh.) Briq. on two phosphorus doses in the soil. The experiment was installed in a greenhouse at the Universidade Estadual de Santa Cruz in a 5x2 factorial design, five mycorrhizal treatments (uninoculated control, inoculation with Acaulospora morrowiae Spain & Schenck, Glomus clarum Nicol. & Schenck, e Scutellospora calospora (Nicol. & Gerd.) Walk. & Sand., and the mixture of inocula) and two doses of phosphorus (60 mg and 120 mg of phosphorus per dm-3 of soil) with five replicates each. After 75 days the plants were collected and evaluated for biomass dry weight, percentage of mycorrhizal colonization, spore numbers in soil and composition of macro and micronutrients of leaves. The AMF increased the production of dry biomass of root, stem, leaf and total. For the colonization G. clarum showed a higher percentage of colonization at the highest P dose and S. calospora obtained higher mycorrhizal root colonization in the lower dose of P. The number of spores present in 50 g of soil was significantly higher for the treatments inoculated with mycorrhizal fungi compared to uninoculated control, however there was no influence of P levels on spore number. The mycorrhizal treatments and phosphorus dose influenced nutrient content and growth (biomass production) of plants. Keywords: Acaulospora, Glomus, Scutellospora, nutrient content, Medicinal Plant.

24
1.1 INTRODUÇÃO
Mentha x piperita L. var. citrata (Ehrh.) Briq. é conhecida popularmente
como hortelã-limão, pertencente à família Lamiaceae tem como componente
principal do seu óleo essencial o linalol e o acetato de linalila (GARLET, 2007;
MURRAY; LINCOLN, 1970). A presença desses compostos aumenta o
interesse econômico pela espécie, pois são utilizados nas indústrias de
perfumaria, cosmética, de alimentos, medicamentos, fragrâncias e tabaco,
principalmente como aromatizante e flavorizante de alimentos e na confecção
de perfumes (GARLET, 2007). Além disso, estes compostos estão associados
à defesa da planta contra patógenos e herbívoros e a atração de polinizadores
(CROTEAU et al., 2000).
De acordo com Marschner (1995) as plantas necessitam de energia
solar (armazenada sob a forma de ATP, NADPH), CO2, água e nutrientes
minerais para seu desenvolvimento. Atualmente são citados dezessete
nutrientes essenciais, ou seja, que desempenham funções vitais no
desenvolvimento do vegetal, sendo a maioria desses nutrientes absorvidos do
solo.
Os fungos micorrízicos arbusculares otimizam a absorção de nutrientes
do solo, aumentando a superfície de contato entre a planta e o solo por meio
de hifas extrarradiculares que colonizam sítios não possíveis de serem
atingidos pela raízes. Para alguns autores, as micorrizas podem ser mais do
que simplesmente uma extensão do sistema radicular, contribuindo na
absorção de nutrientes pela planta em até 80% de fósforo, 60% de cobre, 25%
de nitrogênio, 25% de zinco e 10% de K (MARSCHNER; DELL, 1994). Freitas
et al. (2006) em experimento com M. arvensis detectaram aumento de
nitrogênio, fósforo, potássio e manganês em função da inoculação com FMA e
adubação fosfatada. Gupta et al. (2002) também com espécies do gênero
Mentha comprovaram o aumento na absorção de nitrogênio, fósforo e potássio
em plantas inoculadas com Glomus fasciculatum. Em experimento em casa de
vegetação Rasouli-Sadaghiani et al. (2010) detectaram o incremento do
conteúdo de nitrogênio, fósforo, potássio, cálcio, cobre, manganês e ferro em

25
manjericão (Ocimum basilicum L.) inoculada com três espécies diferentes de
Glomus.
O efeito dos fungos micorrízicos arbusculares sobre o crescimento das
plantas é especialmente significativo com relação aos nutrientes de baixa
mobilidade no solo como por exemplo fósforo, ferro e zinco (MARSCHNER,
1995). A melhora da nutrição fosfatada tem sido reconhecida como um dos
maiores benefícios das micorrizas, as respostas, entretanto, variam
especialmente com o nível de fósforo, com a espécie de FMA presente e com a
cultivar ou espécie de planta (CARDOSO et al., 2010). Karagiannidis et al.
(2011) em experimento em casa de vegetação observaram que plantas de
orégano e menta não-micorrizadas tiveram níveis menores de macro e micro
nutrientes, e que o orégano inoculado com Glomus etunicatum tiveram
aumento na concentração de fósforo. Com três acessos de Artemisia annua,
sob condições controladas Chaudhary et al. (2008) observaram que houve um
aumento na absorção de fósforo, ferro e zinco quando inoculada com duas
espécies diferentes de Glomus.
As micorrizas são geralmente inibidas em condições de elevada
fertilidade tendo a taxa de colonização micorrízica intrarradical uma diminuição
drástica, com níveis elevados de fósforo no solo. Porém em solos deficientes
ou extremamente fixadores de fósforo, adições moderadas de fósforo podem
favorecer o efeito de fungos micorrízicos sobre a nutrição e crescimento da
planta (CARDOSO et al., 2010). A dependência micorrízica tem estreita relação
com a dose de fósforo. Balota et al. (2010) avaliaram o efeitos de FMA em
culturas de girassol, amendoim e mamona submetidas a diferentes doses de
fósforo e concluíram que o girassol e o amendoim apresentaram moderada
dependência micorrízica, enquanto que a mamona apresentou alta
dependência aos FMA, e que a colonização micorrízica radicular nas
oleaginosas foi inibida significativamente com o aumento da dose de P
adicionada ao solo.
Entre os fatores que interferem no crescimento em plantas, a nutrição
merece destaque, pois a deficiência ou excesso de nutrientes pode intervir na
produção da biomassa e consequentemente na quantidade de óleos essenciais
(MORAIS, 2009). Desta maneira, qualquer fator que possa interferir na

26
absorção de nutrientes pela planta, como é o caso das micorrizas, deve ser
estudado com vistas ao aumento da produtividade de massa vegetal.
O objetivo deste trabalho foi avaliar o efeito dos fungos micorrízicos
arbusculares no crescimento e composição de macro e micronutriente em
Mentha x piperita L. var. citrata (Ehrh.) Briq. sob duas doses de fósforo.
1.2 MATERIAL E MÉTODOS
O experimento foi conduzido em casa de vegetação no campus da
Universidade Estadual de Santa Cruz, Ilhéus – BA, sob condições naturais de
luminosidade e temperatura. O delineamento experimental foi constituído por
um fatorial 5x2, sendo utilizados três inóculos de FMA: Acaulospora morrowiae
(Am) Spain & Schenck (A79 CNPAB 037), Glomus clarum (Gc) Nicol. &
Schenck (A5 CNPAB 005) e Scutellospora calospora (Sc) (Nicol. & Gerd.)
Walk. & Sand. (A80 CNPAB 038), além de um controle formado por plantas
não inoculadas (C) e um tratamento com a mistura de todos os três inóculos
(M), perfazendo um total de cinco tratamentos micorrízicos, e duas doses
diferentes de fósforo 60 mg (P60) e 120 (P120) mg dm-3 sendo cinco repetições
cada. A unidade experimental foi composta por um vaso de plástico contendo
quatro dm3 de solo e uma planta por vaso. Os inóculos foram gentilmente
cedidos pela coleção de fungos micorrízicos da Embrapa Agrobiologia
(Seropédica, RJ).
O solo utilizado no experimento foi um argissolo amarelo distrófico típico,
coletado da camada de 0 a 20 cm em área de pastagem de Brachiaria
humidicola com remanescente de dendê na Estação Experimental do Almada
(39º10’ W e 14º38’ S), localizada em Uruçuca (BA) e pertencente à UESC. O
solo foi seco ao ar e tamisado em peneira com malha de 5 mm e suas
características químicas (Tabelas 1) e físicas (Tabela 2) foram determinadas
segundo Embrapa (1997). Esse solo foi autoclavado por uma hora a 121 °C e
1,5 atm por dois dias consecutivos para esterilização.

27
Tabela 1. Características químicas iniciais (antes do início do experimento) do Argissolo Amarelo Distrófico típico coletado na Estação
Experimental da Almada, localizada no município de Uruçuca, BA
Amostra pH M.O. P K Ca Mg Al H + Al SB CTC V B Cu Fe Mn Zn
CaCl2 g dm-3 mg dm-3 ------------------------mmolc dm-3---------------------- % ----------------------mg dm-3--------------------
Solo inicial 4,0 17 ˂ 2 0,4 5 3 5,4 47 7,9 54,8 14 0,49 ˂ 0,3 171 2,0 0,9
P, K, Ca e Mg = Resina trocadora de íons; H + Al = pH SMP; M. O. = dicromato/colorimetria; B = água quente/microndas; Cu, Fe, Mn e Zn = extração DTPA-TEA pH 7,3.
Tabela 2. Características físicas iniciais (antes do início do experimento) do Argissolo Amarelo Distrófico típico coletado na Estação
Experimental do Almada, localizada no município de Uruçuca, BA
Amostra Areia grossa Areia fina Areia total Silte Argila Classificação textural
-------------------------------g kg-1------------------------------
Solo inicial 142 720 861 13 125 Arenosa

28
Com o solo coletado foi realizada uma curva de incubação usando cinco
doses de calcário dolomítico com PRNT de 82,02 %. Os resultados foram
analisados por regressão para ajustar o pH em torno de 6,5. Além do calcário,
que também serviu de fonte para o cálcio e o magnésio, o solo contido nos
vasos recebeu uma adubação individual com base na análise do solo, na
produtividade da espécie e no comportamento do fungo, constituída por 100
mg dm-3 de nitrogênio na forma de sulfato de amônio; duas doses de fósforo 60
e 120 mg dm-3 de fósforo como superfosfato simples; 80 mg dm-3 de potássio
sob a forma de cloreto de potássio; 1,55 mg dm-3 de ferro na forma de Ferro
EDTA; 0,75 mg dm-3 de boro na forma de ácido bórico; 1,33 mg dm-3 de cobre
sob a forma de sulfato de cobre; 0,15 mg dm-3 de molibdênio na forma de
molibdato de amônio; 4 mg dm-3 de zinco e de manganês na forma de sulfato
de zinco e sulfato de manganês, respectivamente. A adubação foi subdividida
em duas partes uma anterior a autoclavagem onde foi adicionado ao solo o
calcário, o superfosfato simples, cloreto de potássio em solução e os
micronutrientes (boro, cobre, molibdênio, zinco e manganês) também em
solução e outra posterior a autoclavagem que recebeu a adição do nitrogênio
sob forma de solução preparado com água autoclavada e o ferro também em
solução que foi autoclavada separada. Antes da transferência do solo para os
vasos, estes foram lavados com água e desinfestados superficialmente com
etanol 70 % e com hipoclorito de sódio comercial (2 a 2,5 %).
A propagação da M. x piperita var. citrata foi vegetativa, mudas foram
obtidas junto ao Horto de Plantas Medicinais da Universidade Estadual de
Santa Cruz, Ilhéus-BA, e estacas de aproximadamente 5 cm de comprimento
foram e colocadas para enraizar em bandejas de polipropileno com substrato
areia e vermiculita (2:1) autoclavado por uma hora a 121 °C e 1,5 atm, onde
permaneceram por quatorze dias até a data do transplante.
No transplante, os inóculos de FMAs foram concomitantemente
adicionados ao solo junto das raízes das mudas e por fim pedriscos
autoclavados por 20 minutos a 121 °C e 1,5 atm foram colocados para diminuir
a superfície de contato do solo com o ar e minimizar as possíveis
contaminações. Durante a condução do experimento as plantas foram irrigadas
com água deionizada autoclavada.

29
Após 75 dias de crescimento na casa de vegetação, as plantas foram
coletadas, separadas em raiz, caule e folhas, embaladas em sacos de papel
Kraft devidamente identificados para determinação de biomassa e avaliação
nutricional.
1.2.1 Determinação da biomassa
Após a coleta e separação, o material foi seco a 70 °C em estufa de
circulação forçada e em seguida foi efetuada a pesagem em balança semi
analítica até peso constante para a obtenção da biomassa seca da raiz, caule,
folha e total.
1.2.2 Teor de nutrientes
Para a análise da composição de macro e micronutrientes (conforme
Embrapa, 1997, modificado) o material sofreu digestão com ácido nítrico. Após
a determinação da biomassa seca as folhas foram moídas em moinho de bola
por 2 min e armazenados em frascos plásticos em desumidificador, depois
pesadas aproximadamente 0,2010 mg do material em balança analítica dentro
dos tubos digestores previamente descontaminados em ácido nítrico 10 %. Em
seguida os tubos foram transferidos para o bloco digestor onde o processo de
digestão foi realizado, na primeira parte do processo foi adicionado 3 ml de
ácido nítrico nos tubos que permaneceram por 30 min a 50 °C, depois a
temperatura foi elevada para 120 °C por uma hora e meia. Ao final do processo
foi adicionado 1 ml de peróxido de hidrogênio por três vezes consecutivas, a
120 °C por 20 min. Posteriormente o material contido no tubo digestor foi
vertido para tubos falcon e volumado pra 12 ml e armazenado na geladeira até
leitura em espectrofotômetro óptico (ICP AOS) Variant 710-ES do Centro de
Microscopia Eletrônica da UESC.
1.2.3 Porcentagem de colonização micorrízica
A estimativa da porcentagem de colonização micorrízica foi realizada em
triplicata a partir de amostras aleatórias das raízes que foram clarificadas e
coradas de acordo com o método de Phillips e Haymann (1970) modificado.
Durante a coleta as raízes mais finas e claras foram selecionadas e

30
armazenadas em álcool 50 %, depois foram lavadas em água corrente e
clareadas, primeiro com a remoção do protoplama utilizando solução de KOH
10 % em banho Maria a aproximadamente 60°C durante 30 min., lavadas com
água em abundância, e depois o clareamento foi finalizado com peróxido de
hidrogênio a 10 % por 20 min em temperatura ambiente. Em seguida as raízes
foram lavadas com água destilada e coradas com corante acidófilo a base de
tinta de caneta preta e ácido acético a 5 % e armazenadas em lactoglicerol na
proporção 1 : 1 : 1 (v/v/v) de ácido láctico, glicerina e água.
A estimativa da colonização foi calculada pelo método de placa riscada
modificado (McGONIGLE et al., 1990). Foram feitas 10 listras verticais em
lâminas e as raízes foram depositadas horizontalmente e na interseção entre a
listra e a raiz foi observado a presença ou a ausência de micorrização bem
como o tipo de estrutura detectada.
1.2.4 Número de esporos de FMAs
A determinação da quantidade de esporos de FMAs também foi
realizada em triplicata, utilizando o método de peneiramento úmido de
Gerdemann e Nicolson (1963) modificado. Amostras de solo dos vasos, ao final
do experimento, foram coletadas e armazenadas em refrigerador. Dessas
amostras foram utilizadas 50 g, que após serem peneiradas foram
centrifugadas por duas vezes, a primeira vez com água destilada por três
minutos a 2.500 rpm depois com sacarose 50 % por um minuto também à
2.500 rpm, e por fim os esporos foram contados utilizando estereomicroscópio
no aumento de 40X.
1.2.5 Análise estatística
Todos os dados obtidos foram submetidos à análise de variância, os
tratamentos fosfatados foram comparados pelo teste de F a 5% de
probabilidade. Para as médias dos tratamentos fúngicos foi utilizado o teste t
de contrastes ortogonais (Tabela 3) a 5% de probabilidade. No caso de
significância da interação FMA x dose de fósforo foi realizado o seu
desdobramento. As análises estatísticas foram realizadas pelo programa
estatístico SISVAR (Ferreira, 2000).

