universidade estadual de santa cruz programa...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DE SANTA CRUZ
PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUÇÃO VEGETAL
DIVERSIDADE DE FUNGOS NO AMBIENTE DE SERINGAIS NO SUDESTE DA
BAHIA E RESISTÊNCIA DE GENÓTIPOS DE SERINGUEIRA À ANTRACNOSE
FOLIAR
TACILA RIBEIRO SANTOS
ILHÉUS – BAHIA 2018

TACILA RIBEIRO SANTOS
DIVERSIDADE DE FUNGOS NO AMBIENTE DE SERINGAIS NO SUDESTE DA
BAHIA E RESISTÊNCIA DE GENÓTIPOS DE SERINGUEIRA À ANTRACNOSE
FOLIAR
ILHÉUS – BAHIA
2018
Tese apresentada à Universidade Estadual de
Santa Cruz como parte das exigências para a
obtenção do título de doutora em Produção
Vegetal.
Área de concentração: Proteção de Plantas
Orientador: Dr. José Luiz Bezerra
Coorientadora: Dra. Edna Dora Martins
Newman Luz

TACILA RIBEIRO SANTOS
DIVERSIDADE DE FUNGOS NO AMBIENTE DE SERINGAIS NO SUDESTE DA
BAHIA E RESISTÊNCIA DE GENÓTIPOS DE SERINGUEIRA À ANTRACNOSE
FOLIAR
Ilhéus, 28 de fevereiro de 2018.
_________________________________________
Dr. José Luiz Bezerra
Biólogo – Ph.D em Fitopatologia
PPGPV – UESC
(Orientador)
_________________________________________
Dr. Marcos Vinícius Oliveira dos Santos
Engenheiro Agrônomo – Doutor em Biologia de Fungos
CEPLAC, Ilhéus, Bahia
_________________________________________
Dr. Adonias de Castro Virgens Filho
Engenheiro Agrônomo – Doutor em Agronomia-Fitotecnia
CEPLAC, Ilhéus, Bahia
_________________________________________
Dr. Cláusio Antônio Ferreira de Mello
Biólogo – Doutor em Genética e Biologia Molecular
PPGPV – UESC
_________________________________________
Dr. Álvaro Figueredo dos Santos
Engenheiro Agrônomo – Doutor em Fitopatologia
EMBRAPA FLORESTAS
Tese apresentada à Universidade Estadual
de Santa Cruz como parte das exigências
para a obtenção do título de doutora em
Produção Vegetal.

DEDICATÓRIA
A Deus, pela saúde e oportunidade que foram concedidas a mim nessa longa jornada.
Aos meus orientadores e ao meu filho.

AGRADECIMENTOS
A Deus, por estar presente em todos os momentos de minha vida.
Ao meu orientador, Dr. José Luiz Bezerra pela oportunidade, paciência e
ensinamentos.
A minha coorientadora, Dra. Edna Dora Martins Newman Luz, pelo amor fraterno,
pela amizade, confiança, orientações, respeito, fé, oportunidade e paciência.
Ao Dr. Antônio Alves Pimenta Neto, Dr. Marcos Vinícius Oliveira dos Santos, Dr.
Adonias de Castro Virgens Filho pela amizade, carinho, colaboração, apoio e por estarem
sempre presentes nos momentos mais difíceis dessa caminhada.
Ao Dr. Cláusio Antônio Ferreira de Melo, pela dedicação, ensinamentos, amizade e
colaboração nas análises moleculares.
Ao Dr. Ronan Xavier, pela oportunidade.
Ao Dr. José Luiz Pires, pelas orientações nas análises estatísticas.
As Plantações Michelin da Bahia e a engenheira agrônoma Lívia Fernanda Lavrador
Toniasso, pela colaboração, disponibilizando todo material vegetal utilizado para a realização
deste trabalho.
A todos os funcionários da CEPLAC, em especial aos funcionários de campo, pela
brilhante colaboração e pela amizade cultivada.
A CAPES, pelo apoio financeiro, pela concessão da bolsa de doutorado, possibilitando
o desenvolvimento da tese.
Aos meus companheiros e amigos, Magui, Catarino, Gisele, Francis, Irina, Lurdinha,
Tita, Cenilda, Deny, Dilze, Elisangela, Ohana, também ao Leonardo pela cooperação,
amizade, paciência, amor e carinho demonstrado.
A minha família amada, pela paciência e apoio.
A meu FILHO querido e amado Davi Vinício, ser único e gracioso, apenas por
EXISTIR.

vi
DIVERSIDADE DE FUNGOS NO AMBIENTE DE SERINGAIS NO SUDESTE DA
BAHIA E RESISTÊNCIA DE GENÓTIPOS DE SERINGUEIRA À ANTRACNOSE
FOLIAR
EXTRATO
É vasta a diversidade de fungos relatados no ambiente dos seringais, alguns deles patogênicos
outros decompositores e/ou antagonistas. O estudo dessas espécies é de grande importância no
sentido de subsidiar o manejo de doenças que acometem a seringueira. A heveicultura brasileira
tem enfrentado diversos problemas fitossanitários nos quais se destacam a alta incidência da
Antracnose, causada por Colletotrichum spp. É comum encontrar várias espécies de
Colletotrichum patogênicas a um mesmo hospedeiro, assim como é comum uma mesma
espécie causar sintomas da doença em diversas plantas hospedeiras, destacando assim a
dificuldade em se elaborar estratégias de manejo para a doença. A correta identificação do
agente causal, o conhecimento da etiologia da doença, a diversidade genética e estrutura da
população do patógeno, bem como avaliação de genótipos para resistência são de grande
importância. Este trabalho foi desenvolvido com os seguintes objetivos: i) conhecer a
diversidade fúngica presente no folhedo de seringueiras adultas através de coletas e
identificação morfométrica das espécies; ii) Coletar e identificar os complexos de
Colletotrichum encontrados nos seringais em estudo, e iii) testar genótipos desenvolvidos no
programa de melhoramento genético das Plantações Michelin da Bahia, no município de
Igrapiúna, quanto à reação a isolados dos principais complexos de espécies do patógeno
presentes na região Sudeste da Bahia. No estudo da diversidade fúngica em alguns seringais da
região, foram identificados 29 gêneros, dentre eles, uma nova espécie do gênero
Spermosporella. Vinte e nove isolados de Colletotrichum spp. foram coletados e identificados
em nível de espécie por caracterização morfocultural, amplificação e sequenciamento,
comparando sequencias genômicas das regiões ITS1/4 e do gene GAPDH. A análise da
diversidade genética foi avaliada pela obtenção dos marcadores moleculares ISSR. Constatou-
se a presença de três complexos de espécies C. gloeosporioides, C. acutatum e C. boninense¸
distantes entre si. Trinta e nove genótipos de seringueira foram avaliados quanto a sensibilidade
a quatro isolados pertencentes aos três complexos de espécies (C. gloeosporioides, C.
boninense e C. acutatum) através do método de inoculação em folíolos destacados com
suspensão de conídios (3x105), porém, nem todos os genótipos foram testados com os quatro
isolados. As reações dos genótipos variaram com o isolado inoculado, demostrando haver
variabilidade entre eles. Em todos os genótipos houve a formação de lesões em algum
momento após a inoculação. O clone FDR 5788 se destacou por apresentar isoladamente ou
como parental de progênies os melhores resultados para resistência a Colletotrichum spp. Este
trabalho é pioneiro para o patossistema Colletotrichum x seringueira por apresentar uma
metodologia de inoculação prática, reprodutível e eficiente na diferenciação de genótipos
resistentes e suscetíveis ao patógeno, além de demostrar que a variabilidade do complexo de
espécies de Colletotrichum interfere na busca da resistência.
Palavras-chave: Diversidade, Colletotrichum, caracterização, resistência.

vii
DIVERSITY OF FUNGI IN THE RUBBER TREE FROM SOUTHEAST BAHIA AND
AND GENOTYPE RESISTANCE TO FOLIAR ANTHRACNOSIS
ABSTRACT
The diversity of fungi reported in environments of rubber tree plantations is wide some of their
wich are pathogenic while others are decomposers and/or antagonists. The study of these
species has a huge importance to support the management of diseases that affect the rubber
tree. Brazilian rubber tree cultivation has faced several phytosanitary problems, especially high
incidence of Anthracnosis caused by Colletotrichum spp. It is common to find several
Colletotrichum species pathogenic to the same host, and also the same species causing
symptoms of the disease in different host plants, thus highlighting the difficulty to establish
management strategies for this disease. Correct identification of causal agent, knowledge on
disease etiology, genetic diversity and pathogen population structure, as well as evaluation of
genotypes for resistance, are of great importance. This study aimed at: i) to know the fungal
diversity present in the leaves of adult rubber trees, based on collections and morphometric
identification of the species; ii) collect and identify the Colletotrichum complexes found in the
rubber tree plantations studied, and iii) test accessions developed in the genetic improvement
program of the Plantações Michelin da Bahia, in the city of Igrapiúna, for the reaction to
isolates of the main complexes of the pathogen species present in the Southeastern region of
Bahia. The study on fungal diversity in some rubber tree plantations of the region identified 29
genera, including a new species of the genus Spermosporella. Twenty-nine isolates of
Colletotrichum spp. were collected and identified at species level by morpho-cultural
characterization, amplification and sequencing by comparing genomic sequences of the ITS1/4
regions and GAPDH gene. Genetic diversity analysis was evaluated by obtaining the ISSR
molecular markers. Three species complexes were found distant from one another: C.
gloeosporioides, C. acutatum and C. boninense. Thirty-nine rubber tree accessions were
evaluated for the sensitivity to four isolates belonging to the three species complexes (C.
gloeosporioides, C. boninense and C. acutatum) by the method of inoculation in detached
leaflets with conidia suspension (3x105), but not all accessions were tested with the four
isolates. Reactions of the accessions varied according to the isolate inoculated, demonstrating
that there is variability between them. In all accessions, lesions were formed at some time after
inoculation. The clone FDR 5788 stood out for showing, individually or as parent of progenies,
the best results for resistance to Colletotrichum spp. This study is precursor in the
Colletotrichum x rubber tree pathosystem, and presents an inoculation methodology that is
practical, reproducible and able to distinguish resistant and susceptible accessions to the
pathogen, besides demonstrating that the variability in the Colletotrichum species complex
interferes with the search for resistance.
Key words: Diversity, Colletotrichum, characterization, resistance.

viii
LISTA DE FIGURAS
CAPÍTULO I
Figura 1. Arthrobotrys arthrobotryoides (A-B). A. conídios e conidióforos; B.
conídios. Aschersonia cubensis (C-D) C. conídios (seta) e conidióforos; D.
estroma. Cladosporium cladosporioides. E. conídio e conidióforo.
Curvularia eragrostidis. F. conídios e conidióforo. Curvularia lunata var.
aeria. G. conídios. Dimerosporiella cf. paulistana. H. ascósporos expulsos
através do ostíolo. Gliocladiopsis tenuis. I. conidióforo ramificado e
conídios. Irenopsis vincensii. J. primórdio peritecial, hifas, hifopódios,
ascósporo germinando (seta). Leptomeliola uvariae. L. setas periteciais,
ascos e ascósporos. Barras: A = 37 µm; B = 20 µm; C = 15 µm; D = 20
µm; E = 3,5 µm; F = 20 µm; G = 20 µm; H = 15 µm; I = 43 µm; J = 12
µm; L = 40 µm................................................................................................
43
Figura 2. Nodulisporium ochraceum (A-B) A. conídios; B. conidióforos. Nigrospora
sphaerica (C-D) C. conídios; D. célula conidiogênica. Cylindrocarpon cf.
candidulum. E. conídios e células conidiogênicas. Dimerina mindanaensis.
F. peritécio rompido, ascos e ascósporos. Pestalotiopsis suffocata. G.
conídio. Trichoderma atroviride. H. conidióforo e conídios. Lasiodiplodia
theobromae. I. conídios uniseptados. Spiropes helleri. J. conídio. Fusarium
decemcellulare. L. conídios. Barras: A = 12 µm, inset = 14 µm; C = 20 µm;
E = 50 µm; F = 25 µm; G = 5 µm; H = 10 µm; I = 25 µm; J = 14 µm; L = 23
µm...................................................................................................................... 44
Figura 3. Atractilina parasitica. A. Hifas de A. parasitica associadas com hifas de
Irenopsis vincensii. B. Conídio com extremidade capitada. C. Sinêmios.
Barra: B = 10 µm; C = 70 µm........................................................................... 45
CAPÍTULO II
Figure 1. Spermosporella irenopsidis. A. Conidia. B. Conidiogenous cells formed in
the hyphae. C. Conidiogenous cells. Bar = 12.7μm ......................................... 53
Figure 2. Spermosporella irenopsidis A. Conidiogenous cells formed in the hyphae.
B. Conidia. Bar = 10.6 μm................................................................................ 53
Figure 3. Spermatoloncha maticola. A. Conidia. B. Hyphal aggregates and
conidiogenous cells. C. Hyphae of S. maticola contact hyphae of Irenopsis
vincensii. Bar = 13 μm...................................................................................... 54
Figure 4. Spermatoloncha maticola. A, B and C. Conidia and hyphal aggregates. Bar
A = 38 μm, B and C =12 μm……………………….………………….….. 54
CAPÍTULO III
Figura 1. Colônias de Colletotrichum spp. apresentando alta variabilidade
morfocultural em meio BDA............................................................................ 67
Figura 2. Diferentes formas de conídios de Colletotrichum spp. isolados de folíolos
de seringueira (Hevea brasiliensis) em meio BDA. (a) cilíndrico, (b)
oblongo (c) clavado (d) oblongo (e) oblongo-cilindráceo (f) oblongo (g)
oblongo-cilindráceo (h) fusoide (i) oblongo................................................... 68

ix
Figura 3. Análise filogenética dos isolados com base nas sequências ITS1/4 e
GAPDH.............................................................................................................
72
Figura 4. Produto da amplificação do primer UBC- 835 separados por eletroforese em
gel de agarose 1,8% corados com SYBR .................................................... 73
Figura 5. Dendrograma baseado no polimorfismo associados aos loci ISSR de
isolados de Colletotrichum spp. Coeficiente de similaridade de Simple
Matching e agrupamento pelo método UPGMA............................................. 74
Figura 6. Análise de coordenadas de componentes principais geradas pela distribuição
dos isolados baseadas em uma matriz de similaridade genética de Simple
Matching............................................................................................................ 75
CAPÍTULO IV
Figura 1. Folíolo do clone PMB 1 com lesões circulares, concêntricas de coloração
amarronzadas, proveniente de inoculação com gotas de 20 µL da suspensão
de esporos de Colletotrichum gloeosporioides................................................. 96
Figura 2. Dendrograma da análise de agrupamento obtido pelo método de UPGMA
(distância Euclidiana), a partir das medidas de dissimilaridade entre 38
genótipos de seringueira (Hevea sp.) avaliados quanto a severidade da
Antracnose na superfície abaxial de folíolos, quando inoculados com o
isolado 19 de Colletotrichum acutatum. Grupos formados após corte na
altura de θ = 10: I, II, III, IV, V......................................................................... 103
Figura 3. Dendrograma da análise de agrupamento obtido pelo método de UPGMA
(distância Euclidiana), a partir das medidas de dissimilaridade entre 12 gen
ótipos de seringueira (Hevea sp.) avaliados quanto a severidade da
Antracnose na superfície abaxial de folíolos, quando inoculados com o
isolado 13 de Colletotrichum gloeosporiiodes. Grupos formados após corte
na altura de θ = 6: I, II, III, IV,
V........................................................................................................................ 104
Figura 4. Dendrograma da análise de agrupamento obtido pelo método de UPGMA
(distância Euclidiana), a partir das medidas de dissimilaridade entre 10
genótipos de seringueira (Hevea sp.) avaliados quanto a severidade da
Antracnose na superfície abaxial de folíolos, quando inoculados com o
isolado 14 de Colletotrichum acutatum. Grupos formados após corte na
altura de θ = 10: I, II, III................................................................................. 105
Figura 5. Dendrograma da análise de agrupamento obtido pelo método de UPGMA
(distância Euclidiana), a partir das medidas de dissimilaridade entre 18
genótipos de seringueira (Hevea sp.) avaliados quanto a severidade da
Antracnose na superfície abaxial de folíolos, quando inoculados com o
isolado 06 de Colletotrichum boninense. Grupos formados após corte na
altura de θ = 20: I, II, III, IV,
V........................................................................................................................ 106
APÊNDICE A
Figura 1. Oidium heveae. A. Muda de seringueira com sintomas da doença. B. Folíolo
com micélio de oídio. C, D. Conidióforo e conídios. E. Conídios em
cadeia. Barra = 37 μm..................................................................................... 130

x
LISTA DE TABELAS
CAPÍTULO II
Table 1. Comparative morphology of species of Spermosporella, including the new
species, S. irenopsidis…….…………………………………………..……… 52
CAPÍTULO III
Tabela 1. Designação, origem, órgão de isolamento e ano de coleta de isolados de
Colletotrichum spp. utilizados neste estudo........................................................ 62
Tabela 2. Características morfoculturais de 29 isolados de Colletotrichum spp. obtidos
de folíolos de seringueira (Hevea brasiliensis) em meio BDA........................... 69
Tabela 3. Primers com suas sequências amplificadas para a obtenção dos marcadores
moleculares ISSR. Temperatura de anelamento e número de bancas que
representam os alelos polimórficos obtidos e utilizados em análise................... 73
CAPÍTULO IV
Tabela 1. Descrição dos 39 genótipos de Hevea spp. testados para resistência à
Antracnose foliar da seringueira.......................................................................... 94
Tabela 2. Diâmetro médio da lesão (DML) em mm, em folíolos destacados de 38
genótipos de seringueira (Hevea brasiliensis) inoculados com suspensão de
3x105 conídios/mL do isolado 19 de Colletotrichum
acutatum............................................................................................................. 98
Tabela 3. Diâmetro médio da lesão (DML) em mm, em folíolos destacados de 18
genótipos de seringueira (Hevea brasiliensis) inoculados com suspensão de
3x105 conídios/mL do isolado 06 de Colletotrichum
boninense............................................................................................................ 99
Tabela 4. Diâmetro médio da lesão (DML) em mm, em folíolos destacados de 12
genótipos de seringueira (Hevea brasiliensis) inoculados com suspensão de
3x105 conídios/mL do isolado 13 de Colletotrichum
gloeosporioides.................................................................................................. 100
Tabela 5. Diâmetro médio da lesão (DML) em mm, em folíolos destacados de 10
genótipos de seringueira (Hevea brasiliensis) inoculados com suspensão de
3x105 conídios/mL do isolado 14 de Colletotrichum acutatum.......................... 100

xi
SUMÁRIO
EXTRATO..................................................................................................................... VI
ABSTRACT................................................................................................................... VII
LISTA DE FIGURAS................................................................................................... IX
LISTA DE TABELAS................................................................................................... X
1. INTRODUÇÃO...................................................................................................... 13
2. REVISÃO DE LITERATURA............................................................................. 15
2.1 Diversidade de fungos em seringais....................................................................... 15
2.2 A cultura da seringueira......................................................................................... 16
2.3 Antracnose foliar da seringueira........................................................................... 18
2.4 Características do gênero Colletotrichum............................................................. 20
2.5 Epidemiologia e sintomas....................................................................................... 21
2.6 Controle da Antracnose.......................................................................................... 22
2.7 Identificação de Colletotrichum spp....................................................................... 25
2.8 Diversidade genética............................................................................................... 26
CAPITULO I
3. TAXONOMIA DE FUNGOS EM SERINGAIS DO SUDESTE DA
BAHIA............................................................................................................................ 28
RESUMO....................................................................................................................... 28
ABSTRACT.................................................................................................................. 29
3.1 INTRODUÇÃO....................................................................................................... 30
3.2 MATERIAL E MÉTODOS.................................................................................... 32
3.2.1 Locais de coletas.................................................................................................... 32
3.2.2 Isolamento dos fungos............................................................................................ 32
3.2.2.1 Isolamento direto................................................................................................. 32
3.2.2.2 Isolamento indireto.............................................................................................. 32
3.2.2.3 Isolamento de amostras de solo........................................................................... 33
3.3 RESULTADOS E DISCUSSÃO............................................................................ 34
3.3.1 Isolamento e identificação de fungos..................................................................... 34
3.3.2 Taxonomia.............................................................................................................. 34
3.4 REFERÊNCIAS BIBLIOGRÁFICAS................................................................. 46
M
CAPITULO II
4. Spermosporella irenopsidis sp. nov. and Spermatoloncha maticola, parasitic
on black mildew (Irenopsis vincensii) of rubber in Bahia, Brazil……...…..….… 49
ABSTRACT………………………….………………………………………………. 49
4.1 INTRODUCTION……………………………………………………………… 49
4.2 MATERIALS AND METHODS………………………………………………. 50
4.3 RESULTS…………………………...…………………………….……............ 51
4.3.1 Taxonomy 51
4.4 DISCUSSION…………………………………………………………………… 55
4.5 ACKNOWLEDGEMENTS……………………………………………...…….. 55
4.6 REFERENCES…………………………………………………………………. 55
M
CAPITULO III
5. CARACTERIZAÇÃO MORFOCULTURAL, MOLECULAR E DIVERSIDADE
GENÉTICA DE Colletotrichum spp. AGENTE CAUSAL DA ANTRACNOSE

xii
FOLIAR EM SERINGUEIRA (Hevea brasiliensis)...............................................57
RESUMO....................................................................................................................... 57
ABSTRACT................................................................................................................... 58
5.1 INTRODUÇÃO....................................................................................................... 59
5.2 MATERIAL E MÉTODOS.................................................................................... 61
5.2.1 Coleta e isolamento de Colletotrichum spp............................................................ 61
5.2.2 Caracterização morfocultural dos isolados de Colletotrichum.............................. 61
5.2.3 Teste de esporulação das espécies de Colletotrichum............................................ 63
5.2.4 Teste de patogenicidade......................................................................................... 63
5.2.5 Identificação molecular por sequenciamento......................................................... 63
5.2.6 Diversidade genética em regiões ISSR.................................................................. 65
5.3 RESULTADOS........................................................................................................ 66
5.3.1 Caracterização morfocultural................................................................................. 66
5.3.2 Sequenciamento e identificação dos níveis taxonômicos dos isolados.................. 71
5.3.3 Análise da diversidade genética em loci ISSR................................................................... 72
5.4 DISCUSSÃO............................................................................................................ 76
5.5 REFERÊNCIAS BIBLIOGRÁFICAS.................................................................. 83
CAPÍTULO IV
6. RESISTÊNCIA DE GENÓTIPOS À ANTRACNOSE FOLIAR........................ 89
RESUMO....................................................................................................................... 89
ABSTRACT................................................................................................................... 90
6.1 INTRODUÇÃO....................................................................................................... 91
6.2 MATERIAL E MÉTODOS.................................................................................... 93
6.2.1Organismos e condições de cultivo......................................................................... 93
6.2.2 Inoculação e avaliação de sintomas........................................................................ 93
6.2.3 Análise dos dados..................................................................................................... 94
6.3 RESULTADOS........................................................................................................ 96
6.4 DISCUSSÃO............................................................................................................ 107
6.5 REFERÊNCIAS BIBLIOGRÁFICAS.................................................................. 112
M
7. CONSIDERAÇÕES FINAIS................................................................................... 116
M
8. REFERÊNCIAS BIBLIOGRÁFICAS.................................................................... 117
M
APÊNDICE A
9. OCORRÊNCIA DE OÍDIO EM MUDAS DE SERINGUEIRA NO ESTADO
DA BAHIA, BRASIL.............................................................................................. 127
RESUMO....................................................................................................................... 128
ABSTRACT................................................................................................................... 128
REFERÊNCIAS BIBLIOGRÁFICAS........................................................................ 130

13
1. INTRODUÇÃO
A seringueira é uma planta natural da Bacia Amazônica, pertencente ao gênero Hevea
Aubl,, família Euphorbiaceae, e compreende onze espécies, entre as quais a Hevea
brasiliensis (Wild. ex. A. Juss) Muell. Arg. é a mais plantada comercialmente, por apresentar
a mais alta produtividade, produzir borracha natural de superior qualidade, além de possuir
uma ampla base genética (GONÇALVES et al., 1997; COSTA et al., 2001).
No Brasil, a heveicultura é uma atividade de importância no agronegócio, pelo fato de
o país possuir uma moderna indústria de pneumáticos e artefatos, ser um importante
consumidor de elastômeros e apresentar condições favoráveis para ampliar a oferta de
borracha natural (VIRGENS FILHO, 2013).
No mercado mundial, cerca de 75 % do consumo de borracha natural é realizado pela
indústria de pneumáticos, sendo o restante demandado pelos diferentes segmentos das
indústrias de artefatos de borracha. Em 2017, a produção mundial de borracha natural foi
correspondente a 12.081 milhões de toneladas, enquanto o consumo foi de 12.596 milhões de
toneladas. O Brasil produziu 184 mil toneladas de borracha natural em 2015, em uma área
explorada de 163.000 hectares e consumiu 403 mil toneladas, sendo as importações
correspondentes a 208 mil toneladas (ESPERANTE, 2017). Neste mercado, de acordo com
dados do IBGE (2017), a Bahia participou com uma produção de 28.590 toneladas em uma
área colhida de 33.595 hectares, mantendo a sua participação como segundo estado maior
produtor de borracha natural do país e o segundo em área plantada.
A introdução da seringueira no Estado da Bahia data de 1908, mas a expansão do
cultivo em grande escala só teve início no final dos anos 1950 e início dos anos 1960, quando
o Governo do Estado fomentou o plantio de mais de 10.000 hectares. A partir desta fase, o
setor privado empreendeu grandes projetos, estimulado pelos incentivos fiscais concedidos
pelo governo. Mais tarde, o Governo Federal promoveu o fomento da heveicultura através do
Programa de Incentivo a Produção de Borracha Vegetal – Probor, sendo a seringueira
expandida em toda a região Litoral Sul da Bahia (VIRGENS FILHO, 2003).
Mais de dois terços dos seringais baianos são explorados em sistemas agroflorestais
com o cacaueiro, modelo que tem a vantagem de aumentar as receitas por unidade de área, ao
ofertar produtos de fácil comercialização como a borracha e o cacau (MARQUES, 2000;
VIRGENS FILHO, 2007), além de formar um ambiente ecologicamente favorável à
sobrevivência de pequenos animais, pássaros, insetos e fungos.

14
Nos seringais da Bahia, as preocupações foram sempre com o manejo fitossanitário
dirigido ao controle das doenças fúngicas, especialmente o Mal das Folhas causado por
Microcyclus ulei P. Henn, a Requeima e a queda anormal das folhas (Phytophthora spp.),
além do cancro do tronco (Phytophthora capsici Leonian) e do mofo cinzento (Ceratocystis
fimbriata) (CANDEIAS et al., 2014; CERQUEIRA et al., 2011). A partir de meados da
década 2000, a Antracnose causada por Colletotrichum spp., passou a constituir um dos
principais problemas pelos danos de importância econômica que as plantações sofriam em
função dos ataques de patógeno. Este fato passou a merecer atenção especial no manejo da
seringueira, principalmente por causar a morte descendente dieback em plantas jovens,
promover o desfolhamento em seringais adultos e causar lesões nos ramos. Mesmo nos anos
de baixa incidência de outras doenças fúngicas, a Antracnose vem se manifestando de
maneira epidêmica, prejudicando a atividade fotossintética na copa da seringueira.
Os seringais constituem um sistema que simula algumas funções ecológicas da floresta
tropical (IAC, 2017), sendo um ambiente bastante propício ao crescimento e sobrevivência de
fungos, embora seja um agrossistema pouco explorado quanto à sua micodiversidade. Os
fungos que habitam a filosfera das plantas desempenham papéis diversos atuando
beneficamente como decompositores da matéria orgânica, promotores da ciclagem de
nutrientes, hiperparasitas ou biocontroladores de patógenos (ALEXOPOULOS et al., 1996;
KENDRICK, 2000).
Diante da nova preocupação, com o aumento da incidência da Antracnose, torna-se
necessário identificar corretamente as espécies de Colletotrichum (Corda) ou os complexos de
espécies existentes nos seringais da região, estudar a sua distribuição e testar a reação de
resistência dos materiais genéticos mais promissores, em relação a esta enfermidade,
principalmente aqueles já avaliados quanto à resistência a M. ulei. A correta identificação do
patógeno é de suma importância para a seleção de genótipos resistentes, sendo necessário o
uso de ferramentas moleculares no caso do gênero Colletotrichum onde existem espécies
crípticas impossíveis de serem distinguidas morfologicamente (WEIR et al., 2012; SUTTON,
1992).
Assim os objetivos desta pesquisa foram: i) conhecer a diversidade fúngica presente
em folhas da copa e da serapilheira de seringais em produção; ii) identificar os complexos de
espécies de Colletotrichum encontrados nesses seringais; e iii) testar genótipos desenvolvidos
no programa de melhoramento genético das Plantações Michelin da Bahia, (cooperação
CIRAD/Michelin) e outras instituições, quanto a reação a isolados do patógeno presentes na
região em estudo.

15
2. REVISÃO DE LITERATURA
2.1 Diversidade de fungos em seringais
Os fungos podem ser considerados os organismos mais importantes encontrados no
meio ambiente, devido ao seu papel no funcionamento do ecossistema e influência nas
atividades relacionadas aos seres humanos. Eles são essenciais na decomposição da matéria
orgânica, reciclagem e transporte de nutrientes, sendo indispensáveis para alcançar o
desenvolvimento sustentável (PALM; CHAPELA, 1998). Os fungos também possuem
importância médica e biotecnológica, como por exemplo, na produção de enzimas e
antibióticos úteis ao tratamento de enfermidades e aplicáveis em outras atividades humanas.
Estima-se que existam 1,5 milhões de espécies fúngicas no mundo (HAWKSWORTH,
1991), no entanto, menos de 5% foram descritas. A capacidade dos fungos em produzirem
enzimas permite que eles atuem em praticamente todos os tipos de substratos. As folhas estão
entre os substratos mais utilizados e relevantes para a colonização dos fungos por ser um
material mais facilmente degradável e encontrado em maior abundância nos ecossistemas
naturais (DIX; WEBSTER, 1995). Considerando a variedade de ambientes que eles ocupam,
praticamente não existem barreiras geográficas para a sua distribuição (MAIA, 2003).
No interior dos seringais existe uma condição bastante propicia ao crescimento e
sobrevivência de fungos, uma vez que há um ambiente estável, com características que
simulam algumas funções da floresta tropical (IAC, 2017). É vasta a diversidade de fungos
estudados nesses ambientes. Cerca de 323 espécies de fungos associados a Hevea brasiliensis
Muell.-Arg, foram reportados a nível mundial (FARR; ROSSMAN, 2017). Na América do
Sul, foi registrada a ocorrência de 120 táxons em H. brasiliensis, incluindo espécies
patogênicas e não patogênicas, sendo a grande maioria pertencente ao filo Ascomycota
(VIÉGAS, 1961). No Brasil, cerca de 53 espécies fúngicas foram relatadas associadas a
Hevea spp. (MENDES et al., 1998; EMBRAPA, 2017).
Esta grande diversidade de fungos no ambiente do seringal ocupa diferentes micro
habitats onde desempenham papéis ecológicos variáveis: 1) folhas vivas na copa, incluindo
espécies fitopatogênicas, hiperparasitas, biocontroladoras de patógenos, comensais e parasitas
de artrópodes; 2) troncos e galhos abrangendo sapróbios e fungos liquenizados; 3) serapilheira
realizando a decomposição da matéria orgânica e promovendo a ciclagem de nutrientes; 4)
raízes, vivendo como sapróbios ou patógenos radiculares. As espécies que habitam a filosfera,
em sua maioria são leveduras e/ou fungos leveduriformes, enquanto que as espécies da
rizosfera correspondem a fungos que se nutrem de exsudatos secretados pelas raízes. Além

16
desses grupos existem os fungos endofíticos que vivem no interior dos tecidos vegetais sem
causar danos aparentes às plantas. Merece destaque às espécies de fungos considerados como
patógenos vasculares que se desenvolvem tanto no floema como no xilema das plantas
(ALEXOPOULOS et al., 1996; KENDRICK, 2000).
Os fungos não patogênicos associados à cultura da seringueira devem ser estudados
porque, entre outros papéis, interferem na sobrevivência de espécies causadoras de doenças
como o Microcyclus ulei (P. Henn.) v. Arx, agente etiológico do Mal das Folhas e
Colletotrichum spp. (Corda) agentes da Antracnose foliar da seringueira. Além disso, é grande
a possibilidade de existirem espécies desconhecidas para a ciência cujo potencial ainda é
desconhecido.
2.2 A cultura da seringueira
A heveicultura é uma atividade de grande importância social, econômica e ambiental,
pela sua capacidade de geração de emprego e renda (VIRGENS FILHO, 2007;
NARAYANAN e MYDIN, 2011). Contribui para conservação do solo e melhoria do
ambiente; além disso, a seringueira durante seu ciclo de vida contribui para o sequestro de
carbono da atmosfera (LAKSHMAN; MUNASINGHE, 2017) e as árvores adultas podem ser
fonte de madeira constituindo um sistema de duplo propósito (CORLEY, 1983).
Pertencente ao gênero Hevea, família Euphorbiacea, ordem Malpighiales, a
seringueira, tem seu centro de origem na região Amazônica do Brasil e possui onze espécies
conhecidas: H. brasiliensis (Wild. ex. A. Juss) Muell. Arg., H. guianensis Aub., H.
benthamiana Muell. Arg., H. nitida Mart. ex. Muell. Arg., H. pauciflora (Spr. ex. Bth.) Muell.
Arg., H. rigidifolia (Spr. ex. Bth.) Muell. Arg., H. camporum Ducke, H. spruceana (Bth.)
Muell. Arg., H. microphylla Ulle, H. camargoana Pires e H. paludosa Ulle, Jhrb
(GONÇALVES et al., 1990).
A espécie H. brasiliensis produz látex de excepcional qualidade, sendo muito utilizada
em plantios em todo o mundo, pois possibilita a alta produtividade de borracha natural e
possui uma ampla base genética (GONÇALVES et al., 1990; COSTA et al., 2011). O látex é
o principal subproduto da seringueira, e atualmente é a principal fonte de borracha natural do
mundo (ALVARENGA; CARMO, 2008). A borracha natural é utilizada nas indústrias de
pneumáticos e artefatos (balões, materiais cirúrgicos, componentes da indústria
automobilística, de calçados, revestimento de cabos elétricos, isolantes e outros), sendo o

17
abastecimento feito principalmente por países do Sudeste asiático, sendo a contribuição
brasileira de apenas 1,9% do total da produção mundial (VALVERDE et al., 2014).
O Brasil possui uma área plantada estimada em 160.968 ha hectares de seringueira,
dos quais pouco menos da metade encontram-se em produção (IBGE, 2017). Os principais
Estados produtores são: São Paulo, Bahia, Mato Grosso, Espírito Santo e Paraná
(EMBRAPA, 2017). Sendo o estado da Bahia, considerado o segundo maior produtor de
borracha natural do país, possuindo a segunda maior área em exploração estimada em 33.203
ha hectares (IBGE, 2017).
A planta adulta apresenta um processo natural de desfolha, que ocorre geralmente na
estação seca, dura em torno de duas a seis semanas, e é chamado de período de senescência e
queda foliar quando a planta entra na fase de hibernação (MORAES, 1980). Após esse
período, a planta começa a se reenfolhar, merecendo bastante atenção, pois é quando há
lançamento de folíolos jovens com coloração antociânica intensa, o que torna a seringueira
mais suscetível ao ataque de fitopatógenos (MORAES, 1980; GASPAROTTO; PEREIRA,
2012).
Vários fatores afetam o desenvolvimento da heveicultura brasileira, e entre estes,
destacam-se as doenças, principalmente o Mal das Folhas, que só ocorre nas Américas,
principalmente no Brasil (FURTADO; TRINDADE, 2005), causado pelo fungo Microcyclus
ulei (HOLLIDAY, 1970), e a Antracnose causada por Colletotrichum spp., que ocorre na
maioria das regiões heveícolas do mundo (FURTADO; TRINDADE, 2005).
O Mal das Folhas, um dos principais problemas da heveicultura brasileira, é uma
doença devastadora e por isso alguns autores fazem analogias a doenças de grande impacto à
economia em outras culturas, tais como Ferrugem do Cafeeiro (Hemileia vastatrix Berk. &
Broome), a Mela da Batatinha (Phytophthora infestans (Mont.) de Bary) e a Vassoura de
Bruxa do Cacaueiro (Moniliophthora perniciosa (Stahel) Aime & Phillips-Mora) (CHEE;
HOLLIDAY, 1986; GASPAROTTO et al., 1997; RUBINI, 2003; MAKI, 2006). O agente
causal possui grande capacidade de causar danos ao hospedeiro, provocando lesões e
deformação nos folíolos e consequentemente a queda prematura levando a planta ao
desfolhamento parcial ou total (GASPAROTTO et al.,1997; VIRGENS FILHO, 2017 –
Dados não publicados).
Em busca de minimizar as perdas ocasionadas pelo Mal das Folhas, pesquisadores da
Embrapa-CPAA, IAC e CEPLAC, estabeleceram programas de melhoramento genético
visando ao desenvolvimento de clones resistentes à doença (STERLING; RODRIGUEZ,
2011; GONÇALVES; MARQUES, 2104). No entanto, ao longo do tempo, observou-se que

18
muitos genótipos produtivos e parcialmente resistentes ao Mal das Folhas passaram a ser
afetados pela Antracnose (STERLING et al., 2011).
Até o início dos anos 2.000 o gênero Colletotrichum não causava danos de
importância econômica nos seringais. Entretanto, após meados desta década passou-se a
registrar danos frequentes de Antracnose em seringais do Baixo Sul baiano, notadamente nos
anos com condições climáticas mais favoráveis. Desse modo, passou-se a atribuir importância
às espécies deste gênero nos seringais, sendo isso atribuído principalmente à ampla
diversidade e capacidade de adaptação do patógeno ao hospedeiro. Isto possivelmente causará
alterações na dinâmica da relação patógento/hospedeiro, possibilitando surtos epidêmicos de
Antracnose, que aé então, era considerada uma doença secundária para a seringueira.
2.3 Antracnose foliar da seringueira
A Antracnose foliar é uma doença fúngica de grande importância na cultura da
seringueira. É causada por um complexo de fungos pertencentes ao gênero Colletotrichum,
cuja fase teleomórfica corresponde a Glomerella spp. (Stonem.) Spalding & Screnk, fungo da
divisão Ascomycota, ordem Diaphortales (KIRK et al., 2008; HIBBETT et al., 2007).
A doença tem causado sérios prejuízos em seringais de todo o mundo. Alguns
cientistas relatam danos causados pela Antracnose na Índia, Tailândia, Sri Lanka, China,
Oeste da África, Colômbia e também no Brasil (THAMBUGALA; DESHAPPRIYA, 2009;
STERLING et al., 2011; HENZ et al., 1992; SAHA et al., 2002; SIERRA HAYER, 2010). No
continente asiático, a Antracnose é considerada uma das principais causas do declínio nos
rendimentos da borracha natural (CAI et al., 2013).
No Brasil, a Antracnose foi relatada em praticamente todas as regiões onde a
heveicultura está presente, sendo primeiramente observada com grande frequência na região
Amazônica, associada ao Mal das Folhas (FURTADO, 2008).
Virgens Filho; Nakayama (2015) constataram forte ataque da Antracnose em algumas
áreas dos novos plantios dos imóveis associados à Cooperativa Ouro Verde Bahia, localizados
em Igrapiúna, neste estado, notadamente nos clones PMB 1 e CDC 312. No Sudeste da Bahia,
desde 2010, esta enfermidade, já presente na região, vem se constituindo em um dos
principais problemas fitossanitários da heveicultura, notadamente nos novos plantios.
Nos seringais jovens do clone PMB 1 com idade de até três anos, observou-se a morte
da gema apical em algumas plantas provocando desuniformidade no estande, tendo em vista
que o corte dos ramos atacados para tratamento da doença provoca atraso no crescimento das

19
mesmas. Nos seringais adultos foi recomendado o controle da doença com fungicida, através
de pulverizações via aérea ou terrestre, em intervalos semanais nos períodos de baixa
temperatura e alta umidade relativa do ar. Entretanto, nas condições destas plantações, este
método tem custo proibitivo, devido às despesas com pulverizações e eficácia moderada em
razão da complexidade da tecnologia de aplicação, acrescendo a isso o impacto ambiental.
Desse modo conclui-se que a solução mais eficaz para o problema é o controle por meio da
resistência genética (VIRGENS FILHO; NAKAYAMA, 2015 – Dados não publicados).
A respeito dos danos causados pela Antracnose na Bahia, Virgens Filho e Nakayama
(2015), reportaram que entre os anos de 2013 e 2014, ocorreram fortes epidemias em
seringais de 12 fazendas associadas à Cooperativa Ouro Verde (Coopeverde) em Igrapiúna,
Bahia, sendo que nos seringais mais velhos, os clones suscetíveis ao ataque de Phytophthora
spp. e Microcyclus ulei foram também afetados por Colletotrichum spp. O ataque deste
patógeno, assim como as desfolhas recorrentes, causado pelo ácaro (Calacarus heveae Feres e
Tenuipalpus heveae Baker) e percevejo-de-renda (Leptopharsa heveae Drake & Poor),
provocaram sérios danos na produção, sendo agravado pelas condições ambientais locais.
Aproximadamente 6.500 ha das 12 fazendas, além da área comercial das Plantações Michelin
da Bahia, sofreram ataques severos, de maneira que os seringais permaneceram desfolhados
por até seis meses consecutivos. Estes problemas em conjunto, causaram uma redução média
de 35,18% na produção dos seringais das fazendas associadas à Coopeverde, entre os anos
2013 e 2014.
Nos estados da região Norte, a doença ocorre em viveiro, jardim clonal e plantio
definitivo, causando lesões foliares, desfolhamento e mortalidade de ramos e galhos
(FURTADO; TRINDADE, 2005). No estado de São Paulo a doença também já assumiu
proporções epidêmicas, exigindo medidas de controle para evitar danos causados,
principalmente no painel (TRINDADE; FURTADO, 1997). Na Bahia, além da ocorrência no
painel, desde 2006, a Antracnose tem sido observada na forma de surtos esporádicos na parte
aérea e no caule de plantas jovens em reboleiras, no clone PMB 1 (VIRGENS FILHO, 2007).
Desde então, a enfermidade tem avançado de forma significativa, ocasionando danos
econômicos, o que vem preocupando os heveicultores da região.
Gasparotto et al, (2012) registraram lesões foliares, desfolhamento e mortalidade de
ramos e galhos causados pela Antracnose em plantas de Hevea brasiliensis, H. pauciflora, H.
guianensis, H. benthamiana e H. camargoana. Em Hevea brasiliensis a doença ataca diversas
partes da planta como folhas, caule, ramos, pecíolo, frutos, inflorescência, hastes, placas de