31
Tabela 3. Descrição dos contrastes ortogonais para a avaliação da influencia de FMAs
em Mentha x piperita var. citrata submetida a duas doses de fósforo
Contraste ortogonais Comparação
Ŷ1 = 4 1m̂ - 2m̂ - 3m̂ - 4m̂ - 5m̂ Controle vs. Demais - contraste um (1)
Ŷ2 = 2m̂ + 3m̂ + 4m̂ - 3 5m̂ A. morrowiae, G. clarum,S. calospora vs.
Mistura dos inóculos - contraste dois (2)
Ŷ3 = 2m̂ + 4m̂ - 2 3m̂ A. morrowiae e S. calospora vs. G. clarum -
contraste três (3)
Ŷ4 = 2m̂ - 4m̂ A. morrowiae vs. S. calospora - contraste
quatro (4)
Ŷ5 = 4 4m̂ - 1m̂ - 2m̂ - 3m̂ - 5m̂ S. calospora vs. demais – contraste (5)
Ŷ6 = 3 4m̂ - 2m̂ - 3m̂ - 5m̂ S. calospora vs. A. morrowiae,G. clarum e
mistura dos inóculos – contraste (6)
Ŷ7 = 2m̂ - 3m̂ A. morrowiae vs. G. clarum – contraste (7)
1.3 RESULTADOS E DISCUSSÃO
1.3.1 Biomassa
Houve interação entre a dose de P e os FMAs inoculados em Mentha x
piperita para as variáveis biomassa seca da raiz, do caule, da folha e total
conforme observado na Tabela 4.
32
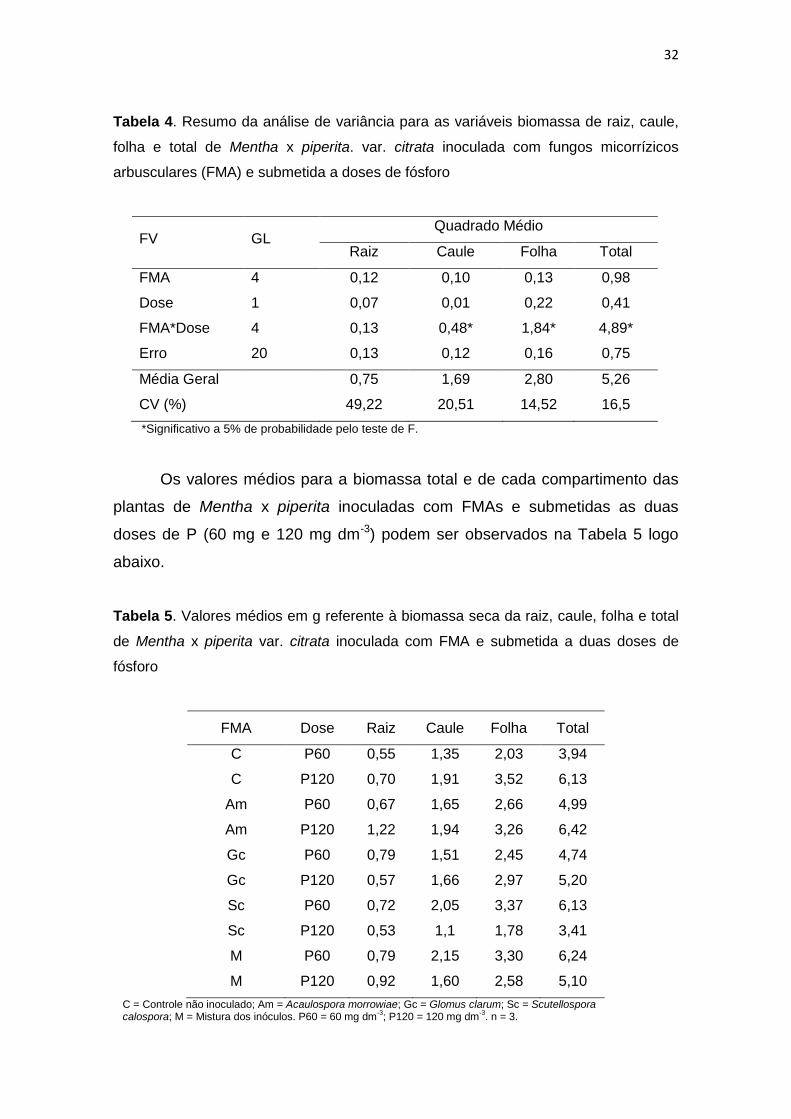
Tabela 4. Resumo da análise de variância para as variáveis biomassa de raiz, caule,
folha e total de Mentha x piperita. var. citrata inoculada com fungos micorrízicos
arbusculares (FMA) e submetida a doses de fósforo
FV GL Quadrado Médio
Raiz Caule Folha Total
FMA 4 0,12 0,10 0,13 0,98
Dose 1 0,07 0,01 0,22 0,41
FMA*Dose 4 0,13 0,48* 1,84* 4,89*
Erro 20 0,13 0,12 0,16 0,75
Média Geral 0,75 1,69 2,80 5,26
CV (%) 49,22 20,51 14,52 16,5
*Significativo a 5% de probabilidade pelo teste de F.
Os valores médios para a biomassa total e de cada compartimento das
plantas de Mentha x piperita inoculadas com FMAs e submetidas as duas
doses de P (60 mg e 120 mg dm-3) podem ser observados na Tabela 5 logo
abaixo.
Tabela 5. Valores médios em g referente à biomassa seca da raiz, caule, folha e total
de Mentha x piperita var. citrata inoculada com FMA e submetida a duas doses de
fósforo
FMA Dose Raiz Caule Folha Total
C P60 0,55 1,35 2,03 3,94
C P120 0,70 1,91 3,52 6,13
Am P60 0,67 1,65 2,66 4,99
Am P120 1,22 1,94 3,26 6,42
Gc P60 0,79 1,51 2,45 4,74
Gc P120 0,57 1,66 2,97 5,20
Sc P60 0,72 2,05 3,37 6,13
Sc P120 0,53 1,1 1,78 3,41
M P60 0,79 2,15 3,30 6,24
M P120 0,92 1,60 2,58 5,10
C = Controle não inoculado; Am = Acaulospora morrowiae; Gc = Glomus clarum; Sc = Scutellospora calospora; M = Mistura dos inóculos. P60 = 60 mg dm
-3; P120 = 120 mg dm
-3. n = 3.
33
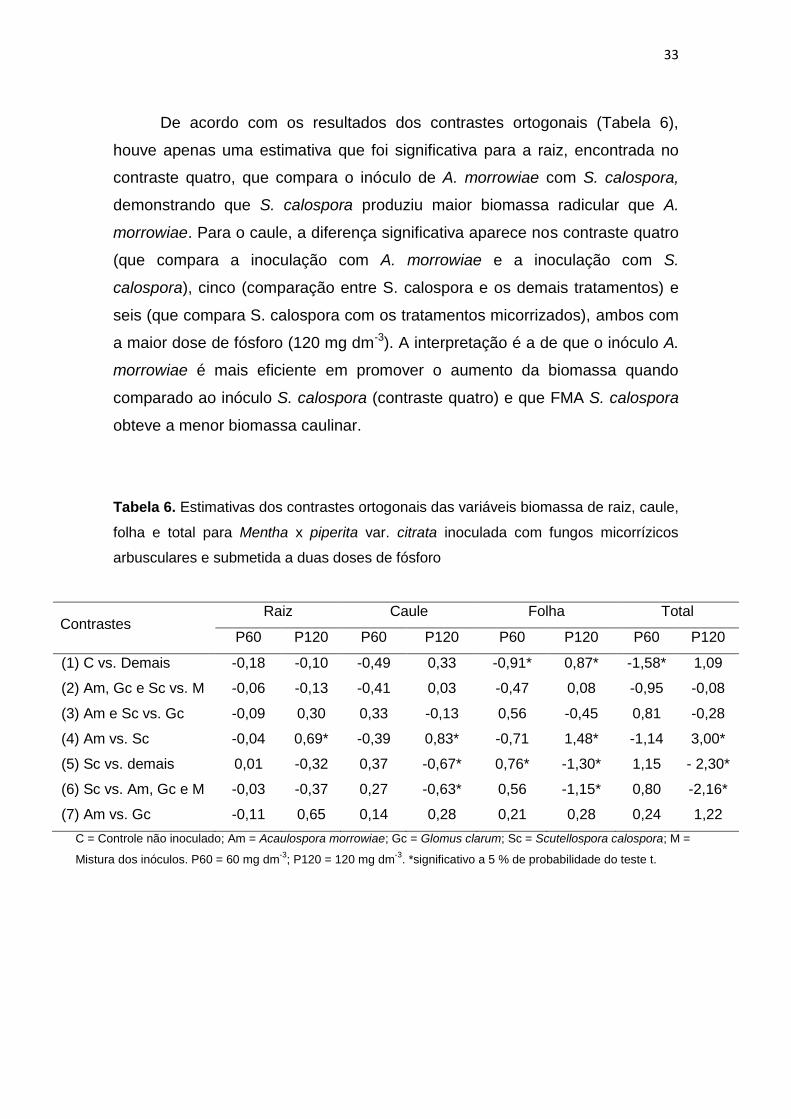
De acordo com os resultados dos contrastes ortogonais (Tabela 6),
houve apenas uma estimativa que foi significativa para a raiz, encontrada no
contraste quatro, que compara o inóculo de A. morrowiae com S. calospora,
demonstrando que S. calospora produziu maior biomassa radicular que A.
morrowiae. Para o caule, a diferença significativa aparece nos contraste quatro
(que compara a inoculação com A. morrowiae e a inoculação com S.
calospora), cinco (comparação entre S. calospora e os demais tratamentos) e
seis (que compara S. calospora com os tratamentos micorrizados), ambos com
a maior dose de fósforo (120 mg dm-3). A interpretação é a de que o inóculo A.
morrowiae é mais eficiente em promover o aumento da biomassa quando
comparado ao inóculo S. calospora (contraste quatro) e que FMA S. calospora
obteve a menor biomassa caulinar.
Tabela 6. Estimativas dos contrastes ortogonais das variáveis biomassa de raiz, caule,
folha e total para Mentha x piperita var. citrata inoculada com fungos micorrízicos
arbusculares e submetida a duas doses de fósforo
Contrastes Raiz Caule Folha Total
P60 P120 P60 P120 P60 P120 P60 P120
(1) C vs. Demais -0,18 -0,10 -0,49 0,33 -0,91* 0,87* -1,58* 1,09
(2) Am, Gc e Sc vs. M -0,06 -0,13 -0,41 0,03 -0,47 0,08 -0,95 -0,08
(3) Am e Sc vs. Gc -0,09 0,30 0,33 -0,13 0,56 -0,45 0,81 -0,28
(4) Am vs. Sc -0,04 0,69* -0,39 0,83* -0,71 1,48* -1,14 3,00*
(5) Sc vs. demais 0,01 -0,32 0,37 -0,67* 0,76* -1,30* 1,15 - 2,30*
(6) Sc vs. Am, Gc e M -0,03 -0,37 0,27 -0,63* 0,56 -1,15* 0,80 -2,16*
(7) Am vs. Gc -0,11 0,65 0,14 0,28 0,21 0,28 0,24 1,22
C = Controle não inoculado; Am = Acaulospora morrowiae; Gc = Glomus clarum; Sc = Scutellospora calospora; M =
Mistura dos inóculos. P60 = 60 mg dm-3
; P120 = 120 mg dm-3
. *significativo a 5 % de probabilidade do teste t.

34
Para a variável biomassa seca da folha o contraste um (que compara o
controle com os demais tratamentos micorrizados) e cinco (que compara a
inoculação com S. calospora com os demais tratamentos) para menor dose de
fósforo (60 mg dm-3) apresentaram diferença significativa, indicando que os
tratamentos micorrizados foram superiores ao controle e que a inoculação com
S. calospora produziu maior massa seca de folhas que os demais tratamentos
micorrízicos. Para a maior dose de fósforo (120 mg dm-3), os contrastes um,
quatro (comparação entre A. morrowiae e S. calospora), cinco e seis (que
compara S. calospora com os demais tratamentos micorrizados) expressaram
diferenças significativas. Isso indica que a utilização dos FMAs reduziu a
produção da biomassa foliar em comparação ao controle. Além disso
evidencia-se a redução de biomassa foliar no tratamento inoculado com S.
calospora quando comparado aos demais tratamentos inoculados com FMAs.
Na biomassa total a diferença significativa surge quando a comparação
é feita entre o controle e os tratamentos micorrizados (contraste um) na menor
dose de fósforo (60 mg dm-3), evidenciando o incremento na produção da
biomassa provocada pela inoculação dos FMAs.
Na maior dose de fósforo (120 mg dm-3) a significância aparece nos
contrastes quatro, que compara o inóculo A. morrowiae com o S. calospora,
cinco, que realiza comparação entre S. calospora com os demais tratamentos e
seis, que compara o inóculo de S. calospora somente com os tratamentos
micorrizados. A interpretação dessas estimativas é a de que o tratamento
inoculado com S. calospora não provocou aumento na biomassa das plantas
de M. x piperita var. citrata quando comparado aos demais. O conceito de um
contínuo que vai do benefício ao prejuízo para a associação micorrízica
(JOHNSON et al., 1997), é baseado na ausência de resposta do crescimento
causada pelo custo da simbiose que supera os benefícios. Nesse caso, a baixa
absorção de P da solução do solo via hifas do FMA e a alta demanda por
carboidratos do fungo provocaria o reduzido crescimento da planta. Essa é
uma hipótese que poderia explicar a pequena produção de biomassa nas
plantas de M. x piperita var. citrata inoculadas com S. calospora na dose de
fósforo de 120 mg dm-3.

35
Entretanto, vale salientar que o fungo não pode ser taxado simplesmente
de parasita, por que a via fúngica de absorção de P é funcional, embora varie
na quantidade do elemento repassado ao vegetal de acordo com a espécie
fúngica (SMITH et al., 2011) e por que diversos outros benefícios podem ser
provocados pela inoculação micorrízica (SMITH; READ, 2008).
Diversos trabalhos comprovam que a micorrização aumenta a
produtividade da massa vegetal de plantas medicinais, como observado por
Karagiannidis et al. (2011) em condições controladas para plantas de orégano
e menta. Para o gênero estudado, Freitas et al., (2006) observaram que plantas
de Mentha arvensis inoculadas com FMAs na ausência de adubação fosfatada
apresentaram acréscimo na produção de biomassa superior às plantas
inoculadas e adubadas com fósforo. Outra informação importante foi a
publicada por Rasouli-Sadaghiani et al. (2010) em pesquisa com plantas
micorrizadas de Ocimum basilicum L., outra espécie produtora de óleo
essencial, as quais apresentaram não somente incremento na biomassa seca,
mas também aumento na altura da planta e massa seca da raiz quando
comparadas ao controle não micorrizado. O aumento da biomassa é,
provavelmente, consequência da melhoria do estado nutricional da planta
micorrizada, em que hifas extracelulares expandem a superfície de contato da
planta com o solo colonizando sítios antes não atingidos pela raiz e assim
otimizando a absorção de nutrientes (MOREIRA; SIQUEIRA, 2006).

36
1.3.2 Teor dos nutrientes
Houve interação entre os FMAs inoculados e as doses de P adicionadas
ao solo para o teor do potássio, cobre e manganês na folha de M. x piperita.
Para o enxofre apenas a inoculação dos fungos micorrízicos influenciou
significativamente. Para o teor de cálcio e de ferro na folha somente a dose de
P propiciou diferenças entre os tratamentos. Já o teor de fósforo na folha
apresentou diferenças significativas para as doses de P e inoculação de FMAs
separadamente não havendo interação entre esses dois fatores. Os teores de
magnésio e zinco na folha não foram significativamente influenciados pela
adubação fosfatada e inoculação dos FMAs (Tabela 7), entretanto nos
desdobramentos das estimativas dos contrastes, foram observadas diferenças
significativas no teor de Zn e Mg foliares entre os tratamentos.
Os valores médios para os teores de macro e micronutrientes nas folhas
de Mentha x piperita var. citrata inoculadas com FMAs e submetidas as duas
doses de P (60 mg e 120 mg dm-3) podem ser observados na Tabela 8.
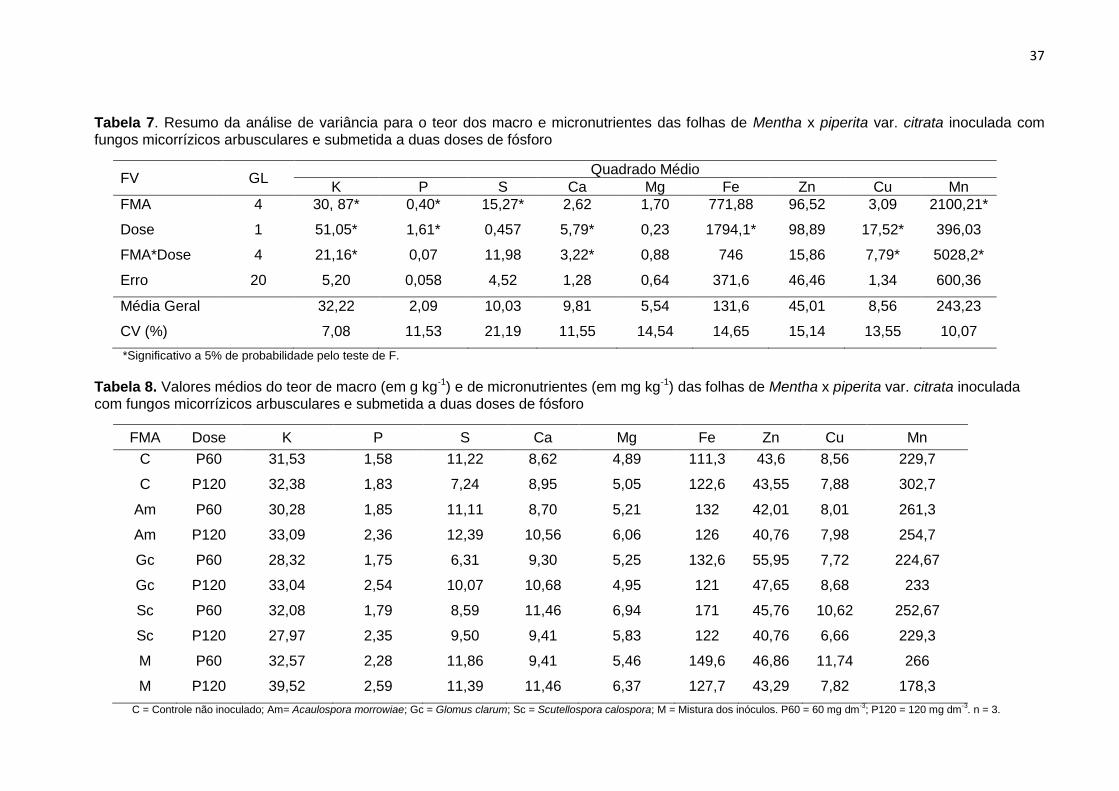
37
Tabela 7. Resumo da análise de variância para o teor dos macro e micronutrientes das folhas de Mentha x piperita var. citrata inoculada com fungos micorrízicos arbusculares e submetida a duas doses de fósforo
FV GL Quadrado Médio
K P S Ca Mg Fe Zn Cu Mn
FMA 4 30, 87* 0,40* 15,27* 2,62 1,70 771,88 96,52 3,09 2100,21*
Dose 1 51,05* 1,61* 0,457 5,79* 0,23 1794,1* 98,89 17,52* 396,03
FMA*Dose 4 21,16* 0,07 11,98 3,22* 0,88 746 15,86 7,79* 5028,2*
Erro 20 5,20 0,058 4,52 1,28 0,64 371,6 46,46 1,34 600,36
Média Geral 32,22 2,09 10,03 9,81 5,54 131,6 45,01 8,56 243,23
CV (%) 7,08 11,53 21,19 11,55 14,54 14,65 15,14 13,55 10,07
*Significativo a 5% de probabilidade pelo teste de F.
Tabela 8. Valores médios do teor de macro (em g kg-1) e de micronutrientes (em mg kg-1) das folhas de Mentha x piperita var. citrata inoculada com fungos micorrízicos arbusculares e submetida a duas doses de fósforo
FMA Dose K P S Ca Mg Fe Zn Cu Mn
C P60 31,53 1,58 11,22 8,62 4,89 111,3 43,6 8,56 229,7
C P120 32,38 1,83 7,24 8,95 5,05 122,6 43,55 7,88 302,7
Am P60 30,28 1,85 11,11 8,70 5,21 132 42,01 8,01 261,3
Am P120 33,09 2,36 12,39 10,56 6,06 126 40,76 7,98 254,7
Gc P60 28,32 1,75 6,31 9,30 5,25 132,6 55,95 7,72 224,67
Gc P120 33,04 2,54 10,07 10,68 4,95 121 47,65 8,68 233
Sc P60 32,08 1,79 8,59 11,46 6,94 171 45,76 10,62 252,67
Sc P120 27,97 2,35 9,50 9,41 5,83 122 40,76 6,66 229,3
M P60 32,57 2,28 11,86 9,41 5,46 149,6 46,86 11,74 266
M P120 39,52 2,59 11,39 11,46 6,37 127,7 43,29 7,82 178,3
C = Controle não inoculado; Am= Acaulospora morrowiae; Gc = Glomus clarum; Sc = Scutellospora calospora; M = Mistura dos inóculos. P60 = 60 mg dm-3; P120 = 120 mg dm
-3. n = 3.

38
De acordo com as estimativas do contraste um, que compara o controle
com os demais tratamentos micorrizados (Tabela 9), a utilização dos inóculos
aumentou a absorção de fósforo e de enxofre em comparação ao controle na
maior dose de fósforo (120 mg dm-3). No entanto, o contraste dois (comparação
entre os inóculos separados com a mistura) mostrou que a utilização dos
inóculos em conjunto aumentou a absorção do P e de S na menor dose de
fósforo (60 mg dm-3) comparativamente aos inóculos utilizados em separado.
Na maior dose de P (120 mg dm-3), o contraste dois (comparação entre
os inóculos separados com a mistura) indicou que a utilização dos inóculos em
conjunto aumentou a absorção do K comparativamente aos inóculos aplicados
em separado.
Na menor dose de P (60 mg dm-3) o teor de S nas folhas de M. x piperita
foi maior nos tratamentos inoculados com A. morrowiae e com S. calospora
comparado com o tratamento inoculado com G. clarum, como ilustra o
contraste três.
A absorção de potássio foi significativamente baixa para o tratamento
com S. calospora na maior dose de fósforo (120 mg dm-3) quando comparada
A. morrowiae (contraste quatro), na comparação com os demais tratamentos
(contraste cinco) e quando comparada com os tratamentos que receberam
adição dos inóculos de FMAs (contraste seis). Porém o inóculo de S. calospora
na menor dose de fósforo (60 mg dm-3) aumentou o teor de cálcio e magnésio
na folha de M. x piperita quando comparada com as plantas inoculadas com A.
morrowiae como pode ser observado no contraste quatro, quando comparado
aos demais tratamentos (contraste cinco) e quando comparado somente com
os tratamentos que receberam adição de FMA (contraste seis).
O contraste sete (comparação entre A. morrowiae versus G. clarum) foi
significativo apenas no teor de enxofre na menor dose de P (60 mg dm-3),
evidenciando que o inóculo de A. morrowiae é mais eficiente em promover a
absorção desse nutriente.

39
Com relação aos micronutrientes (Tabela 9) o contraste um (que
compara o controle com os tratamentos micorrizados) mostra que os FMAs
aumentaram a absorção de ferro com a menor dose de fósforo (60 mg dm-3),
comportamento inverso foi observado com relação à absorção de manganês
onde o controle mostrou-se mais eficiente na maior dose de P (120 mg dm-3). A
utilização dos isolados individualmente quando comparada com a mistura de
FMAs (contraste dois) reduziu a absorção de cobre na menor dose de fósforo
(60 mg dm-3). As espécies de FMAs inoculadas individualmente (contrate dois)
aumentaram a absorção de manganês quando comparadas ao tratamento
inoculado com a mistura de FMAs. S. calospora incrementou a absorção de
ferro e cobre (contraste quatro e cinco) e somente o cobre (contraste seis) na
menor dose de P (60 mg dm-3), já G. clarum (contraste três e sete) aumentou a
absorção de zinco também na menor dose de P (60 mg dm-3).
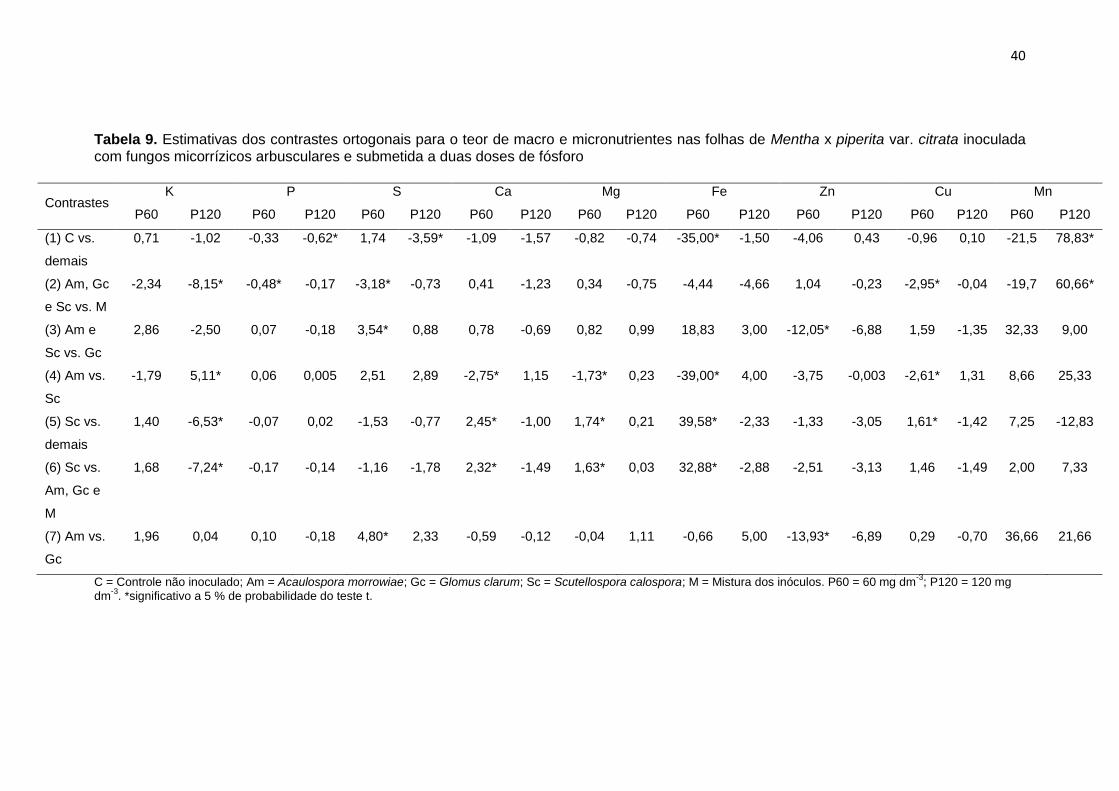
40
Tabela 9. Estimativas dos contrastes ortogonais para o teor de macro e micronutrientes nas folhas de Mentha x piperita var. citrata inoculada com fungos micorrízicos arbusculares e submetida a duas doses de fósforo
Contrastes K P S Ca Mg Fe Zn Cu Mn
P60 P120 P60 P120 P60 P120 P60 P120 P60 P120 P60 P120 P60 P120 P60 P120 P60 P120
(1) C vs.
demais
0,71 -1,02 -0,33 -0,62* 1,74 -3,59* -1,09 -1,57 -0,82 -0,74 -35,00* -1,50 -4,06 0,43 -0,96 0,10 -21,5 78,83*
(2) Am, Gc
e Sc vs. M
-2,34 -8,15* -0,48* -0,17 -3,18* -0,73 0,41 -1,23 0,34 -0,75 -4,44 -4,66 1,04 -0,23 -2,95* -0,04 -19,7 60,66*
(3) Am e
Sc vs. Gc
2,86 -2,50 0,07 -0,18 3,54* 0,88 0,78 -0,69 0,82 0,99 18,83 3,00 -12,05* -6,88 1,59 -1,35 32,33 9,00
(4) Am vs.
Sc
-1,79 5,11* 0,06 0,005 2,51 2,89 -2,75* 1,15 -1,73* 0,23 -39,00* 4,00 -3,75 -0,003 -2,61* 1,31 8,66 25,33
(5) Sc vs.
demais
1,40 -6,53* -0,07 0,02 -1,53 -0,77 2,45* -1,00 1,74* 0,21 39,58* -2,33 -1,33 -3,05 1,61* -1,42 7,25 -12,83
(6) Sc vs.
Am, Gc e
M
1,68 -7,24* -0,17 -0,14 -1,16 -1,78 2,32* -1,49 1,63* 0,03 32,88* -2,88 -2,51 -3,13 1,46 -1,49 2,00 7,33
(7) Am vs.
Gc
1,96 0,04 0,10 -0,18 4,80* 2,33 -0,59 -0,12 -0,04 1,11 -0,66 5,00 -13,93* -6,89 0,29 -0,70 36,66 21,66
C = Controle não inoculado; Am = Acaulospora morrowiae; Gc = Glomus clarum; Sc = Scutellospora calospora; M = Mistura dos inóculos. P60 = 60 mg dm-3
; P120 = 120 mg
dm-3
. *significativo a 5 % de probabilidade do teste t.