20
enxerto em jardim clonal e viveiros e até painéis de sangria, podendo causar a dieback
(FURTADO; SILVEIRA, 1992; SILVEIRA; CARDOSO, 1987; SILVEIRA et al., 1992a).
2.4 Características do gênero Colletotrichum
Colletotrichum (Glomerellaceae, Glomerellales, Ascomycota) é classificado como o
oitavo gênero de fungos fitopatogênicos mais importantes no mundo, sendo de grande
interesse econômico e científico, pois atinge uma grande variedade de culturas relevantes para
o mercado ao redor do mundo (DEAN et al., 2012). Possui distribuição cosmopolita, capaz de
crescer em uma ampla variedade de plantas hospedeiras (CANNON et al., 2012). O fungo
pode atuar como endofítico (ROCHA et al., 2011; MORENO, 2016) ou saprófito associado a
outros patógenos (FURTADO; SILVEIRA, 1992). O patógeno causa uma infinidade de
doenças em frutos pós-colheita, como Antracnose, Podridão de pedúnculo, Varicela em
manga, abacate e mamão (BAILEY; JEGER, 1992), sendo, portanto, notabilizado por causar
doenças de importância econômica para a agricultura no Brasil (SERRA; SILVA, 2004). Na
seringueira, pode causar a Antracnose foliar e Antracnose do painel de sangria.
O gênero Colletotrichum pode ter vários hospedeiros, como também mais de uma
espécie pode estar presente em um único hospedeiro (ROBERTS et al., 2012). O abacate e o
maracujá, por exemplo, podem ser infectados por C. gloeosporioides (Penz.) Penz. & Sacc.,
C. acutatum J.H. Simmonds e por C. boninense Moriwaki, Toy. Sato & Tsukib. (AVILA-
QUEZADA et al., 2007; ALMEIDA; COÊLHO, 2007; TOZZE JUNIOR et al., 2010). A
manga e o pêssego são hospedeiros tanto de C. gloeosporioides como de C. acutatum
(ADASKAVEG; HARTIN, 1997; AFANADOR-KAFURI et al., 2003; BERNSTEIN et al.,
1995; FITZELL, 1979; PLOETZ, 1994). O pimentão é hospedeiro de cinco espécies distintas
de Colletotrichum (TOZZE JÚNIOR, 2006). Em Anonna muricata foram isolados C.
theobromicola, C. tropicales, C. siamense, C. gloeosporioides, C. karstii, e uma espécie
indeterminada no complexo C. boninense, bem como uma espécie indeterminada no
complexo C. acutatum (ALVAREZ et al., 2014). Em feijão lima Sousa et al. (2018)
encontraram entre nove isolados obtidos nos estados do Piauí e Alagoas três espécies: C.
truncatum, C. cliviae e C. fructicola classificadas por análises de multilocus.
Ao estudar o agente causal da Antracnose na seringueira, percebe-se haver incertezas
quanto à identidade do mesmo. Isso porque as espécies de Colletotrichum possuem uma alta
plasticidade fenotípica podendo assim ocorrer erros de identificação e classificação quando

21
baseadas em caracteres morfológicos (MENEZES, 2006). Pode também ocorrer uma variação
na virulência, decorrente da ação de complexos de espécies associados à doença (BROWN;
SOEPENA, 1994; FERNANDO et al., 2000; GUYOT et al., 2001; SAHA et al., 2002;
GUYOT et al., 2005).
Alguns autores relatam que os isolados provenientes de Hevea sp. são pertencentes aos
complexos C. acutatum, C. gloeosporioides e C. boninense, descritos como os agentes causais
da doença na seringueira (PETCH, 1921; CARPENTER; STEVENSON, 1954;
JAYASINGHE et al., 1997; SAHA et al., 2002; GAZIS; CHAVERRI 2010; GAZIS et al.,
2011; DAMM et al., 2012a). O complexo C. acutatum contém mais de 29 espécies
estreitamente relacionadas, o complexo C. gloeosporioides compreende mais de 22 espécies e
o complexo C. boninense tem mais 18 espécies relatadas (DAM et al., 2012; WEIR et al.,
2012).
No Brasil, o estudo relacionado à identificação de espécies de Colletotrichum
associadas à Antracnose foliar da seringueira foi realizado com o uso de ferramentas
moleculares corroborando com a presença dos três complexos anteriormente relatados. Isso
permitiu concluir que os indivíduos do complexo de C. acutatum foram encontrados mais
frequentemente associados à doença (SARMIENTO, 2013).
2.5 Epidemiologia e sintomas
A seringueira está propicia ao ataque de Colletotrichum praticamente em todas as
fases de seu desenvolvimento; no entanto, Gasparotto e Pereira (2012) afirmam que o
patógeno afeta folíolos com até aproximadamente 15 dias de idade, ou seja, a planta está mais
suscetível nos estádios ontogênicos B (duração média são de dez dias), fase correspondente ao
alongamento celular, a qual apresenta dois sub-estádios; o B1, quando os folíolos estão
posicionados verticalmente com o ápice voltado para cima, fortemente carregados com
antocianina; e o B2, quando os ápices dos folíolos são voltados para baixo, de coloração
antociânica menos intensa, esta é a fase de maior velocidade de alongamento do eixo caulinar;
e estádio C (duração média de oito dias), quando os folíolos estão pendentes, flácidos e de cor
verde (HALLÉ et al., 1978).
As infecções são geralmente causadas pela dispersão intensa de conídios, embora
ascósporos também possam causar infecções. A disseminação é feita principalmente por

22
respingos de chuva, ventos e sementes infectadas, que ao serem semeadas, poderão induzir os
sintomas de damping–off.
Uma vez disseminados, os esporos do fungo entram em contato com o tecido do
hospedeiro germinam (4 a 6 horas), penetram nas células do hospedeiro através dos
apressórios, colonizam os tecidos da planta, e posteriormente surgem os primeiros sintomas
visíveis em folhas, inflorescências e frutos (MENEZES, 2002).
O complexo de espécies de Colletotrichum que ataca a seringueira induz diferentes
sintomas. Os sintomas iniciais da Antracnose foliar manifestam-se nas folhas novas,
brotações e frutos. Nas folhas, as lesões são arredondadas, diminutas com 1 a 3 mm de
diâmetro, geralmente numerosas e dispersas no limbo. Estas lesões apresentam a porção
central escura e margem estreita de coloração marrom-avermelhada. Quando o ataque é
intenso, as lesões podem interligar-se acarretando enrugamento do folíolo e consequente
queda.
As lesões podem ocorrer também nos pecíolos e frutos, causando rachaduras e
apodrecimento na casca. Nos ramos verdes, as infecções geralmente ocorrem nas suas
inserções com o galho, podendo causar quebra de ramos na base. Pode ocorrer desfolhamento,
morte da gema apical e seca descendente dos ramos, provocando desuniformidade no estande,
tendo em vista que o corte dos ramos atacados para tratamento da doença provoca atraso no
crescimento das mesmas (VIRGENS FILHO, 2017 – Dados não publicados). Uma
característica bastante peculiar da doença é a abundante esporulação conidial do patógeno
envolta por uma massa alaranjada que sai dos acérvulos (GASPAROTTO, 1997). Como o
patógeno é comum a diversos hospedeiros, isso favorece sua sobrevivência e dificulta seu
controle.
2.6 Controle da Antracnose
No controle da Antracnose, fungicidas a base de chlorothalonil, ou oxicloreto de cobre
têm sido utilizados de forma preventiva em viveiros e jardim clonal. No painel de sangria, o
controle pode ser efetuado através do pincelamento, ou pulverizações com fungicidas a base
de chlorothalonil, chlorothalonil + tiofanato metílico, zineb + óleo vegetal, propiconazele e
tebuconazole (FURTADO; TRINDADE, 2005). Em seringais adultos, devido ao porte
elevado das plantas, o uso de fungicidas é limitado pela complexidade de emprego da
tecnologia de aplicação e custo elevado notadamente nas áreas com topografia declivosa
(GASPAROTTO et al.,1985).

23
Apesar de utilizado, o controle químico além de caro e dispendioso, constitui sério
risco ao ambiente e à saúde humana, principalmente pela presença de resíduos tóxicos,
desequilíbrio biológico, eliminação de organismos benéficos e redução da biodiversidade
(BETTIOL et al., 2014).
Algumas tentativas utilizando o controle biológico em doenças da seringueira foram
feitas. No entanto, todos os trabalhos relacionados ao controle biológico de patógenos
altamente destrutivos, não conseguiram desenvolver uma tecnologia eficiente
(GASPAROTTO; PEREIRA, 2012). Pesquisas com o fungo Dicyma pulvinata (Berk & Curt),
no controle biológico do Mal das Folhas, não apresentaram resultados satisfatórios quando
realizadas em seringais monoclonais altamente suscetíveis (JUNQUEIRA; GASPAROTTO,
1991; TRINDADE; FURTADO, 1997; GASPAROTTO; PEREIRA, 2012).
São poucos os trabalhos envolvendo o controle biológico da Antracnose na
seringueira. Ogbebor e Adekunle (2005) estudaram o efeito de extratos vegetais na inibição
da germinação de esporos e no crescimento micelial de Corynespora cassiicola da
seringueira. Evueh e Ogbebor (2008) estudaram o efeito antagônico in vitro de fungos do
filoplano da seringueira no controle biológico de Colletotrichum spp. e observaram que os
fungos Trichoderma sp., Aspergillus sp., Gliocladium sp., Pleurothecium sp., Botrytis sp.,
Staphylotrichum sp., Trichocladium sp., Gonatorrhodiella sp., Trichophyton sp. e
Syncephalastrum sp. mostraram níveis diferentes de zonas de inibição do patógeno.
Uma das alternativas mais viáveis, práticas e econômicas para o controle de doenças
em culturas perenes de árvores altas é o controle genético (GASPAROTTO et al., 2012). Os
estudos genéticos relacionados à cultura da seringueira iniciaram no final da década de 1930
com a seleção de plantas resistentes ao Mal das Folhas, na cidade de Fordlândia, onde foram
desenvolvidos trabalhos voltados à melhoria de caracteres econômicos importantes como
rendimento e vigor da planta (GONÇALVES et al., 1996). No entanto, nenhuma ênfase foi
dada à Antracnose foliar, relacionado aos aspectos genéticos da resistência à doença naquele
momento pelo fato desta doença não apresentar importância econômica. Somente quando a
Antracnose do painel, causada por Colletotrichum spp., foi detectada em painéis de corte do
clone RRIM 600 por Silveira et al. (1992), foram iniciados estudos genéticos.
No Brasil, ainda são escassos os trabalhos relacionados à seleção de clones resistentes
à Antracnose. Furtado et al. (1994) avaliaram a queda foliar decorrente do ataque de
Colletotrichum no estado de São Paulo, e constataram que os clones GT1, PR 225 e IAN 873,
GT1, IAN 873, RRIM 600 e PR 107 sofreram menores perdas foliares e os clones RRIM 701,
PR 261, PB 217 e PB 235 sofreram maior desfolhamento. Gonçalves et al. (2002) objetivando

24
selecionar clones de seringueira promissores para a região do planalto do estado de São Paulo,
estudaram 16 clones da série IAC 300. Os autores concluíram que os clones IAC 330, IAC
331, IAC 333, IAC 336, IAC 338, IAC 339 e IAC 343 apresentaram alta resistência à
Antracnose do painel, já os clones IAC 340 e RRIM 600 foram suscetíveis.
O conhecimento do comportamento dos clones locais quanto à resistência ou
suscetibilidade ao patógeno é de extrema importância, sendo isto primordial para o sucesso de
um programa de melhoramento.
Clones de seringueira estão sendo desenvolvidos para a resistência a M. ulei, porém
não se sabe a reação dos mesmos a Colletotrichum spp. e nem se conhece ao certo as espécies
do gênero que podem estar envolvidas no patossistema local.
Programas de melhoramento genético da seringueira têm sido desenvolvidos no IAC
no estado de São Paulo e Ceplac em Ilhéus na Bahia, onde visam ganhos em produção, vigor
e resistência principalmente ao Mal das Folhas (GONÇALVES; MARQUES, 2014).
Segundo alguns estudiosos, os esforços no melhoramento para resistência ao M. ulei
falharam devido ao emprego da resistência vertical (monogênica), que se expressa contra
algumas raças do patógeno (HO, 1986; RIVANO et al., 2013). Este tipo de resitência propicia
o aparecimento de novos patótipos do patógeno, virulentos ao germoplasma melhorado
(CHEE et al., 1986; GASPAROTTO et al., 1990; KALIL FILHO; JUNQUEIRA, 1989;
MILLER, 1986).
Para Rivano et al. (2013) o mais preconizado é a obtenção da resistência horizontal
(poligênica), que é uniformemente eficiente contra todas as raças do patógeno (BERGAMIN
FILHO; AMORIM, 1996).
Alguns trabalhos conduzidos para a resistência ao Mal das Folhas, por exemplo,
inferiram que o clone MDF 180 expressa resistência horizontal. Oito clones foram
selecionados como possuidores de resistência ao M. ulei em experimentos conduzidos na
Bahia por Garcia et al. (2004). Tentativas para o uso dessas resistências por meio de
cruzamento com H. brasiliensis tem produzido numerosos clones resistentes com pouca
permanência, ou seja, resistência não durável (PINHEIRO; LIBONATI, 1971)
Devido ao intenso ataque de Colletotrichum spp., a seleção de materiais genéticos de
Hevea sp. para plantio em larga escala deve ser acompanhada de avaliações também quanto à
resistência à Antracnose. No entanto, pode ser que haja resposta diferenciada dos genótipos
do hospedeiro às distintas espécies do patógeno (SAHA et al., 2002; THAMBUGALA;
DESHAPPRIYA, 2009). Segundo Ho (1986), um dos exemplos pode ser obtido do

25
melhoramento para a resistência à Antracnose na Malásia, onde observou-se níveis variantes
de resistência aos diferentes ambientes experimentais.
2.7 Identificação de Colletotrichum spp.
Para se montar estratégias de manejo de uma determinada doença, é importante o
conhecimento prévio de funcionamento do patossistema envolvido. Para isso, o estudo da
caracterização e identificação do agente etiológico, constitui uma das principais etapas.
Na identificação de fungos fitopatogênicos em geral, algumas pesquisas utilizam
técnicas tradicionais de morfologia, como tamanho e formato de conídios. O gênero
Colletotrichum, especificamente, foi caracterizado por vários autores utilizando basicamente
as ferramentas da taxonomia clássica e aspectos culturais como a morfologia da colônia,
forma dos conídios, presença e forma de setas, hospedeiros (CANNON et al., 2012; LENNÉ
et al., 1984; SMITH; BLACK, 1990; MILLS, 1992; MENEZES, 2002). Jayasinghe (1998)
estudou os caracteres fisiológicos na identificação das espécies C. acutatum e C.
gloeosporioides, relacionados ao crescimento micelial, diferentes temperaturas de
crescimento, fontes de carbono e sensibilidade a fungicidas.
Devido à alta plasticidade fenotípica do gênero, suposta existência de formas
intermediárias entre espécies e alta variabilidade das espécies envolvidas, é muito difícil a
identificação baseada unicamente em caracteres morfológicos e culturais, os quais são
considerados insuficientes para a definição das espécies de Colletotrichum (WEIR et al.,
2012; SIERRA-HAYER, 2010; CANNON et al., 2000).
Existem limitações para a identificação de espécies muito próximas como C. acutatum
e C. gloeosporioides, principalmente, porque isolados destas espécies apresentam uma ampla
variação genética e morfológica (SUTTON, 1992). Weir et al. (2012) relataram uma variação
intrínseca de isolados, ao observarem que um isolado monospórico de C. kahawae subsp.
cigarro apresentou dois tipos de colônias. Os autores discutem também que um único isolado
pode apresentar setores na colônia e variar a aparência em resposta ao tempo e método de
preservação. Sendo assim, a utilização de ferramentas moleculares tem sido cada vez mais
utilizada no processo de identificação e caracterização de espécies pertencentes ao gênero
Colletotrichum (BUENO, 2005; NGUYEN et al., 2010).
O uso de marcadores moleculares tem sido bastante estudado, principalmente os
marcadores baseados nas reações de PCR devido à praticidade e à necessidade de pequenas

26
quantidades de DNA da amostra. O uso desses marcadores tem auxiliado na taxonomia
clássica de fungos (Williams et al., 1990).
Reação em cadeia de polimerase (PCR) permite a amplificação de regiões específicas
do genoma, como o Espaço Interno Transcrito (ITS), localizado entre regiões altamente
conservadas. Dentre as regiões do genoma estudadas para Colletotrichum, a ITS é a que
possui maior número de sequências depositadas, sendo considerada útil para a identificação
preliminar de espécie ou para colocá-lo em um complexo de espécie (CAI et al., 2009).
Vários estudiosos utilizaram a técnica de PCR na identificação de espécies de
Colletotrichum em várias culturas. Sreenivasaprasad et al., (1996) utilizaram a PCR para
detectar C. acutatum em frutos de morango. Sarmiento, (2013) baseou-se no PCR
convencional, para o estudo de espécies de Colletotrichum associados à Antracnose foliar da
seringueira. Freemam et al., (1998); Tozze, (2007) utilizaram essa técnica para diferenciar
populações de C. gloeosporioides, C. coccodes, C. kahawae, C. magna e C. orbiculare.
Regiões do genoma de fungos do gênero Colletotrichum têm sido analisadas
juntamente com o ITS utilizando múltiplos marcadores genéticos que levem a uma
identificação mais confiável das espécies, tais como as sequências dos genes que codificam
actina (ACT), quitina sintetase (CHS1), β-tubulina (TUB2), calmodulina (CAL),
gliceraldeído-3-fosfato desidrogenase (GADPH), histona 3 (HIS3), glutamina-sintetase (GS) e
manganês-superóxido desmutase (SOD2) (Cannon et al. 2012; Damm et al., 2012a; Damm et
al., 2012b; Weir et al., 2012). As análises utilizando estes marcadores revelaram que
Colletotrichum compreende 10 complexos de espécies principais, bem como uma série de
pequenos grupos e espécies únicas.
A obtenção de segmentos amplificados em geral obedece as seguintes etapas; a)
extração de DNA molde, b) escolha do seguimento a ser amplificado e obtenção de primers
específicos para o reconhecimento desse seguimento, c) amplificação com a utilização de um
termociclador e d) leitura do produto amplificado após eletroforese e coloração.
2.8 Diversidade genética
O conhecimento prévio da diversidade genética dentro e entre populações de uma
espécie obtida por meio da aplicação de marcadores moleculares é uma etapa inicial
importante para o desenvolvimento de programas de melhoramento genético, uma vez que a
ausência da variabilidade genética inviabiliza sua execução (GEPST, 1993).

27
Técnicas moleculares têm sido utilizadas para estudar a diversidade de espécies de
Colletotrichum, dentre elas o uso do marcador molecular do tipo sequências simples repetidas
internas (ISSR) (SILVEIRA et al., 2016). Os marcadores ISSR tem-se mostrado muito
eficiente em estudos de diversidade e variabilidade genética de microrganismos,
principalmente com o gênero Colletotrichum spp. (RAMPERSAD, 2013; SHARMA;
KATOCH, 2014; MAHMODI et al., 2014), pois não necessitam de conhecimento prévio do
DNA a ser avaliado (BARTH et al., 2002) além de ser uma técnica de baixo custo, fácil uso e
de grande reprodutibilidade (MATTHEWS et al., 1999). Os marcadores ISSR são
amplificados via PCR e não necessitam do sequenciamento da região, resultando ainda na
obtenção de padrões altamente polimórficos (NAGAOKA; OGIHARA, 1997).
Nghia et al. (2008), utilizaram o marcador ISSR para estudar isolados de Corynespora
cassiicola provenientes de folhas de seringueira na Malásia. Guozhong et al. (2004)
estudaram a diversidade de isolados endofíticos obtidos de folhas de árvores na Guiana,
através do marcador ISSR e relataram que as populações de Colletotrichum spp. foram
altamente variáveis, observando diferenças genéticas inclusive entre isolados obtidos da
mesma porção da folha da qual foram obtidos.
Marques, (2008) estudaram a diversidade genética entre isolados de Colletotrichum
spp. obtidos de cafeeiro no estado do Paraná por meio de marcadores ISSR e concluíram que
a técnica foi capaz de mostrar diversidade genética entre os isolados. Ratanacherdchai et al.
(2010) observaram alta variabilidade de C. gloeosporioides e C. capsici em três genótipos de
pimenta utilizando marcadores ISSR. Parreira et al. (2016) também utilizaram o marcador
ISSR e enfatizaram a grande variabilidade genética entre isolados de C. graminicolla isolados
de milho. Silveira (2015), concluiu em seu estudo, que os marcadores ISSR foram eficientes
na caracterização taxonômica de isolados de C. acutatum e C. gloeosporioides associados a
flores de plantas cítricas com sintomas de Podridão Floral dos Citros e que existe alta
diversidade genética entre os isolados.

28
CAPÍTULO 1
TAXONOMIA DE FUNGOS EM SERINGAIS DO SUDESTE DA BAHIA
RESUMO
Muitas espécies de fungos têm sido relatadas no ambiente dos seringais cujos papéis ecológicos
são importantes como decompositoras e antagonistas, além daquelas comprovadamente
patogênicas à seringueira, tais como: Microcyclus ulei, agente etiológico do Mal das Folhas e
Colletotrichum spp. causadoras da Antracnose. As espécies não patogênicas à seringueira
merecem ser estudadas, pois, contribuem para a reciclagem de nutrientes e afetam a
sobrevivência de patógenos, além de haver a possibilidade da existência de espécies ainda
desconhecidas para ciência e/ou ainda não relatadas em seringueira. Foram realizadas coletas
de folíolos da copa e da serapilheira, onde procedeu-se ao isolameto direto e indireto dos
fungos, com objetivo de identificar mediante a taxonomia clássica, fungos presentes em
seringais no Sudeste da Bahia. As pesquisas revelaram a ocorrência de 29 gêneros de fungos,
dentre estes, alguns patógenos da seringueira e agentes biocontroladores. Os espécimes
fúngicos coletados encontram-se depositados no herbário Cepec-Fungi da Ceplac, Bahia.
Palavras-chaves: Diversidade de fungos, Hevea brasiliensis, Ascomycota

29
ABSTRACT
Many species of fungi have been reported in the environment of the rubber plantations where
they act as decomposers and antagonists, in addition to those proven to be pathogenic to
rubber trees, such as Microcyclus ulei, that causes SALB (South American Leaf Bligth) and
Colletotrichum spp. agent of Anthracnose. The species that are not pathogenic to rubber trees
deserve to be studied because they contribute to the recycling of nutrients and affect the
survival of pathogens, besides the possibility of the existence of species still unknown for
science and / or not yet reported in rubber tree. Collection of leaf of the tree top and leaf litter
were carried out, where direct and indirect fungal isolamets were carried out, aiming to
identify, through classical taxonomy, fungi present in rubber plantations in the Southeast of
Bahia. The researches revealed the occurrence of 29 genera of fungi, among them, some
rubber pathogens and biocontrol agents. The collected fungal specimens are deposited in
Cepec-Fungi Herbarium in Ceplac, Ilhéus, Bahia.
Key words: Fungal diversity, Hevea brasiliensis, Ascomycota

30
3.1 INTRODUÇÃO
A seringueira [Hevea brasiliensis (Wild. ex Adr. de Juss.) Muell-Arg] pertencente à
família Euphorbiaceae, é a principal fonte de borracha natural do mundo (SECCO, 2008).
Nativa da bacia amazônica, bioma de clima tropical úmido, é hospedeira de uma grande
diversidade de fungos, patogênicos e não patogénicos.
A seringueira adaptou-se muito bem as condições edafoclimáticas da região Sudeste
da Bahia, onde predomina clima quente e úmido, ambiente bastante propício ao crescimento e
sobrevivência dos fungos, pois constitui-se um sistema estável apresentando características de
floresta tropical (IAC, 2017).
Estima-se que existam cerca de 1,5 milhões de espécies fúngicas no mundo
(HAWKSWORTH, 2012), no entanto, menos de 5% foram descritas. A capacidade que os
fungos têm de produzir enzimas permite que eles atuem em praticamente todos os tipos de
substratos. Dentre estes, as folhas estão entre os mais relevantes para a colonização dos fungos,
por ser um material mais facilmente degradável e encontrado em maior abundância nos
ecossistemas naturais (DIX; WEBSTER, 1995).
O estudo da diversidade de fungos nos ambientes de seringais tem crescido
significativamente. Na Tailândia, foram identificados 447 espécies de fungos, provenientes da
serapilheira e troncos de seringueiras naquela região (SEEPHUEAK et al., 2010, 2011). Cerca
de 323 espécies de fungos foram reportados em plantas de Hevea brasiliensis, a nível mundial
(FARR; ROSSMAN, 2017). Na América do Sul, foi registrada a ocorrência de 120 táxons em
H. brasiliensis, incluindo espécies patogênicas e não patogênicas, sendo a grande maioria
pertencente ao filo Ascomycota (VIÉGAS, 1961). No Brasil, cerca de 53 espécies fúngicas
foram relatadas associadas a Hevea spp. (MENDES; URBEN, 2017; EMBRAPA, 2017).
Muitos destes fungos têm papéis ecológicos importantes dentro do seringal, alguns deles
são espécies hiperparasitas, comensais, biocontroladoras de patógenos e endófitos. Outros são
comprovadamente patogênicos à seringueira, como o Microcyclus ulei (P. Henn.) v. Arx, agente
etiológico do Mal das Folhas e Colletotrichum spp. (Corda) agentes da Antracnose. Os fungos
não patogênicos associados à cultura da seringueira são estudados porque, entre outros papéis,
interferem na sobrevivência de espécies causadoras de doenças na seringueira e atuam na
ciclagem de nutrientes. Além disso, é grande a possibilidade de existirem espécies desconhecidas
para a ciência cujo potencial ainda é desconhecido.
À medida que novas áreas estão sendo exploradas para o cultivo de seringueiras, a
obtenção de informações científicas sobre a micobiota associadas a este cultivo, torna-se de

31
grande importância. Diante do incipiente conhecimento a cerca da diversidade de fungos em
seringais no Sudeste da Bahia, o presente trabalho teve como principal objetivo isolar e
identificar fungos presentes em seringais desta região.

32
3.2 MATERIAL E MÉTODOS
3.2.1 Locais de coletas
As coletas foram realizadas nos períodos de abril de 2015 a julho de 2107, em seringais
adultos das Plantações Michelin da Bahia, Estação Experimental Djalma Bahia e Área
Experimental do CEPEC, localizados nos municípios de Igrapiúna, Una e Ilhéus,
respectivamente.
Foram coletadas, aleatoriamente, amostras de folíolos maduros da serapilheira (em
decomposição), da copa e amostras de solos. O material coletado foi etiquetado, armazenado
em sacos de papel, transportados ao Laboratório de Diversidade de Fungos do Centro de
Pesquisas do Cacau (CEPEC), localizado na Comissão Executiva do Plano da Lavoura
Cacaueira (CEPLAC), município de Ilhéus-BA, onde foram feitos os isolamentos dos fungos
associados às lesões e também a sinais típicos de fungos. As amostras foram fotografadas,
secadas em uma prensa de plantas e posteriormente examinadas. Amostras representativas
foram depositadas no herbário do CEPEC/CEPLAC.
3.2.2 Isolamento dos fungos
3.2.2.1 Isolamentos direto
Os microfungos foram retirados com o auxílio de uma agulha histológica e colocados
em meio de montagem permanente contendo resina PVLG (SILVA; GRANDI, 2011) para a
caracterização morfológica, mediante a consulta de literatura especializada. As espécies
encontradas nas regiões do folíolo também foram inoculadas em placas de Petri contendo meio
de cultura Batata-Dextrose-Ágar (BDA Difco®) para obtenção de culturas axênicas. Todos os
isolados identificados foram mantidos em Castellani (1967) e incorporados à coleção de fungos
do Laboratório de Diversidade de Fungos.
3.2.2.2 Isolamento indireto
Procedeu-se a desinfestação superficial das folhas pelo método de Pereira et al. (1993).
Primeiramente, as folhas foram imersas em álcool a 70 % por 30 segundos, em seguida em
hipoclorito de sódio a 2,5% por 1 minuto e posteriormente imersas em água destilada estéril por

33
duas vezes. Logo, os fragmentos foram secos em papel toalha e depositados em placas de Petri
contendo o meio de cultura Batata-Dextrose-Ágar (BDA) juntamente com o ácido tartárico a
0,1 % para a eliminação de bactérias e isolamento somente de fungos. As placas foram
mantidas em câmara BOD com temperatura de 24±1 °C, durante 8 dias.
3.2.2.3 Isolamento de amostras de solo
Para esse procedimento utilizou-se o método de diluição seriada. Amostras de solo
obtidas da rizosfera de seringais adultos foram coletadas em quatro pontos equidistantes,
formando desse modo uma amostra composta. Em um erlemeyer adicionou-se 1 g da amostra
de solo e 9 mL de água destilada, homogeneizou (diluição 10-1
). Em seguida com uma pipeta
retirou 1 mL da solução e adicionou em 9 mL de água contida em um tubo de ensaio, diluição
de 10-2
, e a partir da diluição 10-2
retirou-se novamente 1 mL e adicionou em 9 mL de água
contida em um outro tubo de ensaio, desta forma obteve a diluição 10-3
, até a diluição 10 -8
.
Para cada amostra, foram espalhados em placas de Petri contendo BDA acidificado, 100
µL da solução. As placas foram incubadas a 24 ºC e observadas diariamente até o aparecimento
dos primeiros sinais de fungos. Logo, foram repicadas e após o crescimento da cultura pura,
feitas lâminas para posterior identificação do fungo.

34
3.3 RESULTADOS E DISCUSSÃO
3.3.1. Isolamento e identificação de fungos
Foram identificados com base nas caracterizações morfológicas 29 táxons, sendo vinte e
seis determinados em nível de espécie (Arthrobotrys arthrobotryoides, Aschersonia cubensis,
Atractilina parasitica, Cladosporium cladosporioides, Cylindrocarpon cf. candidulum,
Corynespora cassiicola, Curvularia eragrostidis, Curvularia lunata var. aeria, Dimerina
mindanaensis, Dimerosporiella cf. paulistana, Fusarium decemcellulare, Gliocladiopsis tenuis,
Hansfordia pulvinata, Irenopsis vincensii, Lasiodiplodia theobromae, Leptomeliola uvariae,
Nigrospora sphaerica, Nodulisporium ochraceum, Oidium heveae, Periconia cookei,
Pestalotiopsis suffocata, Phyllachora huberi, Spermatoloncha maticola, Spermosporella
irenopsidis sp. nov., Spiropes helleri e Trichoderma atroviride).
3.3.2. Taxonomia
1. Arthrobotrys arthrobotryoides (Berl.) Lindau (1906). Figura 1A-B.
Colônias de crescimento lento, hialinas. Conidióforos elevados sobre o substrato, não
ramificados ou ramificados ocasionalmente, 300–450 μm. Conídios elipsoides, hialinos, lisos,
com 1 septo central, 20–30 x 10,5–16 μm, clamidósporos presentes.
Material examinado: BRASIL. BAHIA: Ilhéus, Bahia, CEPLAC/CEPEC, sobre folhas em
decomposição de Hevea brasiliensis (clone SIAL 1005). 10/05/2015, T.R.Santos / J.L.Bezerra
(CEPEC 2503).
Comentários: Espécie cosmopolita comum em solos tropicais, bastante estudada no controle
biológico de fitonematoides (KENDRICK, 2000; ALEXOPOULOS et al., 1996).
2. Aschersonia cubensis Berk. & M.A. Curtis 1868, J. Linn. Soc., Bot. 10 (nº 46): 351. Figura
1C-D.
Estromas circulares, alaranjados, carnosos, pluriloculares, formados por hifas densamente
entrelaçadas, hialinas, lisas, de paredes grossas. Lóculos periféricos, subglobosos. Conidióforos
hialinos, simples ou ramificados, alongados. Fiálides delgadas, 10–20 × 1,5–2,0 μm. Paráfises
ausentes. Conídios unicelulares, hialinos, fusoides, de paredes lisas, 9–12, 5 x 3–5 μm.

35
Material examinado: BRAZIL. BAHIA: Igrapiúna, Plantações Michelin da Bahia, sobre
folhas de Hevea brasiliensis, 29/07/17, T.R. Santos e J.L. Bezerra (CEPEC 2493).
Distribuição geográfica: Taiwan, USA, Brasil.
Comentários: Espécie comum no Brasil, exercendo controle biológico natural principalmente
sobre cochonilhas em várias plantas.
3. Atractilina parasitica (G.Winter) Deighton & Piroz., Mycol. Pap. 128: 34 (1972). (Fig. 3).
Hifas hialinas a oliváceas, cilíndricas, 10–15 x 1,5–3,0 µm associadas às colônias de Irenopsis
vincensii, ramificadas, septadas, aderidas às hifas hospedeiras. Sinêmios cilindráceos, pouco
fasciculados, não ramificados, 100–584 µm de altura, de textura porrecta, 24–48 µm diâm. na
parte mediana, formados por hifas paralelas, anastomosadas, castanhas claras 1–4 µm de diâm;
Conidióforos não ramificados, 2,5–4 µm de diâmetro. Células conidiógenas poliblásticas,
simpodiais, 2–3 µm de diâmetro, retas, cilíndricas, de paredes uniformemente delgadas, com 2–
3 cicatrizes pouco conspícuas por célula, não pigmentadas. Conídios oliváceos a castanhos
claros, 26–40 x 4–6 µm, de paredes levemente rugosas na parte mediana, fusoides a levemente
obclavados, rostrados, retos ou ligeiramente recurvados, com três septos transversais; célula
distal atenuada, 10–16 µm de comprimento, capitada, até 2 µm de diâm. na parte dilatada; base
atenuada, obtusa.
Material examinado: BRAZIL. BAHIA: Igrapiúna, Plantações Michelin da Bahia, sobre
Irenopsis vincensii em jardim clonal Hevea brasiliensis, 06/08/16, T.R. Santos e J.L. Bezerra
(CEPEC 2487).
Distribuição geográfica: Panamá, Trópicos e subtropicos.
Comentário: Os caracteres do espécime estudado são típicos de A. parasitica com algumas
variações quanto às dimensões dos conídios que apresentaram dimensões menores do que as
registradas em Ellis (1971) que foi de (35–65 x 5–8 µm). Trata-se de uma espécie comumente
associada a colônias de Asterinella, Balladyna, Irenopsis e Meliola na Amazônia (ELLIS,
1971).
4. Cladosporium cladosporioides (Fresen.) G.A. de Vries, Contrib. Knowledge of the Genus
Cladosporium Link ex Fries: 57 (1952). Figura 1E.
Colônias efusas, verdes-olivas a marrons oliváceas. Conidióforos macronematosos, marrons
oliváceos, lisos, 350 x 2–6 µm. Ramoconídios 0-1 septado, oliváceos, lisos, 30 x 2–5 µm.
Conídios não septados, elipsoides, oliváceos, lisos, 3–9 x 2,5–4 µm, em cadeias longas
ramificadas.

36
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, folhas de Hevea
brasiliensis. 05/08/2016, T.R. Santos / J.L. Bezerra (CEPEC 2504).
Distribuição geográfica: Cosmopolita.
Comentários: É uma espécie bastante comum, ocorrendo como patógeno secundário em
plantas. Espécie abundante em seringueira.
5. Corynespora cassiicola (Berk. & M.A. Curtis) C.T. Wei, Mycol. Pap. 34: 5 (1950).
Colônias efusas, marrons acinzentadas, micélio superficial e também imerso; estromas
ausentes. Conidióforos marrons claros com até nove proliferações suscessivas, 100–700 x 5–10
µm. Conídios solitários ou em cadeias, obclavados a cilíndricos, retos ou incurvados, marrons
oliváceos, lisos com até 20 pseudoseptos (40–220 x 9–22 µm).
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, Associado a
Irenopsis vincensii 11/07/2016, T.R.Santos / J.L.Bezerra (CEPEC 2505).
Distribuição geográfica: Cosmopolita.
Comentários: Espécie cosmopolita muito comum nos trópicos, causando manchas foliares em
inúmeros hospedeiros (ELLIS, 1977). Patógeno de seringueira.
6. Curvularia eragrostidis (Henn.) J.A. Mey., Publ. Inst. nat. Étude agron. Congo belge, Sér.
sci. 75: 183 (1959). Figura 1F.
Conidióforos simpodiais, marrons, não ramificados. Conídios elipsoides, 3-septados, marrons,
lisos, com o septo central mais escuro, 19–33 x 10–16 µm.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, face adaxial de
folhas da copa de Hevea brasiliensis (clone 322). 29/07/2017, T.R.Santos / J.L.Bezerra
(CEPEC 2506).
Distribuição geográfica: Austrália, Ceilão, Congo, Gana, Guiné, Hong Kong, India, Jafa,
Malásia, Nigéria, Sabah, Serra Leona, Ilhas Salomão, Trinidade, USA, Brasil.
Comentários: É uma espécie polífaga, isolada de inúmeros hospedeiros vegetais e do solo.
Espécie com pouca variação quanto ao tamanho dos conídios relatados na literatura (ELLIS,
1976). Patógeno em pupunheira.
7. Curvularia lunata var. aeria (Bat., J.A. Lima & C.T. Vasconc.) M.B. Ellis, Mycology
Paper 106: 34 (1966). Figura 1G.

37
Conidióforos formando corêmios, negros, ramificados, conídios curvos, marrons, com uma
célula central mais desenvolvida e mais escura, 3-septados, lisos, não constrictos nos septos,
15–30 x 7,5–14 µm.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, face adaxial de
folhas da copa de Hevea brasiliensis (clone Fx 3864). 29/07/2017, T.R.Santos / J.L.Bezerra
(CEPEC 2507).
Distribuição geográfica: Países tropicais.
Comentários: Isolada do ar atmosférico, plantas e outros substratos. Segundo a descrição de
Ellis (1976), os conídios de C. lunata var. aeria apresentam dimensões de 18-32 x 8-16 µm, ou
seja, com pouca variação quando comparadas com as encontradas no presente estudo.
8. Cylindrocarpon cf. candidulum (Sacc.) Wollenw., Z. Parasitkde 1(1): 160 (1928). Figura
2E.
Peritécios não observados. Macroconídios hialinos, cilíndricos, retos ou encurvados, às vezes
falciformes, 50–82 x 5–7 µm, 1–4 septados, com extremidades obtusas. Microconídios
hialinos, elipsoides, unicelulares, abundantes. Fiálides ramificadas.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, folhas em
decomposição de Hevea brasiliensis (clone FDR 5788). 09/05/2016, T. R. Santos / J. L.
Bezerra (CEPEC 2508).
Distribuição geográfica: Estados Unidos, Caribe, Equador, Europa, Índia, Japão, Brasil
(presente trabalho).
Comentários: A fase teleomórfica Neonectria veuillotiana (Sacc. & Roum.) Samuels &
Mantiri não foi encontrada, por essa razão há necessidade da confirmação através de novas
coletas com a presença do teleomorfo ou por análise molecular. Não foi encontrada referência
deste táxon para o Brasil.
9. Dimerina mindanaensis (Henn.) Hansf., Mycological Paper 15: 56 (1946). Figura 2F.
Micélio fino, desenvolvido sobre as hifas de Irenopsis vincensii, reticulado, formado de hifas
subhialinas à oliváceas, espaçamente septadas, não hifopodiadas. Peritécios superficiais,
globosos, negros, ostiolados, não setosos, 70–126 x 60–110 µm; paredes pseudo
parenquimáricas, formados por uma camada de células escuras poligonais. Ascos numerosos,
clavados, bitunicados, sésseis a curto pedicilados, octospóricos (oito esporos), 50 x 12,5 µm.
Paráfises delgadas, filiformes, evanescentes. Ascósporos oblongos à subclavados, arredondados

38
nas extremindades, hialinos, lisos, 1–septados, levemente constrictos, 10–17,5 x 4–5 µm, com
célula superior, mais larga.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, folhas em
decomposição de Hevea brasiliensis. 15/01/2016, T. R. Santos / J. L. Bezerra (CEPEC 2509).
Distribuição geográfica: Uganda.
10. Dimerosporiella cf. paulistana Speg., Revista Mus. La Plata 15: 10(1908). Figura 1H.
Ascomas superficial sobre micélio de Irenopsis vincensii. Subglobosos, piriformes,
alaranjados, glabros, ostiolados, 170–212 µm de diâmetro, de paredes finas, textura
epidermoidea. Ascos clavados de ápice espessado, com anel apical. Paráfises não observadas.
Ascósporos elipsoides, hialinos, bisseriados, 1–septados, lisos, 11–16 x 3–3,5 µm.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, face abaxial de
folhas de Hevea brasiliensis (clone CDC 312). 14/06/17, T. R. Santos / J. L. Bezerra (CEPEC
2498).
Comentários: As duas espécies mais próximas pela chave de Rossmam et al., (1999) são
Dimerosporiella sensitiva (Rehm) Rossmam & Samuels e D. paulistana. A diagnose original
da primeira espécie descreve peritécios setosos, diminutos de até 60 µm, de ocorrência
conhecida apenas para o estado do Rio de Janeiro; a segunda espécie apresenta características
mais semelhantes ao material em estudo, embora os ascósporos sejam mais largos (12–14 x
4–4,5 µm) e o seu hospedeiro seja do gênero Schiffnerula Höhn, sendo uma espécie referida
apenas para o estado de São Paulo, Brasil.
11. Fusarium decemcellulare Brick, Jber. Vereinig. Angew. Bot. 6: 227 (1908). Figura 2L.
Colônias de pigmentação rosea, microconídios ovais, hialinos, 0-1 septados produzidos em
falsas cabeças ou longas cadeias, variando de 7,5–10 x 2,5–4 µm. Monofiálides e esporodóquis
presentes. Macroconídios curvos a cilíndricos com 6–10 septos, variando de 76 – 95 x 5–6 µm.
Célula apical arredondada e célula pé característica.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, face abaxial de
folhas de Hevea brasiliensis. 04/04/15, T. R. Santos / J. L. Bezerra (CEPEC 2510).
Comentários: Não é conhecida como patogênica à seringueira.
12. Gliocladiopsis tenuis (Bugnic.) Crous & M.J. Wingf., Mycol. Res. 97(4): 446 (1993).
Figura 1I.

39
Colônias marrons a marrons douradas, com odor desagradável. Vesículas ausentes.
Conidióforos penicilados, com ramos primários, secundários, terciários e quaternários.
Fiálides cilíndricas à doliformes, com colaretes presentes. Eixo central do conidióforo
terminando em uma fiálide fértil alongada frequentemente ultrapassando os ramos fialídicos.
Conídios 0–1 septados, cilíndricos, hialinos, lisos, 11–22 x 2–4 µm.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, folhas de Hevea
brasiliensis (clone Fx 3864). 24/11/2016, T. R. Santos / J. L. Bezerra (CEPEC 2501).
Distribuição geográfica: Índia, Sudeste Asiático.
Comentários: Espécie relatada em Indigofera sp., Psidium guajava, Shorea robusta, Camelia
sinensis (CROUS; WINGFIELD, 1993).
13. Hansfordia pulvinata (Berk. & M.A. Curtis) S. Hugles, Can. J. Bot. 36: 771 (1958).
Colônias acinzentadas. Conidióforos muito variáveis no tamanho 2–5 µm de espessura,
ramificados; ramos terminando em célula conidiógena apical. Células conidiogênicas
subhialinas, lisas 20 x 1,5–4 µm. Conídios esféricos ou subesféricos, marrons pálidos,
verrucosos, 6–7 µm de diâmentro.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, face abaxial de
folhas de Hevea brasiliensis. 11/08/2016, T. R. Santos / J. L. Bezerra (CEPEC 2511).
Distribuição geográfica: Cosmopolita.
Comentários: Espécie cosmopolita, hiperparasita, com potencial para uso como agente
biocontrolador de Microcyclus ulei, causador do Mal das Folhas da seringueira (MELLO et al,
2006).
14. Irenopsis vincensii D.B. Pinho & O.L. Pereira, in Pinho, Honorato, Firmino, Hora,
Mizubuti & Pereira, Tropical Plant Pathology 39(1): 90 (2014). Figura 1J.
Colônias anfígenas, coalescentes, negras. Hifas marrons escuras, septadas, ligeiramente
onduladas, hifopodiadas, marrons escuras, 22–36 x 3–6 µm. Hifopódios (apressórios) alternos,
antrorsos, retos ou ligeiramente curvos; célula do pedicelo marrom escura, cilíndricas a
cuneadas, 8–10 × 7–10 µm; células capitadas marrons escuras, arredondadas a angulosas,
raramente inteiras, globosas, oblongas a piriformes, 12–20 x 10–15 µm. Fiálides mucronadas,
marrons misturadas com os hifopódios, opostas ou alternas, 17–22 x 5–7 µm. Peritécios negros,
pouco agregados ou dispersos, globosos, verrucosos, 120–270 μm de diâmetro. Setas periteciais
marrons claras a marrons escuras, 2 a 8 por peritécio, lisas, retas, de ápice retorcido ou
uncinulado, 40–70 x 5–9 µm. Ascos evanescentes. Ascósporos hialinos quando imaturos,

40
castanhos a castanhos escuros quando maduros, oblongos, lisos contrictos no septo, 4–septados,
34–48 x 13–22 μm.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, faces adaxial e
abaxial de folhas de Hevea brasiliensis do clone CDC 312. 29/07/2016, T. R. Santos / J. L.
Bezerra (CEPEC 2483).
Distribuição geográfica: Peru e Brasil.
Comentários: Espécie bastante comum nas áreas coletadas, comumente hiperparasitada,
descritas sobre folhas de H. brasiliensis no estado de Espírito Santo e Pará, sem registro até o
momento para Bahia. Irenopsis vincensii foi proposto como novo nome para o mofo negro de
H. brasiliensis (PINHO et al., 2014).
15. Lasiodiplodia theobromae (Pat.) Griffon & Maubl. 1909. Figura 2I.
Colônias negras flocosas, picnídios estromáticos negros. Conidióforos cilíndricos hialinos;
paráfises filiformes presentes. Conídios marrons, 1-septado, estriados, 22,5 –31,4 x 13,8 – 16,6
μm.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, faces adaxial e
abaxial de folhas de Hevea brasiliensis do clone CDC 312. 29/07/2016, T. R. Santos / J. L.
Bezerra (CEPEC 2512).
Distribuição geográfica: Cosmopolita.
Comentários: É um patógeno comum em numerosos hospedeiros nos trópicos e subtrópicos
(ALVES et al., 2008). Patógeno de seringueira.
16. Leptomeliola uvariae (Rehm) S. Hughes, Mycological Papers 166: 203 (1993). Figura 1L.
Colônias anfígenas, superficiais, compostas por hifas subhialinas a marrons pálidas, com
espessura de 3–4 μm. Pseudotécios isolados, marrons, globosos a subglobosos, 190–230 ×
148–253 μm, ostiolados, setosos; Setas simples, retas, cilíndricas, ápice obtuso, septadas,
marrons escuras, 105–212 × 7–8 μm; Perídio composto por muitas camadas de células
angulares de paredes grossas. Ascos bitunicados, curto-estipitados, clavados, 8-esporos, 92–
115 × 35–45 μm; Paráfises ausentes. Ascósporos clavados, hialinos, tornando-se marrons
claros, 27,5–47,5 × 9–15 μm, 3–septados, fortemente constrictos septo mediano, com as duas
células superiores mais largas.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, faces adaxial e
abaxial de folhas de Hevea brasiliensis. 13/12/2016, T.R.Santos / J.L.Bezerra (CEPEC 2513).
Distribuição geográfica: Filipinas, Uganda, Gana, Serra Leona, Congo, Porto Rico, Brasil.

41
Comentários: O material estudado corresponde a descrição atualizada da espécie
(SILVÉRIO et al., 2011). Este é o primeiro registro de L. uvariae parasitando Irenopsis
vincensii, e também a primeira ocorrência citada para a Bahia.
17. Nigrospora sphaerica (Sacc.) E.W. Mason, Trans. Br. mycol. Soc. 12(2-3): 158 (1927).
Figura 2C-D.
Colônias cinza negras, flocosas. Micélio aéreo bem desenvolvido. Conidióforos curtos,
cilíndricos, 4–8 µm de largura. Células conidiógenas infladas, marrons claras, 8–11 µm de
diâmetro. Conídios marrons escuros, unicelulares, elipsoides a subglobosos, lisos, 14–20 µm
de diâmetro.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, sob folhas da copa
de Hevea brasiliensis (clone FDR 5788). 18/06/17, T. R. Santos / J. L. Bezerra (CEPEC 2502).
Distribuição geográfica: Cosmopolita.
Comentários: É uma espécie polífaga encontrada frequentemente em países tropicais.
Espécie com grande potencial para controle biológico de patógenos de planta (KIM et al.,
2001).
18. Nodulisporium ochraceum Preuss, Klotzschii Herb. Viv. Mycol.: nº. 1272 (1849). Figura
2A-B.
Colônias efusas, marrons a negras, velutinas, micélio parcialmente superficial, estromas
ausentes, setas e hifopódios ausentes, conidióforos macronematosos, não sinematosos,
ramificados próximo ao ápice, flexuosos, marrons e lisos. Células conidiógenas poliblásticas,
íntegradas, simpodiais, denticuladas. Conídios solitários, secos, acropleurógenos, simples,
unicelular, elipsoides, hialinos, lisos, 0-septados, 3,1–9,5 x 2,3–4,5 μm.
Material examinado: Ilhéus, Bahia, CEPLAC/CEPEC, sobre folhas em decomposição de
Hevea brasiliensis (clone SIAL 1005). 12/01/17, T. R. Santos / J. L. Bezerra (CEPEC 2500).
Distribuição geográfica: Cosmopolita.
Comentários: Este táxon corresponde ao morfo assexuado de Xylariaceae (ELLIS, 1971; JU e
ROGERS, 1996).
19. Periconia byssoides Pers., Syn. meth. fung. (Göttingen) 1: 18 (1801).
Colônias efusas, marrons compactas, micélio parcialmente imerso e parcialmente superficial;
estromas não observados em cultura; hifopódios ausentes; conidióforos macronematosos,
estiptados com a cabeça esférica, não ramificados, 470–700 µm; células conidiogênicas

42
elipsoides, distintas; conídios catenulados, subglobosos, marrons, verrucosos, variando de 12–
16 x 12–17 µm.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, sob folhas da copa
de Hevea brasiliensis. 09/07/2017, T. R. Santos / J. L. Bezerra (CEPEC 2514).
Distribuição geográfica: Cosmopolita.
Comentários: Espécie extremamente comum, em diversos hospedeiros e substratos.
20. Pestalotiopsis suffocata (Ellis & Everh.) J.G. Wei & T. Xu, Wei, Xu, Guo & Pan,
Mycosystema 24(4): 488 (2005). Figura 2G.
Conídios cinco celulares, fusiformes, estreitos, Concolor, 18–28 x 5–7 µm; células
intermediárias uniformemente oliváceas; apêndices apicais em número de três, 28–30 µm.
Material examinado: BRASIL. BAHIA: Cepec, Ilhéus sob folhas da copa de Hevea
brasiliensis. 29/11/2016, T. R. Santos / J. L. Bezerra (CEPEC 2515).
Distribuição geográfica: Texas e Brasil.
Comentários: Esta espécie pertence à secção Quinqueloculatae de GUBA (1961), que utiliza o
nome Pestalotia ao invés de Pestalotiopsis. Este autor separa as espécies em base
morfológicas, levando em consideração forma e tamanho de conídios e dos apêndices conidiais.
Este critério tem sido questionado em função de estudos moleculares modernos (YU et al.,
2013; LIU et al., 2017).
21. Phylachora huberi Henn., Hedwigia 39 (Beibl.): (78) 1900.
Estromas hipófilos, negros, até 1 cm de diâmetro. Peritécios elipsoides, 280–380 x 70–135
µm. Clípeo negro de 60–70 µm, intraepidérmico. Ascos clavados, unitunicados, 8 esporos,
50–65 x 15–20 µm. Paráfises filiformes. Ascósporos oblongos a elipsoides, hialinos,
unicelulares variando de 14–18 x 8–10 µm.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, sob folíolos de
Hevea brasiliensis em viveiro. 09/02/2017, T. R. Santos / J. L. Bezerra (CEPEC 2516).
Comentários: Patógeno de seringueira.
22. Trichoderma atroviride P. Karst., Bidr. Kann. Finl. Nat. Folk 51:363 (1892). Figura 2H.
Colônias esverdeadas de crescimento rápido. Conidióforos ramificados crucialmente. Células
conidiógenas ampuliformes. Conídios verdes escuros, subglobosos, lisos, 2–3,5 x 2–3 µm, odor
característico, semelhante a coco ralado.
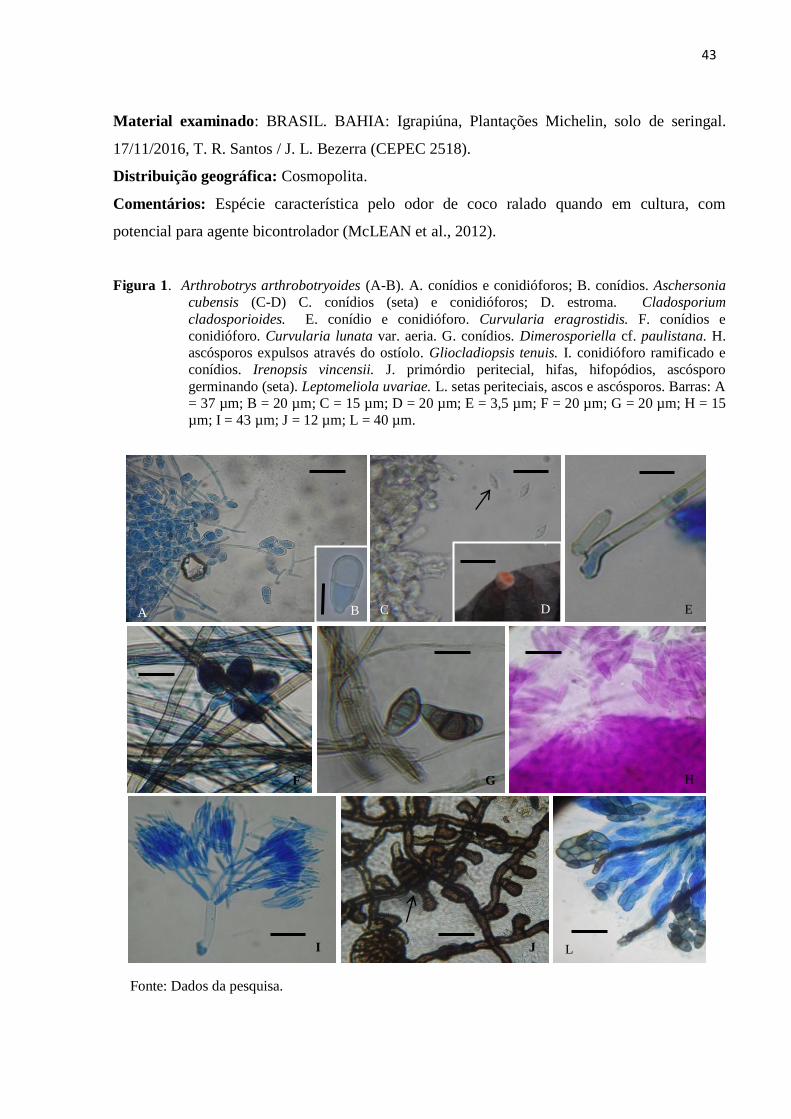
43
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin, solo de seringal.
17/11/2016, T. R. Santos / J. L. Bezerra (CEPEC 2518).
Distribuição geográfica: Cosmopolita.
Comentários: Espécie característica pelo odor de coco ralado quando em cultura, com
potencial para agente bicontrolador (McLEAN et al., 2012).
Figura 1. Arthrobotrys arthrobotryoides (A-B). A. conídios e conidióforos; B. conídios. Aschersonia
cubensis (C-D) C. conídios (seta) e conidióforos; D. estroma. Cladosporium
cladosporioides. E. conídio e conidióforo. Curvularia eragrostidis. F. conídios e
conidióforo. Curvularia lunata var. aeria. G. conídios. Dimerosporiella cf. paulistana. H.
ascósporos expulsos através do ostíolo. Gliocladiopsis tenuis. I. conidióforo ramificado e
conídios. Irenopsis vincensii. J. primórdio peritecial, hifas, hifopódios, ascósporo
germinando (seta). Leptomeliola uvariae. L. setas periteciais, ascos e ascósporos. Barras: A
= 37 µm; B = 20 µm; C = 15 µm; D = 20 µm; E = 3,5 µm; F = 20 µm; G = 20 µm; H = 15
µm; I = 43 µm; J = 12 µm; L = 40 µm.
Fonte: Dados da pesquisa.
I J
L
L
A B D C E
F
G
H

44
Figura 2. Nodulisporium ochraceum (A-B) A. conídios; B. conidióforos. Nigrospora sphaerica
(C-D) C. conídios; D. célula conidiogênica. Cylindrocarpon cf. candidulum. E.
conídios e células conidiogênicas. Dimerina mindanaensis. F. peritécio rompido, ascos
e ascósporos. Pestalotiopsis suffocata. G. conídio. Trichoderma atroviride. H.
conidióforo e conídios. Lasiodiplodia theobromae. I. conídios uniseptados. Spiropes
helleri. J. conídio. Fusarium decemcellulare. L. conídios. Barras: A = 12 µm, inset = 14
µm; C = 20 µm; E = 50 µm; F = 25 µm; G = 5 µm; H = 10 µm; I = 25 µm; J = 14 µm; L
= 23 µm.
Fonte: Dados da pesquisa.
A B C ED
F G H
I J L
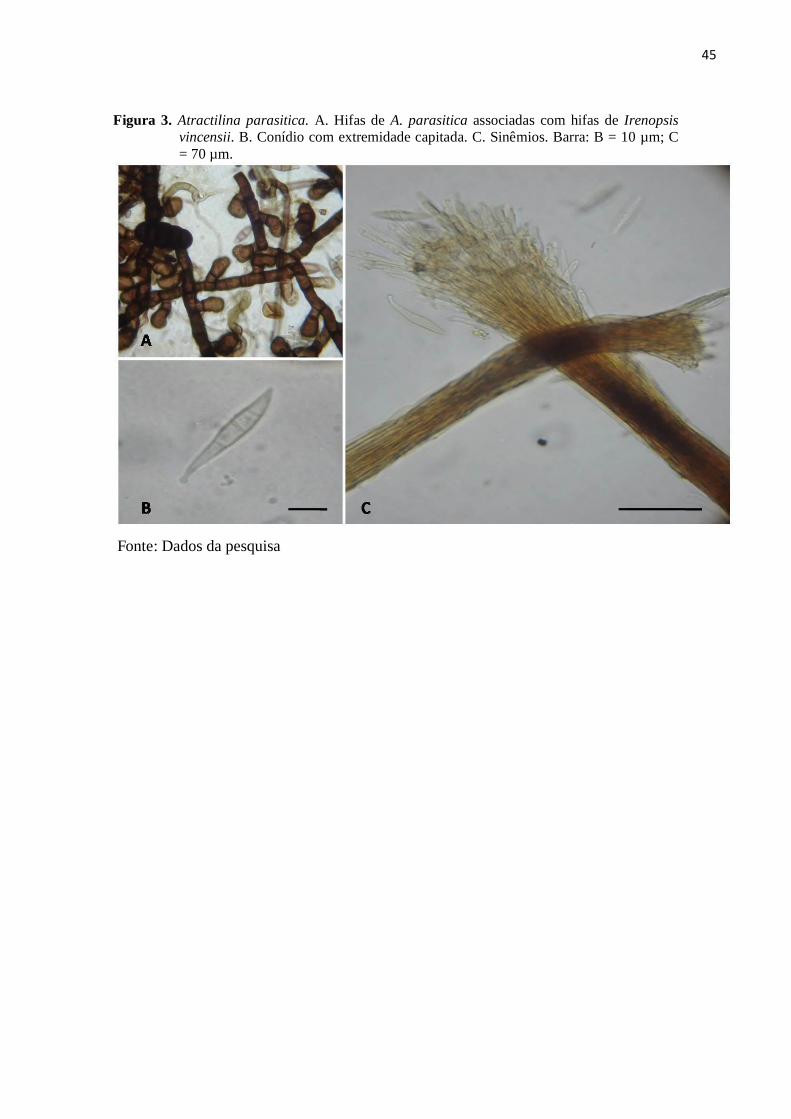
45
Figura 3. Atractilina parasitica. A. Hifas de A. parasitica associadas com hifas de Irenopsis
vincensii. B. Conídio com extremidade capitada. C. Sinêmios. Barra: B = 10 µm; C
= 70 µm.
Fonte: Dados da pesquisa

46
3.4 REFERÊNCIAS BIBLIOGRÁFICAS
ALEXOPOULOS, C.J.; MIMS, C.W.; BLACKWELL, M. Introductory Micology. 4th
edition. New York, John Wiley e Sons, Inc. 1996.
ALVES, A.; CROUS, P.W.; CORREIA, A.; PHILLIPS, A. Morphological and molecular
data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Diversity.28: 1-13. 2008.
BERKELEY, M.J.; CURTIS, M.A. Fungi Cubenses (Hymenomycetes). Journal of the
Linnean Society.10:280-392. 1869.
BRICK, J. Vereinig. Angew. Bot. 6: 227, 1908.
CROUS, P.W.; WINGFIELD, M.J.A re-evaluation of Cylindrocladiella, and a comparison
with morphologically similar genera. Mycological Research. 97(4):433-448. 1993.
DEIGHTON, F.C.; PIROZYNSKI, K.A. More hyperparasitic hyphomycetes. Microfungi
V.Mycological Papers. 128:1-110. 1972.
DIX, N.J.; WEBSTER, J. Fungal Ecology. London, Chapman & Hall. 1995.
ELLIS, M.B. Dematiaceous hyphomycetes. Kew, Common wealth Mycological Institute.
1971.
ELLIS, M.B. Dematiaceous hyphomycetes.VII. Curvularia, Brachysporium, etc.
Mycological Papers. 106:1-571966.
ELLIS, M.B. Dematiaceous hyphomycetes. IX. Spiropes and Pleurophragmium.
Mycological Papers. 114:1-44. 1968.
EMBRAPA, 2017. Disponível em:
<http://pragawall.cenargen.embrapa.br/aiqweb/michtml/micbanco01a.asp>. Acesso: 8 dez. 2017.
FARR, D.F.; ROSSMAN, A.Y. Fungal Databases. Systematic Mycology and Microbiology
Laboratory, ARS, USDA. Disponível em: <from http://nt.ars-grin.gov/fungaldatabases/>.
Acesso em: 22 out. 2017. 2017.
GRIFFON, E.; MAUBLANC, A. Sur unemaladie du cacaoyer. Bulletin de La Société
Mycologique de France. 25:51-58. 1909.
GUBA, E.F. Monograph of Pestalotia and Monochaetia. Harvard University Press.
Cambridge, Massachusetts USA, 1961, 342 p..
HANSFORD, C.G. The foliicolous Ascomycetes, their parasites and associated fungi.
Mycological Papers. 15:1-240. 1946.
HAWKSWORTH, D.L. Global species numbers of fungi: are tropical studies and
molecular approaches contributing to a more robust estimate? Biodiversity and
Conservation 21: 2425-2433. 2012.
HENN, Hedwigia 39 (Beibl.): (78) 1900.

47
HUGHES, S.J. Revisiones Hyphomycetum aliquot cum appendice de nominibus
rejiciendis. Canadian Journal of Botany. 36(6):727-836.1958.
HUGHES, S.J. Meliolina and its excluded species. Mycological Papers. 166:1-255. 1993.
IAC, 2017. Disponível em: <http://www.iac.sp.gov.br/areasdepesquisa/seringueira/ > Acesso
em: 8 dez. 2017.
JU, M.Y.; ROGERS, J.D. A revision of the genus Hypoxylon. Edição.St. Paul, Minn.: APS
Press, the American Phytopathological Society, 365 p. 1996.
KARSTEN, P.A. Finland smögels vampar (Hyphomyctes fennici). Bidrag till Kännedomav
Finlands Naturoch Folk. 51:343-534. 1892.
KENDRICK, B. The Fifth Kingdom. 3rd edition. Newburyport, Focus Publishing. 2000.
KIM, J. C.; CHOI, G.J.; PARK, J.H.; CHO, K. Y. Activity against plant pathogenic fungi
of phomalactone isolated from Nigrospora sphaerica. Pest Management Science . 57 (6):
554-559. 2001.
LINDAU, G. Rabenhorst's Kryptogamen-Flora. Pilze-Fungi imperfecti. 1(8):177-432.
1905.
LIU, F.; HOU, L.; RAZA, M.; CAI, L. Pestalotiopsis and allied genera from Camellia, with
description of 11 new species from China. Scientific Reports. 7:866. doi:10.1038/s41598-
017-00972-5. 2017.
MASON, E.W. On species of the genus Nigro-spora Zimmermann recorded on
monocotyledons. Transactions of the British Mycological Society. 12(2-3):152-165. 1927.
MCLEAN, K.L.; HUNT,J. S.; STEWART, A.; WITE, D.; PORTER, I.J.; VILLALTA, O.
Compatibility of a Trichoderma atroviride biocontrol agent with management practices
of Allium crops. CropProtection. Vol. 33. Pp. 94-100. 2012.
MELLO, S.C. M.; SANTOS, M.F.; SILVA, J. B.T. Isolados de Dicyma pulvinata em
estromas de Microcyclus ulei em seringueira. Pesquisa Agropecuária
Brasileira, Brasília, v. 41, n. 2, p. 359-363. 2006.
MENDES, M.A.S.; URBEN, A.F. Fungos relatados em plantas no Brasil. Laboratório de
Quarentena Vegetal. Brasília, DF: Embrapa Recursos Genéticos e Biotecnologia. Disponível
em: <http://pragawall.cenargen.embrapa.br/aiqweb/michtml/fgbanco01.asp.>. Acesso em: 13
nov. 2017. 2017.
MEYER, J.A. Moisissuresdu sol et deslitières de larégion de Yangambi (Congo Belge).
Publications de l'Institut Agronomiquedu Congo Belge. 75:1-211. 1959.
PEREIRA, J.O.; AZEVEDO, J.L.; PETRINI, O. Endophytic fungi of Stylosantes.
Micologya, v.85, p. 362-364. 1993.
PERSOON, C.H. Synopsis methodica fungorum. 1-706. 1801.

48
PINHO, D.B.; JUNIOR HONORATO, J.; FIRMINO, A.L.; JUNIOR HORA, B.T.;
MIZUBUTI, E.S.G.; PEREIRA, O.L. Reappraisal of the black mildews (Meliolales) on
Hevea brasiliensis. Tropical Plant Pathology, vol.39. Pp 89-94. 2014.
PREUSS. Klotzsch Herb. Mycol: no. 1272, 1849.
ROSSMAM, A.Y.; SAMUELS, G.J.; ROGERSON, C.T.; LOWEN, R. Genera of
Bionectriaceae, Hypocrateaceae and Nectriaceae (Hipocreales, Ascomycetes). Studies of
Mycology. V. 42, 248p. 1999.
SECCO, R.S.de. A botânica da seringueira (Hevea brasiliensis (Wild. ex. Adr. Jussieu) Muell.
Arg.(Euphorbiaceae). IN: ALVARENGA, A.P.; CARMO, C.A.F. de S. (Cood.) Seringueira.
Viçosa, MG, EPAMIG-Viçosa, 2008, 3-24p.
SEEPHUEAK, P.; PHONGPAICHIT, S.; HYDE, K.; PETCHARAT, V. Diversity of
saprobic fungi on decaying rubber logs (Hevea brasiliensis). Sydowia. 63. 249-282. 2011.
SEEPHUEAK, P.; PETCHARAT, V.; PHONGPAICHIT, S. Fungi associated with leaf
litter of rubber (Hevea brasiliensis). Mycology. 1. 213-227. 2010.
SILVA, P.da.; GRANDI, R.A.P. A new species of Thozetella (anamorphic fungi) from
Brazil.Cryptogamie Mycologie, 32(4): 359-363. 2011.
SILVÉRIO, M. L.; CAVALCANTI, M. A. Q.; BEZERRA, J. L. First record of
Leptomeliola uvariae for South America. Mycotaxon, v. 116, p. 1-5, 2011.
SPEGAZZINI, C. Fungi aliquot paulistani. Revista del Museo de La Plata. 15:7-48. 1908.
SPEGAZZINI, C. Hongos de layerba mate. Anales Del Museo Nacional de Historia Natural
Buenos Aires. 17(10):111-141. 1908.
STEINMANN, A. De ziekten em plagen van Hevea brasiliensis in Nederlandsch-Indië. 1-
90. 1925.
VIÉGAS, A.P. Índice de Fungos da América do Sul. Campinas, Instituto Agronômico.
1961.
VRIES, G.A. de. Contribution to the knowledge of the genus Cladosporium. 1-121p. 1952.
WEI, C.T. Notes on Corynespora. Mycological Papers. 34:1-10. 1950.
WEI, J.G.; XU, T. Mycosystema. 24(4): 488, 2005.
WOLLENWEBER, H.W. Über Fruchtformen der krebserregenden Nectriaceen.
Zeitschrift für Parasitenkunde. 1:138-173. 1928.
YU, S.; KUN, G.; HYDE, K.D.; ZHAO, W.S.; WEI, J.; KANG, J.; WANG, Y. Two new
species of Pestalotiopsis from Southern China. Phytotaxa. Auckland, New Zealand:
Magnolia Press. 126 (1): 22–30. 2013.

49
CAPÍTULO II
4. Spermosporella irenopsidis sp. nov. and Spermatoloncha maticola, parasitic on black
mildew (Irenopsis vincensii) of rubber in Bahia, Brazil
Artigo submetido à revista Rodriguésia
TACILA RIBEIRO SANTOS1, EDNA DORA MARTINS NEWMAN LUZ
2, HARRY C.
EVANS3, JOSÉ LUIZ BEZERRA
1*
1 Universidade Estadual de Santa Cruz, PPGPV, Ilhéus, BA, Brasil.
2 Comissão Executiva do Plano da Lavoura Cacaueira, Ceplac/Cepec, Km 22, Rod.
Ilhéus/Itabuna Caixa Postal 7, Cep: 45.600-000, BA, Brazil.
3 CAB International, Egham, Surrey TW20 9TY, U.K.
*Corresponding author: José Luiz Bezerra, [email protected].
Abstract
A new species of the genus Spermosporella and a new record of Spermatoloncha maticola are
reported parasitizing the black mildew (Irenopsis vincensii) on leaves of Hevea brasiliensis in
the Bahia, Brazil. Both species are described and illustrated and all Spermosporella species
are compared.
Key words
Fungicolous fungi, Hevea brasiliensis, Meliolales, taxonomy.
4.1 Introduction
In Brazil, commercial rubber Hevea brasiliensis (Willd. ex A.Juss.) Müll. Arg., is
attacked by several pathogenic fungi. Among them, Irenopsis vincensii D.B. Pinho & O.L.
Pereira causes black mildew disease of rubber. Irenopsis vincensii was reported and well
documented by Pinho et al. (2014) in the Brazilian states of Espírito Santo and Pará having as
heterotypic synonyms the binomials Meliola heveae Vincens and Irenopsis heveae Hansf.
During a survey of fungi on rubber trees, conducted in August 2016 in Michelin
Plantations of Bahia, municipality of Igrapiúna, leaf samples were collected showing colonies
of the black mildew (I. vincensii).

50
In the laboratory, it was found that I. vincensii colonies were parasitized by two
fungicolous fungi, corresponding to Spermatoloncha maticola Speg. and a new species of the
genus Spermosporella Deighton.
The genus Spermosporella is a known parasite of Meliola Fr. species in Africa, Asia
and North America (Deighton 1969; Deighton & Pirozinsky 1972; Sutton 1973; Hawksworth
1981; McKenzie 1992; Hughes 1993; Matsushima & Matsushima 1995; Seifert & Gam,
2011), but has not yet been recorded on the genus Irenopsis F. Stevens. Four species of the
genus Spermosporella are known (Deighton 1969; Deighton & Pirozinsky 1972; Sutton 1973;
Matsushima & Matsushima 1995) which differ morphologically from the species studied.
The monotypic species Spermatoloncha maticola has been reported in association
with Meliola clerodendricola Henn. on leaves of IIex paraguariensis A. St.-Hil. (yerba maté)
in Argentina (Spegazzini 1908; Hawksworth 1981) and later it was found in Uganda on
Clerodendrun capitatum (Willd.) Schumcch. & Thonn. parasitizing the same species of
Meliola (IMI 102852b).
Species of the Meliolales are subject to parasitism by several genera of fungicolous
fungi. Phaeostigme Syd. & P. Syd; Eriocercospora Deighton, Ramichloridium Stahel ex de
Hoog and Spiropes Cif. have all been reported from Brazil (Batista et al. 1966; Caliman
2015). However, there are no references to Spermosporella and Spermatoloncha associated
with Meliolales in Brazil.
Here, we describe a new species of the genus Spermosporella and report a new record
of Spermatoloncha maticola associated with Irenopsis vincensii on Hevea brasiliensis leaves
in Bahia, Brazil.
4.2 Materials and Methods
Leaf samples from the rubber tree clone CDC 312, colonized by a black mildew in a
clonal garden located in Michelin Plantations of Bahia, Igrapiúna city, Bahia, Brazil, were
collected in August 2016.
In the Fungal Diversity Laboratory, at the Cocoa Research Center (Cepec),
municipality of Ilhéus, Bahia, the material was examined under a stereoscopic microscope
(MOTIC SMZ -168). Reproductive and somatic structures of the fungi associated with the
black mildew were morphologically characterized and photographed using a LeicaDM500
microscope coupled with a digital camera Sony Cyber-shot DSC-WX30. The measurements
of the structures were performed using a calibrated ocular micrometer. For observation of

51
intact mycelia, a modified Callan & Carris (2004) technique was used, with the cellulose
acetate being replaced with fast drying colorless nail enamel (Risqué Technology), and
followed by mounting in PVLG (polyvinyl alcohol + lactic acid + glycerol) (Hosagoudar &
Kapoor 1985). The micrographs were done by camera lucida drawings, copied on drawing
paper with India ink and scanned.
For taxonomic identification, keys and descriptions of genera and species of
mycoparasitic fungi on Meliolales were consulted in the literature (Deighton 1969; Deighton
& Pirozinsky 1972; Sutton 1973; McKenzie 1992; Matsushima & Matsushima 1995; Seifert
& Gam, 2011). Samples were deposited in the CEPEC-Fungi Fungarium (Ceplac, Ilhéus-
Bahia, Brazil).
4.3 Results
4.3.1 Taxonomy
1. Spermosporella irenopsidis J.L. Bezerra, T. R. Santos, E.D. Luz sp. nov. (Figs. 1 and 2a-b).
MycoBank 819220
Etymology: Referring to the host fungus Irenopsis.
Diagnosis: Colonies whitish, effuse. Mycelium of repent, hyaline, septate hyphae, 2–2.5 µm
diam. Conidiogenous cells intercalary, continuous, 1-celled, ampulliform, 8–20 x 4–8 µm.
Conidia hyaline, smooth, fusiform, straight to slightly curved, 3-septate, attenuated at both
ends, 24–32 x 2–3 µm.
Whitish colonies, effuse, thin, overgrowing colonies of I. vincensii. Superficial
mycelium composed of repent, hyaline, smooth, septate, branched hyphae, 2–2.5 µm in diam.
Conidiogenous cells produced by intercalary growth in the hyphae; hyaline, unicellular,
simple, smooth, ampulliform, measuring 8–20 x 4–8 µm with an attenuated apical beak where
the spores are formed. Scars not observed. Conidia hyaline, smooth, thin-walled, fusiform,
straight to slightly bent, 3-septate, not constricted at the septa, 24–32 x 2–3 µm, attenuated at
both ends.
Material examined: BRAZIL. BAHIA: Igrapiúna, Michelin Plantations of Bahia, on Hevea
brasiliensis leaves (clone CDC 312). 29.07.2016, Santos et al. (CEPEC, Holotype 2483).
Comments: Spermosporella aggregata Deighton (Tab. 1) is the closest species to S.
irenopsidis which is distinguished by having intercalary conidiogenous cells and not rostrate,
recurved and constricted conidia.

52
TABLE 1. Comparative morphology of species of Spermosporella, including the new species, S. irenopsidis.
Species
Conidial shape
Septation
Conidial size (µm)
Conidiogenous cells
(µm)
Host
Country
Reference
S. aggregata Deighton Fusoid-
obclavate; 2–3 17–29 x 3.5-4 6–8 x 2–4
Meliola geniculata var.
eggelingii
Sierra
Leone
Deighton,
1969
S. elegans Sutton Obclavate-
elongate 6–9 41.5–78.5 x 3.5-4 3.5–5 x 2.5–3.5 Meliola spp. Canada Sutton, 1973
S. flagelliformis Matsush. &
Matsush.
Cylindrical-
caudate 7–12 50–180 x 4–5.5 18 x 2–2.5
Pleurothecium
recurvatum Japan
Matsushima
&
Matsushima,
1995
S. pulvinata Deighton &
Piroz.
Obclavate-
narrow 2 25–40 x 2.5–3 6–12 x 2.5–3 Meliola deinbolliae
Sierra
Leone
Pirozynski,
1972
S. irenopsidis nov.sp. Fusiform 3 24–32 x 2–3 8–20 x 4–8 Irenopsis vincensii Brazil This work
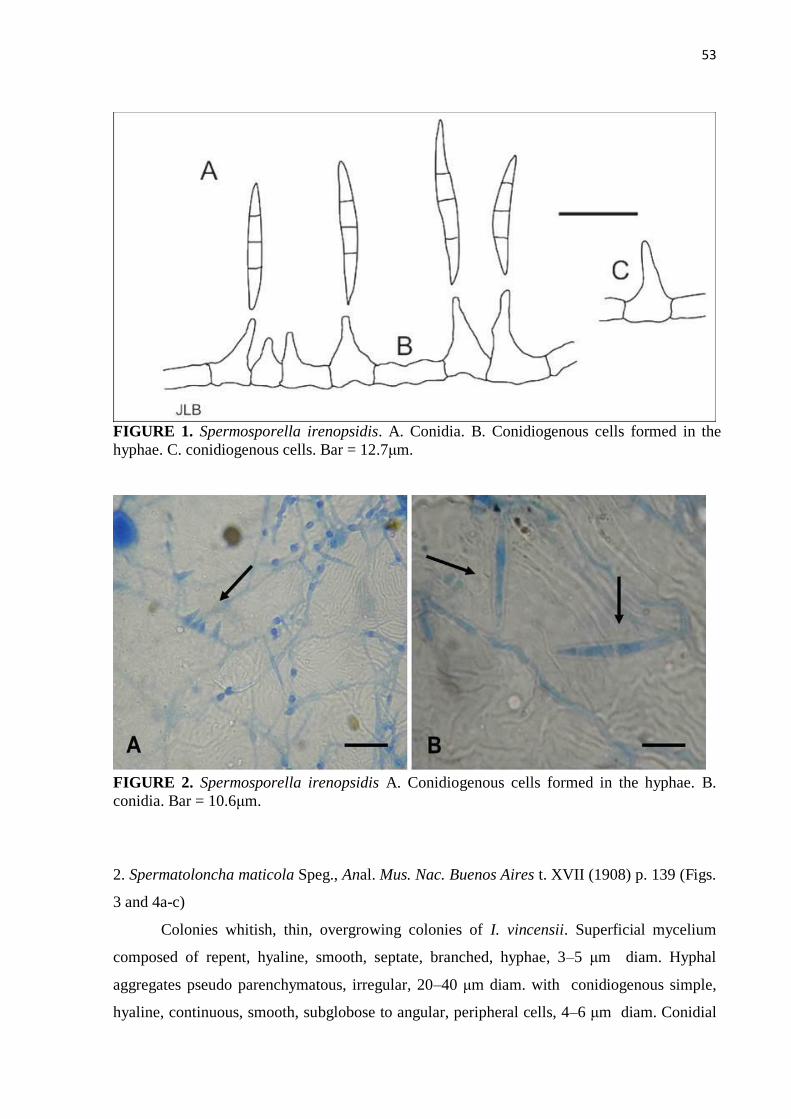
53
FIGURE 1. Spermosporella irenopsidis. A. Conidia. B. Conidiogenous cells formed in the
hyphae. C. conidiogenous cells. Bar = 12.7μm.
FIGURE 2. Spermosporella irenopsidis A. Conidiogenous cells formed in the hyphae. B.
conidia. Bar = 10.6μm.
2. Spermatoloncha maticola Speg., Anal. Mus. Nac. Buenos Aires t. XVII (1908) p. 139 (Figs.
3 and 4a-c)
Colonies whitish, thin, overgrowing colonies of I. vincensii. Superficial mycelium
composed of repent, hyaline, smooth, septate, branched, hyphae, 3–5 μm diam. Hyphal
aggregates pseudo parenchymatous, irregular, 20–40 μm diam. with conidiogenous simple,
hyaline, continuous, smooth, subglobose to angular, peripheral cells, 4–6 μm diam. Conidial
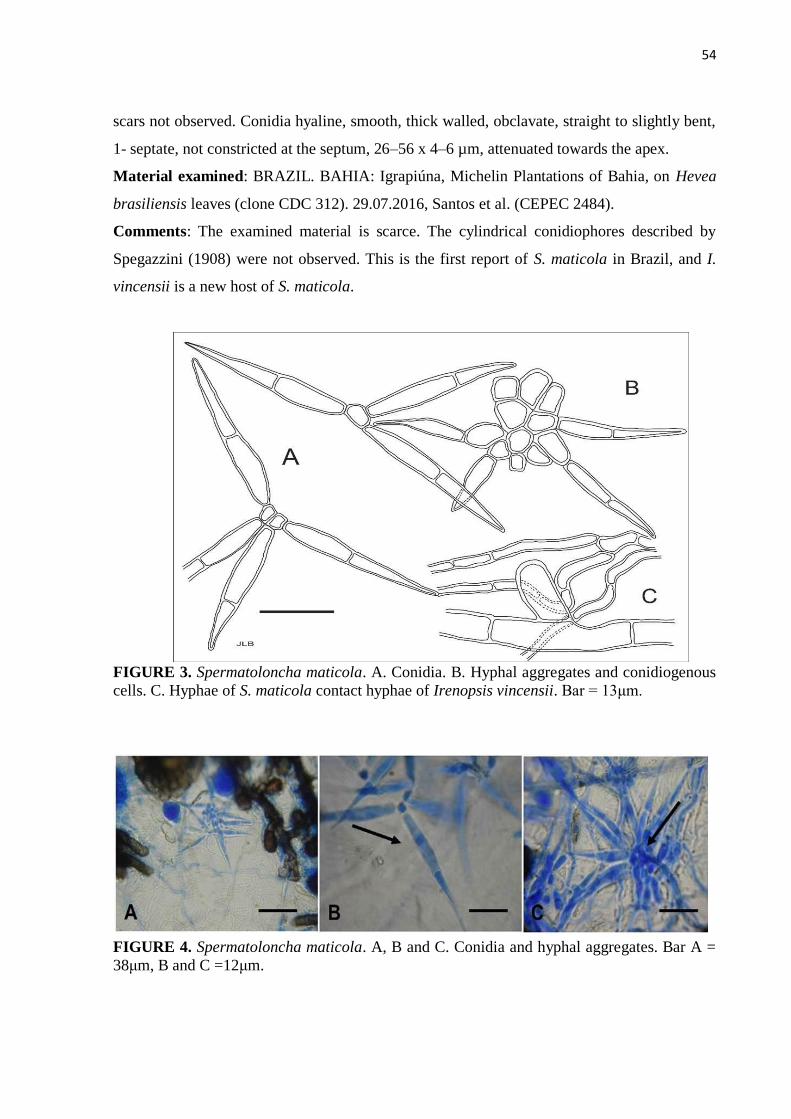
54
scars not observed. Conidia hyaline, smooth, thick walled, obclavate, straight to slightly bent,
1- septate, not constricted at the septum, 26–56 x 4–6 µm, attenuated towards the apex.
Material examined: BRAZIL. BAHIA: Igrapiúna, Michelin Plantations of Bahia, on Hevea
brasiliensis leaves (clone CDC 312). 29.07.2016, Santos et al. (CEPEC 2484).
Comments: The examined material is scarce. The cylindrical conidiophores described by
Spegazzini (1908) were not observed. This is the first report of S. maticola in Brazil, and I.
vincensii is a new host of S. maticola.
FIGURE 3. Spermatoloncha maticola. A. Conidia. B. Hyphal aggregates and conidiogenous
cells. C. Hyphae of S. maticola contact hyphae of Irenopsis vincensii. Bar = 13μm.
FIGURE 4. Spermatoloncha maticola. A, B and C. Conidia and hyphal aggregates. Bar A =
38μm, B and C =12μm.