41
Geralmente os FMAs melhoram a captação de nutrientes para o vegetal,
mas a eficiência da absorção de cada um dos elementos depende de
mecanismos intrínsecos da interação planta e FMAs (ORTAS, 2010; GUPTA et
al., 2002; SAILO; BAGYARAJ, 2005).
O efeito dos fungos micorrízicos arbusculares sobre o crescimento das
plantas é especialmente significativo com relação aos nutrientes de baixa
mobilidade (MARSCHNER, 1995). Neste grupo situam-se primordialmente o
macronutriente fósforo e os micronutrientes zinco e cobre (CARDOSO et al.,
2010). A explicação mais aceita é a de que as micorrizas exploram uma área
maior de solo, em relação às raízes (CARDOSO et al., 2010). Porém as
micorrizas podem ser mais do que simplesmente uma extensão do sistema
radicular, contribuindo na absorção de nutrientes pela planta em até 80% de
fósforo, 60% de cobre, 25% de nitrogênio, 25% de zinco e 10% de K
(MARSCHNER; DELL, 1994).
Plantas respondem de maneira diferente à micorrização, e diferentes
espécies de fungos micorrízicos induzem efeitos diversos, não somente no
nível de colonização, mas também na absorção de nutrientes e crescimento do
vegetal. Em alguns casos a resposta molecular e fisiológica da planta ao fungo
pode ser explicada pela habilidade do fungo em suprir o hospedeiro com
fosfato inorgânico (BURLEIGH, et al., 2002; BURLEIGH; BECHMANN, 2002).
A análise do solo, no fim do experimento (Tabela 10), indica que os
tratamentos com a maior dose de superfosfato simples apresentaram maiores
concentrações de P solúvel no solo comparativamente com o solo que recebeu
a menor dose de superfosfato, confirmando o estabelecimento do tratamento
de fósforo.
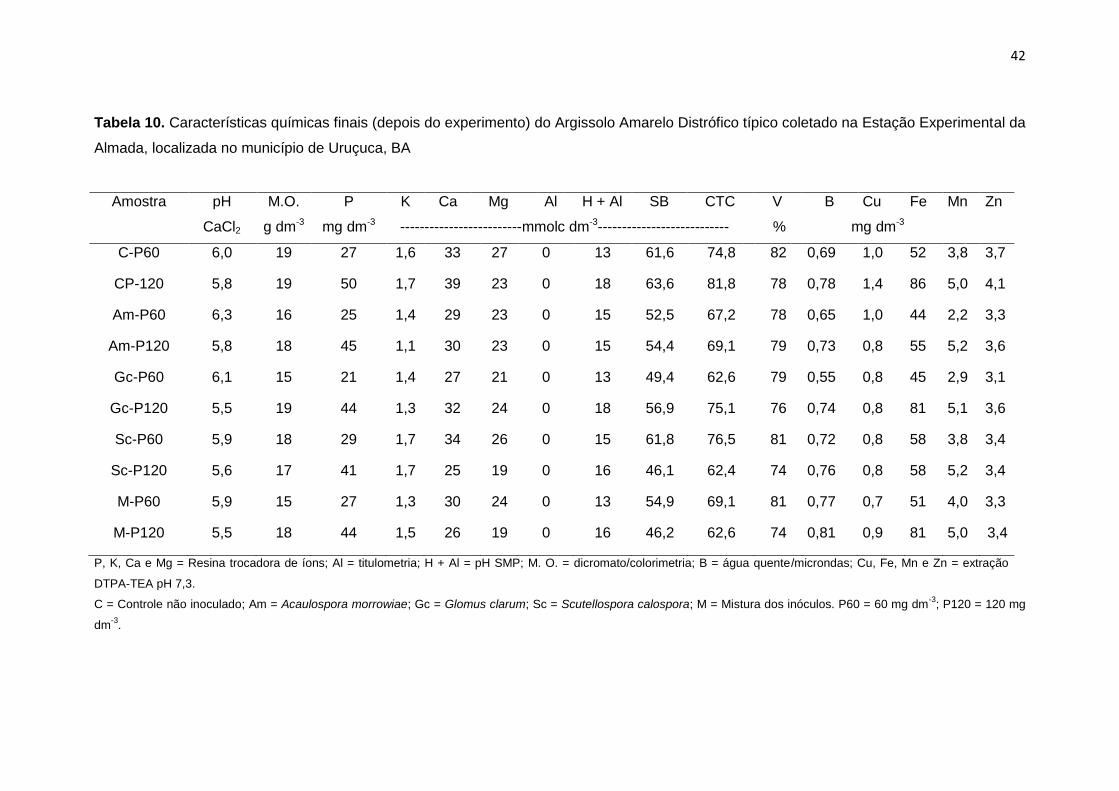
42
Tabela 10. Características químicas finais (depois do experimento) do Argissolo Amarelo Distrófico típico coletado na Estação Experimental da
Almada, localizada no município de Uruçuca, BA
Amostra pH M.O. P K Ca Mg Al H + Al SB CTC V B Cu Fe Mn Zn
CaCl2 g dm-3 mg dm-3 -------------------------mmolc dm-3--------------------------- % mg dm-3
C-P60 6,0 19 27 1,6 33 27 0 13 61,6 74,8 82 0,69 1,0 52 3,8 3,7
CP-120 5,8 19 50 1,7 39 23 0 18 63,6 81,8 78 0,78 1,4 86 5,0 4,1
Am-P60 6,3 16 25 1,4 29 23 0 15 52,5 67,2 78 0,65 1,0 44 2,2 3,3
Am-P120 5,8 18 45 1,1 30 23 0 15 54,4 69,1 79 0,73 0,8 55 5,2 3,6
Gc-P60 6,1 15 21 1,4 27 21 0 13 49,4 62,6 79 0,55 0,8 45 2,9 3,1
Gc-P120 5,5 19 44 1,3 32 24 0 18 56,9 75,1 76 0,74 0,8 81 5,1 3,6
Sc-P60 5,9 18 29 1,7 34 26 0 15 61,8 76,5 81 0,72 0,8 58 3,8 3,4
Sc-P120 5,6 17 41 1,7 25 19 0 16 46,1 62,4 74 0,76 0,8 58 5,2 3,4
M-P60 5,9 15 27 1,3 30 24 0 13 54,9 69,1 81 0,77 0,7 51 4,0 3,3
M-P120 5,5 18 44 1,5 26 19 0 16 46,2 62,6 74 0,81 0,9 81 5,0 3,4
P, K, Ca e Mg = Resina trocadora de íons; Al = titulometria; H + Al = pH SMP; M. O. = dicromato/colorimetria; B = água quente/microndas; Cu, Fe, Mn e Zn = extração
DTPA-TEA pH 7,3.
C = Controle não inoculado; Am = Acaulospora morrowiae; Gc = Glomus clarum; Sc = Scutellospora calospora; M = Mistura dos inóculos. P60 = 60 mg dm-3
; P120 = 120 mg
dm-3
.

43
1.3.3 Porcentagem de colonização micorrízica e número de esporos
Houve interação entre a dose de P e os FMAs inoculados em Mentha x
piperita para a colonização micorrízica das raízes, conforme pode ser
observado na Tabela 11. Para a variável número de esporos de FMAs em 50 g
de solo, somente o fator inoculação de FMAs resultou em efeito significativos
(Tabela 11).
Tabela 11. Resumo da Análise de Variância para o número de esporos e porcentagem de colonização micorrízica em Mentha x piperita var. citrata submetida a duas doses de fósforo
FV GL Quadrado Médio
Esporos Colonização
FMA 4 13889,08* 1807,5*
Dose 1 13,33 316,03*
FMA*Dose 4 4474,91 338,5*
Erro 20 3391,56 32,25
Média Geral 160,66 32,28
CV (%) 36,25 16,10
*Significativo a 5% de probabilidade pelo teste de F.
Os valores médios do número de esporos em 50 g de solo e da
porcentagem de colonização micorrízica e da presença de estruturas nas
raízes de Mentha x piperita, inoculadas com FMAs e submetidas as duas doses
de P (60 mg e 120 mg dm-3), podem ser observados na Tabela 12.
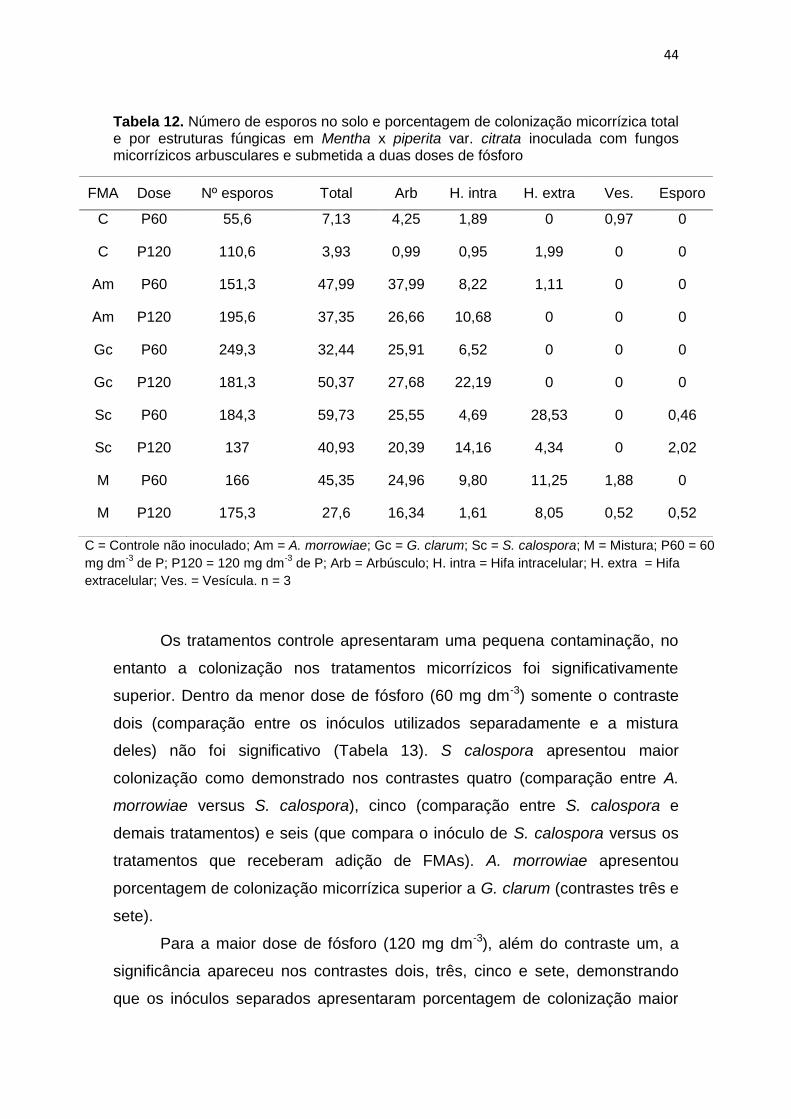
44
Tabela 12. Número de esporos no solo e porcentagem de colonização micorrízica total e por estruturas fúngicas em Mentha x piperita var. citrata inoculada com fungos micorrízicos arbusculares e submetida a duas doses de fósforo
C = Controle não inoculado; Am = A. morrowiae; Gc = G. clarum; Sc = S. calospora; M = Mistura; P60 = 60
mg dm-3
de P; P120 = 120 mg dm-3
de P; Arb = Arbúsculo; H. intra = Hifa intracelular; H. extra = Hifa
extracelular; Ves. = Vesícula. n = 3
Os tratamentos controle apresentaram uma pequena contaminação, no
entanto a colonização nos tratamentos micorrízicos foi significativamente
superior. Dentro da menor dose de fósforo (60 mg dm-3) somente o contraste
dois (comparação entre os inóculos utilizados separadamente e a mistura
deles) não foi significativo (Tabela 13). S calospora apresentou maior
colonização como demonstrado nos contrastes quatro (comparação entre A.
morrowiae versus S. calospora), cinco (comparação entre S. calospora e
demais tratamentos) e seis (que compara o inóculo de S. calospora versus os
tratamentos que receberam adição de FMAs). A. morrowiae apresentou
porcentagem de colonização micorrízica superior a G. clarum (contrastes três e
sete).
Para a maior dose de fósforo (120 mg dm-3), além do contraste um, a
significância apareceu nos contrastes dois, três, cinco e sete, demonstrando
que os inóculos separados apresentaram porcentagem de colonização maior
FMA Dose Nº esporos Total Arb H. intra H. extra Ves. Esporo
C P60 55,6 7,13 4,25 1,89 0 0,97 0
C P120 110,6 3,93 0,99 0,95 1,99 0 0
Am P60 151,3 47,99 37,99 8,22 1,11 0 0
Am P120 195,6 37,35 26,66 10,68 0 0 0
Gc P60 249,3 32,44 25,91 6,52 0 0 0
Gc P120 181,3 50,37 27,68 22,19 0 0 0
Sc P60 184,3 59,73 25,55 4,69 28,53 0 0,46
Sc P120 137 40,93 20,39 14,16 4,34 0 2,02
M P60 166 45,35 24,96 9,80 11,25 1,88 0
M P120 175,3 27,6 16,34 1,61 8,05 0,52 0,52
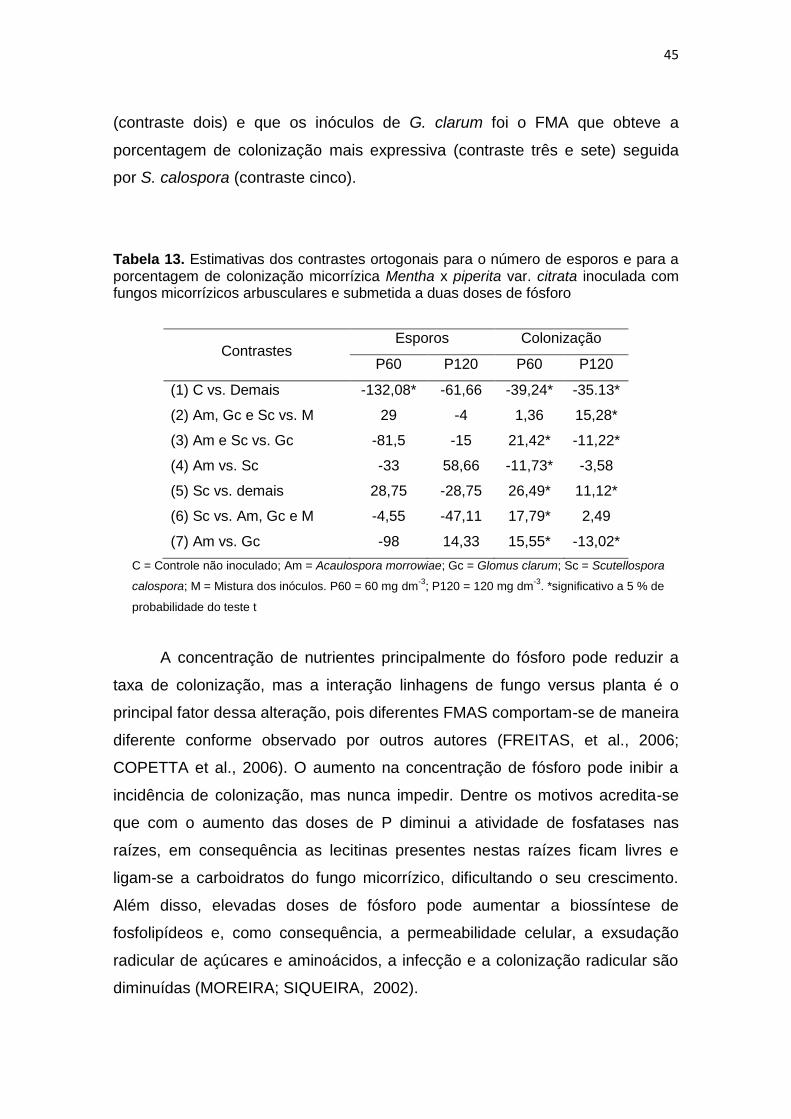
45
(contraste dois) e que os inóculos de G. clarum foi o FMA que obteve a
porcentagem de colonização mais expressiva (contraste três e sete) seguida
por S. calospora (contraste cinco).
Tabela 13. Estimativas dos contrastes ortogonais para o número de esporos e para a porcentagem de colonização micorrízica Mentha x piperita var. citrata inoculada com fungos micorrízicos arbusculares e submetida a duas doses de fósforo
Contrastes Esporos Colonização
P60 P120 P60 P120
(1) C vs. Demais -132,08* -61,66 -39,24* -35.13*
(2) Am, Gc e Sc vs. M 29 -4 1,36 15,28*
(3) Am e Sc vs. Gc -81,5 -15 21,42* -11,22*
(4) Am vs. Sc -33 58,66 -11,73* -3,58
(5) Sc vs. demais 28,75 -28,75 26,49* 11,12*
(6) Sc vs. Am, Gc e M -4,55 -47,11 17,79* 2,49
(7) Am vs. Gc -98 14,33 15,55* -13,02*
C = Controle não inoculado; Am = Acaulospora morrowiae; Gc = Glomus clarum; Sc = Scutellospora
calospora; M = Mistura dos inóculos. P60 = 60 mg dm-3
; P120 = 120 mg dm-3
. *significativo a 5 % de
probabilidade do teste t
A concentração de nutrientes principalmente do fósforo pode reduzir a
taxa de colonização, mas a interação linhagens de fungo versus planta é o
principal fator dessa alteração, pois diferentes FMAS comportam-se de maneira
diferente conforme observado por outros autores (FREITAS, et al., 2006;
COPETTA et al., 2006). O aumento na concentração de fósforo pode inibir a
incidência de colonização, mas nunca impedir. Dentre os motivos acredita-se
que com o aumento das doses de P diminui a atividade de fosfatases nas
raízes, em consequência as lecitinas presentes nestas raízes ficam livres e
ligam-se a carboidratos do fungo micorrízico, dificultando o seu crescimento.
Além disso, elevadas doses de fósforo pode aumentar a biossíntese de
fosfolipídeos e, como consequência, a permeabilidade celular, a exsudação
radicular de açúcares e aminoácidos, a infecção e a colonização radicular são
diminuídas (MOREIRA; SIQUEIRA, 2002).