55
4.4 Discussion
Spermosporella is biologically and morphologically very similar to
Annellospermosporella P.R. Johnst. The two genera are distinguished by sympodial
conidiogenesis in Spermosporella and percurrent-anellidic conidiogenesis in
Annellospermosporella. Thus far, no molecular studies have been done with the genera
Spermosporella and Spermatoloncha and hence there are no DNA sequences available.
Because neither of these genera have been cultured in vitro, DNA extraction depends on
processing fungal fragments from fresh specimens. However, during the present study,
attempts to extract DNA were unsuccessful.
4.5 Acknowledgements
Thanks are due to Comissão Executiva do Plano da Lavoura Cacaueira (Ceplac),
Bahia, Brazil, Michelin Plantations of Bahia, Brazil and the Universidade Estadual de Santa
Cruz, Bahia, Brazil for research support; to Marília Leniuza Soares Ribeiro for the
illustrations; to the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for financial
support/scholarships to J.L. Bezerra, T. R. Santos and E.D.M.N. Luz; and, finally, to Armínio
Santos for help with the literature search.
4.6 References
Batista, A.C; Bezerra, J.L.; Maia, H.da. S. & Garnier, R. 1966. Alguns hiperparasitas
de Meliolaceae e outros Ascomycetes. In: Instituto de Micologia, Universidade do
Recife 470 (Atas do Instituto de Micologia). Universidade Federal de Pernambuco, Recife.
Vol. 3. pp. 10-30.
Calimam, M.E. 2015. Aspectos ecológicos e taxonômicos de Prillieuxina winteriana
(Asterinaceae) e sua interação com Annona muricata. In: Dissertação de mestrado.
Universidade Estadual de Santa Cruz, Ilhéus, Bahia. 60p.
Callan, B.E. & Carris, L.M. 2004. Fungi on living plant substrata, including fruits. In: Mueller, G.M.; Bills, G.F. & Foster, M.S. (eds), Biodiversity of Fungi–Inventory and
Monitoring Methods, Elsevier, California. Pp 105-126.
Deighton, F.C. 1969. Microfungi IV: Some hyperparasitic Hyphomycetes, and a note on
Cercosporella uredinophila Sacc. Mycological Papers. Vol.118. Pp 1-41.

56
Deighton, F.C. & Pirozynski, K.A. 1972. Microfungi V. More hyperparasitic Hyphomycetes.
Mycological Papers. Vol.128. Pp 1-110.
Hawskswork, D.L. 1981. A survery of the fungicolous conidial fungi. In: Cole, G.T &
Kendrik, B. (eds), Biology of Conidial Fungi, Vol. I. Academic Press, New York. Pp 171-
244.
Hosagoudar, V.B & Kapoor, J.N. 1985. New technique of mounting Meliolaceous fungi.
Indian Phytopathology. Vol. 38. Pp 548-549.
Hughes, S.J. 1993. Meliolina and its excluded species. Mycological Papers. Vol.166. Pp 1-
255.
IMI 102852b. Available from: http://www.herbimi.info/herbimi/home.htm
Accessed: 19 January 2017.
Matsushima, K. & Matsushima, T. 1995. Fragmenta Mycologica - I. Matsushima.
Mycological Memoirs. Vol.8. Pp 45-54.
McKenzie, E.H.C. 1992. Fungi of the Kermadec Islands. Mycotaxon. Vol.45. Pp 149-170.
Pinho, D.B.; Junior Honorato, J.; Firmino, A.L.; Junior Hora, B.T.; Mizubuti, E.S.G. &
Pereira, O.L. 2014. Reappraisal of the black mildews (Meliolales) on Hevea brasiliensis.
Tropical Plant Pathology. Vol.39. Pp 89-94.
Pirozynski, K.A. 1972. Microfungi V. More hyperparasitic hyphomycetes. Mycological
Papers. Vol. 128. Pp 1-110.
Seifert, K.A. & Gams, W. 2011. The genera of Hyphomycetes – 2011 update. Persoonia. Vol.
27. Pp 119-129.
Sutton, B.C. 1973. Hyphomycetes from Manitoba and Saskatchewan, Canada. Mycological
Papers. Vol.132. Pp 1-143.
Spegazzini, C.L. 1908. Hongos de layerba mate. Anales del Museo Nacional de Historia
Natural de Buenos Aires Series 3, Vol.17. Pp 111-141.

57
CAPÍTULO III
5. CARACTERIZAÇÃO MORFOCULTURAL, MOLECULAR E DIVERSIDADE
GENÉTICA DE Colletotrichum spp. AGENTE CAUSAL DA ANTRACNOSE FOLIAR
EM SERINGUEIRA (Hevea brasiliensis)
RESUMO
A incidência de espécies do gênero Colletotrichum tem sido amplamente relatada como um
dos principais fitopatógenos que acometem a seringueira (Hevea brasiliensis), causando
grandes perdas na cadeia produtiva da borracha. Devido à grande diversidade de espécies
presentes no patossistema Colletotrichum spp. x H. brasiliensis, os estudos da diversidade
morfológica, genética e a identificação de isolados em nível de espécies, contribuem para as
ações de manejo nas estratégias de controle e melhoramento para a resistência à Antracnose.
Neste contexto, o presente trabalho objetivou a caracterização morfocultural e molecular em
29 isolados de Colletotrichum isolados de folíolos de seringueira. Os isolados foram
identificados em nível de espécie com a amplificação e sequenciamento comparativo de suas
sequências genômicas ITS1/4 e o gene GAPDH. A análise da diversidade genética foi
avaliada pela obtenção dos marcadores moleculares Inter-Simple Sequence Repeats (ISSR).
A avaliação morfológica das colônias revelou heterogeneidade em todos os descritores
analisados, dificultando a certificação em nível de complexo e espécie. Contudo, a
identificação das espécies neste patossistema revelou a presença de cinco espécies
pertencentes a três complexos: C. gloeosporioides, C. acutatum e C. boninense. No grupo
amostral, o maior número de espécies agruparam-se no complexo C. gloeosporioides. A
análise filogenética revelou, com elevado bootstrap, que o complexo C. acutatum diverge do
complexo C. gloeosporioides, porém, com semelhanças filogenéticas com o complexo C.
boninense. A análise da diversidade genética distribuiu os isolados de dois grupos principais
pelos dois métodos de agrupamento e representação gráfica. Pelo método UPGMA
(Unweighted Pair Group Method using Arithmetic Averages) houve a formação de um
subgrupo exclusivo com os isolados referentes à espécie C. acutatum. A análise da
diversidade genética em loci ISSR demonstrou-se eficaz, sugerindo ampla diversidade,
refletindo tanto na variação morfológica, como nos eventos epidemiológicos que acometem a
cultura da seringueira.
Palavras-chave: filogenia molecular, GAPDH, marcadores moleculares, ITS, ISSR.

58
ABSTRACT
The incidence of Colletotrichum species has been widely reported as one of the main
phytopathogens that affect rubber tree (Hevea brasiliensis), causing several losses in the
rubber production chain. Due to the great species diversity in the Colletotrichum spp. x H.
brasiliensis interation, studies of morphological diversity, genetics and the identification of
isolates in the level of species, may contribute to management actions in control and breeding
strategies for resistance to Anthracnose. In this context, the present work aimed at the
morphocultural and molecular characterization of 29 isolates from Colletotrichum genera
from rubber leaves. The isolates were identified at the species level with the amplification and
comparative sequencing of their ITS1/4 genomic sequences and the GAPDH gene. The
analysis of genetic diversity was evaluated by obtaining the molecular markers Inter-Simple
Sequence Repeats (ISSR). The morphological evaluation of the colonies revealed
heterogeneity in all the analyzed descriptors, making difficult the certification at the level of
complex and species. However, the identification of the species in this pathosystem revealed
the presence of five species belonging to three complexes: C. gloeosporioides, C. acutatum
and C. boninense. In the sample group, the largest number of species were of the C.
gloeosporioides complex. The phylogenetic analysis revealed, with high bootstrap, that the C.
acutatum complex diverges from the C. gloeosporioides complex, however, with
phylogenetic similarities with the C. boninense complex. The analysis of genetic diversity
distributed the isolates of two main groups by the two methods of grouping and graphical
representation. By the UPGMA (Unweighted Pair Group Method using Arithmetic Averages)
method, the formation of an exclusive subgroup with C. acutatum isolates was performed.
The analysis of the genetic diversity in ISSR loci proved to be effective, suggesting a wide
diversity, reflecting both the morphological variation and the epidemiological events that
affect the rubber tree culture.
Key word: molecular phylogeny, GAPDH, molecular markers, ITS, ISSR.

59
5.1 INTRODUÇÃO
O gênero Colletotrichum Corda é de interesse econômico e científico, pois acomete
uma grande variedade de culturas de valor econômico a nível mundial (DEAN et al., 2012;
FARR; ROSSMAN 2017). Espécies pertencentes ao gênero estão frequentemente associadas à
seringueira (Hevea brasiliensis Mull. Arg.), causando a Antracnose que acarreta sérios danos
e redução da produtividade, tornando-se uma das mais importantes doenças foliares da
cultura. A Antracnose é constatada praticamente em todos os países onde a seringueira é
cultivada, e no estado da Bahia tem se destacado por atingir plantas de todas as idades. Em
Hevea brasiliensis o fungo ataca folhas, formando pequenas lesões arredondadas de coloração
marrom-avermelhada, ramos, inflorescência, hastes, placas de enxerto em jardins clonais,
viveiros e painéis de sangria (FURTADO; SILVEIRA, 1992; SILVEIRA; CARDOSO, 1987;
SILVEIRA et al., 1992a).
O estudo de identificação das espécies de Colletotrichum de um determinado
hospedeiro, bem como a determinação de sua variabilidade, é de fundamental importância
para o desenvolvimento de estratégias mais eficientes de controle, além de propiciar um
melhor entendimento da epidemiologia da doença e utilização em programas de
melhoramento visando à caracterização da resistência ao patógeno.
A caracterização das espécies do gênero Colletotrichum foi por muito tempo
baseada principalmente em descritores morfológicos associados à taxonomia, e também a
aspectos culturais como a ontogênese, forma dos conídios, presença e forma de setas e
apressórios, pigmentação e textura das colônias, relação com seus hospedeiros,
patogenicidade e taxa de crescimento micelial (CANNON et al., 2012; THAUNG, 2008).
Essas características foram utilizadas principalmente, para diferenciar espécies
morfologicamente próximas, como C. gloeosporioides (Penz.) Penz. & Sacc. e C. acutatum
J.H. Simmonds (SUTTON, 1992; FREEMAN et al., 1998). No entanto, existe uma grande
dificuldade na identificação dessas espécies, principalmente, pela grande diversidade
fenotípica, influência de fatores ambientais na estabilidade dos caracteres morfoculturais e
alta variabilidade das espécies envolvidas (FREEMAN et al., 1998; LOPEZ, 2001). Uma
abordagem polifásica, combinando técnicas moleculares e morfológicas, é recomendada para
identificação mais precisa das espécies do gênero (CAI et al., 2011).
As técnicas de biologia molecular permitem a resolução de problemas de identificação
fúngica, tendo em vista a análise comparativa em sequências de DNA conservadas (MACLEAN
et al., 1993). Contudo, diversas abordagens moleculares têm sido utilizadas para caracterizar

60
espécies de Colletotrichum e complementar estudo de diferenciação de populações do gênero
(BUENO, 2005; LOPEZ, 2001; NGUYEN et al., 2010). A utilização de primers específicos para
espécies de Colletotrichum obtidos a partir da análise de sequência da região ITS são utilizados
para complementar e resultados de morfologia e patogenicidade (FREEMAM et al., 2001). A
comparação de sequências da região ITS 1 do DNA entre C. gloeosporioides e outras espécies
do gênero levaram ao desenvolvimento de primers específicos para a diferenciação entre C.
gloeosporioides e C. acutatum via PCR (MILLS et al., 1992). Regiões do genoma de fungos do
gênero Colletotrichum têm sido analisadas juntamente com o ITS, utilizando múltiplos
marcadores genéticos que aumentam o grau de confiabilidade na classificação das espécies, tais
como as sequências dos genes que codificam actina (ACT), quitina sintetase (CHS1), β-tubulina
(TUB2), calmodulina (CAL) e gliceraldeído-3-fosfato desidrogenase (GADPH) (CANNON et al.
2012; DAMM et al., 2012a; DAMM et al., 2012b; WEIR et al., 2012).
Técnicas moleculares também têm sido utilizadas para estudar a diversidade e
variabilidade genética entre espécies de Colletotrichum (SILVEIRA et al., 2016; ANDERSON et
al., 2013; RAMPERSAD, 2013; SHARMA et al., 2014; MAHMODI et al., 2014). Dentre elas, o
uso do marcador molecular do tipo sequências simples repetidas internas (ISSR) tem-se
mostrado bastante eficiente, pois não necessitam de conhecimento prévio do DNA a ser avaliado
(BARTH et al., 2002) além de ser uma técnica de baixo custo, fácil uso e de grande
reprodutibilidade (MATTHEWS et al., 1999). Os marcadores ISSR são amplificados via PCR e
não necessitam do sequenciamento da região, resultando ainda na obtenção de padrões altamente
polimórficos (NAGAOKA; OGIHARA, 1997).
Estudos revelam que a Antracnose foliar da seringueira é atribuída às espécies
pertencentes aos complexos Colletotrichum gloeosporioides, C. acutatum e C. boninense
(PETCH, 1921; CARPENTER; STEVENSON, 1954; JAYASINGHE et al., 1997, SAHA et al.,
2002, GAZIS; CHAVERRI, 2010, GAZIS et al., 2011, DAMM et al., 2012a). No entanto, não há
estudos sobre a caracterização morfológica e molecular e a ocorrência das espécies de
Colletotrichum associadas à Antracnose foliar na região Sudeste da Bahia, importante pólo
heveícola brasileiro.
O presente trabalho objetivou a caracterização morfológica e molecular de 29 isolados de
Colletotrichum coletados na região Sudeste da Bahia em folhas de seringueira. Em paralelo, os
isolados foram identificados pelo sequenciamento comparativo de duas sequências genômicas, e
em seguida foram analisados quanto à filogenia e diversidade genética em regiões genômicas.

61
5.2 MATERIAL E MÉTODOS
5.2.1 Coleta e isolamento de Colletotrichum spp.
Foram coletadas folhas de clones de seringueira (Tabela 1) com sintomas da
Antracnose, em áreas distintas de seringais nas Plantações Michelin da Bahia (PMB),
localizada no município de Igrapiúna (9º13’ S, 2º39’ W), Estação Experimental Djalma Bahia
(EDJAB), em Una (14º47’ S, 39º03’ W) e Centro de Pesquisa do Cacau (Ceplac/Cepec),
Ilhéus, Bahia (14º47’ S, 39º02’ W). Os materiais coletados foram acondicionados em sacos de
papel e levados para o laboratório de Diversidade de fungos do Cepec/Ceplac em Ilhéus,
Bahia, onde foram lavados superficialmente com água corrente. Foram cortados pequenos
fragmentos compreendidos entre a área necrosada e o tecido assintomático, onde procedeu-se
a desinfestação utilizando álcool a 70% por 30 segundos, solução de hipoclorito de sódio a
2% por um minuto, água destilada estéril por duas vezes e secagem em papel toalha.
Os fragmentos foram colocados em placas de Petri de 8 cm de diâmetro contendo
meio Batata-Dextrose-Ágar (BDA Difco®), suplementados com ácido tartárico a 0,1 %. Todo
o procedimento foi realizado sob condições assépticas, em câmara de fluxo laminar, e as
placas foram mantidas em bancada sob temperatura de 24±1 °C, durante 3 dias. Após esse
período, selecionaram-se as colônias típicas de Colletotrichum spp., seguida de repicagem e
obtenção de culturas puras. No total foram obtidos 29 isolados de Colletotrichum spp., que
foram preservados em óleo mineral e pelo método de Castellani (1967).
5.2.2 Caracterização morfocultural dos isolados de Colletotrichum.
Foram caracterizados morfologicamente 29 isolados de Colletotrichum spp. pela
forma e tamanho de 40 conídios visualizados aleatoriamente ao microscópio óptico no
aumento de 40x. Os isolados foram classificados de acordo com os seguintes formatos:
oblongos, oblongo cilindráceos, cilindráceos, clavados e fusoides.
A caracterização cultural baseou-se na coloração e textura das colônias, sendo
consideradas colônias com crescimento micelial flocoso, feltroso, velutino e liso e coloração
em tons acinzentados, esbranquiçados, alaranjados, rosados e creme amarelado (KIRK et al.,
2008).
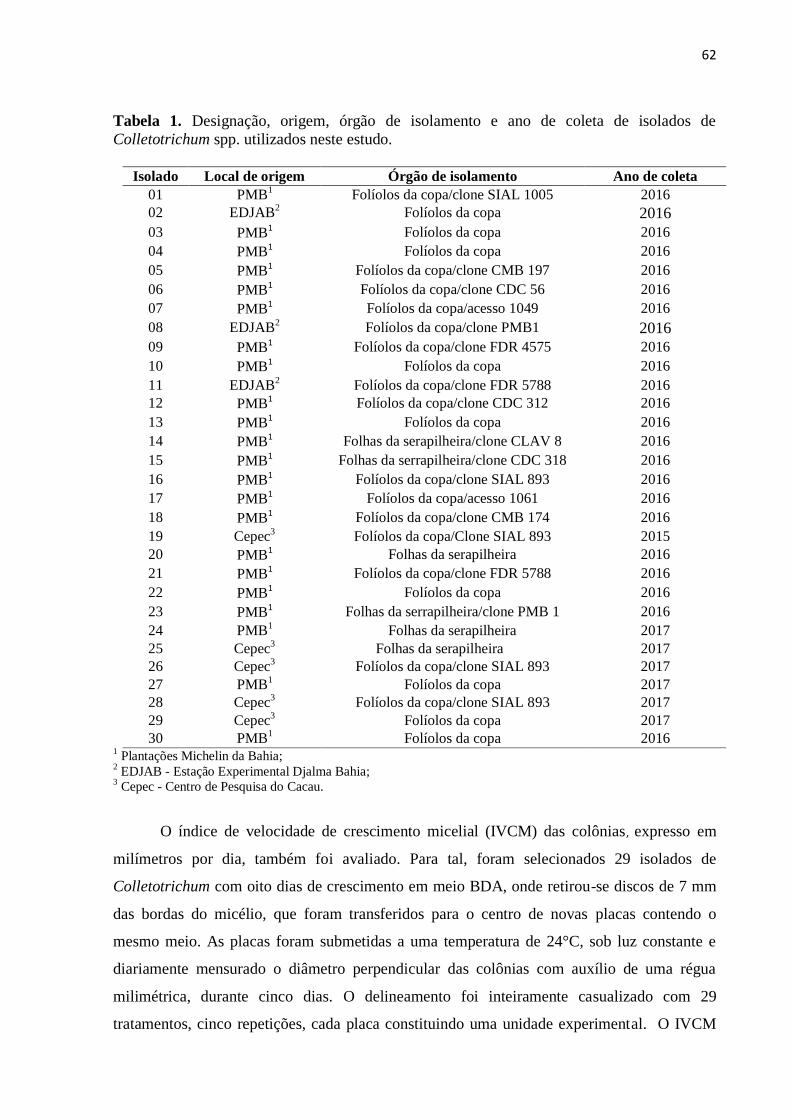
62
Tabela 1. Designação, origem, órgão de isolamento e ano de coleta de isolados de
Colletotrichum spp. utilizados neste estudo.
Isolado Local de origem Órgão de isolamento Ano de coleta
01 PMB1 Folíolos da copa/clone SIAL 1005 2016
02 EDJAB2 Folíolos da copa 2016
03 PMB1 Folíolos da copa 2016
04 PMB1 Folíolos da copa 2016
05 PMB1 Folíolos da copa/clone CMB 197 2016
06 PMB1 Folíolos da copa/clone CDC 56 2016
07 PMB1 Folíolos da copa/acesso 1049 2016
08 EDJAB2 Folíolos da copa/clone PMB1 2016
09 PMB1 Folíolos da copa/clone FDR 4575 2016
10 PMB1 Folíolos da copa 2016
11 EDJAB2 Folíolos da copa/clone FDR 5788 2016
12 PMB1 Folíolos da copa/clone CDC 312 2016
13 PMB1 Folíolos da copa 2016
14 PMB1 Folhas da serapilheira/clone CLAV 8 2016
15 PMB1 Folhas da serrapilheira/clone CDC 318 2016
16 PMB1 Folíolos da copa/clone SIAL 893 2016
17 PMB1 Folíolos da copa/acesso 1061 2016
18 PMB1 Folíolos da copa/clone CMB 174 2016
19 Cepec3
Folíolos da copa/Clone SIAL 893 2015
20 PMB1 Folhas da serapilheira 2016
21 PMB1 Folíolos da copa/clone FDR 5788 2016
22 PMB1 Folíolos da copa 2016
23 PMB1 Folhas da serrapilheira/clone PMB 1 2016
24 PMB1 Folhas da serapilheira 2017
25 Cepec3 Folhas da serapilheira 2017
26 Cepec3 Folíolos da copa/clone SIAL 893 2017
27 PMB1 Folíolos da copa 2017
28 Cepec3 Folíolos da copa/clone SIAL 893 2017
29 Cepec3 Folíolos da copa 2017
30 PMB1 Folíolos da copa 2016
1 Plantações Michelin da Bahia;
2 EDJAB - Estação Experimental Djalma Bahia;
3 Cepec - Centro de Pesquisa do Cacau.
O índice de velocidade de crescimento micelial (IVCM) das colônias, expresso em
milímetros por dia, também foi avaliado. Para tal, foram selecionados 29 isolados de
Colletotrichum com oito dias de crescimento em meio BDA, onde retirou-se discos de 7 mm
das bordas do micélio, que foram transferidos para o centro de novas placas contendo o
mesmo meio. As placas foram submetidas a uma temperatura de 24°C, sob luz constante e
diariamente mensurado o diâmetro perpendicular das colônias com auxílio de uma régua
milimétrica, durante cinco dias. O delineamento foi inteiramente casualizado com 29
tratamentos, cinco repetições, cada placa constituindo uma unidade experimental. O IVCM

63
foi medido segundo Nechet (1999), adaptada por Oliveira (1991): IVCM= S (D-Da)/N, em
que IVCM= Índice de velocidade de crescimento micelial, D = Diâmetro médio atual, Da =
Diâmetro médio do dia anterior, e N = Número de dias após a inoculação. Os dados obtidos
foram submetidos ao teste de F a 5% de probabilidade, e as médias comparadas
pelo teste de Scott-Knott a 5%.
5.2.3 Teste de esporulação das espécies de Colletotrichum.
Para verificação da capacidade de esporulação, os isolados foram crescidos em BDA
por oito dias a 24º C. Após esse período, depositou-se 10 mL de água destilada estéril em
cada placa, onde procedeu-se a raspagem do micélio com o auxílio de uma alça de Drigalski.
O micélio foi filtrado em uma camada dupla de gaze estéril, retirando-se uma alíquota para
quantificação em câmara de Neubauer.
5.2.4 Teste de patogenicidade
O teste de patogenicidade foi realizado com todos 29 isolados de Colletotrichum,
utilizando folíolos do clone Fx 3864, no estádio B2 de desenvolvimento (HALLÉ et al.,
1978), com nove repetições para cada isolado. O inóculo foi preparado a partir de culturas
com sete dias de crescimento em meio BDA. Uma suspensão com 105 conídios/mL foi
inoculada utilizando uma alíquota de 20 µm sobre a superfície abaxial dos folíolos. No
tratamento controle, nenhum isolado foi inoculado.
5.2.5 Identificação molecular por sequenciamento
Para caracterização molecular, foi feita a extração de DNA genômico de 29 isolados
de Colletotrichum e um isolado de Phomopsis sp. (grupo externo), com sete dias de cultivo
em meio BDA à 24°C, sob luz constante, de acordo com o método CTAB 2% (Cetiltrimetil
Brometo de Amônio) para fungos filamentosos modificado por Muller et al. (1992) e Geraldo
et al. (2004). O DNA extraído foi quantificado por eletroforese gel de agarose 1% em tampão
TBE 1X (Tris 89 mM, ácido bórico 89 mM, EDTA 2,5 mMpH 8,3), por 90 volts por 30
minutos. Como padrão de quantificação foi utilizado o DNA Lambda com 100 ng
(Fermentas®). As bandas de DNA foram visualizadas em transiluminador de luz ultravioleta e

64
fotografadas pelo uso do sistema Kodak®
Electrophoresis Documentation and Analysis
System, cuja comparação do brilho das bandas dos DNAs com o Lambda DNA padrão foi
realizada para a inferência da concentração das amostras. Os DNAs extraídos foram diluídos
para a concentração de 10ng/ μ e utilizados nas amplificações em cadeia da polimerase (PCR).
O DNA genômico anteriormente extraído foi utilizado para a identificação dos isolado
pelo sequenciamento das sequências ITS1/ITS4 e da região gênica parcial do gene
gliceraldeído-3-fosfato desidrogenase (GAPDH). O isolamento das sequências foram
realizados por reação em cadeia da polimerase com o uso dos primers ITS1
(TCCGTAGGTGAACCTGCGG) e ITS4 (TCCTCCGCTTATTGATATGC) para a sequência
ITS1/ITS4 que amplificam um fragmento com cerca de 600 pb (WHITE et al., 1990;
SREENIVASAPRASAD et al., 1996). A outra região genômica isolada foi o gene parcial
GAPDH gerando fragmentos com cerca de 250 pb com o uso dos primers
(GCCGTCAACGACCCCTTCATTGA) e (GGTGGAGTCGTACTTGAGCATGT)
(GUERBER et al., 2003).
As amplificações foram realizadas utilizando um volume final de 50 µL, com mistura
de reação contendo 33,55 µL de água ultrapura, 5,0 µL de tampão de reação 10x, 3,0 µL de
MgCl2 (25mM), 3,2 µL de dNTPs (2,5 mM cada), 1 µL do primer R (10 p.mol) e 1 µL do
primer F (10 p.mol), 0,25 µL de DNA Taq-polimerase (5,0 U μl-1) e 3,0 µL de DNA diluído
em água (1:100). Utilizou-se o termociclador PTC100 (MJ Research), com a seguinte
programação: desnaturação inicial 5 min a 94°C, seguido por 35 ciclos de 45s a 94°C, 1 min a
56°C e 1 min a 72°C. A extensão final foi a 72°C por 7 min.
Os produtos da amplificação foram observados com a eletroforese em gel de agarose
1,2% contendo o corante SYBR (Invitrogen®) feito com em tampão TBE 1X (Tris 89 mM,
ácido bórico 89 mM, EDTA 2,5 mM pH 8,3). Foi utilizado o padrão de tamanho molecular
para 100 pb ―DNA Ladder Plus (Sinapse Biotecnologia®
)‖. A eletroforese foi realizada com a
constante de 90 volts por 45 minutos. As bandas de DNA foram visualizadas em
transiluminador de ultravioleta e fotografadas pelo uso do sistema Kodak®
Electrophoresis
Documentation and Analysis System.
Os fragmentos amplificados foram purificados com mistura enzimática ExoI
(Fermentas®) e SAP (Fermentas
®) utilizando 0,8 μL de cada enzima, 1,65 μL de H2O ultra
pura e 20 μL das reações de PCR com incubação a 37°C durante 30 min, seguido de
inativação enzimática a 80°C durante 15 min. Após a purificação o produto foi precipitado
com a adição de 10% do volume da reação de Acetato de Sódio 10%, mais 200% do volume
final de etanol P.A. gelado.

65
O sequenciamento foi realizado em sequenciador ABI3500 XL (AppliedBiosystems®).
utilizando-se o kit BigDye® terminator v 3.1 cycle sequencing conforme as recomendações
do fabricante. As sequências foram editadas e alinhadas usando o programa BioEdit Sequence
Alignment Editor (1997-2005). As sequências ITS e o gene GAPDH foram concatenadas com
o uso do programa Mega 5.05. A árvore filogenética foi construída na plataforma Phylogeny
pelo método ―one click‖ usando o modelo de comparação Neighbor-Joining e ―p-distance‖
(www.phylogeny.fr). A árvore filogenética gerada foi editada com o uso do software
Photoshop CS5 (Adobe®).
5.2.6 Diversidade genética em regiões ISSR
A escolha dos primers para a obtenção dos marcadores ISSR foi baseada em estudos
anteriores realizados no gênero Colletotrichum em relação à capacidade de revelar o
polimorfismo nessas regiões genômicas (GUPTA et al., 1994; RATANACHERDCHAI et al.,
2010; RAMPERSAD, 2013). As reações de PCR foram realizadas para um volume final de
20 µL, contendo 1X de tampão de reação para DNA Taq-polimerase 10X; 75 µM de MgCl2,
5 mM de dNTPs; 10 p.mol do primer; 1 U de DNA Taq-polimerase e 30 ng de DNA. As
condições de amplificação foram: desnaturação inicial 5 min a 94°C, seguido por 35 ciclos de
45s a 94°C, 1 min a temperatura específica para o anelamento do primer, e 1 min a 72°C. A
extensão final foi a 72°C por 7 min; em termociclador PTC100 (MJ®Research).
Os produtos das amplificações foram observados com a eletroforese em tampão TBE
1X (Tris 89 mM, ácido bórico 89 mM, EDTA 2,5 mMpH 8,3), utilizando gel de agarose 1,8%
contendo o corante SYBR (Invitrogen®
) com a constante de 90 volts por 50 minutos. Foi
utilizado o padrão de tamanho molecular para 100 pb ―DNA Ladder Plus (Sinapse
Biotecnologia®)‖. As bandas de DNA foram visualizadas em transiluminador de ultravioleta
e fotografadas pelo uso do sistema Kodak®
Electrophoresis Documentation and Analysis
System.
A partir dos produtos de amplificação (bandas claras e evidentes) foi construída uma
matriz binária de dados com informações de presença (1) e ausência de bandas (0). Para a
análise de similaridade entre os isolados, foi empregado o coeficiente de Simple Matching,
através do programa NTSYS-pc 2.1 software (ROHLF, 2000). O dendrograma foi construído
baseado no agrupamento estatístico UPGMA (Unweighted Pair Group Method with
Arithmetic mean) e editado no software Photoshop CS5 (Adobe®). A análise de correlação

66
cofrenética foi também realizada, juntamente com a distribuição dos isolados em um gráfico
de coordenadas principais baseando-se na diversidade avaliada pelo coeficiente de Simple
Matching. Adicionalmente, foi realizada a análise de componentes principais PCA com o uso
do software NTSYS-pc 2.1 (ROHLF, 2000)
5.3 RESULTADOS
5.3.1 Caracterização Morfocultural
Todos os 29 isolados avaliados, apresentaram padrões morfoculturais que se
enquadravam dentre aqueles descritos para o gênero Colletotrichum (SUTTON, 1992). Em
meio BDA, os isolados apresentaram hifas septadas hialinas, células conidiógenas alongadas a
ampuliformes, conídios hialinos, unicelulares, de formas variadas. Não foram observadas
setas e clamidósporos. O teleomorfo não foi observado.
As colônias de Colletotrichum spp. apresentaram-se bastante heterogêneas quanto aos
aspectos morfoculturais, sendo observados cinco tipos de coloração, quatro texturas miceliais
(Figura 1) e cinco formas predominantes de conídios (Figura 2). Culturas de coloração
acinzentadas foram visualizadas em nove isolados, esbranquiçada em oito isolados, alaranjada
em seis isolados, rosácea em apenas um isolado, e creme amarelada em cinco isolados, dentre
o total avaliado. Seis isolados apresentaram micélio com textura predominante flocosa, 12
isolados com textura feltrosa, sete isolados com textura velutina e um isolado com textura
lisa. Ao microscópio foram observados isolados com conídios predominantemente oblongos
(15), oblongo cilindráceos (5), cilindráceo (7), clavados (1) e fusoides (1) (Tabela 2).
As dimensões médias do comprimento dos conídios variaram de 11,0 µm (isolado 29)
a 16,9 µm (isolado 01); e largura de 1,9 µm (isolado 14) a 5,7 µm (isolado 02). A relação
comprimento/largura (C/L) dos conídios também foi utilizada na caracterização das espécies
do gênero, e variou de 2,5 µm (isolado 06) a 6,8 µm (isolado 14). Os resultados da capacidade
de esporulação das culturas de Colletotrichum em meio BDA variaram de 2,5 x 105
(isolado
15) a 166 x 105 conídios/mL (isolado 19). Todos os isolados testados mostraram-se
patogênicos a folíolos de seringueira. O índice da velocidade de crescimento micelial (IVCM)
apresentou variação de 0,38 (isolado 14) a 1,15 mm/dia (isolado 02) (Tabela 2).

67
Figura 1. Colônias de Colletotrichum spp. apresentando alta variabilidade morfocultural em
meio BDA.
Fonte: Dados da pesquisa.
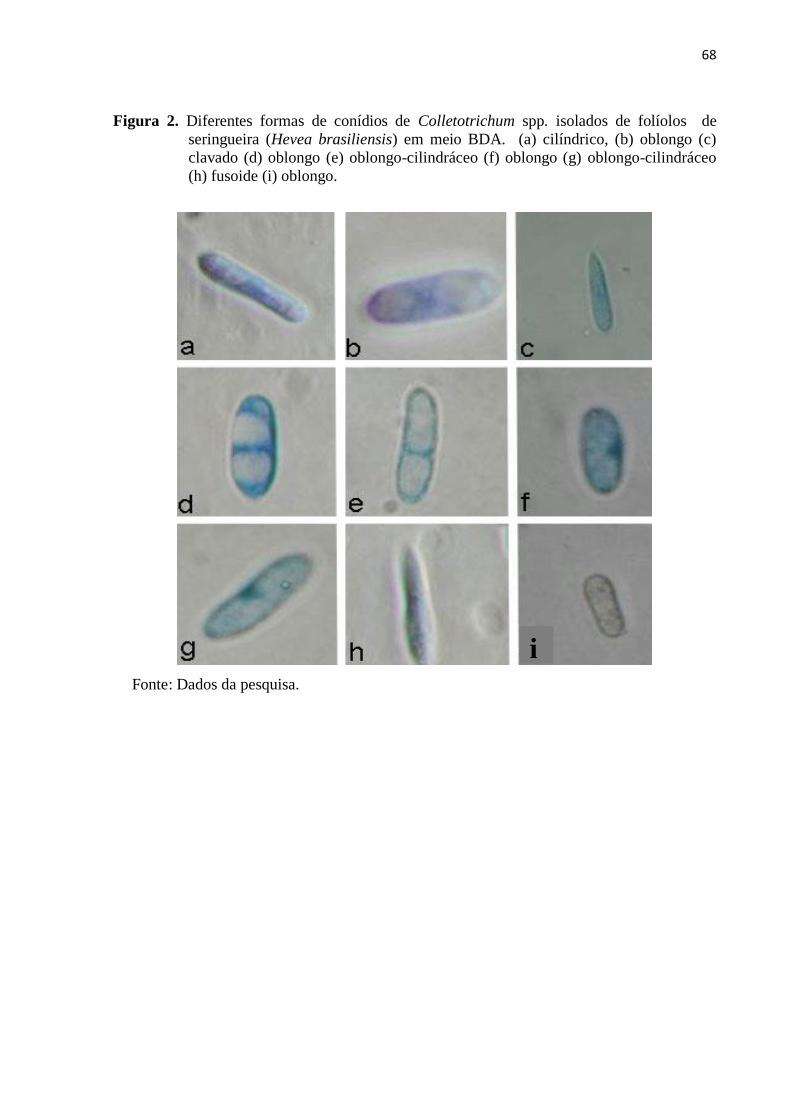
68
Figura 2. Diferentes formas de conídios de Colletotrichum spp. isolados de folíolos de
seringueira (Hevea brasiliensis) em meio BDA. (a) cilíndrico, (b) oblongo (c)
clavado (d) oblongo (e) oblongo-cilindráceo (f) oblongo (g) oblongo-cilindráceo
(h) fusoide (i) oblongo.
Fonte: Dados da pesquisa.
i
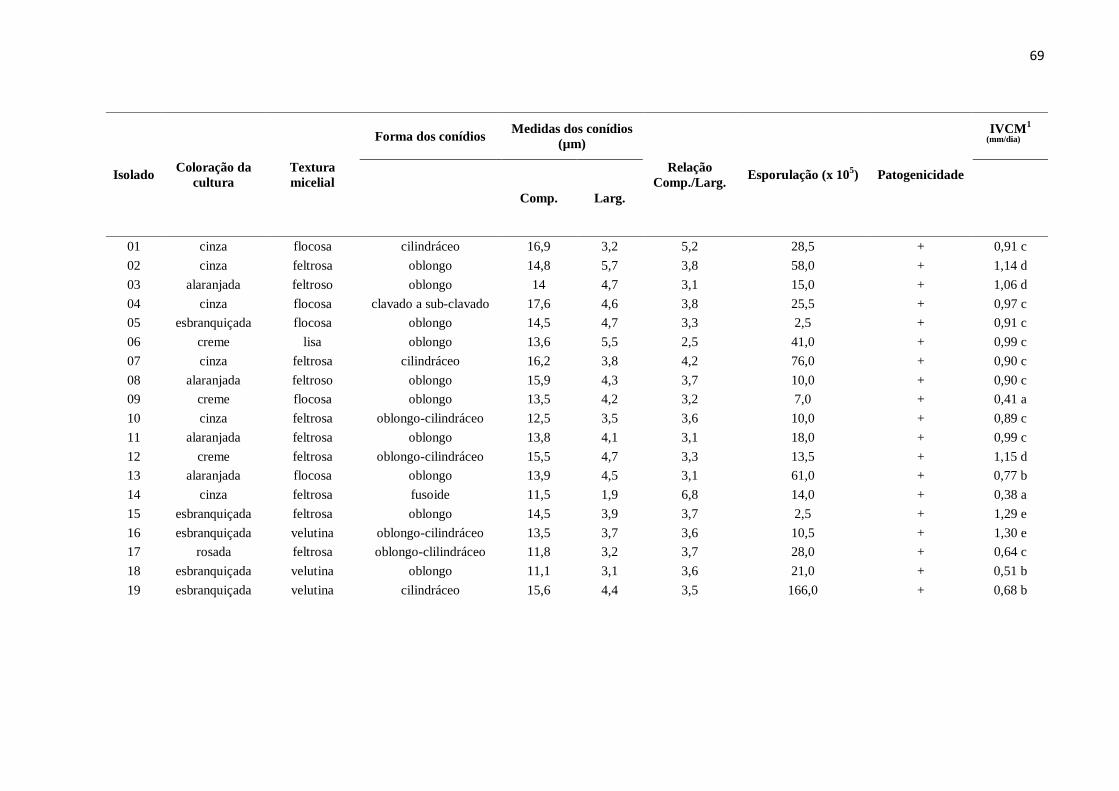
69
Isolado Coloração da
cultura
Textura
micelial
Forma dos conídios Medidas dos conídios
(µm)
Relação
Comp./Larg. Esporulação (x 10
5) Patogenicidade
IVCM1
(mm/dia)
Comp. Larg.
01 cinza flocosa cilindráceo 16,9 3,2 5,2 28,5 + 0,91 c
02 cinza feltrosa oblongo 14,8 5,7 3,8 58,0 + 1,14 d
03 alaranjada feltroso oblongo 14 4,7 3,1 15,0 + 1,06 d
04 cinza flocosa clavado a sub-clavado 17,6 4,6 3,8 25,5 + 0,97 c
05 esbranquiçada flocosa oblongo 14,5 4,7 3,3 2,5 + 0,91 c
06 creme lisa oblongo 13,6 5,5 2,5 41,0 + 0,99 c
07 cinza feltrosa cilindráceo 16,2 3,8 4,2 76,0 + 0,90 c
08 alaranjada feltroso oblongo 15,9 4,3 3,7 10,0 + 0,90 c
09 creme flocosa oblongo 13,5 4,2 3,2 7,0 + 0,41 a
10 cinza feltrosa oblongo-cilindráceo 12,5 3,5 3,6 10,0 + 0,89 c
11 alaranjada feltrosa oblongo 13,8 4,1 3,1 18,0 + 0,99 c
12 creme feltrosa oblongo-cilindráceo 15,5 4,7 3,3 13,5 + 1,15 d
13 alaranjada flocosa oblongo 13,9 4,5 3,1 61,0 + 0,77 b
14 cinza feltrosa fusoide 11,5 1,9 6,8 14,0 + 0,38 a
15 esbranquiçada feltrosa oblongo 14,5 3,9 3,7 2,5 + 1,29 e
16 esbranquiçada velutina oblongo-cilindráceo 13,5 3,7 3,6 10,5 + 1,30 e
17 rosada feltrosa oblongo-clilindráceo 11,8 3,2 3,7 28,0 + 0,64 c
18 esbranquiçada velutina oblongo 11,1 3,1 3,6 21,0 + 0,51 b
19 esbranquiçada velutina cilindráceo 15,6 4,4 3,5 166,0 + 0,68 b
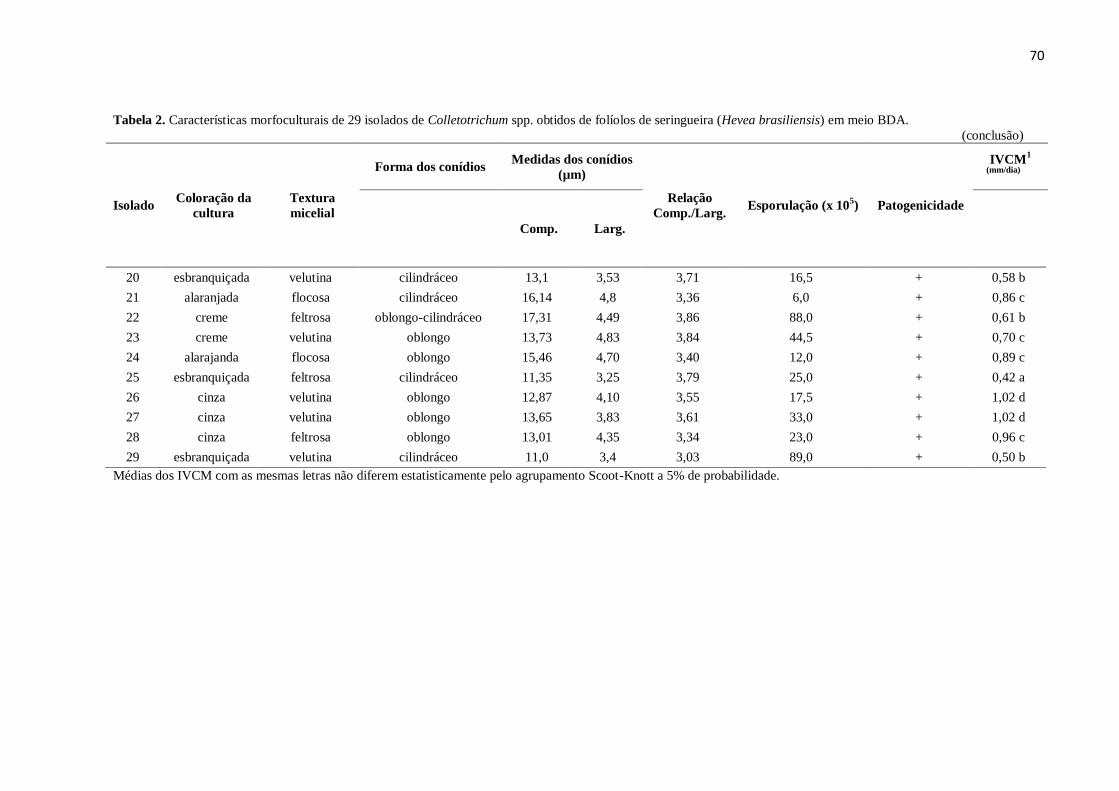
70
Tabela 2. Características morfoculturais de 29 isolados de Colletotrichum spp. obtidos de folíolos de seringueira (Hevea brasiliensis) em meio BDA.
(conclusão)
Isolado Coloração da
cultura
Textura
micelial
Forma dos conídios Medidas dos conídios
(µm)
Relação
Comp./Larg. Esporulação (x 10
5) Patogenicidade
IVCM1
(mm/dia)
Comp. Larg.
20 esbranquiçada velutina cilindráceo 13,1 3,53 3,71 16,5 + 0,58 b
21 alaranjada flocosa cilindráceo 16,14 4,8 3,36 6,0 + 0,86 c
22 creme feltrosa oblongo-cilindráceo 17,31 4,49 3,86 88,0 + 0,61 b
23 creme velutina oblongo 13,73 4,83 3,84 44,5 + 0,70 c
24 alarajanda flocosa oblongo 15,46 4,70 3,40 12,0 + 0,89 c
25 esbranquiçada feltrosa cilindráceo 11,35 3,25 3,79 25,0 + 0,42 a
26 cinza velutina oblongo 12,87 4,10 3,55 17,5 + 1,02 d
27 cinza velutina oblongo 13,65 3,83 3,61 33,0 + 1,02 d
28 cinza feltrosa oblongo 13,01 4,35 3,34 23,0 + 0,96 c
29 esbranquiçada velutina cilindráceo 11,0 3,4 3,03 89,0 + 0,50 b
Médias dos IVCM com as mesmas letras não diferem estatisticamente pelo agrupamento Scoot-Knott a 5% de probabilidade.