46
Os arbúsculos foram as estruturas mais abundantes em todos os
tratamentos, independente da dose indicando que todos os tratamentos
formaram simbioses funcionais. As hifas intracelulares obtiveram maiores
porcentagens nos tratamentos micorrizados com exceção da mistura dos
inóculos, já as hifas extracelulares tiveram maior desenvolvimento na menor
dose de fósforo, com exceção do inóculo G. clarum. Vesículas e esporos foram
as estruturas com menor incidência e podem ser observadas na Fig. 1.
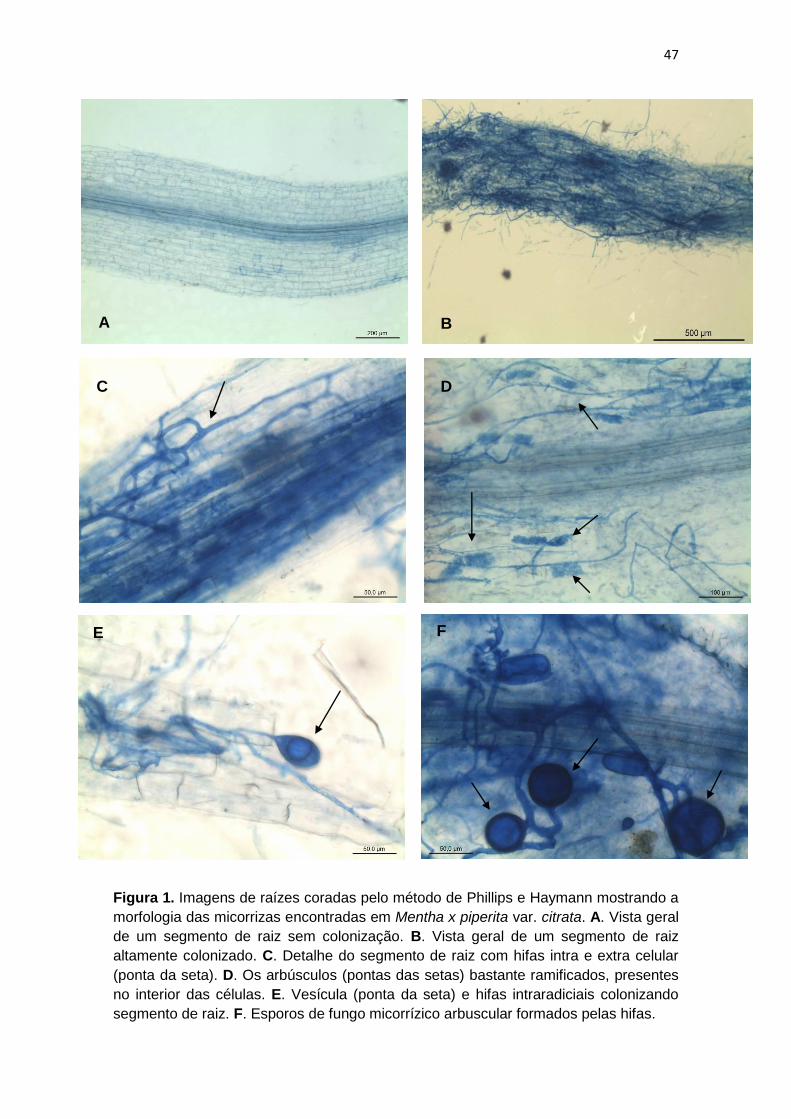
47
E F
A B
C D
Figura 1. Imagens de raízes coradas pelo método de Phillips e Haymann mostrando a
morfologia das micorrizas encontradas em Mentha x piperita var. citrata. A. Vista geral
de um segmento de raiz sem colonização. B. Vista geral de um segmento de raiz
altamente colonizado. C. Detalhe do segmento de raiz com hifas intra e extra celular
(ponta da seta). D. Os arbúsculos (pontas das setas) bastante ramificados, presentes
no interior das células. E. Vesícula (ponta da seta) e hifas intraradiciais colonizando
segmento de raiz. F. Esporos de fungo micorrízico arbuscular formados pelas hifas.
A B
C D
E F

48
Morfologicamente as micorrizas de Mentha x piperita var. citrata
apresentaram semelhança aos da série Arun (Fig. 1), a qual predomina na
maioria das angiospermas herbáceas (SMITH; SMITH, 1997). Nesse tipo
micorrízico ocorre frequentemente arbúsculos bem desenvolvidos, os quais são
as estruturas onde ocorre a troca de nutrientes entre o fungo e a planta
(SMITH; READ, 1997). Vesículas intracelulares, que são características da
subordem Glomineae (MORTON et al, 1996) também foram observadas (Fig.
1). Nessas vesículas ocorre o armazenamento de lipídios, fonte de reserva
para desenvolvimento do fungo micorrízico.
1.4 CONCLUSÕES
Os FMAs incrementaram a produção de biomassa seca da raiz, caule,
folha e total na dose de 60 mg dm-3 P adicionada ao solo.
Para a colonização micorrízica G. clarum apresentou a maior
porcentagem na maior dose, enquanto S. calospora obteve a maior
colonização na menor dose de P aplicada.
Os tratamentos micorrízicos e as doses de fósforo influenciaram na
absorção de macro e micronutrientes nas folhas de Mentha x piperita.
REFERÊNCIAS
BALOTA, E. L.; MACHINESKI, O.; TRUBER, P. V.; CEREZINI, P.; MILANI, K. L.; SCHERER, A.; HONDA, C.; LEITE, L. G. Efeito dos fungos micorrízicos arbusculares em culturas oleaginosas. Congresso brasileiro de mamona, 4 & simpósio internacional de oleaginosas energéticas, João Pessoa. Inclusão Social e Energia: Anais... Campina Grande: Embrapa Algodão, 2010. p. 680-684. BARBIERI JUNIOR, D.; BRAGA, L. F.; ROQUE, C. G.; SOUSA, M. P. Análise de crescimento de Hymenaea courbaril L. sob efeito da inoculação micorrizica e adubação fosfatada. Revista de Ciências Agro-Ambientais, v. 5, p. 1-15, 2007.

49
BERBARA, R.L.L.; SOUZA, F.A.; FONSECA, H.M.A.C. Fungos Micorrízicos Arbusculares: muito além da nutrição 2006. In: FERNANDES, M. S. Nutrição Mineral de Plantas. Viçosa, MG: Sociedade Brasileira de Ciências do Solo. 2006. 432 p. BURLEIGH, S. H.; BECHMANN, I. E. Plant nutrient transporter regulation in arbuscular mycorrhizas. Plant and Soil, v. 244, p. 247-251, 2002. BURLEIGH, S. H.; CAVAGNARO, T.; JAKOBSEN, I. Functional diversity of arbuscular mycorrhizas extends to the expression of plant genes involved in P nutrition. Journal of Experimental Botany, v. 53, p. 1593-1601, 2002. CARDOSO, E. J. B. N.; CARDOSO, I. M.; NOGUEIRA, M. A.; BARRETA, C. R. D. M.; de PAULA, A. M. Micorrizas Arbusculares na aquisição de nutrientes pelas plantas. In: SIQUEIRA, J. O.; de SOUZA, F. A.; CARDOSO, E. J. B. N.; TSAI, S. M. Micorrizas: 30 anos de pesquisas no Brasil, Lavras: UFLA, 2010. 716 p. CHAUDHARY, V.; KAPOOR, R.; BHATNAGAR, A. K. Effectiveness of two arbuscular mycorrhizal fungi on concentrations of essential oil and artemisinin in three accessions of Artemisia annua L. Applied Soil Ecology, v. 40, p. 174-181, 2008. COPETTA, A.; LINGUA, G.; BERTA, G. Effects of three AM fungi on growth, distribution of glandular hairs, and essential oil production in Ocimum basilicum L. var. Genovese. Mycorrhiza, v. 16, p. 485-494, 2006. CROTEAU , R.; KUTCHAN, T. M.; LEWIS, N. G. Natural Products (Secondary Metabolites) In: Buchanan, B.; Gruissem, W.; Jones, R. (Eds). Biochemistry & Molecular Biology of Plants. Rockville: Courier Companies, 2000, p. 1250-1318. EMBRAPA. Manual de métodos de análise de solo. Rio de Janeiro: Centro Nacional de Pesquisa em Solos, 1997. 212p. FERREIRA, D. F. Manual do sistema Sisvar para análises estatísticas. Lavras, Universidade Federal de Lavras, 2000. 66p. FREITAS M. S. M; MARTINS M. A.; CARVALHO A. J. C. Crescimento e composição da menta em resposta à inoculação com fungos micorrízicos arbusculares e adubação fosfatada. Horticultura Brasileira, v. 24, p.11-16, 2006. GARLET, T. M. B. Produtividade, teor e composição do óleo essencial de espécies de Mentha L. (Lamiaceae) cultivadas em hidroponia com variação de potássio. 113f. Tese de doutorado, 2007. Pós-Graduação em Agronomia, Universidade Federal de Santa Maria, RS. 113 folhas. 2007.

50
GEORGE, E. Nutrient uptake. In: KAPULNIK, Y. and DOUDS Jr., D.D. Arbuscular Mycorrhizas: Physiology and Function. Netherlands, 2000. p. 307-343. GERDEMANN, J. W.; NICOLSON, T. H. Spores of mycorrhizal edogone species extracted from soil by wet sieving and decanting. Transactions of British Mycological Society, v. 46, p. 235-44, 1963. GIOVANETTI, M.; MOSSE, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytologist, v. 84, p. 489-500, 1980. GUPTA, M. L.; PRASAD, A.; RAM, M.; KUMAR, S. Effect of tha vesicular-arbuscular mycorrhizal (VAM) fungus Glomus fasciculatum on the essential oil yield related characters and nutrient acquisition in the crops of different cultivars of menthol mint (Mentha arvensis) under fild conditions. Bioresource Technology, v. 81, p. 77-79, 2002. JOHNSON, N. C.; GRAHAM, J.H.; SMITH, F. A. Functioning of mycorrhizal associations along the mutualism-parasitism continuum. New Phytologist, v. 135, p. 575-586, 1997. KARAGIANNIDIS, N.; THOMIDIS, T.; LAZARI, D.; PANOU-FILOTHEOU, E.; KARAGIANNIDOU, C. Effect of three Greek arbuscular mycorrhizal fungi in improving the growth, nutrient concentration, and production of essential oils of oregano and mint plants. Scientia Horticulturae, v. 129, p. 329-334, 2011. KOTHARI, S.K.; MARSCHNER, H; GEORGE, E. Effect of VA mycorrhiza and rhizosphere microorganisms on root and shoot morphology, growth and water relations of mayse. New Phytologist, v. 116, p. 303-311. 1990. MARSCHNER, H. Mineral nutrition of higher plants. San Diego, Academic Press, 1995. 889p. McGONIGLE, T.P. MILLER, M.H.; EVANS, D.G.; FAIRCHILD, G.L. SWANA, J.A. New method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytologist, v. 115, p. 495–501, 1990. MORAIS, L. A. S. Influência dos fatores abióticos na composição química dos óleos essenciais. Horticultura Brasileira. 27: S4050-S4063, 2009. MOREIRA, F. M. S.; SIQUEIRA, J. O. Microbiologia e bioquímica do solo. Lavras, MG: Editora UFLA, 2006. 729p. MORTON, J. B. et al. Classification and identification of arbusculars mycorrhizal fungi. In: First ICOM Workshop, Canada, 1996. MURRAY, M. J.; LINCOLN, D. E. The genetic basis of acyclic oil constituents in Mentha citrata Ehrh. Genetics, v. 65, p. 457-471, 1970.

51
MELLONI, R.; NOGUEIRA, M. A.; FREIRE, V. F.; CARDOSO, E. J. B. N. Fósforo adicionado e fungos micorrízicos arbusculares no crescimento e nutrição mineral de limoeiro-cravo [Citrus limonia (L.) Osbeck] Revista Brasileira de Ciência do Solo, v. 24, p. 767-775, 2000. NELL, M.; VÖTSCH, M.; VIERHEILIG, H.; STEINKELLNER, S.; ZITTERL-EGLSEER, K.; FRANZA, C.; NOVAK, J. Effect of phosphorus uptake on growth and secondary metabolites of garden sage (Salvia officinalis L.) Journal Science Food Agriculturae, v. 89, p. 1090-1096, 2009. ORTAS, I. Effect of mycorrhiza application on plant growth and nutrient uptake in cucumber production under field conditions. Spanish Journal of Agricultural Research, v. 8 (S1), S116-S122, 2010. PHILLIPS, J.M.; HAYMAN, D.S. Improved procedures for clearing roots and staining parasitic and vesicular arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of British Mycological Society, v. 55, p. 158-161, 1970. RASOULI-SADAGHIANI, M.; HASSANI, A.; BARIN, M.; DANESH, Y. R.; SEFIDKON, F. Effects of arbuscular mycorrhizal (AM) fungi on growth, essential oil production and nutrients uptake in basil. Journal of Medicinal Plants Research, v. 4, p. 2222-2228, 2010. SAILO, G. L., BAGYARAJ, D. J. Influence of different AM-fungi on the growth, nutrition and forskolin content of Coleus forskohlii. Mycological Research, v. 109, p. 795-798, 2005. SMITH, S. E.; READ, D. J. Mycorrhizal Symbiosis, 3ª ed. New York: Academic Press, 2008. 800p. SMITH. F. A.; SMITH, S. E. Structural diversity in (vesicular) - arbuscular mycorrhizal symbioses. New Phytolologist, v. 137, p. 373-388, 1997.

52
2. Capítulo II - INFLUÊNCIA DE FUNGOS MICORRÍZICOS
ARBUSCULARES NO TEOR, RENDIMENTO E COMPOSIÇÃO DO ÓLEO
ESSENCIAL DE MENTHA X PIPERITA L. VAR. CITRATA (EHRH.) BRIQ.
CULTIVADA COM DUAS DOSES DE FÓSFORO
RESUMO
As plantas medicinais assim como os demais organismos recebem influencia de fatores externos e dentre estes a colonização por fungos micorrízicos arbusculares (FMAs) nas células radiculares é capaz de aumentar a sua produtividade. No caso das plantas aromáticas, os FMAs podem afetar principalmente o rendimento de óleo essencial, inclusive alterar a sua composição química. O objetivo do presente trabalho foi avaliar a influência dos fungos micorrízicos arbusculares e de duas doses de fósforo na produção do óleo essencial em Mentha x piperita L. var. citrata (Ehrh.) Briq. O experimento foi conduzido em casa de vegetação na Universidade Estadual de Santa Cruz em fatorial 5x2, sendo cinco tratamentos (controle não inoculado, inoculação de Acaulospora morrowiae Spain & Schenck, Glomus clarum Nicol. & Schenck, e Scutellospora calospora (Nicol. & Gerd.) Walk. & Sand., e a mistura desses inóculos) e duas doses de fósforo (60 mg e 120 mg de fósforo por dm3 de solo) com cinco repetições cada. Após 75 dias as plantas foram coletadas e avaliadas quanto a biomassa de folhas, teor, rendimento e composição do óleo essencial. A inoculação com FMAs aumentou o teor e o rendimento do óleo na menor dose de fósforo. Os inóculos utilizados separadamente foram mais eficientes do que a mistura dos fungos no incremento do teor e do rendimento do óleo essencial na dose maior de fósforo e a espécie G. clarum elevou significativamente o rendimento do óleo quando a dose de 120 mg dm-3 de P foi aplicada ao solo. A colonização com FMAs não alterou a composição do óleo essencial, que teve como componentes majoritários o linalol e o acetato de linalol, mas houve alterações quantitativas (percentuais) dos componentes do óleo essencial de Mentha x piperita var. citrata.
Palavras-chave: Acaulospora, Glomus, Scutellospora, Planta medicinais, Linalol, Acetato de linalol.

53
ABSTRACT
Medicinal plants as well as other plants receive influence of external factors and among these the colonization by arbuscular mycorrhizal fungi (AMF) in root cells is able to increase the productivity of plant and essential oil producing plants primarily affect the oil yield and in some cases can affect the chemical composition. The aim of this study was to evaluate the influence of mycorrhizal fungi and two phosphorus levels in the production of essential oil on Mentha x piperita L. var. citrata (Ehrh.) Briq. The experiment was conducted in a greenhouse at the Universidade Estadual de Santa Cruz in a 5x2 factorial design, with five treatments (uninoculated control, inoculation with Acaulospora morrowiae Spain & Schenck, Glomus clarum Nicol. & Schenck, e Scutellospora calospora (Nicol. & Gerd.) Walk. & Sand. and the mixture of inoculum) and two doses of phosphorus (60 mg and 120 mg of phosphorus per dm-3 soil) with five replicates each. After 75 days the plants were collected and evaluated for leaf biomass, and content, yield and essential oil composition. Inoculation with AMF increased the content and oil yield at the lowest dose of phosphorus. Inoculums used alone were more effective than the mixture of fungi in increasing the content and yield of essential oil in the higher dose of phosphorus, and G. clarum significantly increased oil yield when 120 mg dm-3 of P was applied to the soil. The mycorrhizal colonization did not alter the essential oil composition, which had as its major components linalool and linalool acetate, but there were quantitative changes (percentages) of essential oil components of Mentha x piperita var. citrata.
Keywords: Acaulospora, Glomus, Scutellospora, Medicinal plant, Linalool, Acetato de Linalool.

54
2.1 INTRODUÇÃO
O gênero Mentha é um dos mais importantes dentro família Lamiaceae,
e seu uso é amplamente difundido devido a sua adaptabilidade a diferentes
condições, ao ciclo vegetativo anual e à quantidade de informações já
existentes sobre suas características, desde anatômicas, bioquímicas,
taxonômicas e agronômicas. Espécies pertencentes a esse gênero enquadram-
se como excelentes produtoras de óleo essencial (MONTEIRO, 2009).
Dentre as espécies incluídas nesse gênero Mentha x piperita L. var.
citrata (Ehrh.) Briq. ou hortelã-limão, como é conhecida popularmente, vem
alcançando merecido destaque. Contrário a maioria das mentas, possui como
composto principal do seu óleo essencial o linalol e o acetato de linalila
(GARLET, 2007; MURRAY; LINCOLN, 1970). A presença desses compostos
aumenta o interesse econômico pela espécie, pois são utilizados nas indústrias
de perfumaria, cosmética, de alimentos, medicamentos, fragrâncias e tabaco,
principalmente como aromatizante e flavorizante de alimentos e na confecção
de perfumes (GARLET, 2007). Além disso, ecologicamente estes componentes
estão associados à defesa da planta contra patógenos e herbívoros, e à
atração de polinizadores (CROTEAU et al., 2000).
De toda a produção mundial de óleo essencial, as espécies do gênero
Mentha são as maiores contribuintes, geralmente sendo o mentol o composto
de maior interesse. No entanto, as espécies produtoras de linalol vem
ganhando mercado. Linalol é um dos compostos mais usados em fragrâncias e
estima-se que esteja presente em 60-90 % dos cosméticos existentes no
mercado (GARLET, 2007). Sua produção vem aumentando ano após ano,
devido a sua utilização não somente na indústria de perfumaria, mas também
na produção de vitaminas (STEFFANI et al., 2006).
A biossíntese de óleo essencial em plantas aromáticas é regulada por
diversos fatores, que podem ser genéticos, bioquímicos, fisiológicos e de
origem metabólica. A regulação fisiológica além de ser exercida por um padrão
de desenvolvimento específico é influenciada por fatores ambientais
(SANGWAN et al., 2001). Dentre estes fatores, pode-se ressaltar a interação
entre planta e fungos micorrízicos arbusculares (FMAs), que nas raízes de

55
plantas medicinais e aromáticas que pode influenciar na produção de princípios
ativos através da melhoria do estado nutricional, ou como resposta do fitobionte
ao fungo (MORAIS, 2009; VOLPIN et al., 1994).
A simbiose micorrízica arbuscular tem um importante potencial
biotecnológico e ecológico. Seu maior benefício diz respeito à absorção de
nutrientes no solo principalmente daqueles que possuem baixa mobilidade
como o fósforo, zinco e cobre (MOREIRA; SIQUEIRA, 2006). A ocorrência e o
funcionamento das micorrizas recebe influencia direta ou indireta de fatores
ambientais, podendo ser geralmente menor em condições de elevada
fertilidade pois a taxa de colonização micorrízica intrarradical apresenta uma
diminuição drástica com altos níveis de fósforo no solo (MOREIRA; SIQUEIRA,
2006; CARDOSO et al., 2010).
Dentre os fatores que podem interferir no teor e no rendimento do óleo
essencial em plantas medicinais, a micorrização merece destaque, pois
participa diretamente do processo de captação de nutrientes cuja deficiência ou
excesso pode intervir na quantidade e qualidade do óleo (MORAIS, 2009). O
objetivo do presente estudo foi avaliar o efeito dos fungos micorrízicos
arbusculares sob duas doses de fósforo no teor, rendimento e composição do
óleo essencial de Mentha x piperita var. citrata.
2.2 MATERIAL E MÉTODOS
O experimento foi conduzido em casa de vegetação no campus da
Universidade Estadual de Santa Cruz, Ilhéus – BA, sob condições naturais de
luminosidade e temperatura. O delineamento experimental foi constituído por
um fatorial 5x2, sendo utilizados três inóculos de FMA: Acaulospora morrowiae
(Am) Spain & Schenck (A79 CNPAB 037), Glomus clarum (Gc) Nicol. &
Schenck (A5 CNPAB 005) e Scutellospora calospora (Sc) (Nicol. & Gerd.)
Walk. & Sand. (A80 CNPAB 038). além de um controle formado por plantas
não inoculadas (C) e um tratamento com a mistura de todos os três inóculos
(M), perfazendo um total de cinco tratamentos micorrízicos, e duas doses
diferentes de fósforo 60 mg (P60) e 120 (P120) mg dm-3 com cinco repetições
cada. A unidade experimental foi composta por um vaso de plástico de cinco

56
litros contendo quatro dm3 de solo e uma planta por vaso. Os inóculos foram
cedidos pela Coleção de Fungos Micorrízicos da Embrapa Agrobiologia
(Seropédica, RJ).
O solo utilizado no experimento foi um argissolo amarelo distrófico típico,
coletado da camada de 0 a 20 cm em área de pastagem de Brachiaria
humidicola com remanescente de dendê na Estação Experimental do Almada
(39º10’ W e 14º38’ S), localizada em Uruçuca (BA) e pertencente à
Universidade Estadual de Santa Cruz. O solo foi seco ao ar e tamisado em
peneira com malha de 5 mm e suas características químicas (Tabela 1) e
físicas (Tabela 2) foram determinados (Embrapa, 1997). Esse solo foi
autoclavado por uma hora a 121 °C e 1,5 atm por dois dias consecutivos para
esterilização.
Com o solo coletado foi realizada uma curva de incubação usando cinco
doses de calcário dolomítico com PRNT de 82,02 %. Os resultados foram
analisados por regressão para ajustar o pH em torno de 6,5. Além do calcário,
que também serviu de fonte para o cálcio e o magnésio, o solo contido nos
vasos recebeu uma adubação individual com base na análise do solo, na
produtividade da espécie e no comportamento do fungo, constituída por 100
mg dm-3 de nitrogênio na forma de sulfato de amônio; duas doses de fósforo 60
e 120 mg dm-3 como superfosfato simples; 80 mg dm-3 de potássio sob a forma
de cloreto de potássio; 1,55 mg dm-3 de ferro na forma de Ferro EDTA; 0,75 mg
dm-3 de boro na forma de ácido bórico; 1,33 mg dm-3 de cobre sob a forma de
sulfato de cobre; 0,15 mg dm-3 de molibdênio na forma de molibdato de amônio;
4 mg dm-3 de zinco e de manganês na forma de sulfato de zinco e sulfato de
manganês, respectivamente. A adubação foi subdividida em duas partes, uma
anterior a autoclavagem onde foi adicionado ao solo o calcário, o superfosfato
simples, cloreto de potássio em solução e os micronutrientes (boro, cobre,
molibdênio, zinco e manganês) também em solução e outra posterior à
autoclavagem que recebeu a adição do nitrogênio sob forma de solução
preparada com água autoclavada e o ferro, também em solução que foi
autoclavada separada. Antes da transferência do solo para os vasos, estes
foram lavados com água e desinfestados superficialmente com etanol 70 % e
com hipoclorito de sódio comercial (2 a 2,5 %).
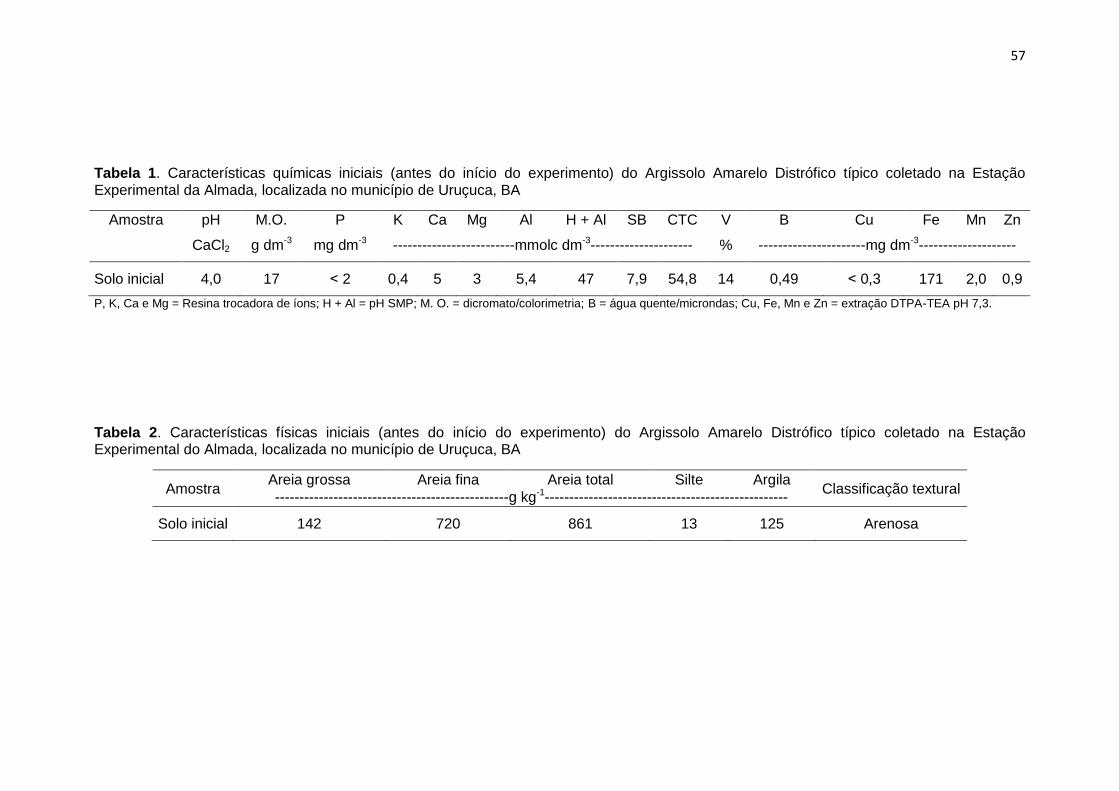
57
Tabela 1. Características químicas iniciais (antes do início do experimento) do Argissolo Amarelo Distrófico típico coletado na Estação Experimental da Almada, localizada no município de Uruçuca, BA
Amostra pH M.O. P K Ca Mg Al H + Al SB CTC V B Cu Fe Mn Zn
CaCl2 g dm-3 mg dm-3 -------------------------mmolc dm-3--------------------- % ----------------------mg dm-3--------------------
Solo inicial 4,0 17 ˂ 2 0,4 5 3 5,4 47 7,9 54,8 14 0,49 ˂ 0,3 171 2,0 0,9
P, K, Ca e Mg = Resina trocadora de íons; H + Al = pH SMP; M. O. = dicromato/colorimetria; B = água quente/microndas; Cu, Fe, Mn e Zn = extração DTPA-TEA pH 7,3.
Tabela 2. Características físicas iniciais (antes do início do experimento) do Argissolo Amarelo Distrófico típico coletado na Estação Experimental do Almada, localizada no município de Uruçuca, BA
Amostra Areia grossa Areia fina Areia total Silte Argila
Classificação textural ------------------------------------------------g kg-1--------------------------------------------------
Solo inicial 142 720 861 13 125 Arenosa

58
A propagação da Mentha x piperita L. var. citrata foi realizada por via
vegetativa. Estacas apicais com aproximadamente 5 cm de comprimento foram
obtidas junto ao Horto de Plantas Medicinais da Universidade Estadual de Santa
Cruz, Ilhéus-BA, e colocadas para enraizar em bandejas de polipropileno com
substrato areia e vermiculita (2:1) autoclavado por uma hora a 121 °C e 1,5 atm,
onde permaneceram por quatorze dias até o transplante.
No transplante, os inóculos de FMAs foram concomitantemente adicionados
ao solo junto das raízes e, por fim, pedriscos autoclavados por 20 minutos a 121 °C
e 1,5 atm foram colocados para diminuir a superfície de contato do solo com o ar e
minimizar as possíveis contaminações. Durante a condução do experimento as
plantas foram irrigadas com água deionizada autoclavada.
2.2.1 Determinação da biomassa
Após 75 dias de cultivo em casa de vegetação as folhas das plantas foram
coletadas e secas a 40 °C em estufa de circulação forçada e posteriormente foram
pesadas em balança semi-analítica até peso constante para a obtenção da
biomassa foliar, utilizada para o cálculo do rendimento.
2.2.2 Determinação do teor, rendimento e composição química do óleo
essencial
O óleo essencial foi extraído com aparelho de Clevenger modificado utilizando
de 2 a 3,6 g de biomassa seca foliar em 400 ml de água por uma hora com quatro
repetições por tratamento, proveniente de uma amostragem composta de todas as
repetições referente a cada tratamento.
O teor do óleo foi determinado com base no peso do óleo extraído do material
vegetal seco (% m/m) em balança analítica. O rendimento do óleo foi calculado por
meio do teor multiplicado pelo valor médio de biomassa seca foliar da planta (mg por
planta).