71
5.3.2 Sequenciamento e identificação dos níveis taxonômicos dos isolados
Utilizando as ferramentas morfológicas e moleculares foi possível identificar mais de
uma espécie de Colletotrichum associadas à Antracnose foliar da seringueira. O
sequenciamento das sequências ITS1/4 e do gene GAPDH, revelaram fragmentos em torno de
600 e 250 pb) respectivamente, para todos os 30 isolados analisados. As sequências obtidas
foram comparadas com as informações existentes no banco de dados de ácidos nucléico NCBI
(http:// blast.ncbi.nlm.nih.gov/Blast.cgi) (HSIANG; GOODWIN, 2001), onde foi possível
identificar os isolados pelas semelhanças entre as respostas das sequências pelo método de
BLAST. Desta forma, foram identificados três complexos de espécies: C. gloeosporioides, C.
acutatum e C. boninense. O grupo externo formado por uma amostra do gênero Phomopsis
previamente conhecida foi identificada com elevada acurária na comparação com o banco de
dados (Figura 3).
Foi observado no complexo C. gloeosporioides, que 60% dois isolados são da espécie
C. gloeosporioides, 28% são C. siamense e apenas 9% C. theobromicola. As espécies desse
complexo foram identificadas com o query cover variando de 87 a 97%. O complexo C.
acutatum foi representado unicamente pela espécie C. acutatum, totalizando sete isolados.
Este complexo foi identificado com elevado query cover (99%). Uma única espécie do
complexo C. boninense foi encontrada, sendo o isolado ―06‖ com query cover de 89%. O
isolado ―30‖ composto pelo grupo externo foi confirmado como do gênero Phomopsis com
query cover de 85%.
A árvore com a distribuição filogenética comparativa entre os isolados (Figura 3)
demonstrou a distribuição das espécies C. acutatum (bootstrap 0.96 e 0.92) e C. boniense
distantes das demais espécies do complexo C. gloeosporioides. Contudo, apesar de C.
acutatum estar próxima de C. boniense, ambas estão em ramos filogenéticos distintos com
bootstrap 0.98. As espécies do complexo C. gloeosporioides foram distribuídas em dois
grupos principais com bootstrap de 0.80 e 0.83. O isolado 04 representando a espécies C.
theobromicola foi distribuída com grande distancia filogenética do isolado 10, sendo a mesma
espécie, contudo essa distribuição gerada com baixo bootstrap de 0.50. O grupo externo,
representado pelo acesso do gênero Phomopsis foi distribuído com elevado bootstrap de 0.93.
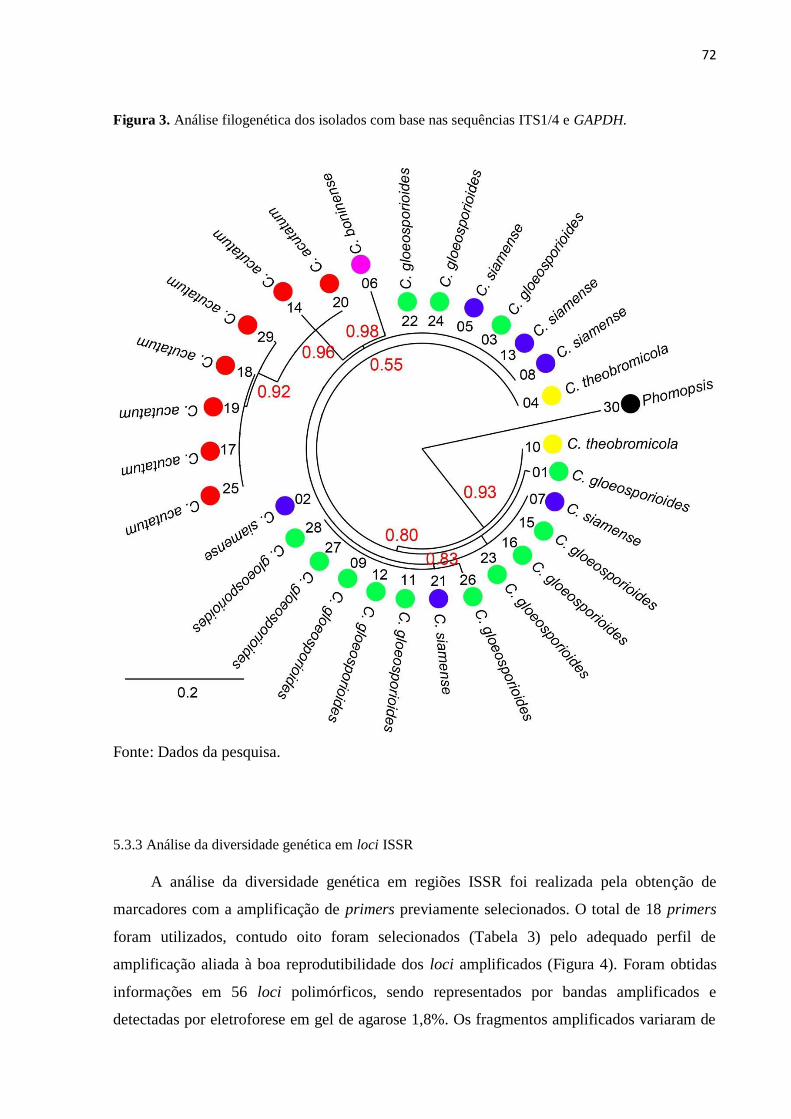
72
Figura 3. Análise filogenética dos isolados com base nas sequências ITS1/4 e GAPDH.
Fonte: Dados da pesquisa.
5.3.3 Análise da diversidade genética em loci ISSR
A análise da diversidade genética em regiões ISSR foi realizada pela obtenção de
marcadores com a amplificação de primers previamente selecionados. O total de 18 primers
foram utilizados, contudo oito foram selecionados (Tabela 3) pelo adequado perfil de
amplificação aliada à boa reprodutibilidade dos loci amplificados (Figura 4). Foram obtidas
informações em 56 loci polimórficos, sendo representados por bandas amplificados e
detectadas por eletroforese em gel de agarose 1,8%. Os fragmentos amplificados variaram de
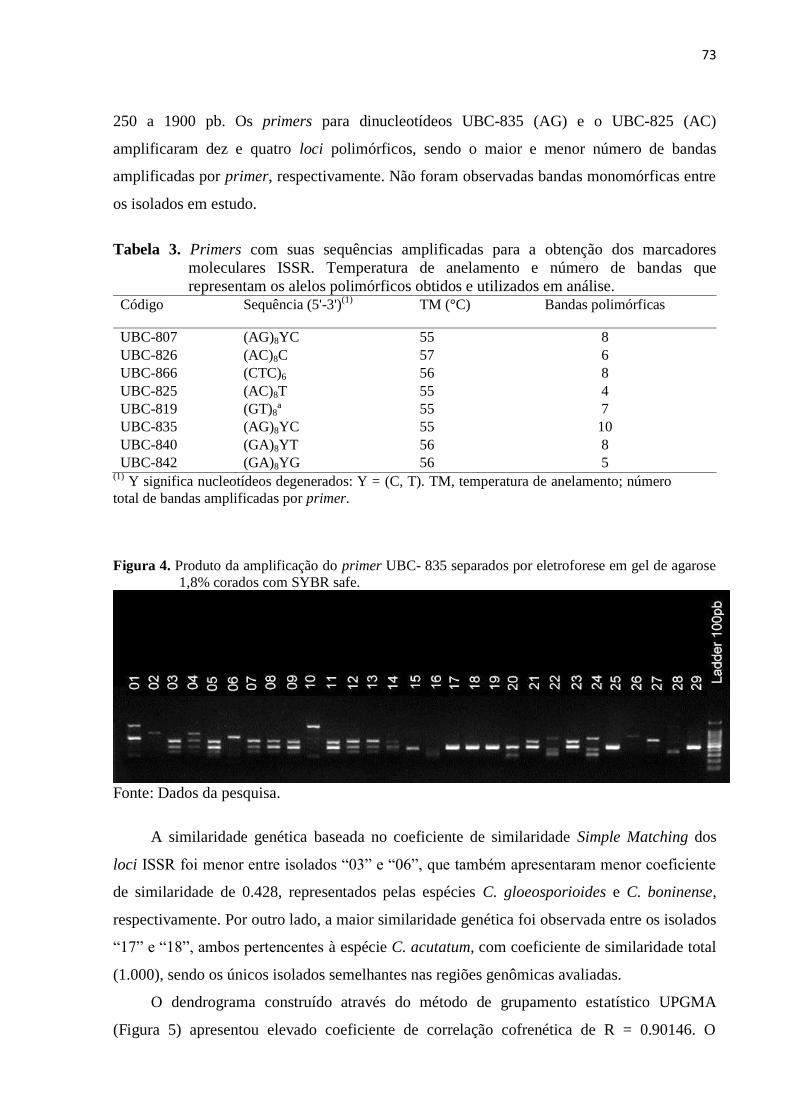
73
250 a 1900 pb. Os primers para dinucleotídeos UBC-835 (AG) e o UBC-825 (AC)
amplificaram dez e quatro loci polimórficos, sendo o maior e menor número de bandas
amplificadas por primer, respectivamente. Não foram observadas bandas monomórficas entre
os isolados em estudo.
Tabela 3. Primers com suas sequências amplificadas para a obtenção dos marcadores
moleculares ISSR. Temperatura de anelamento e número de bandas que
representam os alelos polimórficos obtidos e utilizados em análise. Código Sequência (5'-3')
(1) TM (°C) Bandas polimórficas
UBC-807 (AG)8YC 55 8
UBC-826 (AC)8C 57 6
UBC-866 (CTC)6 56 8
UBC-825 (AC)8T 55 4
UBC-819 (GT)8ª 55 7
UBC-835 (AG)8YC 55 10
UBC-840 (GA)8YT 56 8
UBC-842 (GA)8YG 56 5 (1)
Y significa nucleotídeos degenerados: Y = (C, T). TM, temperatura de anelamento; número
total de bandas amplificadas por primer.
Figura 4. Produto da amplificação do primer UBC- 835 separados por eletroforese em gel de agarose
1,8% corados com SYBR safe.
Fonte: Dados da pesquisa.
A similaridade genética baseada no coeficiente de similaridade Simple Matching dos
loci ISSR foi menor entre isolados ―03‖ e ―06‖, que também apresentaram menor coeficiente
de similaridade de 0.428, representados pelas espécies C. gloeosporioides e C. boninense,
respectivamente. Por outro lado, a maior similaridade genética foi observada entre os isolados
―17‖ e ―18‖, ambos pertencentes à espécie C. acutatum, com coeficiente de similaridade total
(1.000), sendo os únicos isolados semelhantes nas regiões genômicas avaliadas.
O dendrograma construído através do método de grupamento estatístico UPGMA
(Figura 5) apresentou elevado coeficiente de correlação cofrenética de R = 0.90146. O

74
dendrograma separou os isolados de Colletotrichum em dois grandes grupos principais. O
grupo I incluiu oito isolados, subdivididos em dois subgrupos IA e IB. O grupo II também foi
subdividido em dois subgrupos: IIA e IIB. No subgrupo IA dois isolados para a espécie C.
gloeosporioides foram distribuídos juntamente com C. theobromicola (isolado 10). Já no
subgrupo IB o único isolado pertencente a C. boninense foi disposto próximo a quatro da
espécie C. gloeosporioides. No subgrupo IIA o isolado ―02‖ (C. siamense) se apresentou mais
distantes dos demais isolados do mesmo grupo, contudo os isolados que representam C.
gloeosporioides e C. siamense foram dispostos próximos entre si. No subgrupo IIB os
isolados de C. acutatum foram agrupados em um ramo exclusivo, contudo próximo a um C.
theobromicola (―04‖). Os isolados ―017‖ e ―018‖, que representam C. acutatum, foram
distribuídos no mesmo ramo com maior similaridade genética.
Figura 5. Dendrograma baseado no polimorfismo associados aos loci ISSR de isolados de
Colletotrichum spp. Coeficiente de similaridade de Simple Matching e agrupamento pelo
método UPGMA.
Fonte: Dados da pesquisa.
O coeficiente de similaridade genética de Simple Matching gerado entre os isolados
analisados foi utilizado para a distribuição dos genótipos em um gráfico de análise de
componentes principais (Figura 6). Nesta análise foram observados dois grupos principais
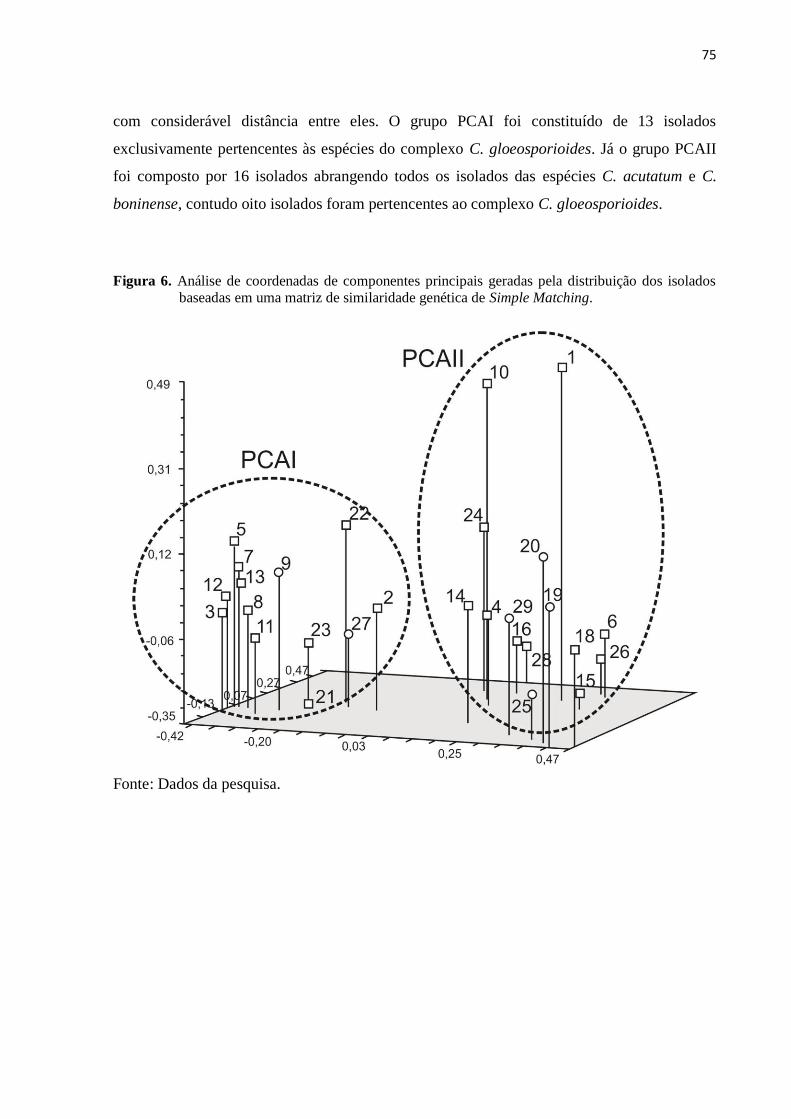
75
com considerável distância entre eles. O grupo PCAI foi constituído de 13 isolados
exclusivamente pertencentes às espécies do complexo C. gloeosporioides. Já o grupo PCAII
foi composto por 16 isolados abrangendo todos os isolados das espécies C. acutatum e C.
boninense, contudo oito isolados foram pertencentes ao complexo C. gloeosporioides.
Figura 6. Análise de coordenadas de componentes principais geradas pela distribuição dos isolados
baseadas em uma matriz de similaridade genética de Simple Matching.
Fonte: Dados da pesquisa.

76
5.4 DISCUSSÃO
A caracterização e análise dos descritores morfológicos revelaram a alta variabilidade
entre os isolados obtidos. Apesar da predominância das colorações acinzentadas e
esbranquiçadas, a variação observada nos isolados avaliados coaduna com resultados obtidos
que indicam alta variabilidade nas colorações culturais de Colletotrichum spp. (ANDRADE et
al., 2007; PIMENTA, 2009).
Sutton (1992) descreve colônias de coloração alaranjadas (salmão) como típicas da
espécie Colletotrichum acutatum, e alta variabilidade colorimétrica dentro com complexo C.
gloeosporioides. As variações apresentadas pelas colônias dessa espécie podem ser atribuídas
ao genótipo de cada isolado, cuja expressão fenotípica está associada ao ambiente de cultivo,
assim como à biossíntese de metabólitos secundários, fatores esses que contribuem para as
grandes variações culturais (BAILEY; JERGER, 1992; BONETT et al., 2010). Segundo essa
característica, as culturas alaranjadas (isolados 03, 08, 11, 13, 21 e 24) seriam identificadas
como C. acutatum, muito embora, segundo a análise molecular, esses isolados tenham sido
classificados como C. gloeosporioides, comprovando que os métodos baseados em análises
culturais têm se mostrado insuficientes para a classificação da espécie de Colletotrichum,
conforme ressaltado por Freeman et al. (1998) e Stracier (2015).
A heterogeneidade morfocultural e a presença de características comuns a mais de
uma espécie ou complexo de espécies de Colletotrichum descritas neste trabalho, corroboram
com os resultados encontrados nos diversos trabalhos referentes à caracterização
morfocultural do gênero Colletotrichum (ANDRADE, 2007; PERES, 2002; ALAM et al.,
2017). A ausência de setas reportadas neste estudo pode estar associada à composição do
meio de cultura além de fatores ambientais, tais como aeração, umidade do ar, temperatura e
fotoperíodo (CARVALHO, 1997).
O isolado 14 foi o único que apresentou a forma dos conídios fusiformes, sendo esta,
uma característica predominante da espécie C. acutatum (SUTTON, 1992), sendo confirmada
a identificação desse isolado segundo a caracterização molecular. No entanto, os demais
isolados identificados via análise comparativa de sequências, como sendo pertencentes ao
complexo C. acutatum (17, 18, 19, 20, 25 e 29) apresentaram formatos variados, com
predominância de conídios cilindráceos, formato descrito como sendo predominante nas
espécies do complexo C. gloeosporioides (SUTTON, 1992; CANNON et al., 2008).

77
Os isolados apresentaram dimensões dentro da faixa de variação descritas por Sutton
(1992), para as espécies de C. acutatum e C. gloeosporiodes. No entanto, a diferenciação com
bases nas dimensões dos conídios, não foi possível devido às proximidades entre os valores, o
que também foram observadas por Afanador-Kafuri et al. (2003), Andrade (2007) e Stracieri
(2015).
O isolado 06 que apresentou a menor relação comprimento/largura (2,5 mm/dia) foi
identificado como C. boninense. Moriwaki et al. (2003) também relataram baixos valores
médios na relação comprimento/largura quando estudaram a espécie C. boninense em
anonáceas no estado de Alagoas. Uma característica útil na distinção desta espécie, é que
alguns conídios apresentam uma pequena cicatriz ou protuberância na base, o que não foi
observado no presente trabalho.
Em BDA, os isolados que mais cresceram foram 02, 03, 12, 15, 16, 26 e 27, com
índice de crescimento micelial acima de 1 mm/dia, todos esses confirmados pela análise
molecular com sendo pertencentes ao complexo C. gloeosporioides. Foram formados cinco
grupos distintos com relação a variável IVCM, os isolados que apresentaram as menores
médias foram identificados como do complexo C. acutatum. Colletotrichum gloeosporioides
é descrito como tendo o crescimento micelial mais rápido, quando comparado à C. acutatum
(GUNNEL; GUBLER, 1992; SMITH; BLACK, 1990; ANDRADE, 2007), o que foi
confirmado em nosso estudo.
Todos os isolados testados mostraram-se patogênicos a folíolos da seringueira, e
apresentaram padrões diferenciados com relação à esporulação. Diferenças tanto na
patogenicidade quanto na virulência têm sido relatadas em espécies do gênero Colletotrichum
nos complexos C. gloeosporioides e C. acutatum (SAXENA et al., 2014).
As espécies de ocorrência mais comum deste gênero formam complexos de espécies
filogenéticas que pela análise morfológica são indistinguíveis ou de difícil distinção umas das
outras (DU et al., 2005; CROUCH et al., 2009). A alta variabilidade cultural e a presença de
características comuns a mais de uma espécie ou complexo de espécies de Colletotrichum
descritas na literatura (MENEZES, 2006; SUTTON, 1992), como coloração, forma e tamanho
de conídios, impossibilitou a clara identificação específica dos isolados com base
exclusivamente nas características avaliadas.
A identificação molecular dos isolados de Colletotrichum foi realizada pela
comparação de sequências depositadas no banco de dados NCBI. Uma porção parcial da
sequencia ITS (Internal Transcribed Spacer) e do gene gliceraldeído-3-fosfato desidrogenase
(GAPDH) foram utilizadas para a comparação e análise. A sequência ITS1/4 é uma região

78
interna entre transcritos do DNA nuclear constituindo parte do DNA ribossomal. Essa região
possui grande conservação em sequência, sendo amplamente utilizada na identificação de
táxons e análise filogenética nos complexos de Colletotrichum (SANGPUEAK et al., 2017).
O gene GAPDH é responsável pela produção de uma enzima que catalisa a oxidação de
glicerol-3-fosfato em diidroxiacetona fosfato (GOUVEIA et al., 2011), a região parcial deste
gene também tem sido amplificada e utilizada na identificação de espécies do complexo
Colletotrichum (FREEMAN et al., 2001; TALHINHAS et al., 2002; NELSON et al., 2013).
As amplificações dos fragmentos utilizados na identificação dos isolados neste estudo
apresentaram tamanhos esperados em pares de base com relação a trabalhos anteriores com o
isolamento destas sequências em espécies do gênero (FREEMAN et al., 2001; TALHINHAS
et al., 2002; NELSON et al., 2013; SANGPUEAK et al., 2017).
A identificação de isolados em nível de espécies utilizando sequências gênicas e
regiões conservadas não expressas pode ser realizada pela simples comparação do produto de
sequenciamento com a homologia nas sequências depositadas em bancos de dados de
informação genética, como por exemplo, o NCBI GenBank data library (http://
blast.ncbi.nlm.nih.gov/Blast.cgi) (HSIANG; GOODWIN, 2001). Este método de identificação
pela comparação com informações prévias das sequências é altamente recomendado para a
visão preliminar da distribuição de espécies em um grupo amostral, contudo quando a
comparação com os bancos de dados atingem similaridade em sequências altas (querry cover
>98) a identificação de espécies possui uma maior confiabilidade (SANGPUEAK et al.,
2017). A identificação de espécies também pode ser realizada com a amplificação diagnóstica
de espécies com o uso de primers específicos (XIE et al., 2010; SARMIENTO, 2013). Esse
método utiliza primers que apenas amplificam uma espécie em particular, se o fragmento for
amplificado significa que o isolado pertence à espécie em questão. O uso de primers
específicos para a identificação diagnóstica não foi utilizado neste estudo, pois é recomendado
no caso de suposições que respondam quais são as possíveis espécies que ocorrem no
patossistema (SARMIENTO, 2013; ANDRADE et al., 2007).
No presente trabalho a análise de sequências indicou a ocorrência de espécies
pertencentes aos três complexos: C. gloeosporioides, C. acutatum e C. boninense. Estudos
relacionados ao patossistema seringueira x Colletotrichum têm demonstrado a interação entre
os três complexos com o citado hospedeiro (SARMIENTO, 2013; SIERRA HAYER, 2014).
Maior predominância de espécies do complexo C. gloeosporioides foi relatada, sendo a mais
predominante, o que difere do observado por Sarmiento (2013) que relatou maior ocorrência
de espécies do complexo C. acutatum ocorrendo com maior frequência em seringueira.

79
Estudando espécies exclusivas do complexo C. acutatum observou-se que esse complexo
possui grande distribuição neste patossistema (HUNUPOLAGAMA, et al., 2017). Apenas
uma espécie foi encontrada como sendo do complexo C. boninense, sendo representada por
um único isolado. Em seringueira, este complexo foi primeiramente identificado em H.
guainensis (GAZIS et al., 2011). Em estudos conduzidos com esse patossistema, C. boninense
têm sido observado com menor frequência quando comparados com os complexos C.
gloeosporiodes e C. acutatum (SARMIENTO, 2013; CAI et al., 2016).
Colletotrichum siamense e C. theobromicola, espécies pertencentes ao complexo C.
gloeosporioides, também foram identificadas, porém, com baixa frequência. Neste contexto,
pode-se indicar que o aumento do grupo amostral demonstra o real perfil populacional tanto
da ocorrência de espécies dos complexos como a diversidade associada a essa ocorrência,
para diferentes patossistemas (SIERRA HAYER, 2014).
Neste estudo, a distribuição filogenética demonstrou que o complexo C. acutatum é
filogeneticamente próximo do complexo C. boninense. Similaridades entre esses complexos
foram indicadas, do ponto de vista morfológico, quando estudou-se um isolado de
Colletotrichum coletado de amêndoas na Austrália (MCKAY et al., 2009). Os autores
identificaram por morfologia, uma espécie de C. acutatum, contudo investigações moleculares
afirmaram que se tratava da espécie C. boninense. Essa incongruência sugere que as
semelhanças morfológicas entre esses complexos refletem também nas proximidades
genômicas. Em contra partida, as espécies do complexo C. gloeosporioides apresentaram
maior distância filogenética entre os demais complexos analisados. A identificação de
espécies desse complexo tem sido dificultada em função das similaridades morfológicas e
variações atribuídas a fatores ambientais (DAMM et al., 2012). Contudo, a identificação
molecular e análise filogenética têm revelado que o perfil de distribuição das espécies de C.
gloeosporioides são os mais diversificado em termos de distância genética, mas também
formam um clado fortemente suportado na análise da diversidade filogenética (WEIR et al.,
2012).
A análise filogenética revelou a distribuição dos dois isolados referentes a espécies C.
theobromicola em ramos distintos. Apesar de identificados como a mesma espécie, segundo a
identificação molecular com base na comparação entre as sequências, erros de identificação
podem estar incluídos em análises das informações contidas nos bancos de dados,
favorecendo consequentemente erros de identificação. Trabalhos anteriores indicam que a
espécie C. theobromicola é filogeneticamente distante das outras espécies do complexo C.
gloeosporioides, informação confirmada no resultado deste estudo, onde é possível observar

80
que C. theobromicola apresenta-se distribuída em um ramo distinto de C. siamense e C.
gloeosporioides (NELSON et al., 2013). Apesar das incoerências em agrupamentos
observados em árvores filogenéticas em relação à identificação de espécies, a análise
filogenética pela distribuição dos isolados e realizada pela inclusão de dados de sequência
sem informações da identificação de espécies, gerando a distribuição da informação de modo
imparcial. Adicionalmente, a confiabilidade nas distribuições dos isolados é dada pelo
elevado bootstrap aliado ao perfil esperado na distribuição do grupo externo, o qual deve ser
filogeneticamente distante dos demais isolados em análise. Neste caso, tanto a identificação
de espécies como a análise filogenética deve ser preferencialmente realizada pela
concatenação de diversos genes (SANGPUEAK et al., 2017).
A análise da diversidade genética com a amplificação de marcadores moleculares,
além de permitir a visualização da variação e de sua distribuição intra e interespecífica,
contribui também nas inferências acerca da variação epidemiológica (patogenicidade e
virulência) de isolados de Colletotrichum em populações de culturas importantes, como por
exemplo, cana-de açúcar, sorgo, pimenta e leguminosas (SSERUMAGA et al., 2013;
MAHMODI et al., 2014; SEXENA et al, 2014; SILVEIRA, 2015; PATEL et al., 2017). Neste
âmbito, as informações obtidas pela análise da diversidade são úteis para as definições de
manejo e melhoramento visando à resistência a fitopatógenos.
Os marcadores moleculares ISSR forneceram informações sobre a variabilidade
multiloci para a verificação da diversidade genética entre os isolados de Colletotrichum
amplamente coletados em seringueiras. Este marcador tem se mostrado eficiente na detecção
da variação genômica em regiões entre sequências microssatélites em isolados deste gênero,
sendo superior a outros marcadores dominantes como o RAPD (Random Amplified
Polymorphic DNA) em relação à capacidade de detecção do polimorfismo em análises
interespecíficas (MAHMODI et al., 2014). Em cana-de-açucar a utilização combinada de dois
marcadores moleculares foi altamente eficiente para a análise da variabilidade intra-específica
entre isolados da espécie C. falcutum (PATEL et al., 2017).
De modo geral, a presença e distribuição de dinucleotídeos microssatélites no genoma
de eucariotos são maiores em relação a monômeros com três, quatro, cinco ou mais bases.
Neste sentido, os primers para motivos microssatélites com monômeros dinucleotídicos
amplificam mais loci revelando níveis de polimorfismo a depender da estrutura genética da
população. O estudo demonstrou que sete dos oito primers utilizados, amplificaram com bom
perfil loci baseados em dinucleotídeos reforçando hipótese da maior ocorrência destes
microssatélites no genoma. Maior polimorfismo é observado na análise inter-específica,

81
principalmente quando os isolados que formam o grupo amostral em análise são provenientes
de hospedeiros distintos (MAHMOD et al., 2014). Neste caso, as variações na resistência das
culturas a patógenos ampliam a diversidade genômica deste último pelo fato dos eventos
evolutivos e de seleção. Contudo, as variações ambientais também possuem efeito nas
modificações genômicas ao longo do tempo, contribuindo com o aumento da variabilidade e
diminuição de monomorfismos observados em populações intraespecíficas
(RATANACHERDCHAI et al., 2010).
O número de loci polimórficos mesmo com a utilização de poucos primers tem sido
suficiente para analisar as distâncias genéticas entre os isolados de Colletotrichum
previamente identificados. A utilização de apenas sete iniciadores ISSR da série UBC foram
utilizados por Mahmor et al. (2014) revelando polimorfismo suficientemente discriminatório
para três espécies distintas: C. gloeosporioides, C. dematium e C. truncatum. Contudo, o
aumento do número de primers e o número de loci polimórficos avaliados geram grupos mais
fidedignos a variação genética entre espécies e também entre variantes inter-específicos
(PATEL et al., 2017).
A distribuição dos isolados no dendrograma revelou a formação de dois grandes
grupos, sendo as espécies de C. acutatum distribuídas em um subgrupo específico. A
formação de grupos e subgrupos específicos a espécies é relatado em trabalhos com
marcadores dominantes (SAXENA et al., 2014; SILVEIRA et al., 2016; MAHMOD et al.,
2017). O elevado coeficiente de correlação cofrenética certifica a distribuição correta dos
isolados na representação gráfica, uma vez que o compara com os dados da matriz de
presença e ausência de loci na análise (SOUSA et al., 2015). Contudo, a distribuição do
isolado ―04‖ (C. theobromicola) no grupo de específico do complexo C. acutatum,
juntamente com a distribuição em ramos distintos dos isolados de C. theobromicola na árvore
filogenética (isolado ―10‖ e ―04‖), pode indicar erros de identificação do isolado ―04‖. A
identificação dos isolados na denominação de espécies é realizada pela identificação
molecular via comparação de muitas sequências gênicas (MAHMOD et al., 2017). Por outro
lado, o diagnóstico de espécies com primers específicos é o método de identificação mais
efetivo (SAXENA et al., 2014; SILVEIRA et al., 2016). Dificuldades e erros de identificação
de isolados são comuns quando poucas sequências conservadas são analisadas, principalmente
quando diversas espécies são sugeridas pela análise comparativa com dados depositados em
banco de dados. Apesar desses erros na identificação a análise da diversidade genética de
isolados revelam o perfil da variação no patossistema.

82
Nesse trabalho a separação entre os isolados pertencentes ao complexo C.
gloeosporioides e C. acutatum demonstram a grande dissimilaridade genética entre os dois
complexos. Dissimilaridades entre esses complexos foram observadas com o uso de
marcadores moleculares dominantes (SILVEIRA et al., 2016). Observou-se que a única
espécie do complexo C. boninense foi distribuída juntamente com isolados pertencentes ao
complexo C. gloeosporioides, sugerindo maior relação entre os loci ISSR entre os complexos
de C. gloeosporioides e C. boninense em relação ao complexo C. acutatum.
A análise de componentes principais (PCA) fornece uma visão distinta da distribuição
dos dados de distância genética em relação ao observado por outros métodos de distribuição e
agrupamento, como o método UPGMA (SOUSA et al., 2015; PATEL et al., 2017). O gráfico
de PCA distinguiu os isolados em dois grandes grupos, separando C. acutatum do grupo de C.
gloeosporioides, sugerindo que a estrutura genética em regiões ISSR é variável entre os
genótipos, com dois domínios de diversidade entre o grupo amostral de isolados aqui
analisados. A distribuição da variação em loci ISSR pelo gráfico PCA tem sido considerada
um suporte adicional ao estudo da variação, reforçando a variabilidade demonstrada por
outros métodos de agrupamento (PATEL et al., 2017).

83
5.5 REFERÊNCIAS BIBLIOGRÁFICAS
AFANADOR-KAFURI, L.; MINZ, D.; MAYMON, M.; FREEMAN, S. Characterization
of Colletotrichum isolates from tamarillo, passiflora and mango in Colombia and
identification of a unique species from the genus. Phytopathology 93, 579– 87. 2003.
ALAM, A.; ADHIKARY, S.K.; AHMED, M. Morphological Characterization
of Colletotrichum gloeosporioides Identified from Anthracnose of Mangifera indica L. Asian Journal of Plant Pathology. Vol. 1. Pp. 102-117. 2017.
ANDERSON, J.M.; AITKENB, E. A. B.; DANNC, E. K.; COATESA, L. M. Morphological
and molecular diversity of Colletotrichum spp. causing pepper spot and anthracnose of
lychee (Litchi chinensis) in Australia. Plant Pathology. Vol.62. Pp. 279–288. 2013.
ANDRADE, EM.; UESUGI CH.; UENO, B, FERREIRA, MASV. Caracterização
morfocultural e molecular de isolados de Colletotrichum gloeosporioides patogênicos ao
mamoeiro. Fitopatologia brasileira. v. 32, n.1. 2007.
BAILEY, A. J.; JEGER, J. M. Colletotrichum: biology, pathology and control. Oxford:
British Society for Plant Pathology, 1992. 388p.
BARTH, S.; MELCHINGER, A.E.; LÜBBERSTEDT, T. Genetic diversity in Arabidopsis
thaliana L. Heynh. Investigated by cleaved amplified polymorphic sequence (CAPS) and
inter-simple sequence repeat (ISSR) markers. Molecular Ecology, v.11, p.495-505, jul.
2002.
BONETT, L. P.; ALMEIDA, M.; GONÇALVES, R. G. A.; AQUINO, T. F.; WENZEL, J.B.
Caracterização morfocultural e infecção cruzada de Colletotrichum gloeosporioides
agente causal da antracnose de frutos e hortaliças em pós-colheita. Ambiência,
Guarapuava - PR. v.6 n.3 p. 451 – 463, 2010.
BUENO, C. R. N. C. Identificação e caracterização das espécies de Colletotrichum
causadoras de antracnose em hortaliças e solanáceas. 2005. 61 f. Dissertação (Mestrado em
Agronomia/Fitopatologia)-Escola Superior de Agricultura Luiz de Queiroz, Universidade de
São Paulo, Piracicaba, 2005.
CAI, L.; GIRAUD, T; ZHANG, N.; BEGEROW, D.; CAI, G.; SHIVAS, R. G. 2011. The
evolution of species concepts and species recognition criteria in plant pathogenic fungi. Fungal Diversity, v.50, p. 121–133. 2011.
CAI, Z. Y.; LIU, Y. X.; SHI, Y. P.; UM, H.J.; LI, G.H. First Report of Leaf Anthracnose
Caused by Colletotrichum karstii of Rubber Tree in China. Plant Disease, v. 100, n. 12.
2016.
CANNON, P. F.; BUDDIE, A. G.; BRIDGE, P. D. The typification of Colletotrichum
gloeosporioides. Mycotaxon, Ithaca, 104:189-204, 2008.
CANNON, P.F.; DAMM, U.; JOHNSTON, P.R.; WEIR, B.S. Colletotrichum – current
status and future directions. Studies in Mycology,73:181–213. 2012.