59
Os óleos foram caracterizados quimicamente pelo índice de refração (IR), e
posteriormente por cromatografia gasosa acoplada a um espectrômetro de massa
de alta resolução (CGAR). Os diversos constituintes químicos dos óleos essenciais
foram identificados por meio da comparação computadorizada com a biblioteca do
aparelho, literatura e índice de retenção de Kovats (ADAMS, 1995). Os índices de
retenção de Kovats (IK) foram calculados através da injeção de uma série de
padrões de n-alcanos (C8-C26) injetados nas mesmas condições cromatográficas
das amostras.
2.2.3 Análise estatística
Todos os dados foram submetidos à análise de variância, os tratamentos
fosfatados foram comparados pelo teste de F a 5% de probabilidade. Para as
médias dos tratamentos fúngicos foi utilizado o teste t de contrastes ortogonais
(Tabela 3) a 5% de probabilidade. No caso de significância da interação FMA x dose
de fósforo foi realizado o seu desdobramento. As análises estatísticas foram
realizadas pelo programa estatístico SISVAR (FERREIRA, 2000).
Tabela 3. Descrição dos contrastes ortogonais para a avaliação da influencia de FMAs em Mentha x piperita var. citrata submetida a duas doses de fósforo
Contraste ortogonais Comparação
Ŷ1 = 4 1m̂ - 2m̂ - 3m̂ - 4m̂ - 5m̂ Controle vs. Demais - contraste um (1)
Ŷ2 = 2m̂ + 3m̂ + 4m̂ - 3 5m̂ A. morrowiae, G. clarum, S. calospora vs.
Mistura dos inóculos - contraste dois (2)
Ŷ3 = 2m̂ + 4m̂ - 2 3m̂ A. morrowiae e S. calospora vs. G. clarum -
contraste três (3)
Ŷ4 = 2m̂ - 4m̂ A. morrowiae vs. S. calospora - contraste
quatro (4)
Ŷ5 = 4 4m̂ - 1m̂ - 2m̂ - 3m̂ - 5m̂ S. calospora vs. demais – contraste (5)
Ŷ6 = 3 4m̂ - 2m̂ - 3m̂ - 5m̂ S. calospora vs. A. morrowiae, G. clarum e
mistura dos inóculos – contraste (6)
Ŷ7 = 2m̂ - 3m̂ A. morrowiae vs. G. clarum – contraste (7)
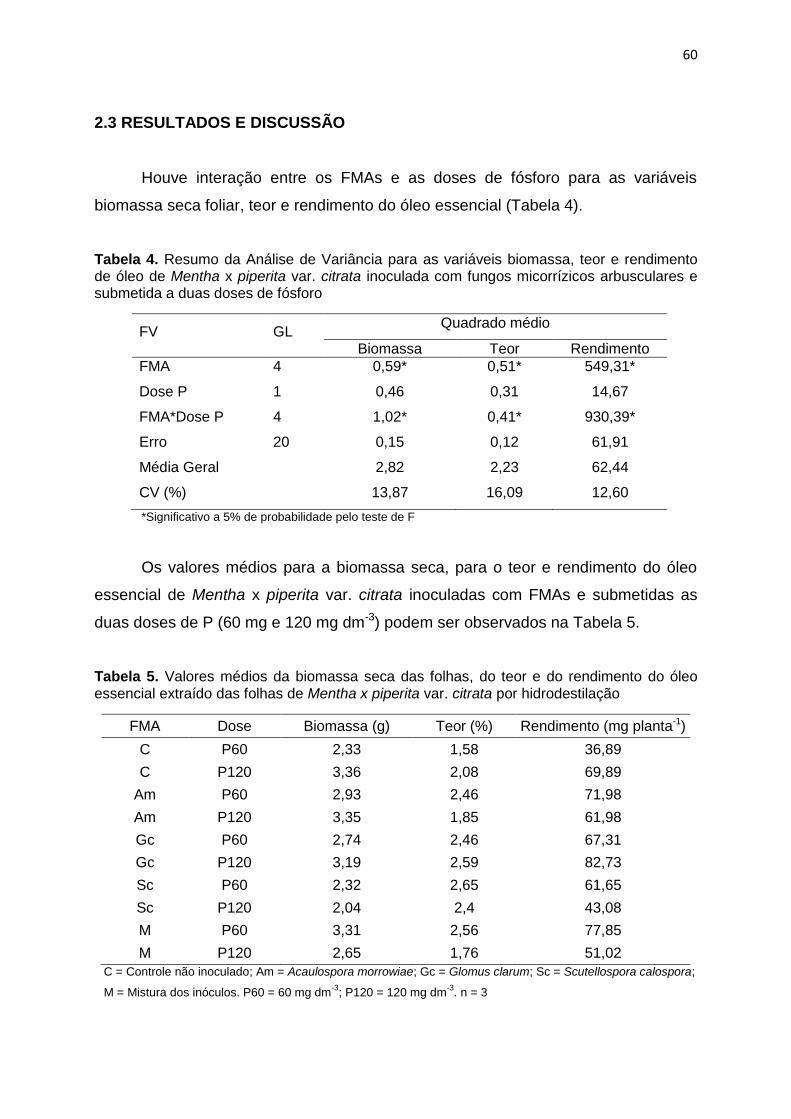
60
2.3 RESULTADOS E DISCUSSÃO
Houve interação entre os FMAs e as doses de fósforo para as variáveis
biomassa seca foliar, teor e rendimento do óleo essencial (Tabela 4).
Tabela 4. Resumo da Análise de Variância para as variáveis biomassa, teor e rendimento de óleo de Mentha x piperita var. citrata inoculada com fungos micorrízicos arbusculares e submetida a duas doses de fósforo
FV GL Quadrado médio
Biomassa Teor Rendimento
FMA 4 0,59* 0,51* 549,31*
Dose P 1 0,46 0,31 14,67
FMA*Dose P 4 1,02* 0,41* 930,39*
Erro 20 0,15 0,12 61,91
Média Geral 2,82 2,23 62,44
CV (%) 13,87 16,09 12,60
*Significativo a 5% de probabilidade pelo teste de F
Os valores médios para a biomassa seca, para o teor e rendimento do óleo
essencial de Mentha x piperita var. citrata inoculadas com FMAs e submetidas as
duas doses de P (60 mg e 120 mg dm-3) podem ser observados na Tabela 5.
Tabela 5. Valores médios da biomassa seca das folhas, do teor e do rendimento do óleo essencial extraído das folhas de Mentha x piperita var. citrata por hidrodestilação
FMA Dose Biomassa (g) Teor (%) Rendimento (mg planta-1)
C P60 2,33 1,58 36,89
C P120 3,36 2,08 69,89
Am P60 2,93 2,46 71,98
Am P120 3,35 1,85 61,98
Gc P60 2,74 2,46 67,31
Gc P120 3,19 2,59 82,73
Sc P60 2,32 2,65 61,65
Sc P120 2,04 2,4 43,08
M P60 3,31 2,56 77,85
M P120 2,65 1,76 51,02
C = Controle não inoculado; Am = Acaulospora morrowiae; Gc = Glomus clarum; Sc = Scutellospora calospora;
M = Mistura dos inóculos. P60 = 60 mg dm-3
; P120 = 120 mg dm-3
. n = 3

61
De acordo com as estimativas dos contrastes (Tabela 6), na variável
biomassa seca, dentro da menor dose de P (60 mg dm-3), a mistura dos inóculos foi
significativamente superior à utilização dos inóculos separados (contraste dois) e o
tratamentos com A. morrowiae, G. clarum e a mistura dos inóculos foi superior ao
tratamento com S. calospora (contraste seis) . Já na maior dose de fósforo (120 mg
dm-3), o contraste um (comparação entre o controle e os demais tratamentos), o
contraste quatro (comparação entre o A. morrowiae e S. calospora), o cinco
(comparação entre o FMA S. calospora versus os demais tratamentos) e o contraste
seis (que compara o S. calospora com os tratamentos micorrizados) apresentaram
diferença significativa, mostrando que o controle apresentou massa seca superior
aos tratamentos micorrizados, e que o FMA S. calospora obteve resultado inferior
para esse parâmetro.
Para o teor as estimativas de contrastes (Tabela 6) apresentaram
significância apenas para o contraste um na dose de 60 mg de fósforo por dm3 que
compara o controle não inoculado com os demais tratamentos micorrizados, e nos
contrastes dois e sete na dose de 120 mg de fósforo, a primeira comparação, que
corresponde ao contraste dois, é entre os inóculos utilizados separados versus a
mistura de todos os inóculos e o contraste sete compara o FMA A. morrowiae com
G. clarum. Sendo assim, os tratamentos micorrizados se mostraram mais eficientes
na produção de óleo, comparado ao controle não micorrizado, dentro da menor dose
de fósforo (60 mg dm-3 de P), e na maior dose de fósforo (120 mg dm-3) para esse
mesmo parâmetro, os inóculos utilizados separadamente foram mais promissores do
que o tratamento com a mistura dos três inóculos, e G. clarum foi superior a A.
morrowiae.

62
Tabela 6. Estimativas dos contrastes ortogonais para teor e rendimento do óleo essencial de Mentha x piperita var. citrata inoculada com fungos micorrízicos arbusculares e submetida a duas doses de fósforo
Contrastes Biomassa Teor Rendimento
P60 P120 P60 P120 P60 P120
(1) C vs. Demais -0,45 0,73* - 0,94* -0,07 - 32,80* 10,18
(2) Am, Gc e Sc vs. M -0,88* 0,42 - 0,03 0,51* -10,86 11,57*
(3) Am e Sc vs. Gc 0,17 -0,43 0,09 - 0,46 -0,493 - 30,20*
(4) Am vs. Sc 0,60 1,02* - 0,19 - 0,55 10,32 18,89*
(5) Sc vs. demais -0,46 -0,87* 0,38 0,32 -1,85 -23,32*
(6) Sc vs. Am, Gc e M -0,61* -0,73* 0,16 0,33 -10,72 -22,16*
(7) Am vs. Gc 0,48 0,07 0 -0,74* 4,67 -20,75*
C = Controle não inoculado; Am = Acaulospora morrowiae; Gc = Glomus clarum; Sc = Scutellospora calospora;
M = Mistura dos inóculos; P60 = 60 mg dm-3
de P; P120 = 120 mg dm-3
. *significativo a 5 % de probabilidade do
teste t.
Alguns trabalhos relatam a influência dos FMA no teor do óleo essencial,
como os realizado por Freitas et al. (2004a) com M. arvensis, e Gupta et al. (2002)
que avaliaram o comportamento de três cultivares de Mentha, e Freitas et al.
(2004b), em seu trabalho com carqueja no qual apesar de não apresentar interação
entre FMA e adubação, a presença de micorrização aumentou a produção de fenóis
totais na carqueja. De uma maneira geral, a colonização de plantas medicinais por
FMA aumenta o teor do óleo essencial, pois os fungos podem majorar as
concentrações dos metabólitos secundários. A frequência e a intensidade com que
isso ocorre dependem muito da interação planta – fungo, como relatado por
Chaudhary et al. (2008) em Artemisia annua, a qual teve o teor do composto
majoritário alterado, sendo que a combinação espécie vegetal e a linhagem de FMA
também provocou variação no metabólito secundário.
Para o rendimento do óleo essencial em M. x piperita var. citrata as
estimativas (Tabela 6) demonstram diferença significativa apenas no contraste um
(comparação entre o controle versus os tratamentos micorrizados) para a menor
dose de fósforo (60 mg dm-3). Esse resultado significa que os tratamentos

63
micorrizados elevaram a produção do óleo essencial quando comparados com o
controle.
Na maior dose de P (120 mg dm-3) a significância para o rendimento do óleo
das folhas M. x piperita var. citrata ocorre em praticamente todas as estimativas dos
contrastes, com exceção do contraste um. No contraste dois percebe-se que a
mistura dos inóculos foi menos eficiente em promover o aumento no rendimento do
óleo essencial do que os inóculos de FMAs utilizados separadamente. Já o contraste
três, evidencia que G. clarum incrementou o rendimento do óleo de M. x piperita var.
citrata em comparação aos outros dois inóculos e mais especificamente ao de A.
morrowiae como elucida o contraste sete. Esse comportamento está em
consonância com os dados da colonização micorrízica, onde tratamento micorrizado
com G. clarum na maior dose de fósforo obteve a maior porcentagem de
colonização (ver Capítulo 1).
Os contrastes quatro, cinco e seis, que foram significativos, demonstram que
o menor rendimento do óleo essencial foi obtido no tratamento inoculado com S.
calospora. Essa tendência de baixo rendimento no óleo foi devida a pequena
produção de biomassa das plantas inoculadas com S. calospora.
O acréscimo no rendimento do óleo essencial deve-se geralmente ao
aumento da produção de biomassa, ocasionado neste caso pela melhoria do estado
nutricional da planta hospedeira em função da micorrização. Pesquisa desenvolvida
por Kapoor et al. (2004) mostrou que Foeniculum vulgare Mill. teve o rendimento do
óleo incrementado em todos os níveis de fósforo utilizados para as duas espécies de
Glomus inoculadas.
Foram identificados treze componentes para o óleo essencial de M. x piperita
var. citrata, e apenas dois compostos não foram determinados (Tabela 7). Os
componentes majoritários para todos os tratamentos foram linalol e acetato de
linalol, confirmando as informações encontradas na literatura (MURRAY; LINCOLN,
1970; GARLET, 2007), onde M. x piperita é tida como espécie promissora na
produção de linalol. Além desses componentes outros nove foram identificados com
porcentagens menos expressivas, foram eles: cis-3- pinanona, α- terpineol, nerol,
acetado de nerila, acetato de geranila, cariofileno, γ-gurjuneno, elemol e guaiol.
Na menor dose de fósforo (60 mg dm-3) o tratamento com Glomus clarum
obteve a maior porcentagem de linalol (51 %) em contrapartida apresentou a menor