84
CARPENTER, J. B; STEVENSON, J. A.A secondary leaf spot of the Hevea rubber tree
caused by Glomerella cingulata. Plant Disease Reporter, v. 38, n. 7, p. 494-499, 1954.
CARVALHO, F.M.S. Caracterização cultural, morfológica e genética de espécies
de Colletotrichum associadas a doenças em macieira. Dissertação de Mestrado. Londrina.
Universidade Estadual de Londrina. 1997.
CASTELLANI A. 1976. Maintenance and cultivation of the common pathogenic fungi of
man in sterile water. Further researches. The American Journal of Tropical Medicine and
Hygiene. v. 70, p. 181 - 184.
CROUCH , J. A.; CLARKE, B. B.; WHITE, J. F. J.; HILLMAN, B. I. Systematic analysis of
the falcate-spored graminicolous Colletotrichum and a description of six new species of
the fungus from warm season grasses. Mycologia 101: 717–732. 2009.
DAMM, U.; CANNON, P.F.; WOUDENBERG, J.H.C.; CROUS, P.W. The Colletotrichum
acutatum species complex. Studies in Mycology. 73, 37–113. 46, 2012a.
DAMM, U.; CANNON, P.F.; WOUDENBERG, J.H.C.; JOHNSTON, P.R.; WEIR, B.S.; TAN,
Y.P.; SHIVAS, R.G.; CROUS, P.W. The Colletotrichum boninense species complex. Studies
in Mycology. 73, 1–36. 2012b.
DEAN R.; VAN KAN, J.A. L.; PRETORIUS, Z.A, et al. The top 10 fungal pathogens in
molecular plant pathology. Molecular Plant Pathology 13: 414–430. 2012.
DU, M.; SCHARDL, C. L.; NUCKLES, E. M.; VAILLANCOURT, L. J. Using mating-type
gene sequences for improved phylogenetic resolution of Colletotrichum species
complexes. Mycologia, Lawrence, v. 97, n. 3, p. 641-658, 2005.
FARR, D.F.; ROSSMAN, A.Y. Fungal Databases. U.S. National Fungus Collections, ARS,
USDA. Disponível em: < https://nt.ars-grin.gov/fungaldatabases/> Acesso em 18 out. 2017.
FREEMAN, S.; KATAN, T.; SHABI, E. Characterization of Colletotrichum Species
Responsible for Anthracnose Diseases of Various Fruits. The American Phytopathological
Society, June, vol. 82, N. 6, 1998.
FREEMAN, S.; MINZ, D.; MAYMON, M.; ZVEIBIL, A. Genetic diversity
within Colletotrichum acutatum sensu Simmonds. Phytopathology, St. Paul. v.91, p.586-
592, 2001.
FURTADO, E. L.; SILVEIRA, A. P. Doencas da seringueira em viveiros e jardins clonais e seu
controle. In: MEDRADO, M. J. S. et al. Formação de mudas e plantio de seringueira.
Piracicaba: ESALQ, p. 52-64. 1992.
GAZIS R.; CHAVERRI, P. Diversity of fungal endophytes in leaves and stems of wild
rubber trees (Hevea brasiliensis) in Peru. Fungal Ecology, 3: 240-254. 2010.
GAZIS, R.; REHNER, S.; CHAVERRI, P. Species delimitation in fungal endophyte
diversity studies and its implications in ecological and biogeographic inferences. Molecular Ecolology, 20, pp. 3001-3013. 2011.

85
GOUVEIA, G.R.; FERREIRA, SC.; SABINO, EC.; SIQUEIRA, SAC.; PEREIRA, J.
Comparação de três protocolos distintos para extração de RNA de amostras fixadas em
formalina e emblocadas em parafina. Jornal Brassileiro de Patologia Medico
Laboratorial, Rio de Janeiro , v. 47, n. 6, p. 649-654, 2011.
GUERBER, J.C.; LIU, B.; CORRELL, J.C.; JOHNSTON, P.R. Characterization of
diversity in Colletotrichum acutatum sensulato by sequence analysis of two gene introns,
mtDNA and intron RFLPs, and mating compatibility. Mycologia, Lancaster, v.95, n.5,
p.872-895, 2003.
GUNNEL, P.; GUBLER, W.D. Taxonomy and morphology of Colletotrichum species
pathogenic to strawberry. Mycologia 84:157-165. 1992.
GUPTA, M.; CHYI, Y.S.; ROMERO-SEVERSON, J. OWEN JL. Amplificação de
marcadores de DNA de genomas evolutivamente variados usando primers simples de
repetições de seqüência simples. Theoretical Applied Genetics. 89: 998-1006. 1994.
HALLÉ, F.; OLDEMAN, R. A.; TOMLINSON, P. B. Tropical trees and forest. Berling:
Springer– Verlag, 1978. 441 p.
HSIANG, T.; GOODWIN, P. H. Ribosomal DNA sequence comparisons of Colletotrichum
graminicola from turfgrasses and other hosts. European Journal of Plant Pathology,
Dordrecht, v. 107, p. 593-599. 2001.
HUNUPOLAGAMA, D. M.; CHANDRASEKHARAN, V.; WIJESUNDERA, W. S. S.;
KATHRIARACHCHI, H. S.; FERNANDO, T. H. P. S.; WIJESUNDERA, R. L. C. Unveiling
Members of Colletotrichum acutatum Species Complex Causing Colletotrichum Leaf
Disease of Hevea brasiliensis in Sri Lanka. Currenty Microbiology. DOI 10.1007/s00284-
017-1238-6. 2017.
JAYASINGHE C.K.; FERNANDO T.H.P.S.; PRIYANKA, U.M.S. Colletotrichum
acutatum is the main cause of Colletotrichum leaf disease of rubber in Sri Lanka. Mycopathologia 137:53-56. 1997.
KIRK, P.M.; CANNON, P.F.; MINTER, D.W.; STALPERS, J.A. Dictionary of the Fungi.
10th edition. Wallingford, CAB International. 2008.
LOPEZ, A.M.Q. Taxonomia, patogênese e controle de espécies do gênero Colletotrichum.
Revisão Anual de Patologia de Plantas, v. 9, p.291-337, 2001.
MACLEAN, D.J., BRAITHWAITE, K.S., MANNERS, J.M. AND IRWING, J.A.G. How do
we identify and classify fungal pathogens in the era of DNA analysis. Advances in. Plant
Pathology.10, 207-244. 1993.
MAHMODI, F.; KADIR, J. B.; PUTEH, A.; POURDAD, S. S. NASEHI, A.; SOLEIMANI,
N. Genetic Diversity and Differentiation of Colletotrichum spp. Isolates Associated. The
Plant Pathology Journal, v.30(1),2014.
MATTHEWS, D.; MCNICOLL, J.; HARDING, K.; MILLAM, S. 5'-anchored simple-
sequence repeat primers are useful for analysing potato somatic hybrids. Plant Cell
Reports, v.19, p.210-212, 1999.

86
MCKAY SF.; FREEMAN S.; MINZ D.; MAYMON M.; SEDGLEY M.; COLLINS GC.;
SCOTT ES. Morphological, genetic, and pathogenic characterization of Colletotrichum
acutatum, the cause of anthracnose of almond in Australia. Phytopathology, Aug;
99(8):985-95. 2009.
MENEZES, M. Aspectos biológicos e taxonômicos de espécies do gênero Colletotrichum.
Anais da Academia Pernambucana de Ciência Agronômica 3:170-179. 2006.
MILLS, P.R.; SREENIVASAPRASAD, S.; BROWN, A.E. Detection and differentiation of
Colletotrichum gloeosporioides isolates using PCR. FEMS Microbiology Letters,
Amsterdam, v.98, n.1/3, p.137-144, 1992.
MORIWAKI, J.; SATO, T.; TSUKIBOSHI, T. Morphological and molecular
characterization of Colletotrichum boninense sp. nov. from Japan. Mycoscience, Tokyo,
v. 44, n. 1, p. 47-53. 2003.
NAGAOKA, T.; OGIHARA Y. Applicability of inter-simple sequence repeat
polymorphisms in wheat for use as DNA markers in comparison to RFLP and RAPD
markers. Theoretical and Applied Genetics, New York, v. 94, p. 597-602, 1997.
NGUYEN, P. T. H.; PETTERSSON, O. V.; OLSSON, P.; LILJEROTH, E. Identification of
Colletotrichum species associated with anthracnose disease of coffee in Vietnam. European Journal of Plant Pathology, Dordrecht, v. 127, p. 73- 87, 2010.
PATEL, P.; RAJKUMAR, B.K.; PARMAR, P.; SHAH, R.; KRISHNAMURTHY, R.
Assessment of genetic diversity in Colletotrichum falcatum Went accessions based on
RAPD and ISSR markers. Journal of Genetic Engineering and Biotechnology. DOI:
https://doi.org/10.1016/j.jgeb.2017.11.006. 2017.
PERES, N.A.R.; KURAMAE, E.E.; DIAS, M.S.C.; SOUZA, N.L. Identification and
characterization of Colletotrichum spp. affecting fruit after harvest in Brazil. Journal of
Phytopathology 150:128-134. 2002.
PETCH, T. The Diseases and Pests of the Rubber Tree. MacMillan. London. 278 p. 1921.
PIMENTA, A. A. Caracterização morfométrica, patogênica e genética de isolados de
Colletotrichum gloeosporioides, agente causal da antracnose em manga (Mangifera indica
L.), 79 f. Dissertaçao (mestrado). Universidade Estadual Paulista Julho de Mesquita Filho,
Jaboticabal. 2009.
RAMPERSAD, S. N. Estrutura genética de Colletotrichum gloeosporioides sensu lato
isola infectando papaya inferido por marcadores ISSR multilocus. Phytopathology 103:
182-189. 2013.
RAMPERSAD, S. N. Genetic structure of Colletotrichum gloeosporioides sensu lato
isolates infecting papaya inferred by multilocus ISSR markers. Phytopathology, Ithaca, v.
103, n. 2, p.182-189, 2013.
RATANACHERDCHAI, K.; WANG, H. K.; LIN, F. C.; SOYTONG, K. ISSR for
comparison of cross-inoculation potential of Colletotrichum capsici causing Chilli
Anthracnose. African Journal of Microbiology. Res. 4:76-83. 2010.

87
ROHLF, F.J. NTSYS.PC: numerical taxonomy and multivariate analysis system. Version
2.1. New York: Exeter, 2000.
SAHA, T.; KUMAR, A.; RAVINDRAN, M.; JACOB, C. K.; ROY, B.; NAZEER, M. A.
Identification of Colletotrichum acutatum from rubber using random amplified
polymorphic DNAs and ribosomal DNA polymorphisms. Mycological Research 106:215-
221. 2002.
SANGPUEAK, R.; PHANSAK, P.; BUENSANTEAI, N. Morphological and molecular
identification of Colletotrichum species associated with cassava anthracnose in Thailand. Journal of Phytopathology. 1–14. 2017.
SARMIENTO, S. S. Identificação de espécies de Colletotrichum associados à Antracnose
Foliar da seringueira. Dissertação (Mestrado em Etiologia; Epidemiologia; Controle) -
Universidade Federal de Viçosa, Viçosa. 83 f. 2013.
SAXENA, A.; RAGHUWANSHI, R.; SINGH, H. B. Molecular, phenotypic and
pathogenic variability in Colletotrichum isolates of subtropical region in north-eastern
India, causing fruit rot of chillies. Journal of Applied Microbiology. Vol. 117, Pp. 1422—
1434. 2014.
SHARMA, P.N.; KATOCH, A.; SHARMA, P.; SHARMA, S.K.; SHARMA, O.P. First
Report on Association of Colletotrichum coccodes with Chili Anthracnose in India. Plant
Disease. 95:12, Pp.1584-1584. 2011.
SIERRA HAYER, J. F. Variabilidade genética de Colletotrichum gloeosporioides e
Colletotrichum acutatum em seringueira (Hevea brasiliensis) no Brasil. Tese (doutorado) -
Universidade Estadual Paulista Júlio de Mesquita Filho Faculdade de Ciências Agronômicas,
55 f. 2014.
SILVEIRA, A. L.; STRACIERI, J.; PEREIRA, F. D.; SOUZA, A.; GOES, A.
Caracterização molecular de isolados de Colletotrichum spp. associados a podridão
floral dos citros. Revista Brasileira de Fruticultura, 38(1), 64-71. 2016.
SILVEIRA, A. P.; CARDOSO, R. M. G. Ocorrência de Colletotrichum gloeosporioides em
Seringueira (Hevea brasiliensis) no Estado de São Paulo. Summa Phytopathologica,
Piracicaba, v. 13, n. 1/2:19, 1987.
SILVEIRA, A. P.; FURTADO, E. L; LOPES, M. E. B. M. Antracnose: nova doença do
painel de sangria da seringueira. São Paulo: Instituto Biológico, v. 18, p. 196-200, 1992a.
SILVEIRA, A.L.; STRACIERI, J.; PEREIRA, F. D.; SOUZA, A.; GOES, A.
Caracterização molecular de isolados de Colletotrichum spp. associados a podridão
floral dos citros. Revista Brasileira de Frutiultura. Jaboticabal, SP, v.38, n. 1. Pp. 064-071.
2016.
SMITH, B. J.; BLACK, L. L. Morphological, cultural and pathogenic variation
among Colletotrichum sp. isolated from strawberry. Plant Disease 74:69-76. 1990.
SOUSA, A. G. R.; SOUZA, M. M.; MELO, C. A. F.; SODRÉ, G. A. ISSR markers in wild
species of Passiflora L. (Passifloraceae) as a tool for taxon selection in ornamental
breeding. Genetics and Molecular Research, v. 14, p. 18534-18545, 2015.

88
SREENIVASAPRASAD, S.; SHARADA, K.; BROWN, A.E.; MILLS, P.R. PCR based
detection of Colletotrichum acutatum on strawberry. Plant Pathology, London, v.45, n.4,
p.650-655, 1996.
SSERUMAGA, J.P.; BIRUMA, M.; AKWERO, A.; OKORI, P.; EDEMA, R. Genetic
characterisation of Ugandan strains of Colletotrichum sublineolum using ISSR makers.
Journal of Agricultural Sciences, Uganda. 14 (1): 111 – 123. 2013.
STRACIERI, J. Caracterização Morfocultural e Molecular De Colletotrichum spp.
Associados a Antracnose em Manga, Mamão e Goiaba. Tese de doutorado. Universidade
Estadual Paulista - UNESP Câmpus De Jaboticabal. 2015.
SUTTON, B.C. The genus Glomerella and its anamorph Colletotrichum. In: BAILEY, J.A.;
JEGER, M.J. (Ed.). Colletotrichum: biology, pathology and control. Oxon: CAB
International, 1992. p. 1-26.
TALHINHAS, P.; SREENIVASAPRASAD, S.; NEVES-MARTINS, J.; OLIVEIRA, H.
Genetic and morphological chacracterization of Colletotrichum acutatum causing
anthracnose of lupins. Phytopathology. v.92, p. 986-996. 2002.
THAUNG, M.M. Biodiversity survey of coelomycetes in Burma. Australas. Mycological.
27: 74-110p. 2008.
WEIR, B.S.; JOHNSTON, P.R.; DAMM, U. The Colletotrichum gloeosporioides species
complex. Studies in Mycology. 73, 115–180. 2012.
WHITE, T.J.; BRUNS, T.; TAYLOR, J. Amplification and direct sequencing of fungal
ribosomal RNA genes for phylogenetics. In: Innis, M.A., Gelfand, D.H., Shinsky, J.J. &
White, T.J. (Eds.) PCR Protocols: A guide to methods and applications. New York.
Academic Press. pp. 315-322. 1990.
XIE, L.; ZHANG, J.; WAN, Y.; HU, D. Identification of Colletotrichum spp. isolated
from strawberry in Zhejiang Province and Shanghai City, China. Journal of Zhejiang
University-SCIENCE B (Biomedicine & Biotechnology). 11(1):61-70. 2010.

89
CAPÍTULO IV
6. RESISTÊNCIA DE GENÓTIPOS DE SERINGUEIRA À ANTRACNOSE FOLIAR
RESUMO
Trinta e nove genótipos de seringueira (Hevea brasiliensis) foram avaliados quanto à
sensibilidade a quatro isolados de Colletotrichum, pertencentes a três complexos de espécies
(C. gloeosporioides, C. boninense e C. acutatum). As inoculações foram realizadas depositando
gotas da suspensão de 3x105 conídios/mL em folíolos destacados dos genótipos, e estes
mantidos em condições controladas durante todo o experimento. As avaliações foram
realizadas diariamente de quatro a sete dias após a inoculação, através da mensuração ortogonal
do diâmetro das lesões para obtenção do diâmetro médio da lesão (DML). As reações de cada
acesso variaram com o isolado, demostrando haver variabilidade entre eles. Nenhum clone
apresentou resistência total, visto que, em algum momento após a inoculação, houve formação
de lesões, que variaram de tamanho. Os resultados permitiram inferir que existe variabilidade
na virulência entre os isolados de Colletotrichum, e também no comportamento dos genótipos
estudados, quanto aos isolados testados. O clone FDR 5788 se destacou por apresentar
isoladamente ou como parental de progênies os melhores resultados para resistência a
Colletotrichum. Este trabalho é pioneiro para o patossistema Colletotrichum x seringueira por
apresentar uma metodologia de inoculação prática, reproduzível e possível de diferenciar
genótipos resistentes e suscetíveis ao patógeno, além de demostrar que a variabilidade do
complexo de espécies de Colletotrichum interfere na busca da resistência.
Palavras-chave: Hevea brasiliensis, Colletotrichum spp., metodologia de inoculação,
patossistema.

90
ABSTRACT
Thirty-nine accessions of rubber tree (Hevea brasiliensis) were evaluated for sensitivity to four
isolates of Colletotrichum, belonging to three species complexes (C. gloeosporioides, C.
boninense and C. acutatum). The inoculations were performed by depositing drops of the
suspension of 3x105 conidia/mL on leaflets detached from the accessions, and kept under
controlled conditions throughout the experiment. The evaluations were performed daily from
four to seven days after inoculation, by orthogonal measurement of the lesion diameter to
obtain the mean diameter of the lesion (DML). The reactions of each access varied with the
isolate, demonstrating that there was variability between them. No clone showed complete
resistance, since at some time after inoculation, lesions formed, which varied in size. The
results allowed inferring that there is variability in the virulence between Colletotrichum
isolates, and also in the behavior of the studied accessions, regarding the tested isolates. FDR
5788 clone was distinguished by presenting the best results for Colletotrichum resistance alone
or as parent of progenies. This work is a pioneer for the Colletotrichum x rubber tree
pathosystem because it presents a methodology of practical, reproducible and possible
inoculation to differentiate resistant and susceptible accessions to the pathogen, besides
showing that the variability of Colletotrichum species complex interferes in the search for
resistance.
Key words: Hevea brasiliensis, Colletotrichum spp., methodology of inoculation, pathosystem.

91
6.1 INTRODUÇÃO
A heveicultura é uma atividade de grande importância econômica, social e ambiental,
pela sua capacidade de geração de emprego e renda e por contribuir para conservação do solo e
melhoria do ambiente, pois possui características típicas de uma floresta tropical (VIRGENS
FILHO, 2007; NARAYANAN e MYDIN, 2011). Além disso, a seringueira durante seu ciclo
de vida tem grande potencial para sequestro de carbono dentro da agroindústria e a madeira das
plantas adultas pode ser utilizada, constituindo assim um sistema de duplo propósito
(LAKSHMAN; MUNASINGHE, 2017; CORLEY, 1983).
O Brasil possui uma área plantada estimada em 160.968 ha hectares de seringueira,
dos quais pouco menos da metade encontra em produção (IBGE, 2017). Os principais Estados
produtores são: São Paulo, Bahia, Mato Grosso, Espírito Santo e Paraná (EMBRAPA, 2017).
Sendo o estado da Bahia, considerado o segundo maior produtor de borracha natural do país,
possuindo a segunda maior área plantada com aproximadamente 33.203 hectares (IBGE,
2017).
Entre os fatores que afetam o desenvolvimento da heveicultura no Brasil, merecem
destaque os danos causados pelas doenças fungicas, entre as quais, menciona-se a Antracnose
que é causada pelo complexo de espécies Colletotrichum, que ocorrem na maioria das regiões
heveícolas do mundo (GAPAROTTO et al., 1990; SANTOS et al., 1989; SILVEIRA et al.,
1986). A Antracnose nos últimos anos, tem se alastrado nos seringais das diversas regiões do
país causando grande prejuizos na produção (FURTADO, 2008), especialmente no Sudeste da
Bahia e no Espírito Santo onde a enfermidade vem aumentando gradativamente sua
importância econômica. O patógeno ataca plantas nas diversas fases de desenvolvimento, desde
mudas no viveiro até seringueiras adultas causando lesões necróticas, enrrugamento e queda
foliar (GASPAROTTO et al., 2012).
As principais formas de controle para esta enfermidade é o controle químico, com
utilização de fungicidas, sendo seu uso dificultado pelo porte elevado das plantas
(GASPAROTTO et al.,1985). O controle genético é uma das alternativas mais práticas e
econômicas para o manejo desta doença (GASPAROTTO et al., 2012). A primeira medida é
evitar o plantio comercial de materiais suscetíveis, e para isso é muito importante o
conhecimento do comportamento de clones comerciais quanto à resistência ou suscetibilidade
aos principais patógenos, sendo, portanto, primordial para o sucesso de um programa de
melhoramento. Na avaliação de resistência contra os patógenos, de modo geral, as variáveis

92
analisadas são diâmetro das lesões, análise do período de incubação e período latente do
patógeno nas folhas (JUNQUEIRA, 1985; BERGAMIN FILHO et al., 2011).
No Brasil, ainda são escassos os trabalhos relacionados à seleção de clones resistentes à
Antracnose. No estado de São Paulo, Furtado et al. (1994) avaliaram a queda foliar decorrente do
ataque de Colletotrichum e constataram que os clones GT1, PR 225 e IAN 873, GT1, IAN 873,
RRIM 600 e PR 107 sofreram menores perdas foliares que os clones RRIM 701, PR 261, PB 217
e PB 235 que foram mais desfolhados.
Gonçalves et al. (2002) objetivando selecionar clones de seringueira promissores para a
região do planalto do estado de São Paulo, avaliaram dentre os principais caracteres
secundários, a incidência da Antracnose do painel em 16 clones da série IAC 300, e observaram
que os clones IAC 330, IAC 331, IAC 333, IAC 336, IAC 338, IAC 339 e IAC 343 apresentaram
alta resistência à doença, ao contrário dos clones IAC 340 e RRIM 600 que foram suscetíveis.
Programas de melhoramento genético da seringueira no Brasil vêm sendo conduzidos no
estado de São Paulo pelo Instituto Agronômico (IAC) e na Bahia pelo Centro de Pesquisa do
Cacau (Cepec) (GONÇALVES; MARQUES, 2014). Em 1992, a Michelin, em colaboração com
o Centre de Coopération Internationale em Recherche Agronomique pour le Développement –
CIRAD, iniciou um programa de melhoramento genético da seringueira com o objetivo de
selecionar clones com bons níveis de resistência a Microcyclus ulei (Henn.) Arx atrelado a uma
elevada produção. Clones de seringueira estão sendo desenvolvidos para a resistência a M. ulei,
no entanto, ao longo do tempo, observou-se que muitos genótipos produtivos e parcialmente
resistentes a esta enfermidade, passaram a ser afetados pela Antracnose (STERLING et al.,
2011).
Considerando o aumento da intensidade da Antracnose nos seringais e a pouca
disponibilidade de informações envolvendo esse patossistema na região Sudeste da Bahia,
importante pólo heveícola, este trabalho foi desenvolvido com o objetivo de analisar as reações
de genótipos de seringueira, oriundos do banco de germoplasma das Plantações Michelin da
Bahia, à infecção por diferentes isolados de Colletotrichum spp., visando a seleção de materiais
resistentes para serem incorporados aos futuros trabalhos de melhoramento genético.

93
6.2 MATERIAL E MÉTODOS
Os ensaios foram realizados no laboratório de Diversidade de fungos na Seção de
Fitopatologia do Centro de Pesquisa do Cacau (Cepec) na sede da Comissão Executiva do
Plano da Lavoura Cacaueira (Ceplac), situada em Ilhéus, BA.
6.2.1 Organismos e condições de cultivo
Trinta e nove genótipos de seringueira (Tabela 1), cultivados em condições controladas,
foram obtidos do banco de germoplasma das Plantações Michelin da Bahia (PMB), localizada
no município de Igrapiúna (Latitude13º 49' 35" S/Longitude: 39º 08' 32" W), região Sudeste da
Bahia.
Quatro isolados de Colletotrichum (C. acutatum, C. gloeosporioides e C. boninense)
obtidos de folíolos da copa de seringueira com sintomas de Antracnose, e mantidos em coleção
do Laboratório de Diversidade de Fungos do Cepec/Ceplac, foram utilizados como patógenos
nos ensaios de avaliação de resistência. Os isolados foram cultivados em meio Batata-
Dextrose-Ágar (BDA), à 24º C, sob luminosidade constante.
Devido à variação na disponibilidade de material vegetal (lançamento foliar no estádio
C) para as inoculações, foram analisados a reação de 38 genótipos para o isolado 19 (C.
acutatum), 12 para o isolado 13 (C. gloeosporioides), 18 para o isolado 06 (C. boninense), e
apenas 10 genótipos para o isolado 14 (C. acutatum).
6.2.2 Inoculação e avaliação de sintomas
Folíolos no estádio C de desenvolvimento foliar (HALLÉ et al., 1978), foram coletados
e armazenados em sacos plásticos, devidamente etiquetados, contendo papel toalha umedecidos
com água destilada e acondicionados em caixas de isopor contendo gelo. No laboratório de
Diversidade de Fungos do Cepec, procedeu-se a lavagem dos folíolos com água corrente e
desinfestação com hipoclorito de sódio a 2%. Os experimentos foram conduzidos em caixas
plásticas (40 x 25 x 10 cm) contendo espumas autoclavadas, umedecidas com água destilada
esterilizada. O desenho experimental foi em blocos casualizados, com 39 tratamentos, quatro
repetições e quatro folíolos destacados por repetição, com dois pontos de inoculação por
folíolo, totalizando 32 unidades experimentais por genótipo.
Procedeu-se a inoculação de cada um dos isolados de Colletotrichum com uma alíquota
de 20 μL da suspensão do inóculo, ajustada para 3x105conídios/ mL e acrescida de Tween 80%.

94
Cada folíolo foi inoculado na face abaxial, no ápice e na base. As caixas contendo os folíolos
foram mantidas a uma temperatura de 24º C ±2, sob luz constante.
As avaliações foram realizadas a partir do quarto dia após a inoculação, até o sétimo
dia, onde foram mensurados diariamente o comprimento e largura das lesões, em milímetro,
com o auxílio de um paquímetro digital. Como testemunha foram utilizados folíolos destacados
inoculados com água destilada estéril.
6.2.3 Análise dos dados
Foram avaliados os parâmetros: período de incubação (PI) e diâmetro médio da lesão
(DML) ao longo de sete dias da incubação (DAI). Os primeiros sintomas considerados foram
às lesões necróticas observadas na superfície foliar.
Os dados da morfometria foram submetidos à análise de variância pelo teste F e as
médias do diâmetro das lesões comparadas pelo teste de Tukey a 5% de probabilidade,
utilizando-se a versão 9 do software SAS®
(Statistical Analysis System). Analisou-se
separadamente o comportamento dos clones para cada isolado utilizado.
Tabela 1. Descrição dos 39 genótipos de Hevea spp. testados para resistência à Antracnose
foliar da seringueira.
(continua)...
Genótipos Parentais Origem
PB 314 RRIM 600 x PB 235 Malásia
MDX 624 AVROS 1581 x MDF? Guatemala - Libéria
SIAL 1005 AVROS 255 x Fx 25 Brasil
SIAL 893 Família de meio-irmão do PB 86 Brasil
PMB 1 Desconhecido Brasil
Fx 2784 F 4542 X AVROS 363 Brasil
Fx 3899 F 4542 X AVROS 363 Brasil
Fx 3864 PB 86 x PB 38 Brasil
Fx 2261 F 1619 x AVROS 183 Brasil
FDR 5788 Harbel8 x MDF 180 Guatemala – Libéria
FDR 5240 Harbel 68 x TU 42-525 Guatemala – Libéria
FDR 4575 Harbel8 x FDR 18 Guatemala – Libéria
IAN 6590 Fx 43- 651 x PB 86 -
PFB 5 PRIMÁRIO Brasil
CDC 312 AVROS 308 x MDX 40 Guatemala – Libéria
CDC 308 AVROS 308 x MDX 40 Guatemala – Libéria
RRIM 600 Tjir1 x PB 86 Malásia
TP 875 IAN 873 x ? Brasil
CMB 197 MDX 608 - 04 Brasil
1711 IRCA 109 x MDX 50 Brasil

95
Tabela 1. Descrição dos 39 genótipos de Hevea spp. testados para resistência à Antracnose
foliar da seringueira.
(conclusão)
Genótipos Parentais Origem
807 RRIM 600 x FDR 5788 Brasil
1031 RRIM 600 x FDR 5788 Brasil
819 RRIM 600 x FDR 5788 Brasil
1013 RRIM 600 x FDR 5788 Brasil
959 RRIM 600 x FDR 5788 Brasil
840 RRIM 600 x FDR 5788 Brasil
1014 RRIM 600 x FDR 5788 Brasil
1030 RRIM 600 x FDR 5788 Brasil
960 RRIM 600 x FDR 5788 Brasil
1601 MDX 50 x CDC 312 Brasil
1634 RRIM 600 x PFB 5 Brasil
1023 RRIM 600 x FDR 5788 Brasil
1640 IRCA 109 x MDX 624 Brasil
1609 FDR 5788 x PFB 5 Brasil
1645 RRIM 600 x PFB 5 Brasil
1666 FDR 5788 x PMB 1 Brasil
1688 RRIM 600 x PFB 5 Brasil
1741 MDX 607 x PMB 1 Brasil
MDX 607 AVROS 1581 x MDF ? -
RRIM: Rubber Research Institute of Malaysia; PB: Prang Besar, Malásia; AVROS: Algemene Vereniging Rubber
planters Oostkust Sumatra, Indonésia; Fx: Ford Cruzamento, Brasil; MDX: Madre de Dios Cruzamento, Guatemala;
Tjir1: Tjirandji; IAN: Instituto de Pesquisas Agropecuárias do Norte, Brasil; IRCA: Institute de Recherches sur le
Caoutchou, Costa do Marfim; FDR: Firestone Dothiella Resistant; PFB: Pé franco de Belterra, Brasil; PMB:
Plantações Michelin da Bahia, Brasil; CDC: Clavellina Dothidella Resistent; Tu: Turrialba.

96
6.3 RESULTADOS
Em todos os genótipos avaliados, observou-se a formação de lesões circulares,
concêntricas de coloração amarronzadas, típicas de Colletotrichum gloeosporioides (Figura 1).
A metodologia utilizada com folíolos destacados possibilitou analisar o comportamento dos
clones em relação à infecção do patógeno ao longo de sete dias.
Houve diferença significativa (p<0,01) para a variável analisada (diâmetro médio da
lesão - DML), demonstrando a existência de variabilidade no comportamento dos genótipos
inoculados com cada um dos isolados testados.
Figura 1. Folíolo do clone PMB 1 com lesões circulares, concêntricas
de coloração amarronzadas, proveniente de inoculação com gotas de
20 µl da suspensão de esporos de Colletotrichum gloeosporioides.
Fonte: Dados da pesquisa.
O período de incubação variou de dois a cinco dias, tendo os genótipos 1601, MDX
607, 1711, SIAL 893, TP 875, PMB1, Fx 2784 e 1634, apresentado o menor período de
incubação. Em contrapartida, o maior número de genótipos, dentre os 39 avaliados, apresentou
lesões pontuais ao 3º DAI. Os genótipos que apresentaram sintomas mais tardios foram 1645,
960, 1666, 1688, 1609, FDR4575, 807, 1014 e 1023.
Houve diferença em relação ao comportamento dos genótipos quando inoculados com o
isolado 19 (Colletotrichum acutatum). A tabela 2 refere-se à avaliação de 38 genótipos de
seringueira quanto à evolução temporal da doença. As variações ocorreram em todas as leituras

97
(4 DAI à 7 DAI), no entanto, para a discussão dos dados será levada em consideração apenas a
variável diâmetro médio das lesões no quarto dia de avaliação, correspondendo ao sétimo DAI.
Os clones Fx 3864, TP 875 e PMB 1 foram os mais suscetíveis ao isolado 19, por
apresentarem respectivamente as maiores médias referentes ao diâmetro médio da lesão (26,61;
26,00; 25,77 mm). Os genótipos 1711, 1601, 1640 e os clones, já considerados comerciais, Fx
2784, SIAL 893 e RRIM 600, foram estatisticamente similares aos três primeiros e tiveram
DML variando de 25,43 a 21,62 mm. A grande maioria dos genótipos testados diferiu
estatisticamente dos mais suscetíveis, apresentando médias variando entre 8,49 mm (1609) e
21,28 (IAN 6590). Neste grupo estão clones comerciais como PFB 5, CDC 308, SIAL 1005,
FDR 5240, Fx 3899, FDR 5788 e CDC 312. Dentre os genótipos mencionados deve ser
destacado o 1609 cuja média não diferiu estatisticamente do grupo mais resistente.
Os genótipos FDR 4575, 1688, 1030, 1645 e 960 apresentaram as menores lesões com
variação de 1,00 a 7,29 mm e retardamento na evolução da doença, como pode ser observado
para as variáveis 4 DAI e 6 DAI, sendo considerados dentre os genótipos testados, os mais
promissores ao caráter resistência. Destaca-se o genótipo 960 que apresentou o menor DML
(1,00 mm).
Na tabela 3 estão expostos os resultados do DML de 18 genótipos quando inoculados
com o isolado 06 de C. boninense. A diferença mínima significativa no último dia de avaliação
foi de 8,08, quando foi possível visualizar pela diferença entre as médias apresentadas, que o
clone TP 875 foi o mais suscetível ao ataque do patógeno, com um valor médio de 24,26 mm,
não diferindo estatisticamente dos genótipos SIAL 1005, 1030 e IAN 6590, que também
apresentaram alta suscetibilidade. Um grupo intermediário foi formado pelos clones já
considerados comercias, PMB 1, Fx 3864, CDC 312, FDR 5788, RRIM 600, todos
estatisticamente semelhantes entre si, porém diferentes dos considerados mais suscetíveis e
resistentes. Para esse isolado, os genótipos mais resistentes ao patógeno, obtiveram médias
entre 1,84 mm (819) e 5,55 (1014), com destaque para os genótipos 819 e 807, que
apresentaram as menores lesões. Além disso, o crescimento das lesões foi lento e bem uniforme
nesse grupo de genótipos.
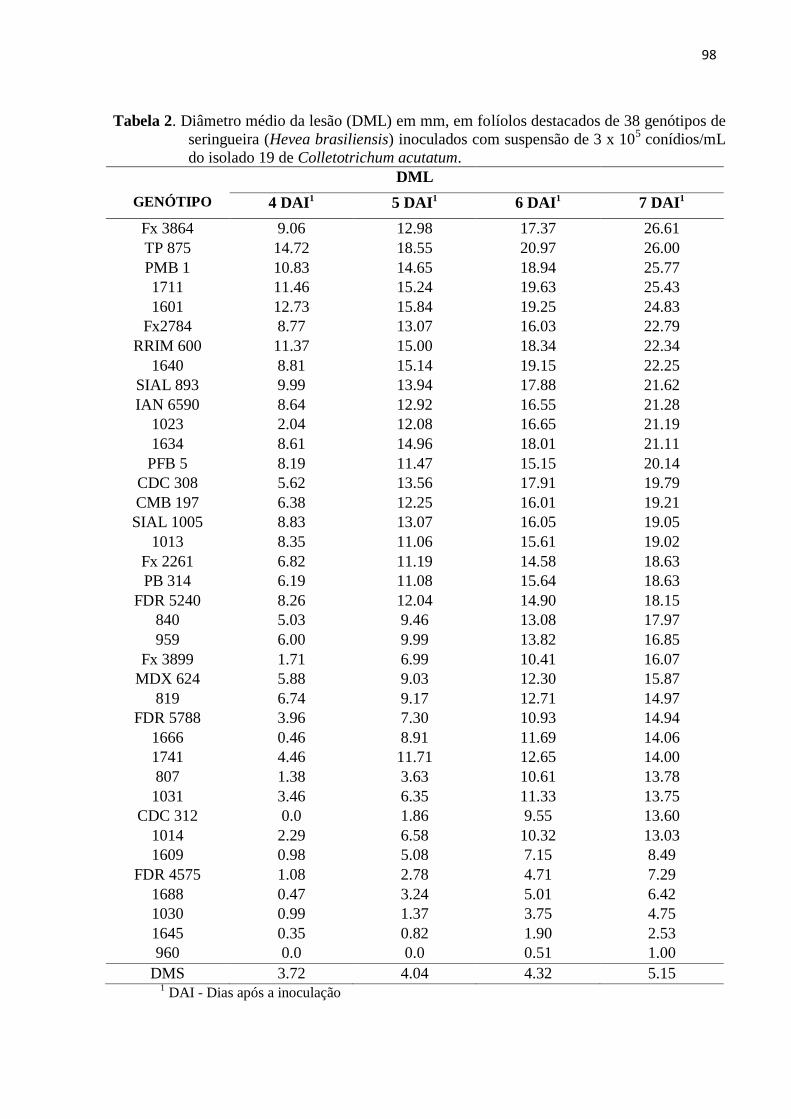
98
Tabela 2. Diâmetro médio da lesão (DML) em mm, em folíolos destacados de 38 genótipos de
seringueira (Hevea brasiliensis) inoculados com suspensão de 3 x 105 conídios/mL
do isolado 19 de Colletotrichum acutatum. DML
GENÓTIPO 4 DAI1 5 DAI1 6 DAI1 7 DAI1
Fx 3864 9.06 12.98 17.37 26.61
TP 875 14.72 18.55 20.97 26.00
PMB 1 10.83 14.65 18.94 25.77
1711 11.46 15.24 19.63 25.43
1601 12.73 15.84 19.25 24.83
Fx2784 8.77 13.07 16.03 22.79
RRIM 600 11.37 15.00 18.34 22.34
1640 8.81 15.14 19.15 22.25
SIAL 893 9.99 13.94 17.88 21.62
IAN 6590 8.64 12.92 16.55 21.28
1023 2.04 12.08 16.65 21.19
1634 8.61 14.96 18.01 21.11
PFB 5 8.19 11.47 15.15 20.14
CDC 308 5.62 13.56 17.91 19.79
CMB 197 6.38 12.25 16.01 19.21
SIAL 1005 8.83 13.07 16.05 19.05
1013 8.35 11.06 15.61 19.02
Fx 2261 6.82 11.19 14.58 18.63
PB 314 6.19 11.08 15.64 18.63
FDR 5240 8.26 12.04 14.90 18.15
840 5.03 9.46 13.08 17.97
959 6.00 9.99 13.82 16.85
Fx 3899 1.71 6.99 10.41 16.07
MDX 624 5.88 9.03 12.30 15.87
819 6.74 9.17 12.71 14.97
FDR 5788 3.96 7.30 10.93 14.94
1666 0.46 8.91 11.69 14.06
1741 4.46 11.71 12.65 14.00
807 1.38 3.63 10.61 13.78
1031 3.46 6.35 11.33 13.75
CDC 312 0.0 1.86 9.55 13.60
1014 2.29 6.58 10.32 13.03
1609 0.98 5.08 7.15 8.49
FDR 4575 1.08 2.78 4.71 7.29
1688 0.47 3.24 5.01 6.42
1030 0.99 1.37 3.75 4.75
1645 0.35 0.82 1.90 2.53
960 0.0 0.0 0.51 1.00
DMS 3.72 4.04 4.32 5.15 1 DAI - Dias após a inoculação
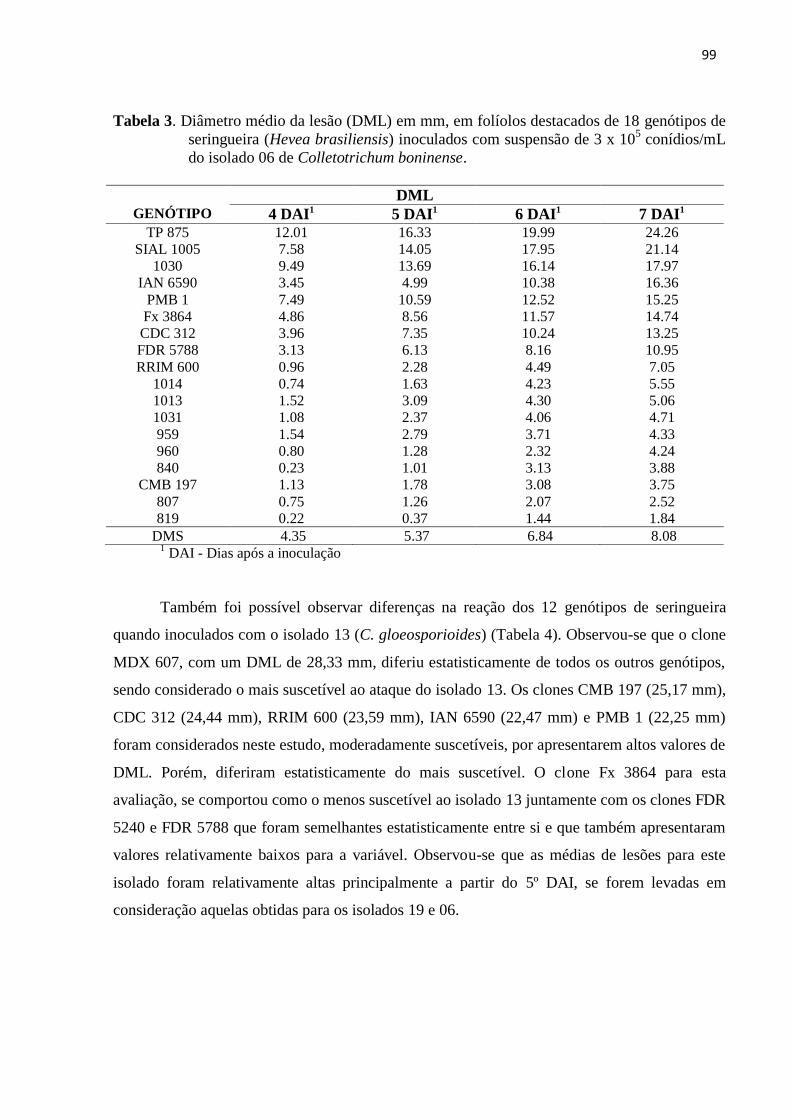
99
Tabela 3. Diâmetro médio da lesão (DML) em mm, em folíolos destacados de 18 genótipos de
seringueira (Hevea brasiliensis) inoculados com suspensão de 3 x 105 conídios/mL
do isolado 06 de Colletotrichum boninense.
DML
GENÓTIPO 4 DAI1 5 DAI1 6 DAI1 7 DAI1 TP 875 12.01 16.33 19.99 24.26
SIAL 1005 7.58 14.05 17.95 21.14
1030 9.49 13.69 16.14 17.97
IAN 6590 3.45 4.99 10.38 16.36
PMB 1 7.49 10.59 12.52 15.25
Fx 3864 4.86 8.56 11.57 14.74
CDC 312 3.96 7.35 10.24 13.25
FDR 5788 3.13 6.13 8.16 10.95
RRIM 600 0.96 2.28 4.49 7.05
1014 0.74 1.63 4.23 5.55
1013 1.52 3.09 4.30 5.06
1031 1.08 2.37 4.06 4.71
959 1.54 2.79 3.71 4.33
960 0.80 1.28 2.32 4.24
840 0.23 1.01 3.13 3.88
CMB 197 1.13 1.78 3.08 3.75
807 0.75 1.26 2.07 2.52
819 0.22 0.37 1.44 1.84
DMS 4.35 5.37 6.84 8.08 1 DAI - Dias após a inoculação
Também foi possível observar diferenças na reação dos 12 genótipos de seringueira
quando inoculados com o isolado 13 (C. gloeosporioides) (Tabela 4). Observou-se que o clone
MDX 607, com um DML de 28,33 mm, diferiu estatisticamente de todos os outros genótipos,
sendo considerado o mais suscetível ao ataque do isolado 13. Os clones CMB 197 (25,17 mm),
CDC 312 (24,44 mm), RRIM 600 (23,59 mm), IAN 6590 (22,47 mm) e PMB 1 (22,25 mm)
foram considerados neste estudo, moderadamente suscetíveis, por apresentarem altos valores de
DML. Porém, diferiram estatisticamente do mais suscetível. O clone Fx 3864 para esta
avaliação, se comportou como o menos suscetível ao isolado 13 juntamente com os clones FDR
5240 e FDR 5788 que foram semelhantes estatisticamente entre si e que também apresentaram
valores relativamente baixos para a variável. Observou-se que as médias de lesões para este
isolado foram relativamente altas principalmente a partir do 5º DAI, se forem levadas em
consideração aquelas obtidas para os isolados 19 e 06.
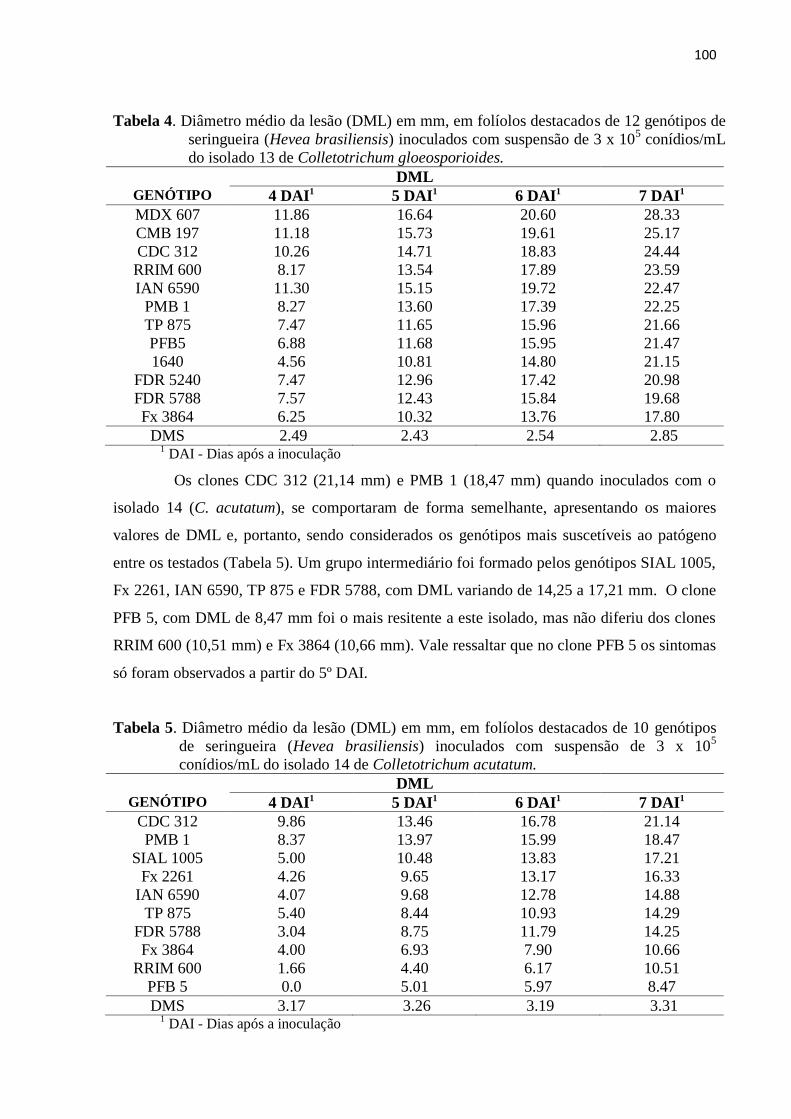
100
Tabela 4. Diâmetro médio da lesão (DML) em mm, em folíolos destacados de 12 genótipos de
seringueira (Hevea brasiliensis) inoculados com suspensão de 3 x 105 conídios/mL
do isolado 13 de Colletotrichum gloeosporioides.
DML
GENÓTIPO 4 DAI1 5 DAI1 6 DAI1 7 DAI1
MDX 607 11.86 16.64 20.60 28.33
CMB 197 11.18 15.73 19.61 25.17
CDC 312 10.26 14.71 18.83 24.44
RRIM 600 8.17 13.54 17.89 23.59
IAN 6590 11.30 15.15 19.72 22.47
PMB 1 8.27 13.60 17.39 22.25
TP 875 7.47 11.65 15.96 21.66
PFB5 6.88 11.68 15.95 21.47
1640 4.56 10.81 14.80 21.15
FDR 5240 7.47 12.96 17.42 20.98
FDR 5788 7.57 12.43 15.84 19.68
Fx 3864 6.25 10.32 13.76 17.80
DMS 2.49 2.43 2.54 2.85 1 DAI - Dias após a inoculação
Os clones CDC 312 (21,14 mm) e PMB 1 (18,47 mm) quando inoculados com o
isolado 14 (C. acutatum), se comportaram de forma semelhante, apresentando os maiores
valores de DML e, portanto, sendo considerados os genótipos mais suscetíveis ao patógeno
entre os testados (Tabela 5). Um grupo intermediário foi formado pelos genótipos SIAL 1005,
Fx 2261, IAN 6590, TP 875 e FDR 5788, com DML variando de 14,25 a 17,21 mm. O clone
PFB 5, com DML de 8,47 mm foi o mais resitente a este isolado, mas não diferiu dos clones
RRIM 600 (10,51 mm) e Fx 3864 (10,66 mm). Vale ressaltar que no clone PFB 5 os sintomas
só foram observados a partir do 5º DAI.
Tabela 5. Diâmetro médio da lesão (DML) em mm, em folíolos destacados de 10 genótipos
de seringueira (Hevea brasiliensis) inoculados com suspensão de 3 x 105
conídios/mL do isolado 14 de Colletotrichum acutatum. DML
GENÓTIPO 4 DAI1 5 DAI1 6 DAI1 7 DAI1
CDC 312 9.86 13.46 16.78 21.14
PMB 1 8.37 13.97 15.99 18.47
SIAL 1005 5.00 10.48 13.83 17.21
Fx 2261 4.26 9.65 13.17 16.33
IAN 6590 4.07 9.68 12.78 14.88
TP 875 5.40 8.44 10.93 14.29
FDR 5788 3.04 8.75 11.79 14.25
Fx 3864 4.00 6.93 7.90 10.66
RRIM 600 1.66 4.40 6.17 10.51
PFB 5 0.0 5.01 5.97 8.47
DMS 3.17 3.26 3.19 3.31 1 DAI - Dias após a inoculação