64
porcentagem de acetato de linalol (24,5 %). Para a maior dose de fósforo (120 mg
dm-3) foi o tratamento com Scutellospora calospora quem obteve a maior
porcentagem de linalol (48 %) e Glomus clarum apresentou a maior porcentagem de
acetato de linalol (37 %).
A maioria dos trabalhos não relata mudança qualitativa na composição do
óleo essencial, mas sim uma alteração quantitativa na porcentagem de alguns
componentes, como foi observado por Copetta et al. (2007) utilizando inóculos de
Gigaspora rosea e G. margarita. Zubek et al. (2010) que detectou uma mudança no
teor de derivados de timol. Karagiannidis et al. (2011) com mudanças nos
componentes principais de espécies de menta e orégano.
Além da presença dos FMAs a adubação fosfatada exerce forte influência
sobre o teor rendimento e composição do óleo essencial como foi proposto por Nell
et al. (2009) e Freitas et al. (2004). No primeiro caso plantas submetidas à adubação
fosfatada tiveram um aumento na concentração de fenóis totais e no segundo caso
os fungos micorrízicos arbusculares e as doses de fósforo influenciaram na
qualidade do óleo com alterações em seu composto majoritário. Sem adubação
fosfatada os FMAs foram benéficos à produção e qualidade do óleo, e com
adubação houve uma tendência à redução do teor e dos componentes majoritários
(FREITAS et al., 2004).
Os mecanismos utilizados pelo fungo que provocam as mudanças nas
concentrações dos compostos fitoquímicos podem ser multidirecionais e não estão
ainda elucidados (TOUSSAINT et al., 2007). Em geral a produção de óleo essencial
é considerada como uma resposta de defesa da planta à colonização pelo FMA, e a
modificação nessa produção pode ser em consequência de mecanismo de
sinalização entre os simbiontes (COPETTA et al., 2006; TOUSSAINT et al., 2007).
O aumento na produção de metabólitos secundários pode envolver vários
processos, e isso pode ser mediado pelo aumento na nutrição de fósforo e de
nitrogênio, por exemplo, causado pelo FMA (KAPOOR et al., 2002; TOUSSAINT et
al., 2007). Além disso, a presença da micorriza pode causar mudanças no nível de
fitormonios do vegetal (COPETTA et al., 2006; KAPOOR et al., 2007).
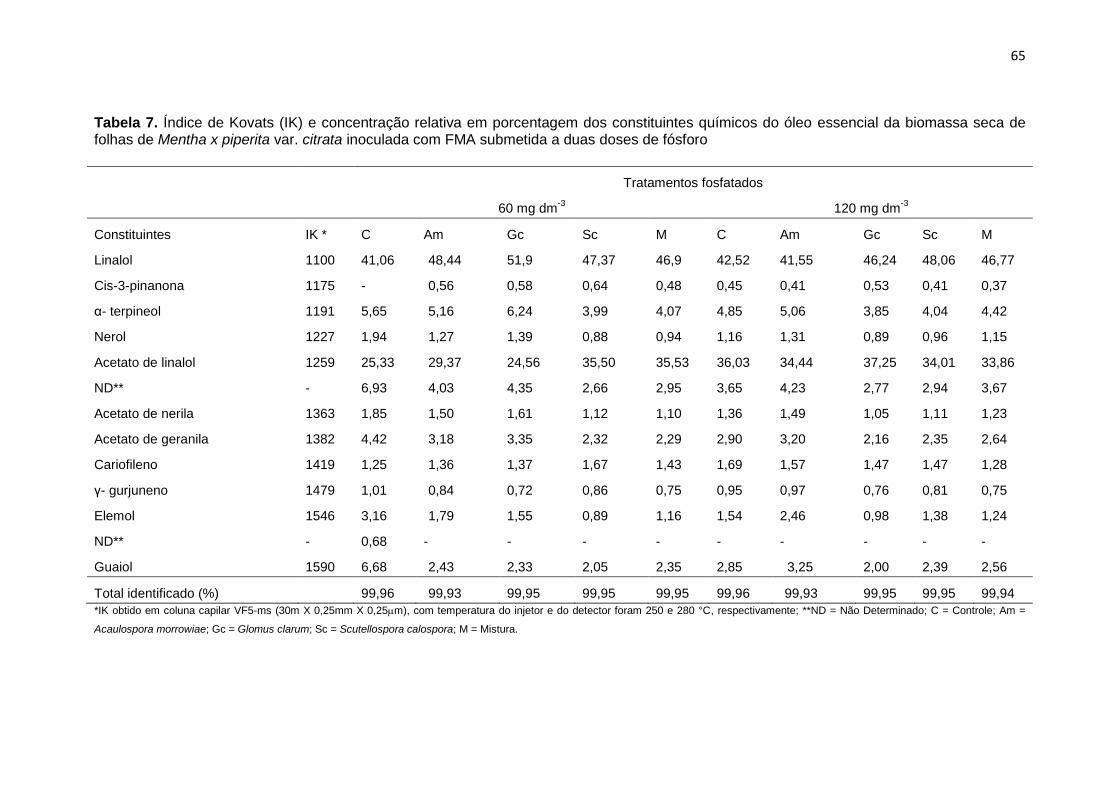
65
Tabela 7. Índice de Kovats (IK) e concentração relativa em porcentagem dos constituintes químicos do óleo essencial da biomassa seca de folhas de Mentha x piperita var. citrata inoculada com FMA submetida a duas doses de fósforo
Tratamentos fosfatados
60 mg dm-3
120 mg dm-3
Constituintes IK * C Am Gc Sc M C Am Gc Sc M
Linalol 1100 41,06 48,44
51,9 47,37 46,9 42,52 41,55 46,24 48,06 46,77
Cis-3-pinanona 1175 - 0,56
0,58 0,64 0,48 0,45 0,41 0,53 0,41 0,37
α- terpineol 1191 5,65 5,16
6,24 3,99 4,07 4,85 5,06 3,85 4,04 4,42
Nerol 1227 1,94 1,27
1,39 0,88 0,94 1,16 1,31 0,89 0,96 1,15
Acetato de linalol 1259 25,33 29,37
24,56 35,50 35,53 36,03 34,44 37,25 34,01 33,86
ND** - 6,93 4,03
4,35 2,66 2,95 3,65 4,23 2,77 2,94 3,67
Acetato de nerila 1363 1,85 1,50
1,61 1,12 1,10 1,36 1,49 1,05 1,11 1,23
Acetato de geranila 1382 4,42 3,18
3,35 2,32 2,29 2,90 3,20 2,16 2,35 2,64
Cariofileno 1419 1,25 1,36
1,37 1,67 1,43 1,69 1,57 1,47 1,47 1,28
γ- gurjuneno 1479 1,01 0,84
0,72 0,86 0,75 0,95 0,97 0,76 0,81 0,75
Elemol 1546 3,16 1,79
1,55 0,89 1,16 1,54 2,46 0,98 1,38 1,24
ND** - 0,68 - - - - - - - - -
Guaiol 1590 6,68 2,43
2,33 2,05 2,35 2,85 3,25 2,00 2,39 2,56
Total identificado (%) 99,96 99,93
99,95 99,95 99,95 99,96 99,93
99,95 99,95 99,94
*IK obtido em coluna capilar VF5-ms (30m X 0,25mm X 0,25m), com temperatura do injetor e do detector foram 250 e 280 °C, respectivamente; **ND = Não Determinado; C = Controle; Am =
Acaulospora morrowiae; Gc = Glomus clarum; Sc = Scutellospora calospora; M = Mistura.

66
2.4 CONCLUSÕES
Os fungos micorrízicos arbusculares aumentaram o teor e o rendimento
do óleo essencial de Mentha x piperita L. var. citrata quando o solo foi adubado
com 60 mg dm-3 de fósforo.
Os fungos inoculados separadamente foram mais eficientes do que o
tratamento com a mistura das espécies de FMAs em promover o incremento no
teor e no rendimento do óleo na dose 120 mg dm-3 de fósforo.
O fungo Glomus clarum aumentou o rendimento do óleo Mentha x
piperita L. var. citrata com 120 mg dm-3 de fósforo.
O óleo essencial teve como componentes majoritários o linalol e o
acetato de linalol, havendo alteração no percentual dos componentes, mas não
na composição do óleo.
2.5 REFERÊNCIAS
ADAMS, R.P. Identification of essential oil components by gas chromatography and mass spectroscopy. Illinois: Allured, 1995. 698p. CARDOSO, E. J. B. N.; CARDOSO, I. M.; NOGUEIRA, M. A.; BARRETA, C. R. D. M.; de PAULA, A. M. Micorrizas Arbusculares na aquisição de nutrientes pelas plantas. In: SIQUEIRA, J. O.; de SOUZA, F. A.; CARDOSO, E. J. B. N.; TSAI, S. M. Micorrizas: 30 anos de pesquisas no Brasil. Lavras: UFLA, 2010. 716 p. CHAUDHARY, V.; KAPOOR, R.; BHATNAGAR, A. K. Effectiveness of two arbuscular mycorrhizal fungi on concentrations of essential oil and artemisinin in three accessions of Artemisia annua L. Applied Soil Ecology, v.40, p. 174-181, 2008. COPETTA, A.; LINGUA, G.; BARDI, L.; MASOERO, G.;BERTA, G. Influence of arbuscular mycorrhizal fungi on growth and essential oil composition in Ocimum basilicum var. Genovese. Caryologia. Vol. 60, n°. 1-2, p. 106-110, 2007. COPETTA, A.; LINGUA, G.; BERTA, G. Effects of three AM fungi on growth, distribution of glandular hairs, and essential oil production in Ocimum basilicum L. var. Genovese. Mycorrhiza, v. 16, p. 485-494, 2006.

67
CROTEAU , R.; KUTCHAN, T. M.; LEWIS, N. G. Natural Products (Secondary Metabolites) In: BUCHANAN, B.; GRUISSEM, W.; JONES, R. (Eds). Biochemistry & Molecular Biology of Plants. Rockville: Courier Companies, 2000, p. 1250-1318. FERREIRA, D. F. Manual do sistema Sisvar para análises estatísticas. Lavras, Universidade Federal de Lavras, 2000. 66p. FREITAS, M. S. M.; MARTINS, M. A.; VIEIRA, I. J. C. Produção e qualidade de óleos essenciais de Mentha arvensis em resposta à inoculação de fungos micorrízicos arbusculares. Pesquisa agropecuária Brasileira, Brasília, v. 39, n°9, p.887-894, set. 2004a. FREITAS, M. S. M.; MARTINS, M. A.; CARVALHO, A. J. C.; CARNEIRO, R.F.V. Crescimento e Produção de fenóis totais em carqueja [Baccharis trimera (Less.) DC.] em resposta à inoculação com fungos micorrízicos arbusculares, na presença e na ausência de adubação mineral. Revista Brasileira de Plantas Medicinais, Botucatu, v.6, n. 3, p.30-34, 2004b. GARLET, T. M. B. Produtividade, teor e composição do óleo essencial de espécies de Mentha L. (Lamiaceae) cultivadas em hidroponia com variação de potássio. 113f. Tese de doutorado, 2007. Pós-Graduação em Agronomia, Universidade Federal de Santa Maria, RS. 113 f. 2007. GUPTA, M.L.; PRASAD, A.; RAM, M.; KUMAR, S. Effect of tha vesicular-arbuscular mycorrhizal (VAM) fungus Glomus fasciculatum on the essential oil yield related characters and nutrient acquisition in the crops of different cultivars of menthol mint (Mentha arvensis) under fild conditions. Bioresource Technology, v. 81 p. 77 – 79, 2002. KAPOOR, R.; GIRI, B.; MUKERJI, K. G. Improved growth and essential oil yield and quality in Foeniculum vulgare mill on mycorrhizal inoculation supplemented with P-fertilizer. Bioresource Technology, v. 93 p. 307 – 311, 2004. KAPOOR, R.; GIRI, B.; MUKERJI, K.G. Mycorrhization of coriander (Coriandrum sativum L.) to enhance the concentration and quality of essential oil. Journal of the Science of Food and Agriculture, v. 82, p. 339-342, 2002. KARAGIANNIDIS, N; THOMIDIS, T; LAZARI, D.; PANOU-FILOTHEOU, E.; KARAGIANNIDOU, C. Effect of three Greek arbuscular mycorrhizal fungi in improving the growth, nutrient concentration, and production of essential oils of oregano and mint plants. Scientia Horticulturae, v. 129, p. 329 – 334, 2011. MONTEIRO, R. Desenvolvimento de menta e produção de óleo essencial sob diferentes condições de manejo. 81f. Dissertacao (Mestrado em Agronomia) Ciencias Agrarias, Universidade Federal do Parana. Curitiba, 2009.

68
MORAIS L. A. S. Influência dos fatores abióticos na composição química dos óleos essenciais. Horticultura Brasileira v. 27, p. S4050 - S4063, 2009. MOREIRA, F.M.S.; SIQUEIRA, J.O. Microbiologia e bioquímica do solo. Lavras, MG: Editora UFLA, 2006. 729p. MURRAY, M.J.; LINCOLN, D. E. The genetic basis of acyclic oil constituents in Mentha citrate Ehrh. Genetics, v. 65, p. 457-471, 1970. NELL, M.; VÖTSCH, M.; VIERHEILIG, H.; STEINKELLNER, S.; ZITTERL-EGLSEER, K.; FRANZA, C.; NOVAK, J. Effect of phosphorus uptake on growth and secondary metabolites of garden sage (Salvia officinalis L.) Journal Science Food Agriculturae, v. 89, p. 1090-1096, 2009. SANGWAN, N. S.; FAROOQI, A.H.A.; SHABIH, F.; SANGWAN, R. S. Regulation of essential oil production in plants. Plant Growth Regulation, v. 34, p. 3–21, 2001. STEFFANI, E.; ATTI-SANTOS, A. C.; ATTI-SERAFINI, L.; PINTO, L. T. Extraction of ho-sho (Cinnamomum camphora Nees and Eberm var. linaloolifera fujita) essential oil with supercritical CO2: experiments and modeling. Brazilian Journal of Chemical Engineering. v. 23, n. 02, p. 259 - 266, April - June, 2006. TOUSSAINT, J. P.; SMITH, F. A.; SMITH, S. E. Arbuscular mycorrhizal fungi can induce the production of phytochemicals in sweet basil irrespective of phosphorus nutrition. Mycorrhiza, v. 17, p. 291–297, 2007. VOLPIN, H.; ELKIND, Y.; OKON, Y.; KAPULNIK, Y. A vesicular arbuscular mycorrhizal fungus (Glomus intraradix) induces a defense response in alfafa roots. Plant Physiology, v. 104, p. 683-689, 1994. ZUBEK, S.; STOJAKOWSKA, A.; ANIELSKA, T.; TURNAU, K. Arbuscular mycorrhizal fungi alter thymol derivative contents of Inula ensifolia L. Mycorrhiza, v. 20, p. 497–504, 2010.

69
CONSIDERAÇÕES FINAIS
A associação simbiótica entre FMAs e o sistema radicular de organismos
vegetais é um dos fatores que podem interferir no desenvolvimento das
plantas, normalmente a micorrização aumenta a produtividade do fitobionte, em
plantas produtoras de óleo essencial, além desse benefício, a simbiose pode
influir no rendimento e no teor do óleo essencial. De maneira semelhante o
estabelecimento e a propagação dos FMAs são influenciados por fatores
edáficos, como por exemplo, a disponibilidade de fósforo, o que
consequentemente altera o comportamento do vegetal. Ambas as fontes de
variação (inoculação de FMAs e doses de P) influenciaram de alguma maneira
nos parâmetros avaliados. Tanto os tratamentos fúngicos como as doses de
fósforo influenciaram no teor de macro e micronutrientes nas folhas de Mentha
x piperita var. citrata.
Na menor dose de fósforo (60 mg dm-3), as plantas dos tratamentos
inoculados com FMAs apresentaram maior biomassa (raiz, caule, folha e total),
e maior teor e o rendimento do óleo essencial. Na maior dose de fósforo (120
mg dm-3) tanto o teor como o rendimento do óleo essencial de Mentha x
piperita var. citrata foram mais altos quando os inóculos de FMAs foram
utilizados separadamente.
A interação planta – microrganismo depende de características
intrínsecas inerentes a cada espécie envolvida na interação, os diferentes
FMAs tiveram comportamento distinto em cada dose de fósforo adicionada ao
solo. A inoculação com S. calospora obteve a maior colonização, na menor
dose de P (60 mg dm-3), enquanto que o FMA G. clarum apresentou a
porcentagem de colonização mais expressiva na maior dose de P (120 mg dm-
3) adicionada e foi o tratamento que proporcionou maior rendimento do óleo
essencial de Mentha x piperita L. var. citrata.
O óleo essencial apresentou como componentes majoritários linalol e
acetato de linalol, compostos utilizados como matéria-prima de diversos

70
produtos processados por indústrias de alimentos, cosmética e farmacêutica, o
que torna a utilização de FMAs importante para produção desse composto já
que, em alguns tratamentos fúngicos, a porcentagem de linalol chegou a mais
de 50 % do total de óleo extraído.
O acima exposto comprova a eficiência dos FMAs e indica a utilização
destes como estimuladores da produtividade do vegetal, do teor e rendimento
do óleo essencial, principal produto e fonte de interesse industrial proveniente
de M. x piperita var. citrata.