101
Uma análise de agrupamento feita pelo método UPGMA, analisou a reação dos
genótipos com relação aos quatro isolados de Colletotrichum spp. considerando o DML nos
quatro dias de avaliação. A figura 2 refere-se ao dendrograma que representa a dissimilaridade
entre 38 genótipos de seringueira quanto à evolução temporal da doença, quando inoculados
com o isolado 19 (Colletotrichum acutatum).
A linha vermelha vertical no dendrograma determina o ponto de corte estabelecido (10),
que equivale a 10 % de distância total determinada pelo método de Mojema (1977).
Foram estabelecidos três grupos distintos A, B e C separando os genótipos em
resistentes, altamente suscetíveis e suscetíveis, respectivamente. Após o corte, evidenciou-se a
divisão dos grupos, em cinco subgrupos (A-I, A-II, B-III, B-IV e C-V) compostos por 3, 5, 7,
14 e 9 genótipos respectivamente. O subgrupo A-II está representado pelo ramo onde estão
presentes os genótipos mais resistentes ao isolado 19, apresentando uma alta similaridade entre
os genótipos 960 e 1645; 1688 e FDR4575. Os genótipos 1609, CDC312 e 807 compõem o
subgrupo A-I, e também fazem parte de um ramo composto por genótipos que apresentaram
resistência ao isolado 19. Os genótipos mais suscetíveis (1601, 1711, PMB1, 1640, RRIM 600,
Fx 3864 e TP875), estão no subgrupo B-III, sendo o TP875 o que se apresentou mais distante
dos demais genótipos do mesmo grupo, em função de ter causado as maiores lesões nas
primeiras avaliações. O grupo B-III é um grupo intermediário, formado pela maioria dos
genótipos avaliados, e o 1023 foi o genótipo que mais distanciou-se dos demais, por apresentar
as menores lesões no 4 DAI.
O dendrograma da dissimilaridade entre 12 genótipos de seringueira quanto à reação ao
isolado 13 de C. gloeosporiodes (Figura 3) é composto por dois grupos A e B, subdivididos em
cinco subgrupos. Os genótipos Fx 3864 e 1640, pertencentes ao grupo B-V, apresentaram um
comportamento resistente para quatro variáveis estudadas, em oposição ao grupo A-I e A-II,
que apresentaram padrões mais altos de suscetibilidade ao isolado, com destaque para o
genótipo MDX 607 que foi o mais suscetível, distanciando-se dos demais genótipos
pertencentes ao grupo. Os clones FDR 5240 e FDR 5788 fazem parte de dois subgrupos (B-III
e B-IV) que apresentaram sensibilidade média ao isolado, sendo que dentro de seus respectivos
subgrupos, estes foram os que mais se distanciaram dos demais genótipos agrupados.
O isolado 14 de C. acutatum (Figura 4) gerou um dendrograma composto por dois
grupos distintos subdivididos em três subgrupos. O grupo B-III separa os genótipos com menor
sensibilidade ao patógeno (Fx 3864, RRIM 600 e PFB 5). Dentro do subgrupo dos genótipos
mais sensíveis (A-I), estão o PMB1 e CDC 312. Um grupo intermediário definido (A-II) foi

102
formado por genótipos que apresentaram sensibilidade média ao isolado, dentre eles, o SIAL
1005 foi o que se mostrou mais distante dos demais genótipos pertencentes ao mesmo grupo.
O dendrograma da análise de agrupamento, a partir das medidas de dissimilaridade
entre 18 genótipos inoculados com o isolado 06 de C. boninense (Figura 5) permitiu a
visualização de dois grupos principais A e B suscetíveis e resistentes respectivamente, que
foram subdivididos em três subgrupos A-I, A-II e B-III. O subgrupo A-I composto por cinco
genótipos (CDC 312, IAN 6590, Fx 3864, FDR 5788 e PMB1), comporta os de menor
suscetibilidade ao patógeno, em oposição ao grupo B-III que contêm os genótipos mais
suscetíveis (1030, SIAL 1005, TP 875). Embora os clones Fx 3864 e PMB1 sejam
considerados suscetíveis ao ataque de Colletotrichum, para esta avaliação eles foram
enquadrados no subgrupo dos mais resistentes, sendo o PMB 1 que mais distanciou-se dos
demais genótipos testados. Um grupo intermediário A-II composto por 10 genótipos apresentou
distância muito curta entre os genótipos testados, como por exemplo, 1013 e 1031, com
destaque para o genótipo 819 que mais se distanciou dos demais testados.
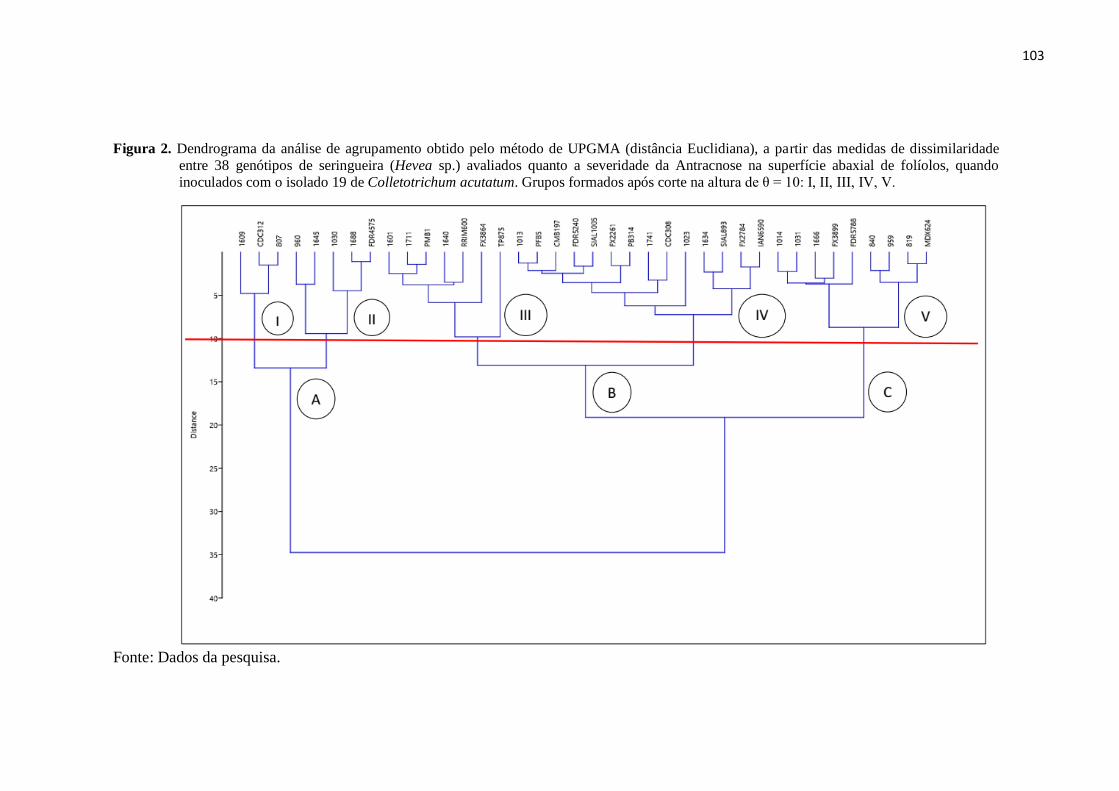
103
Figura 2. Dendrograma da análise de agrupamento obtido pelo método de UPGMA (distância Euclidiana), a partir das medidas de dissimilaridade
entre 38 genótipos de seringueira (Hevea sp.) avaliados quanto a severidade da Antracnose na superfície abaxial de folíolos, quando
inoculados com o isolado 19 de Colletotrichum acutatum. Grupos formados após corte na altura de θ = 10: I, II, III, IV, V.
Fonte: Dados da pesquisa.
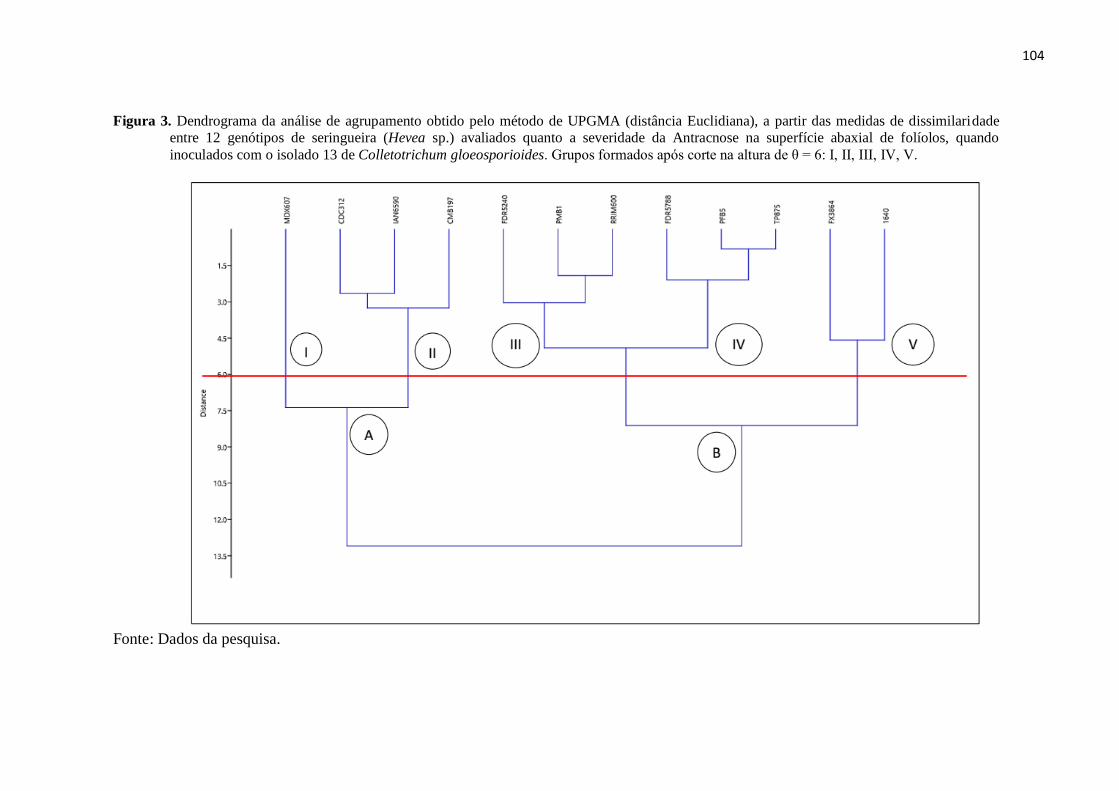
104
Figura 3. Dendrograma da análise de agrupamento obtido pelo método de UPGMA (distância Euclidiana), a partir das medidas de dissimilaridade
entre 12 genótipos de seringueira (Hevea sp.) avaliados quanto a severidade da Antracnose na superfície abaxial de folíolos, quando
inoculados com o isolado 13 de Colletotrichum gloeosporioides. Grupos formados após corte na altura de θ = 6: I, II, III, IV, V.
Fonte: Dados da pesquisa.
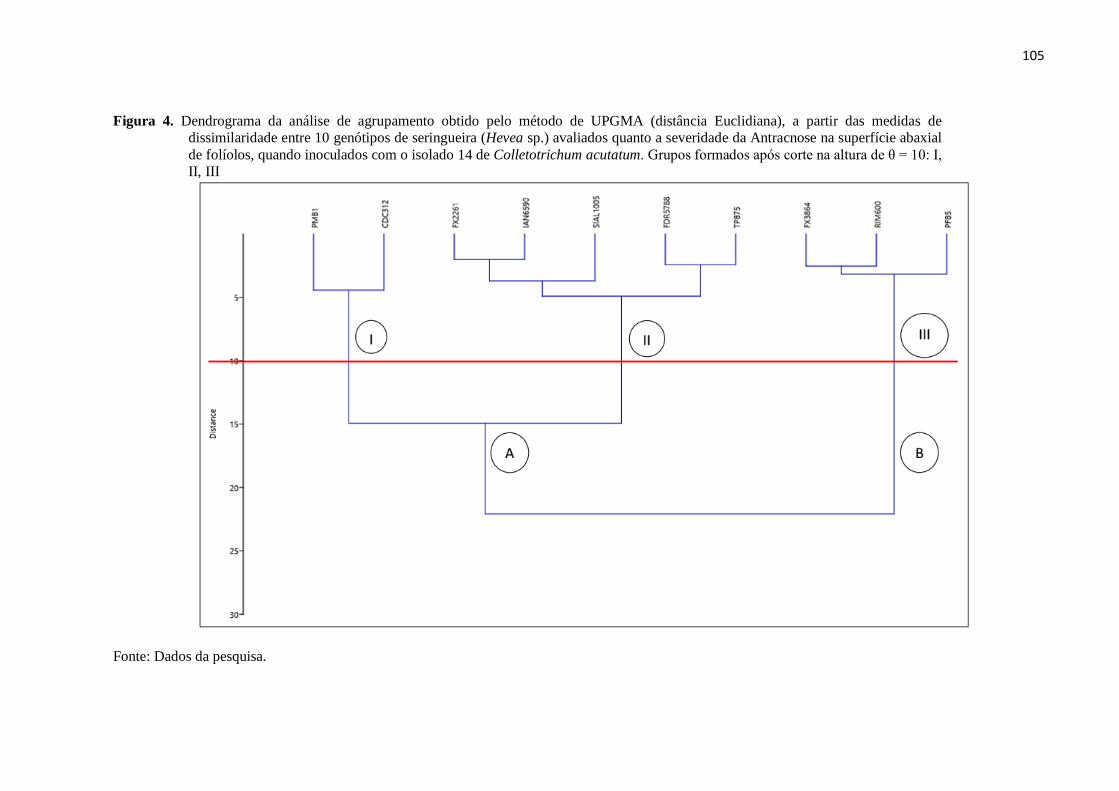
105
Figura 4. Dendrograma da análise de agrupamento obtido pelo método de UPGMA (distância Euclidiana), a partir das medidas de
dissimilaridade entre 10 genótipos de seringueira (Hevea sp.) avaliados quanto a severidade da Antracnose na superfície abaxial
de folíolos, quando inoculados com o isolado 14 de Colletotrichum acutatum. Grupos formados após corte na altura de θ = 10: I,
II, III
Fonte: Dados da pesquisa.
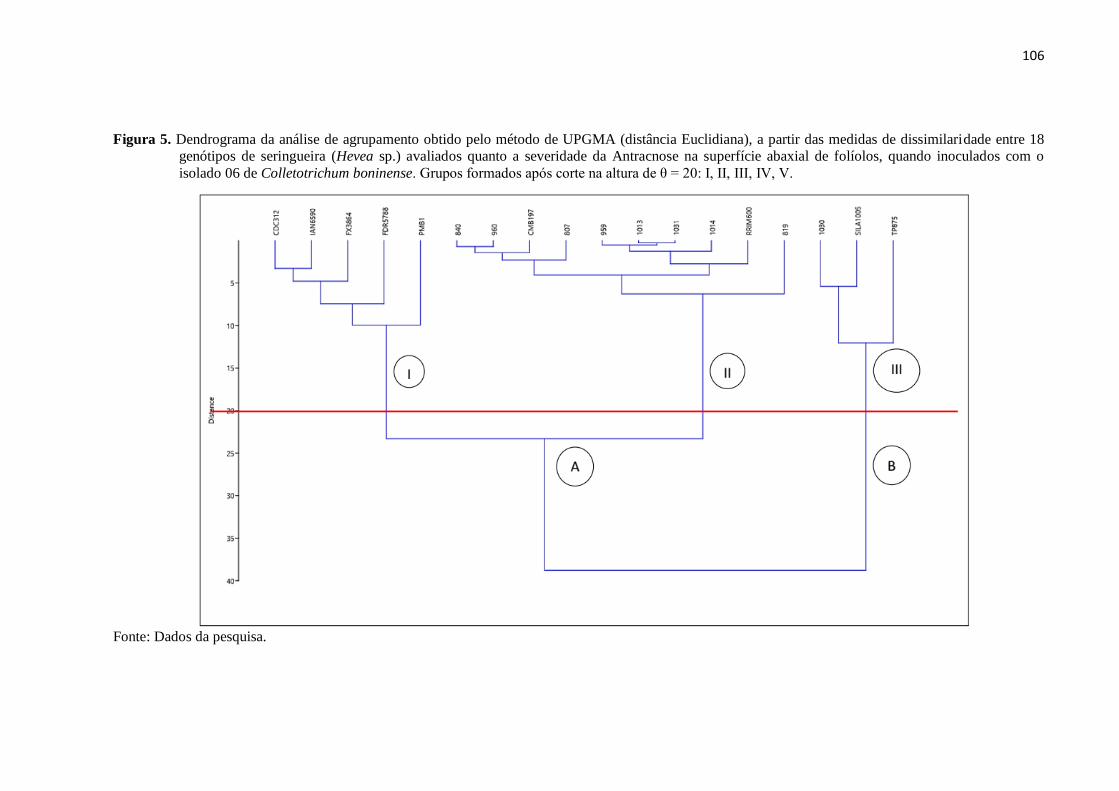
106
Figura 5. Dendrograma da análise de agrupamento obtido pelo método de UPGMA (distância Euclidiana), a partir das medidas de dissimilaridade entre 18
genótipos de seringueira (Hevea sp.) avaliados quanto a severidade da Antracnose na superfície abaxial de folíolos, quando inoculados com o
isolado 06 de Colletotrichum boninense. Grupos formados após corte na altura de θ = 20: I, II, III, IV, V.
Fonte: Dados da pesquisa.

107
6.4 DISCUSSÃO
A metodologia de inoculação com isolados de Colletotrichum spp. utilizando folíolos
destacados foi eficiente na diferenciação dos genótipos testados, quanto à resistência e
suscetibilidade ao patógeno. Até o momento, os trabalhos relacionados à seleção de clones
para resistência à Colletotrichum spp. têm sido feitos por observações visuais no campo em
plantas adultas (GONÇALVES et al., 1999; GONÇALVES, 2002; VIRGENS FILHO, 2017-
dados não publicados). No presente trabalho, os experimentos de seleção de clones para
resistência a esta enfermidade foram realizados através de inoculações do patógeno,
utilizando folíolos destacados no estádio C sob condições controladas, em função de ser um
estádio fenológico intermediário entre o B e D (HALLÉ et al., 1978). No estádio B, os
folíolos apresentam-se com coloração antociânica intensa, e são os mais suscetíveis ao ataque
das doenças foliares (GASPAROTTO et al., 1997), porém nesse período, os folíolos estão
muito frágeis, sendo facilmente degradados, dificultando a avaliação dos sintomas. No estádio
D, os folíolos apresentam-se resistentes à penetração de fungos em geral, dificultando o
aparecimento de sintomas (GASPAROTTO et al., 2012). Os estádios foliares são importantes
para o ciclo das relações patógeno-hospedeiro, pois estes têm correlação direta com as fases
de desenvolvimento do patógeno, suscetibilidade e resistência dos folíolos à infecção
(SAMBUGARO, 2007).
A metodologia de avaliação de clones de seringueira em folíolos destacados, mesmo
quando utilizada em outro patossistema, se mostrou eficiente e consonante com sintomas
encontrados em folíolos presos na planta, independente da idade dos folíolos e do clone. Esta
metodologia foi utilizada para avaliar a resistência da seringueira à Phytophthora capsici
(SANTOS et al., 1993), demonstrando ser um método rápido, reproduzível e não destrutivo,
que se adequa à ensaios em etapas iniciais de seleção, com possibilidade de verificação de
resistência à vários patógenos (TEDFORD et al., 1990; TU, 1986).
As medidas do diâmetro médio das lesões formadas nos genótipos inoculados com os
quatro isolados de Colletotrichum foram realizadas somente a partir do quarto dia após a
inoculação, quando da coalescência de pontos necróticos para a maioria dos genótipos, devido
à possibilidade da reação de hipersensibilidade e consequente formação de pontos necróticos
sem o desenvolvimento de lesão (PASCHOLATI, 1994). No entanto, observações foram feitas
desde 24 horas após a inoculação para a visualização do aparecimento de pontos necróticos nas
folhas inoculadas. Para alguns genótipos isto se deu dois dias após a inoculação (DAI),
corroborando com as informações disponibilizadas na literatura (GASPAROTTO et al., 2012).

108
Utilizou-se neste trabalho a palavra genótipo para designar todo material botânico
testado do Banco de Germoplasma da PMB, por haverem entre eles cruzamentos, clones
comerciais, híbridos e seleções.
Os diâmetros médios das lesões (DML) apresentados nas Tabelas 2 a 5 permitiram
verificar que há variação no comportamento dos genótipos com relação aos diferentes
isolados de Colletotrichum testados. Tal fato é explicado pela grande variabilidade genética
que o gênero Colletotrichum apresenta, conforme dados disponibilizados na literatura (WEIR
et al., 2012; SHARMA et al., 2015) refletindo diretamente no comportamento dos genótipos.
Na tabela 4, observou-se que o tamanho médio das lesões causadas pelo isolado 13,
desde o primeiro dia de avaliação (DML1), foi maior em comparação aos outros três isolados
de Colletotrichum testados, consequentemente refletindo nos valores do DML4, onde foi
possível observar lesões acima de 17,8 mm. O clone Fx 3864, que diferiu estatisticamente dos
demais, sendo considerado o mais resistente para o isolado 13 (C. gloeosporioides),
apresentou lesões maiores em relação aos isolados 14 (C. acutatum) e 06 (C. boninense).
Esses resultados podem ser explicados pela alta variabilidade genética existente dentro do
complexo C. gloeosporioides quando comparado a C. acutatum. Sierra Hayer (2014)
comprovou essa variabilidade quando analisou 79 isolados de C. gloeosporioides e C.
acutatum associados à seringueira, e observou uma grande variabilidade genética entre
isolados de C. gloeosporioides, superando a existente dentro de C. acutatum, como observado
por Freemam (2000). Sierra Hayer (2010) relatou que o complexo C. gloeosporioides, é mais
agressivo quando comparado com as outras espécies de Colletotrichum em seringueira. Gazis
e Chaverri (2010), também comprovaram a alta diversidade genética de C. gloeosporioides
quando comparado com C. boninense.
Para os isolados de C. boninense (06) e C. acutatum (14 e 19) foi possível observar
que o padrão de resistência e suscetibilidade dos genótipos foi evidenciado desde a primeira
avaliação, havendo desde a inexistência de lesões, no DML1, a lesões de 14,72 mm de
diâmetro.
Todos os genótipos foram infectados, embora com níveis de infecção diferenciados o
que indica que o tipo de resistência existente na seringueira em relação à Colletotrichum spp. é
horizontal, podendo ser indicada pela variação de reação dos genótipos (RIBEIRO et al., 1981).
Não foi observada resistência total para Colletotrichum em nenhum dos genótipos de
seringueira avaliados, em contrapartida verificou-se uma variabilidade no progresso da doença,
demonstrando haver mecanismos de defesa que possibilitam o bloqueio ou diminuição do
avanço do patógeno no tecido vegetal. Koop et al. (2016) estudando o patossistema

109
Microcyclus ulei x seringueira, constatou níveis diferenciais da expressão gênica em genótipos
considerados totalmente resistentes, parcialmente resistentes e suscetíveis, bem como
diferenciação em genótipos inoculados e não inoculados com o patógeno. Para os autores,
alguns genes presentes e/ou ativados, de maneira diferencial nestes genótipos, estão envolvidos
na produção de espécies reativas de oxigênio (ERO), sistema de resposta anti-oxidante e
senescência foliar. É possível que no patossistema seringueira x Colletotrichum spp., isto
também ocorra, ou algum outro mecanismo semelhante.
Existem outros mecanismos de defesa envolvidos no processo de resistência das plantas
ao ataque de patógenos. Esses mecanismos podem ser constitutivos ou pré-formados (cutícula
espessada, tecidos lignificados, compostos fenólicos e outros) ou induzidos pela infeção
(proteínas relacionadas à patogênese – PR, fitoalexinas, ligninas, caloses, entre outros)
(GWINN et al., 2010; CHINNAPUN, 2009). Foi verificado por Garcia et al. (1999) que o
maior acúmulo de escopoletina (filtoalexina da seringueira) e de lignina nos sítios de infecção
da doença estavam presentes nos clones mais resistentes ao patógeno. Churngchow e
Rattarasarn (2001) também observaram teores altos de escopoletina em genótipos de
seringueira resistentes à Phytophthora palmivora como também, o aumento da concentração de
escapoletina, proporcional à concentração de esporos no sítio de infecção. Também foi
observado o acúmulo de espécies reativas de oxigênio (ERO) em genótipos resistentes a
Colletotrichum (FIORI, 2011). Esquerré-Tugayé et al. (1992) também sugerem que as
fitoalexinas têm papel fundamental na resistência à Colletotrichum, ao estudarem a resistência
no patossistema C. lindemuthianum x feijão.
Mesmo para os genótipos que não apresentaram lesões nos primeiros dias de avaliação,
após determinado tempo, estas foram observadas. Havendo desta forma, uma suplantação das
defesas do hospedeiro a exemplo da possibilidade de desintoxicação do patógeno frente à
fitoalexinas liberadas pelo hospedeiro, ou inibição de outras moléculas (proteases) liberadas
pelo hospedeiro (GWINN et al., 2010).
Neste estudo, foram observados diferentes níveis de resistência entre os genótipos
testados. Esses resultados eram esperados, principiante pelo fato de serem provenientes de
cruzamentos entre clones interespecíficos, utilizando diferentes materiais como progenitores,
o que indica uma ampla diversidade genética entre eles. Marques et al. (2002) estudaram a
variabilidade genética de clones de seringueira, pertencentes à série SIAL e as variedades
comerciais da série Fx, e observaram a alta variabilidade genética entre os clones da serie
SIAL em relação à série Fx, sendo que os clones da série SIAL apresentaram maiores
distâncias genéticas entre si. Os resultados sugerem que a alta diversidade genética observada

110
entre os clones da série SIAL evidencia que essa ampla base genética pode ser explorada em
programas de melhoramento genético da seringueira. Embora Marques (2010), recomende o
plantio dos clones SIAL 893 e SIAL 1005 por serem clones produtivos, precoces e tolerantes
às principais doenças foliares como Mal das Folhas, no presente estudo esses clones
apresentaram-se como medianamente suscetíveis ao ataque de Colletotrichum. Porém, estes
clones da série SIAL devem continuar sendo explorados em programas de melhoramento
genética da seringueira por apresentarem alta diversidade genética.
Dentre os genótipos avaliados, dez (807, 1031, 819, 1013, 959, 840, 1014, 1030,
960, 1023) são provenientes de um mesmo cruzamento entre um clone oriental altamente
produtivo (RRIM 600) (GONÇALVES; MARQUES, 2008) e um clone que apresenta
resitência ao M. ulei (FDR 5788) (SANDOVAL et al., 2017) (Tabela 1). Mesmo havendo
diferenças no comportamento destes genótipos frente à inoculação de diferentes isolados de
Colletotrichum, estes se encontram dentro do grupo que apresentou os menores diâmentro da
lesão. Segundo Virgens Filho (2017 - dados não publicados) o clone FDR 5788 quando
avaliado em condições de campo pela escala diagramática de densidade foliar, apresentou
resitência ao Colletotrichum spp. Desta forma acredita-se na possibilidade de transferência de
genes envolvidos na resistência a Colletotrichum para suas progênies, na tentativa de conciliar
produtividade com resistência a doenças.
O clone FDR 5788, utilizado como base de resistência a M. ulei em Programas de
Melhoramento Genético (SUAREZ et al., 2015) foi também parental de cruzamentos que
originaram genótipos que apresentaram resistência aos isolados de Colletotrichum, a exemplo
do genótipo 1609 (FDR 5788 x PFB5) e 1666 (FDR 5788 x PMB 1). Originário da Amazônia
Brasileira, o clone PFB 5, mesmo não apresentando um bom potencial produtivo (RIBEIRO,
1988), tem sido utilizado como fonte de resitência ao M. ulei em diversos cruzamentos
(JUNQUEIRA et al., 1988). A característica de resistência a outros patógenos também pode
ser transferida para suas progênies, o que foi evidenciado neste trabalho, nos genótipos 1609,
1645 (RRIM 600 x PFB 5) e 1688 (RRIM600 x PFB 5) para o patossitema Colletotrichum x
seringueira. O clone PMB1 tem sido estudado em campos de clones de grande escala (CCGE)
por apresentar bons potenciais de produção em teste precoce, além de outras características
como vigor, arquitetura da copa, formato do tronco, e uma boa estrutura do sistema laticífero,
no entanto, ao avaliar resistência a Colletotrichum spp. este material apresentou altos índices
de infecção ao patógeno em condições de campo (VIRGENS FILHO, 2017 - dados não
publicados) e também em condições controladas como demonstram os resultados neste
estudo. Apesar do PMB1 apresentar suscetibilidade ao patógeno, quando este é utilizado

111
como progenitor em cruzamentos com outros materiais que possuam genes de resistência a
determinado patógeno, este material poderá gerar progênies com características desejáveis, a
exemplo do resultado obtido pelo genótipo1666.
Outros representantes da série FDR, originários da Guatemala, como o FDR 5240 e
FDR 4575 também foram avaliados e possuem como progenitor comum o clone Harbel 8.
Cubry e colaboradores (2014), ao avaliarem genes candidatos que foram utilizados em estudo
de resistência à M. ulei, identificaram similaridade genética entre clones das series FDR e Fx,
demonstrando haver possibilidade de estarem utilizando recursos genéticos semelhantes para
o combate ao patógeno.
Como representantes de clones amazônicos também se encontram nesse trabalho, o
clone IAN 6590, desenvolvido pelo Instituto Agronômico do Norte (IAN) e integrantes da
série Fx (Fx 2784, Fx 3899, Fx 3864 e Fx 2261), a qual é composta pela maioria de clones
desenvolvidos pela Compahia Ford com base em características agronômicas desejáveis e
resistência moderada a suscetível ao Mal das Folhas (M. ulei) e cancro do painel
(Phytophthora spp.) de acordo com Gonçalves e Marques (2008), mas que vêm se
comportando como altamente suscetíveis a estas enfermidadaes no Sudeste da Bahia. Esses
genótipos quando avaliados em condições de campo apresentaram baixo índice de infecção
em torno de 25%, não diferindo de representantes da série FDR (VIRGENS FILHO, 2017-
Dados não publicados).
O presente trabalho destaca a possibilidade de complementariedade da resistência ao
Mal das Folhas e a Antracnose. É possível que os mecanismos de resistência entre a
seringueira e os patógenos M. ulei e Colletotrichum spp. sejam diferentes do ponto de vista
bioquímico, entretanto o genótipo poderá reunir diferentes populações de genes que estejam
relacionados à resistência a mais de um patógeno. Como evidenciado pelo clone FDR 5788
que se destacou dentre todos os genótipos estudados, por apresentar isoladamente ou como
parental de progênies os melhores resultados para resistência a Colletotrichum.
A metodologia empregada na presente pesquisa é de fácil reprodução, permite
diferenciar genótipos, e também pontua a dificuldade existente em avaliar genótipos de
seringueira em relação aos complexos de espécies de Colletotrichum. Além disso, trata-se de
um trabalho pioneiro que demostra o quanto a variação do complexo do patógeno interfere na
avaliação para resistência à Antracnose.

112
6.5 REFERENCIAS BIBLIOGRÁFICAS
BERGAMIN FILHO, A.; KIMATI, H.; AMORIM, L. Manual de fitopatologia: princípios e
conceitos. 4rd ed. São Paulo: Ed. Agronômica Ceres, 907 p. 1 v. 2011.
CHINNAPUN, D.; TIAN, M.; DAY, B.; CHURNGCHOW, N. Inhibition of a Hevea
brasiliensis protease by a Kazal-like serine protease inhibitor from Phytophthora
palmivora. Physiological and Molecular Plant Pathology. v. 74, Issue 1, P. 27-33. 2009.
CHURNGCHOW, N.; RATTARASARN, M. 2001. Biosynthesis of scopoletin in Hevea
brasiliensis leaves inoculated with Phytophthora palmivora. Journal of Plant Physiology,
Volume 158, Issue 7, 33-8. 2001.
CORLEY, R.V.H. Potential productivity of tropical perennial crops. Experimental
Agriculture 19:217-237. 1983.
CUBRY, P.; RENAUD-PUJADE, V.; GARCIA, D.; ESPEOUT, S.; GUEN, V.L.; GRANET,
F.; SEGUIN, M. Development and characterization of a new set of 164 polymorphic
EST-SSR markers for diversity and breeding studies in rubber tree (Hevea brasiliensis
Müll. Arg.) Plant Breeding. 133.419-426p. 2014.
ESQUERRÉ-TUGAYÉ, M.T.; MAZAU, D.; BARTHE, J.P.; LAFITTE, C.; TOUZÉ, A.
Mechanismo of resistance to Colletotrichum species. In: Control. Wallingford, U.K: CAB
Internacional 1992.p.121-133.
EMBRAPA. 2017. Disponível em: https://www.embrapa.br/busca-de-imagens/-
/midia/746004/seringueira. Acesso em: 12 dez. 2017.
FIORI, M.S. Estresse oxidativo em clones de seringueira sob ataque de antracnose-das-
folhas. Dissertação Mestrado - Faculdade de Ciências Agronômicas da UNESP – Campus de
Botucatu, São Paulo. 97p. 2011.
FREEMAN, S.; MINZ, D.; JURKEVITCH, E.; MAYMON, M.; SHABI, E. Molecular
analyses of Colletotrichum species from almond and other fruits. Phytopathology, St.
Paul, v. 90, n. 6, p. 608-614. 2000.
FURTADO, E.L. Doenças das folhas e do caule da seringueira. In: ALVERENGA,
A.P.;FERREIRA DE SANTANA DO CARMO, C.A: Seringueira. Vicosa, EPAMIG, p. 499-
534. 2008.
FURTADO, E.L.; SILVEIRA, A.R.; GONÇALVES, P.S.; COSTA, J.D.; WINDEL, W.;
SEGNINI JUNIOR, I. Avaliação de cultivares de seringueira quanto ao desfolhamento
causado por Colletotrichum gloeosporioides Penz., no Estado de São Paulo. Fitopatologia
Brasileira, Brasília, DF, vol. 19, p. 339. 1994.
GARCIA, D.; TROISPOUX, V.; GRANGE, N.; RIVANO, F.; D'AUZAC, J. Evaluation of
the resistance of 36 Hevea clones to Microcyclus ulei and relation to their capacity to
accumulate scopoletin and lignins. European Journal of Forest Pathology. 29 (5): 323–338.
1999.

113
GASPAROTTO, L.; ALBUQUERQUE, P. E. P.; D`ANTONA, O. de J. G.; RIBEIRO, I. .A.;
RODRIGUES, F. M.; LIM, T. M. Reabilitacao de seringais de cultivo da Amazonia.
Manaus, EMBRAPA-CNPSD, 27p. 1985.
GASPAROTTO, L.; FERREIRA, F.A.; LIMA, M.I.P.M.; PERREIRA, J.C.R.; SANTOS, A.F
dos. Enfermidades da seringueira no Brasil. Manaus: Embrapa-CPAA, 169p. (EMBRAPA-
CPAA. Circular técnica, 3). 1990.
GASPAROTTO, L.; SANTOS, A. F.dos; PEREIRA, J. C. R.; FERREIRA, F. A. Doenças da
Seringueira no Brasil. Brasília: SPI-EMBRAPA. 168 p. 1997.
GASPAROTTO, L.; FERREIRA, F. Α.; SANTOS, A. F.dos.; PEREIRA, J. C. R.;
FURTADO, E. L. Doenças das Folhas. In: GASPAROTTO, L.; PEREIRA, J.C.R (Ed. Tec.).
Doenças da seringueira no Brasil. 2. ed. rev. e atual. EMBRAPA: Brasília, DF, Brasil,
2012. cap. 3, p. 35-176.
GAZIS, R., CHAVERRI, P. Diversity of fungal endophytes in leaves and stems of wild
rubber trees (Hevea brasiliensis) in Peru. Fungal Ecol. 3, 240– 254. 2010.
GONÇALVES, P.S.; MARQUES, J.R.B. Clones de seringueira: Influencia dos fatores
ambientais na produção e recomendação para o plantio. IN: ALVARENGA, A.P.; do
CARMO, C.A.F. Seringueira. Viçosa, MG. EPAMIG xvi, 894p. 2008.
GONÇALVES, P.S.; MARTINS, A.L.M.; FURTADO, E.L.; SAMBUGARO, R.; OTTATI,
E.L.; ORTOLANI, A.A.; GODOY JÚNIOR, G. Desempenho de clones de seringueira da
série IAC 300 na região do planalto de São Paulo. Pesquisa Agropecuária Brasileira,
Brasília, v. 37, n. 2, p. 131-138. 2002.
GWINM, K.D.; GREENE, S.E.; GREEN, J.F.; TRENTLY,D. Defesa do hospedeiro: Um
enfoque físico e fisiológico. IN: TRIGIANO, R.N.; WINDHAM, M.T.; WINDHAM, A.S.
Fitopatologia: Conceitos e Exercícios de Laboratório. 2. ed., Porto Alegre: Artmed, 576p.
2010.
HALLÉ, F.; OLDEMAN, R. A.; TOMLINSON, P. B. Tropical trees and forest. Berlim,
Springer-Verlag, 441 p. 1978.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA – IBGE. 2017. Disponível
em: < http://www.cidades.ibge.gov.br/>. Acesso em: 29 out. 2017.
JUNQUEIRA, N. T. V. Variabilidade fisiológica de Microcyclus ulei (P. Henn) v. Arx.
1985. 135 f. Tese de Doutorado em Agronomia – Universidade Federal de Viçosa, Viçosa,
1985.
JUNQUEIRA, N.T.V.; CHAVES, G.M.; ZAMBOLIM, L.; ALFENAS,A.C.;
GASPAROTTO, L. Reação de clones de seringueira a vários isolados de Microcyclus ulei.
Fitopatologia Brasileira, Brasília, DF, v.23, n.8, p. 877-893. 1988.
JUNQUEIRA, N. T. V.; PEREIRA, A. V.; PEREIRA, E. B. C.; CARBAJAL, A. C. R.
Eficiência de fungicidas no controle de doenças foliares de seringas adultas e em
formação. Manaus: EMBRAPA-CNPSD, Comunicação Técnico, 57). 1987b.

114
JUNQUEIRA, N.T.V.; CHAVES, G.M; ZAMBOLIN, L; ALFENAS, A; GASPAROTTO, L.
Reação de clones de seringueira a vários isolados de Microcyclus ulei. Pesquisa
Agropecuária Brasileira, n. 23, p. 877-893. 1988.
KOOP, D. M.; RIO, M. ; SABAU, X. ; CARDOSO, S. E. A ; CAZEVIEILLE, C.;
LECLERCQ, J.; GARCIA, D. Expression analysis of ROS producing and scavenging
enzyme-encoding genes in rubber tree infected by Pseudocercospora ulei. Plant
Physiology and Biochemistry (Paris), p. 188-199. 2016.
MARQUES, J.R.B. Melhoramento genético da seringueira para área de ocorrência de
Microcyclus ulei. In: II Encontro Brasileiro de Heveicultura. 2010. Ilhéus-BA. Anais. Ilhéus-
BA, CEPEC/CEPLA, MARS Cacau. 2010.
MARQUES, J.R.B.; FALEIRO, F.G.; ANHERT, D. Diversidade genética entre clones de
seringueira das séries SIAL e Fx com base em marcadores RAPD. Agrotrópica. 14 (3):
159-164. 2002.
MARQUES, J.R.B.; MONTEIRO, W.R.; LOPES, U.V.; VALLE, R.R.M. O cultivo do
cacaueiro em sistemas agroflorestais com a seringueira. In: CIÊNCIA, Tecnologia e
manejo do cacaueiro. Itabuna, BA: Gráfica e Editora Vital, p.272-290. 2007.
MOJEMA, R. Hierarquial grouping methods and stopping rules: an evaluation. The
Computer Journal, 20:359-363. 1977.
NARAYANAN, C.; MYDIN, K.K. Heritability of yield and secondary traits in two
populations of Pará rubber tree (Hevea brasiliensis). Silvae Genetica. 60 (3-4): 132–139.
2011.
PASCHOLATI, S. F. Mecanismos bioquímicos de resistência às doenças. In.: FERNANDES,
J. M.; PRESTES, A. M.; PICININI, E. C. (ed.). Revisão anual de patologia de plantas.
Passo Fundo: Revisão anual de patologia de plantas, 1994, v. 2, cap.1, p. 1-51.
RIBEIRO, I. J. A.; BERGAMIM FILHO, A.; CARVALHO, P. C. T. Avaliação da
resistência horizontal a Hemileia vastatrix Berk et Br. em cultivares de Coffea arabica L.
em condições naturais de epidemia. Summa Phytopathologica, Piracicaba, v. 7, n. 1/2, p.
80-95, abr./jun. 1981.
RIBEIRO, S.I. Avaliação de clones amazônicos de seringueira (Hevea spp.) em Porto
Velho, RO. Porto Velho, RO: EMBRAPA.UEPAE Porto Velho, 15p. EMBRAPA.UEPAE
Porto Velho. (Boletim de Pesquisa, 7) 1988.
RODRIGO, LAKSHMAN.; MUNASINGHE,E. Disponível em:<
http://www.heveabrasil.com/noticias/not0032-Sequestro-de-carbono-na-seringueira-vira-
realidade.pdf . >. Acesso 10 out. 2017. 2017.
SAMBURGO, 2007. Estágios foliares, fenologia da seringueira (Hevea spp.) e interação
com Microcyclus ulei (Mal das Folhas). Disponível em:<
https://www.researchgate.net/publication/35145978_Estagiosfoliares_fenologia_da_seringuei
ra_Hevea_spp_e_interacao_com_Microcyclus_ulei_Mal_das_folhas>. Acesso em: 08 Jan.
2018.

115
SANDOVAL, V.J. C.; SILVA, F.F.; RESENDE, M.D.V.; MACEDO, L.R.; CECON, P.R.
Regressão aleatória Bayesiana para avaliação genética da resistência ao Mal das Folhas
em seringueiras. Revista Ciência Agronômica, Fortaleza, v. 48, n. 1, p. 151-156. 2017.
SANTOS, A.F.dos.; PEREIRA, J.C.R.; FERREIRA, F.A. Doenças da copa da seringueira
causadas por Phytophthora spp. –requeima e queda anormal das folhas. IN: FERREIRA, F.A.
(ed.). Patologia Florestal: principais doenças foliares no Brasil. Viçosa: SIF. P.314-325.
1989.
SANTOS, A. F. dos; MATSUOKA, K.; ALFENAS, A. C.; MAFFIA, L. A. Metodologia
para avaliação de componentes de resistência da seringueira (Hevea spp.) a
Phytophthora capsici. Rev. U.A. Série: Ciências Agrárias, vol. 2, n.2, p.11-20. 1993.
SIERRA HAYER, J.F. Caracterização e controle de Colletotrichum spp. em seringueira
(Hevea brasiliensis). 2010. 68 f. Dissertação (Mestrado em Agronomia/Proteção de Plantas)-
Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Botucatu, 2010.
SIERRA HAYER, J.F. Variabilidade genética de Colletotrichum gloeosporioides e
Colletotrichum acutatum em seringueira (Hevea brasiliensis) no Brasil. 2014. 55 f. Tese
(doutorado) - Universidade Estadual Paulista Júlio de Mesquita Filho-Faculdade de Ciências
Agronômicas. 2014.
SILVEIRA, A.P.; CARDOSO, R.M.G.; FEICHTEMBERGER, E.; OLIVEIRA, D.A..
Patogenicidade de Phytophthora spp. em seringueira (Hevea sp.). Fitopatologia Brasileira.
Brasília, DF. v.11, p.316. 1986.
SHARMA, G.; PINNAKA, A.K.; SHENOY, B.D. Resolving the Colletotrichum siamense
species complex using ApMat marker. Fungal Diversity 71, 247–264. 2015.
STERLING, A.; RODRÍGUEZ, C.; BETANCURT, B.; MAZORRA, A.; DUSSAN, I.;
GALINDO, L.; HERNANDEZ, J.; PLAZA, C.; POLO, F.; GAMBOA, A.; CASTRO, D.;
SHARMA, G.; PINNAKA, A.K.; SHENOY, B.D. Resolving the Colletotrichum siamense
species complex using Ap Mat marker. Fungal Divers. 71, 247–264. 2015.
SUAREZ, Y.Y. J.; MOLINA, J. R.; FURTADO, E. L.2015. Clones de Hevea brasiliensis de
alta productividad caracterizados por resistência a Microcyclus ulei en jardin clonal en
el magdalena médio colombiano. Summa Phytopathologica, Botucatu , v. 41, n. 2, p. 115-
120.
TEDFORD, E.C.; MILLER, T.L.; WIELSEN, M.T. A detached-leaf technique for
detecting resistence to Phytophthora parasistica var. nicotianea in tabacco. Plant Disease.
Vol. 74. P. 313-316. 1990.
TU, J.C. A detachead ttechinique for screening beans (Phaseolus vulgaris L.) in vitro
against anthracnose (Colletotrichum lindemuthiacum ). Canadian Jounal of Plant Science,
vol. 66. P. 805-809. 1986.
VIRGENS FILHO, A. de C. Explotação de seringais. Informe Agropecuário. v. 28, n. 237,
p. 105 - 119, 2007.
WEIR, B.S.; JOHNSTON, P.R.; DAMM, U. The Colletotrichum gloeosporioides species
complex. Studies in Mycology 73, 115–80. 2012.

116
7. CONSIDERAÇÕES FINAIS
Os dados obtidos neste estudo contribuíram para aumentar o conhecimento na
micobiota associada à Hevea brasiliensis mediante a descoberta de um novo táxon
para a ciência e de novos registros fúngicos;
A utilização das duas sequencias ITS1/4 e a sequência parcial do gene GAPDH
permitiram identificar três complexos de espécies de Colletotrichum associados à
Antracnose foliar da seringueira;
Estudos adicionais com um grupo amostral amplo e a inclusão de novas tecnologias de
análise são recomendáveis para maior detalhamento da população fúngica existente;
Foi possível verificar alta variabilidade na virulência entre os isolados de
Colletotrichum, e também no comportamento dos genótipos estudados, quanto aos
isolados testados;
A metodologia utilizada permitiu diferenciar genótipos resistentes e suscetíveis ao
patógeno, além de demostrar que a variabilidade do complexo de espécies de
Colletotrichum interfere na busca da resistência.
Este trabalho proporcionou uma contribuição relevante para a heveicultura brasileira,
por apresentar resultados que poderão ser explorados em programas de melhoramento
genético da seringueira, destacando-se o clone FDR 5788 para recomendação de uso.

117
8. REFERÊNCIAS
ADASKAVEG, J.E.; HARTIN, R.J. Characterization of Colletotrichum acutatum isolates
causing anthracnose of almond and peach in California. Phytopathology, Saint Paul, v.87,
n.9, p.979-987, 1997.
AFANADOR-KAFURI, L.; MINZ, D.; MAYMON, M.; FREEMAN, S. Characterization of
Colletotrichum isolates from tamarillo, passiflora and mango in Colombia and
identification of a unique species from the genus. Phytopathology, Saint Paul, v.93, n.5,
p.579-587, 2003.
ALEXOPOULOS, C.J.; MIMS, C.W.; BLACKWELL, M. 1996. Introductory Micology.
4th edition. New York, John Wiley e Sons, Inc.
ALMEIDA, L.C.C.; COÊLHO, R.S.B. Caracterização da agressividade de isolados de
Colletotrichum de maracujá amarelo com marcadores bioquímico, fisiológico e
molecular. Fitopatologia Brasileira, Brasília, v.32, n.4, p.318-328, 2007.
ALVARENGA, A.de P.; CARMO, C.A.F.S. do. Seringueira. Viçosa: EPAMIG, 2008. 879p.
ALVAREZ, E.; GAÑÁN, L.; TRIVIÑO, A.R.; MEJÍA, J.F.; LLANO, G.A.; GONZÁLEZ, A.
Diversity and pathogenicity of Colletotrichum species isolated from soursop in Colombia. European Journal of Plant Pathology.Vol.139,Issue 2, pp 325–338. 2014.
AVILA-QUEZADA, G.; SILVA-ROJAS, H.V.; TELIZ-ORTIZ, D. First report of the
anamorph of Glomerella acutata causing anthracnose on avocado fruits in Mexico. Plant
Disease, Saint Paul, v.91, p.1200-1200, 2007.
BAILEY, A. J.; JEGER, J. M. Colletotrichum: biology, pathology and control. Oxford:
British Society for Plant Pathology, 1992. 388p.
BARTH, S.; MELCHINGER, A.E.; LÜBBERSTEDT, T. Genetic diversity in Arabidopsis
thaliana L. Heynh. Investigated by cleaved amplified polymorphic sequence (CAPS) and
inter-simple sequence repeat (ISSR) markers. Molecular Ecology, v.11, p.495-505, jul.
2002.
BERGAMIN FILHO, A; AMORIM, L. Doenças de plantas tropicais: epidemiologia e
controle econômico. São Paulo: Ed. Agronômica Ceres, 1996. 299 p.
BERNSTEIN, B.; ZEHR, E.I.; DEAN, R.A.; SHABI, E. Characteristics of Colletotrichum
from peach, apple, pecan, and other hosts. Plant Disease, Saint Paul, v.79, n.5, p.478-482,
1995.
BETTIOL, W.; MAFFIA, L. A.; CASTRO, M. L. M. P. Control biológico de enfermedades
de plantas en Brasil. In: BETTIOL, W.; RIVERA, M. C.; MONDINO, P.; MONTEALEGRE,
J. R.; COLMENAREZ, Y. C. (ed.). Control biológico de enfermedades de plantas en
América Latina y el Caribe. Montevideo: Universidad de la República, 2014. p. 91-138.
BROWN, A.; SOEPENA, H. Pathogenicity of Colletotrichum acutatum and C.
gloeosporioides on leaves of Hevea spp. Mycological Research 98:264-266. 1994.

118
BUENO, C. R. N. C. Identificação e caracterização das espécies de Colletotrichum
causadoras de antracnose em hortaliças e solanaceas. 61 f, 2005. Dissertação (Mestrado
em Agronomia/Fitopatologia)-Escola Superior de Agricultura Luiz de Queiroz, Universidade
de São Paulo, Piracicaba, 2005.
CAI, L.; CHUKEATIROTE, M.; DAYANGA, D.; HYDE, K.D. Endophytic Colletotrichum
from tropical grasses with a new species endophytica. Fungal Divers. 61, 107–115. 2013.
CAI, L.; HYDE, K.D.; TAYLOR, P.W.J.; WEIR, B.S.; WALLER, J. A polyphasic approach
for studying Colletotrichum. Fungal Diversity, Chiang Mai, v.39, p.183-204, 2009.
CANDEIAS, E. L.; LUZ, E. D. M. N.; NIELLA, G. R.; MIRANDA, J. J. C.; SANTOS, C. D.
dos. Microcyclus ulei na Bahia: base para estudo populacional. Agrotrópica (Itabuan), v.
26, n.1.p. 5-16. 2014.
CANNON, P.F.; BRIDGE, P.D.; MONTE, E. Linking the past, present, and future of
Colletotrichum Systematics. In: PRUSKY, D.; FREEMAN, S.; DICKMAN, M.B. (ed.).
Colletotrichum: host specificity, pathology and host-pathogen. Saint Paul: APS Press, 2000.
p. 1-19.
CANNON, P.F.; DAMM, U.; JOHNSTON, P.R.; WEIR, B.S. Colletotrichum– current status
and future directions. Stud. Mycol. 73, 181–213. 2012.
CARPENTER, J. B; STEVENSON, J. A. A secondary leaf spot of the Hevea rubber tree
caused by Glomerella cingulata. Plant Disease Reporter 38: 43-46. (1954)
CHEE, K. H.; DARMONO, T. W.; SANTOS, A. F. Laboratory screening of fungicides
using cellulose film and leaf discs against south american leaf blight pathogen,
Microcyclus ulei. Jounal. Nat. Rubber Resistance. 1 (2): 98-103. 1986.
CERQUEIRA, A. O.; LUZ, E. D. M. N. ; SANTOS, M. V. . Distribuição de Phytophthora
capsici em municípios que cultivam seringueiras no Sul da Bahia. Agrotrópica (Itabuna),
v. 23, p. 65-70, 2011.
CORLEY, R.V.H. Potential productivity of tropical perennial crops. Experimental
Agriculture 19:217-237. 1983.
COSTA, R.B.; GONÇALVES, P. de S.; RÍMOLI, A.O.; ARRUDA, E.J. Melhoramento e
conservação genética aplicados ao desenvolvimento local – o caso da seringueira (Hevea
sp.). Interações - Revista Internacional de Desenvolvimento Local , v.1, p.51- 58, 2001.
DAMM, U.; CANNON, P.F.; WOUDENBERG, J.H.C.; CROUS, P.W. The Colletotrichum
acutatum species complex. Studies Mycology. 73, 37–113. 46. 2012a.
DAMM, U.; CANNON, P.F.; WOUDENBERG, J.H.C.; JOHNSTON, P.R.; WEIR, B.S.;
TAN, Y.P.; SHIVAS, R.G.; CROUS, P.W. The Colletotrichum boninense species complex.
Studies Mycology. 73, 1–36. 2012b.
DEAN, R.; VAN, K.J.; PRETORIUS, Z.A. The top 10 fungal pathogens in molecular plant
pathology. Molecular Plant Pathology 13: 414–430. 2012.

119
DIX, N.J.; WEBSTER, J. Ecologia dos fungos, Chapman & Hall, Londres. 1995.
EMBRAPA, 2017. Disponível em:< https://www.embrapa.br/busca-de-imagens/-
/midia/746004/seringueira >. Acesso: 08 dez. 2017.
ESPERANTE, D. Estatísticas e tendências da borracha natural. APRABOR. Brasília,
2017. Disponivel em: < http://www.agricultura.gov.br/assuntos/camaras-setoriais-
tematicas/documentos/camaras-setoriais/borracha-natural/2017/38a-ro/app-abrabor-38ro-
borracha.pdf>. Acesso em: marco/2018.
EVUEH, G.A.; OGBEBOR, N.O. Uso de Fungos de Phylloplane como Agente de
Biocontrole contra a Doença Colletotrichum em seringueira (Hevea brasiliensis Muell.
Arg.). African Journal of Biotechnology, 15, 2569-2572. 2008.
FARR, D.F.; ROSSMAN, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS,
USDA. Disponivel em : <https://nt.ars-grin.gov/fungaldatabases/ >. Acesso em: 08 out. 2017.
2017.
FERNANDO, T.H.; JAYASINGHE, C.K.; WIJESUNDERA, R.L. Factors affecting spore
production. germination and viability of Colletotrichum acutatum isolates from Hevea
brasiliensis. Mycological Research 104:681-685. 2000.
FITZELL, R.D. Colletotrichum acutatum as a cause of anthracnose of mango in New
South Wales. Plant Disease Reporter, Washington, v.63, p.1067-1070, 1979.
FREEMAN, S.; KATAN, T.; SHABI, E. Characterization of Colletotrichum species
responsible for anthracnose diseases of various fruits. Plant Disease 82:596-605. 1998.
FURTADO, E. L.; SILVEIRA, A. P. Doenças da seringueira em viveiros e jardins clonais e
seu controle. In: MEDRADO, M. J. S. et al. Formação de mudas e plantio de seringueira.
Piracicaba: ESALQ, p. 52-64. 1992.
FURTADO, E. L.; TRINIDADE, D. R. Doenças da seringueira. In: KIMATI, H. et al.
Manual de fitopatologia: doenças das plantas cultivadas. 4. ed. Piracicaba: Agronômica
Ceres, v. 2, p. 559-569. 2005.
FURTADO, E.L.; SILVEIRA, A.R.; GONÇALVES, P.S.; COSTA, J.D.; WINDEL, W.;
SEGNINI JUNIOR, I. Avaliação de cultivares de seringueira quanto ao desfolhamento
causado por Colletotrichum gloeosporioides Penz., no Estado de São Paulo. Fitopatologia
Brasileira, Brasília, DF, vol. 19, p. 339. 1994.
GARCIA, D.; MATTOS, C. R.R. ; GONÇALVES, P.S.; GUEN, V. Selection of rubber
clones for resistance to South American Leaf Blight and latex yield in the germplasm of
the Michelin Plantation of Bahia (Brazil). Journal of Rubber Research, Malaisia, v. 7, n.3,
p. 188-198, 2004.
GASPAROTTO, L.; SANTOS, A.F.dos.; PEREIRA, J.C.R.; FERREIRA, F.A. Doenças da
seringueira no Brasil. Brasilia, DF: EMBRAPA, SPI; EMBRAPA, CPAA, 1997. 168 p.

120
GASPAROTTO, L.; ALBUQUERQUE, P.E.P.; D’ANTONA, O de J.G.; RIBEIRO, I.A.;
RODRIGUES, F.M.; LIM, T.M. Reabilitação de seringais de cultivo da Amazônia.
Manaus, EMBRAPA-CNPSD, 1985. 27p.
GASPAROTTO, L.; FERREIRA, F. Α.; LIMA, Μ. I. P. M.; PEREIRA, J. C. R.; SANTOS,
A. F.dos. Enfermidades da Seringueira no Brasil. Manaus: EMBRAPA-CPAA, 169 p.
(EMBRAPA CPAA, Circular Técnica, 3). 1990.
GASPAROTTO, L.; FERREIRA, F. Α.; SANTOS, A. F.dos.; PEREIRA, J. C. R.;
FURTADO, E. L. Doenças das Folhas. In: GASPAROTTO, L.; PEREIRA, J.C.R (Ed. Tec.).
Doenças da seringueira no Brasil. 2. ed. rev. e atual. EMBRAPA: Brasília, DF, Brasil,
2012. cap. 3, p. 35-176.
GASPAROTTO, L.; PEREIRA, J.C.R.; FURTADO, E.L. Seringueira. In: GASPAROTTO,
L.; PEREIRA, J.C.R (Ed. Tec.). Doenças da seringueira no Brasil. 2. ed. rev. e atual.
EMBRAPA: Brasília, DF, Brasil, 2012. cap.1, p. 15-26.
GAZIS, R.; CHAVERRI, P. Diversity of fungal endophytes in leaves and stems of wild
rubber trees (Hevea brasiliensis) in Peru. Fungal Ecology 3: 240-254. 2010.
GAZIS, R.; REHNER, S.; CHAVERRI, P. Species delimitation in fungal endophyte
diversity studies and its implications in ecological and biogeographic inferences. Molecular Ecolology 20:3001-3013. 2011.
GEPTS, P. The use of molecular and biochemical markers in crop evolution studies.
Evolution Biology., 27: 51-94. 1993.
GONÇALVES, P.S.; ORTOLANI, A.A.; CARDOSO, M. Melhoramento genético da
seringueira: Uma revisão. Documentos IAC. Campinas, SP. 1997. 55p.
GONÇALVES, P.S.; CARDOSO, M.; ORTOLANI, A.A. Origem, variabilidade e
domesticação da Hevea: uma revisão. Pesquisa Agropecuária Brasileira. Brasília, 25(2):135-
156. 1990.
GONÇALVES. P.S.; MARQUES, J.R.B. Melhoramento genético da seringueira: Passado,
presente e futuro. IN: ALVARENGA, A.P.; CARMO, C.A.F.F.do (Coord.). Seringueira. 2º
Edição. Viçosa, MG. p. 247-335. 2104.
GONÇALVES, P.S.; MARTINS, A.L.M.; FURTADO, E.L.; SAMBUGARO, R.; OTTATI,
E.L.; ORTOLANI, A.A; GODOY JÚNIOR, G. Desempenho de clones de seringueira da
série IAC 300 na região do planalto de São Paulo. Pesquisa Agropecuária Brasileira,
Brasília, v. 37, n. 2, p. 131-138. 2002.
GONÇALVES, P.S.; MARTINS, A.L.M.; BORTOLETTO, N.; TANZINI, M.R. Estimates
of genetic parameters and correlations of juvenile characters basead on open pollinated
progenies of Hevea. Revista Brasileira de Genética, 19(1): 105-111. 1996.
GUOZHONG, L.; CANNON, P.F.; REID, A.; SIMMONS, C.M. Diversity and molecular
relationships of Colletotrichum isolates from the Iwokrama Foret. Mycological Research,
Cambridge, v. 108, p. 53-63, 2004.

121
GUYOT, J.; NTAWANGA, E.; NDOUTOUME, E.; OTSAGHE, A.; ENJALRIC, F.;
ASSOUMOU, H. Effect of controlling Colletotrichum leaf fall of rubber tree on epidemic
development and rubber production. Crop Protection 20:581-590. 2001.
GUYOT, J.; NTAWANGA, E.; PINARD, F. Some epidemiological investigations on
Colletotrichum leaf disease on rubber tree. Crop Protection 24:65-77. 66. 2005.
HALLÉ, F.; OLDEMAN, R.A.A.; TOMLINSON, P.B. Tropical trees and forests. Berlin,
Springer, 1978. 441p.
HAWKSWORTH, D.L. The fungal dimension of biodiversity: magnitude, significance,
and conservation. Mycology Research 95:641-655. 1991.
HENZ, G.P.; BOITEUX, L.S.; LOPES, C.A. Outbreak of strawberry anthracnose caused
by Colletotrichum acutatum in central Brazil. Plant Disease 76:212. 1992.
HIBBETT, D.S.; BINDER, M.; BISCHOFF, J.F.; BLACKWELL, M.; CANNON, P.;
ERIKSSON, O.; HUHNDORF, S.; JAMES, T.; KIRK, P.M.; LUCKING, R.; THORSTEN,
H.; LUTZONI, F.; MATHENY, P.; MCLAUGHLIN, D.; POWELL,M.; REDHEAD, S.;
SCHOCH, C.L.; SPATAFORA, J.; STALPERS, J.; VILGALYS, R.; AIME, M.; APTROOT,
A.; BAUER, R.; BEGEROW, D.; BENNY, G.; CASTLEBURY, L.; CROUS, P.; DAI, Y-W.;
GAMS,W.; GEISER, D.; GRIFFITH, G.; GUEIDAN, C.; HAWKSWORTH, D.;
HESTMARK, G.; HOSAKA, K.; HUMBER, R.; HYDE, K.; IRONSIDE, J.; KO LJALG, U.;
KURTZMAN, C.; LARSSON, K.H.; LICHTWARDT, R.; LONGCORE, J.; DLIKOWSKA,
M.; MILLER, A.; MONCALVO, J-M.; MOZLEYSTANDRIDGE, S.; OBERWINKLER, F.;
PARMASTO, E.; REEB, V.; ROGERS, J.; ROUX, C.; RYVARDEN, L.; SAMPAIO, J.P.;
SCHUßLER, A.; SUGIYAMA, J.; THORN, R.; TIBELL, L.; UNTEREINER, W.;
WALKER, C.; WANG, Z.; WEIR, A.; WEISS,M. WHITE, M.; WINKA, K.; YAO, Y-J.;
ZHANG, N. A Higher-level Phylogenetic Classification of the Fungi. Mycological
Research 111: 509-547. 2007.
HO, C.Y. Rubber, Hevea brasiliensis, In: FAO, Breeding for Durable Resistance in
Perennial Crops. FAO Plant Production and Protection Paper 70, p. 85 - 114. 1986.
HOLLIDAY, P. South American leaf blight (Microcyclus ulei) of Hevea brasilienses.
C.M.I. Phytopatological Papers. Nº 12, p. 1-31. 1970.
IAC, 2017. Disponível em: < http://www.iac.sp.gov.br/areasdepesquisa/seringueira/>.
Acesso: 08 dez. 2017.
IBGE, 2017. Disponível em:
<https://ww2.ibge.gov.br/home/estatistica/pesquisas/pesquisa_resultados.php?id_pesquisa=44
>. Acesso em: 11 nov. 2017.
JAYASINGHE, C.K.; FERNANDO, T. H. P. S. Growth at different temperatures and on
fungicide amended media: Two characteristics to distinguish Colletotrichum species
pathogenic to rubber. Mycopathologia 143:93-95. 1998.

122
JAYASINGHE C. K.; FERNANDO, T. H. P. S.; PRIYANKA, U. M. S. Colletotrichum
acutatum is the main cause of Colletotrichum leaf disease of rubber in Sri Lanka. Mycopathologia 137:53-56. 1997.
JUNQUEIRA, N. T. V.; PEREIRA, A. V.; PEREIRA, E. B. C.; CARBAJAL, A. C. R.
Eficiência de fungicidas no controle de doenças foliares de seringas adultas e em
formação. Manaus: EMBRAPA-CNPSD, Comunicação Técnico, 57. 1987b.
KALIL FILHO, A. N.; JUNQUEIRA, Ν. Τ. V. Bases e procedimentos para o programa de
melhoramento de seringueira no CNPSD-Manaus, AM. Manaus, EMBRAPA-CNPSD, 13
p. (EMBRAPA-CNPSD. Documentos, 8). 1989.
KENDRICK, B. The Fifth Kingdom.3rd edition.Newburyport, Focus Publishing. 2000.
KIRK, P.M., CANNON, P.F., MINTER, D.W., STALPERS, J.A. Dictionary of the
Fungi.10th edition. Wallingford, CAB International. 2008.
LAKSHMAN, R.; MUNASINGHE, E. Disponível em
http://www.heveabrasil.com/noticias/not0032-Sequestro-de-carbono-na-seringueira-vira-
realidade.pdf . Acesso em: 10 out. .2017. 2017.
LENNÉ, J. M.; THOMAS, D.; ANDRADE, R. P. de.; VARGAS, A. Anthracnose of
Stylosanthes capitata: implications for future disease evaluations of indigenous tropical
pasture legumes. Phytopathology, St. Paul, MN, v.74, n.9, p.1070-1073, 1984.
MAKI, C. S. Diversidade e potencial biotecnológico de fungos endofíticos de cacau
(Theobroma cacao L.). Tese de doutorado – Escola Superior de Agricultura ―Luiz de
Queiroz‖. Universidade de São Paulo, Piracicaba, 2006.
MARQUES, V.V. Patogenicidade e variabilidade genética de Colletotrichum spp. em
cafeeiro (Coffea arabica L. ) 2008. p.108. Tese (Doutorado em Agronomia) Universidade
Estadual de Londrina, Londrina. 2008.
MAHMODI, F.; KADIR, J. B.; PUTEH, A.; POURDAD, S. S.; NASEHI, A.; SOLEIMANI,
N.Genetic Diversity and Differentiation of Colletotrichum spp. Isolates Associated with
Leguminosae Using Multigene Loci, RAPD and ISSR. Plant Pathology Journal. 30(1): 10-
24, 2014. doi: 10.5423/PPJ.OA.05.2013.0054.
MAIA, L.C. Coleções de fungos nos herbários brasileiros: estudo preliminar. In: PEIXOTO,
A.L., (Org.). Coleções Biológicas de apoio ao inventário, uso sustentável e conservação da
Biodiversidade. Instituto de Pesquisa Jardim Botânico do Rio de Janeiro, Rio de Janeiro,
p.21-40, 2003 Disponível em:
<http://www.iac.sp.gov.br/areasdepesquisa/seringueira/importancia.php>. Acesso:15 abr.
2017.
MATTHEWS, D.; MCNICOLL, J.; HARDING, K.; MILLAM, S. 5'-anchored simple-
sequence repeat primers are useful for analysing potato somatic hybrids. Plant Cell
Reports, v.19, p.210-212, 1999.

123
MENDES, M.A.S.; SILVA, V. L. DA; DIANESE, J. C.; FERREIRA, M. A. S.; V.; SANTOS
, C. E. N. D. O. S.; GOMES NETO E.; URBEN A. F.; CASTRO, C. Fungos em plantas no
Brasil. Brasília: EmbrapaSPI/Embrapa-Cenargen, 569p. 1998.
MENEZES, M. Aspectos biológicos e taxonômicos de espécies do gênero Colletotrichum.
Fitopatologia Brasileira 27:23-24. 2002.
MENEZES, M. Aspectos biológicos e taxonômicos de espécies do gênero Colletotrichum.
Anais da Academia Pernambucana de Ciência Agronômica v. 3:170-179. 2006.
NGHIA, N.A; KADIR, J; SUNDERASAN, E; ABDULLAH, M.P; MALIK, A; NAPIS, S.
Morphological and Inter Simple Sequence Repeat (ISSR) Markers Analyses of
Corynespora cassiicola Isolates from Rubber Plantations in Malaysia. Mycopathologia,
v.166, edição 4, 189-201pp. 2008.
MILLER, J. W. Diferencial clones of Hevea for identifying races of Dothidela ulei. Plant.
Dis. Rept. 30: 187-90. 1966.
MILLS, P. R.; HODSON, A.; BROWN, A. E. Molecular differentiation of Colletotrichum
gloeosporioides isolates infecting tropical crops. In: BAILEY, J. A.; JEGER, M. J. (eds.).
Colletotrichum: Biology, Pathology and Control. Wallingford, United Kingdom: CAB
international, p. 269-288. 1992a.
MORAES. V.H. de F. Fisiologia - Parte 1. Belém : FCAp, 1980. 51 p. Trabalho apresentado
no 7. Curso de Especialização em Heveicultura. Belém. PA. 1980.
MORENO, H.L.A. Diversidade de espécies de Colletotrichum endofíticas da seringueira.
Dissertação de mestrado. Universidade Federal de Viçosa. Departamento de Microbiologia.
Programa de Pó-graduação em Microbiologia Agrícola. 2016.
NAGAOKA, T.; OGIHARA Y. Applicability of inter-simple sequence repeat
polymorphisms in wheat for use as DNA markers in comparison to RFLP and RAPD
markers. Theoretical and Applied Genetics, New York, v. 94, p. 597-602, 1997.
NARAYANAN, C.; MYDIN, K.K. Heritability of yield and secondary traits in two
populations of Pará rubber tree (Hevea brasiliensis). Silva e Genética. 60 (3-4): 132–139.
2011.
NGUYEN, P. T. H.; PETTERSSON, O. V.; OLSSON, P.; LILJEROTH, E. Identification of
Colletotrichum species associated with anthracnose disease of coffee in Vietnam. European Journal of Plant Pathology, Dordrecht, v. 127, p. 73- 87, 2010.
OGBEBOR, N.; ADEKUNLE, A.T. Inibição da Germinação Conidial e Crescimento
Mycelial de Corynespora cassiicola (Berk e Curt) de Borracha (Hevea Brasiliensis Muell.
Arg.) Usando Extratos de Algumas Plantas. African Journal of Biotechnology, 4, 996-
1000. 2005.
PALM, M. E.; CHAPELA, I.H. Mycology in sustainable development: expanding concepts,
vanishing borders. Parkaway Publishers Inc., Boone, N.C. 1998, 400p.

124
PARREIRA, D. F.; ZAMBOLIM, L.; GOMES, E, A.; COSTA, R, V., SILVA, D, D.;
PAULA LANA, U, G.; NEVES, W, S.; FIGUEIREDO, J, E, F.; COTA, L, V. Diversidade
genética estimada através de marcadores ISSR de Colletotrichum graminicola no Brasil.
Revista Brasileira de Milho e Sorgo, v.15, n.2, p. 186-194, 2016.
PETCH, T. The Diseases and Pests of the Rubber Tree. MacMillan. London. 278 p. 1921.
PINHEIRO, E.; LIBONATI, V.F. O emprego da Hevea pauciflora M.A. como fonte
genética de resistência ao Mal das Folhas. Polímeros, Rio de Janeiro, (1): 31-41,1971.
PLOETZ, R.C. Mango diseases caused by fungi: anthracnose. In: Ploetz, R.C.; Zentmeyer,
G.A.; Nishijima, N.T.; Rohrbasch, K.G.; Ohr, H.D. (Ed.). Compendium of Tropical Fruit
Diseases. Saint Paul: APS Press, 1994. p.35-36.
RAMPERSAD, S. N. Genetic structure of Colletotrichum gloeosporioides sensu lato
isolates infecting papaya inferred by multilocus ISSR markers. Phytopathology 103:182-
189, 2013.
RATANACHERDCHAI, K.; WANG, H.; LIN, F.; SOYTONG, K. ISSR para comparação
do potencial de inoculação cruzada de Colletotrichum capsici causando antracnose de
pimenta. Microbiology Research, 4 (1): 76-83. 2010.
RIVANO, F.; MATTOS, C.R.R.; CARDOSO, S.E.A.; MARTINEZ, M.; CEVALLOS, V.;
LE GUEN, V.; GARCÍA, D. Breeding Hevea brasiliensis for yield, growth and SALB
resistance for high disease environments. Industrial Crops and Products, V 44, p. 659-670,
2013.
RIVANO, F.; NICOLAS, D.; CHEVAUGEON, J. Resistance de Hevea a Ia maladie sud -
americaine des levilies. Caoutchouc et Plastiques, n. 690, p189- 206, 1989.
ROBERTS, P.D.; PERNEZNY, K. L.; KUCHAREK, T. Anthracnose on pepper in Florida.
University of Florida, PP-178, 2012.
ROCHA, A.; GARCIA, D.; UETANABARO, A.P.T.; CARNEIRO, R.T.; ARAÚJO, I.S.;
MATTOS, C.R.R.; GÓES-NETO, A. Foliar endophytic fungi from Hevea brasiliensis and
their antagonism on Microcyclus ulei. Fungal Diversity 47:75-84. 2011.
RUBINI, M. R. Microbiota endofítica de cacaueiro (Theobroma cacao L.) e o controle
biológico de Crinipellis perniciosa. Dissertação de Mestrado – Universidade de Caxias do
Sul. São Paulo, 2003. 100 p.
SAHA. T.; KUMAR, A.; RAVINDRAN, M.; JACOB, C.K.; ROY, B.; NAZEER, M.A.
Identification of Colletotrichum acutatum from rubber using random amplified
polymorphic DNAs and ribosomal DNA polymorphisms. Mycological Research 106:215-
221. 2002.
SARMIENTO, S.S. Identificação de espécies de Colletotrichum associadas à antracnose
foliar da seringueira. 2013. 72 f. Dissertação (Mestrado em Ciências) - Programa de Pós-
graduação em Fitopatologia. Universidade Federal de Viçosa, Viçosa (MG). 2013.

125
SERRA, I. M. R. de S.; SILVA, G. S. da. Caracterização Morfofisiológica de Isolados de
Colletotrichum gloeosporioides Agentes de Antracnose em Frutíferas no Maranhão. Summa Phytopathologica, Botucatu, v. 30, n. 4, p. 475-480. 2004.
SHARMA, P. N.; KATOCH, A. Population structure of Colletotrichum gloeosporioides
associated with capsicum anthracnose in North Western Himalayas and evaluation for
disease resistance. Journal of Mycology and Plant Pathology Vol. 44 No. 4 pp. 412-416,
2014.
SIERRA HAYER, J.F. Caracterização e controle de Colletotrichum spp. em seringueira
(Hevea brasiliensis). 2010. 68 f. Tese (Doutorado em Agronomia/Proteção de Plantas)-
Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Botucatu, 2010.
SILVEIRA, A. P.; CARDOSO, R. M. G. Ocorrência de Colletotrichum gloeosporioides em
Seringueira (Hevea brasiliensis) no Estado de São Paulo. Summa Phytopathologica,
Piracicaba, v. 13, n. 1/2:19, 1987.
SILVEIRA, A. S., STRACIERI, J., PEREIRA, F. D., SOUZA, A., GOES. A. Caracterização
molecular de isolados de Colletotrichum spp. associados a Podridão floral dos citros. Revista Brasileira de Fruticultura. 2016, vol.38, n.1, pp.64-71. ISSN 0100-2945.
SILVEIRA, A. P.; FURTADO, E. L.; LOPES, M. E. B. M. Antracnose: nova doença do
painel de sangria da seringueira. Summa Phytopathologica, Piracicaba, v. 18, n. 3/4, p. 195-
200, 1992.
SILVEIRA, A. L. Caracterização molecular de Colletotrichum acutatum e C.
gloeosporioides associados a citros, por marcadores ISSR. Jaboticabal, X, 39 p.2015.
SIMMONDS, N. Breeding horizontal resistance to south American leaf blight of rubber.
Journal Natural Rubber Research, 5(2), p. 102-113. 1990.
SMITH, B. J.; BLACK, L. L. Morphological, cultural, and pathogenic variation among
Colletotrichum species isolated from strawberry. Plant Disease, v. 74, p. 69-76, 1990.
SOUSA, E.S.; SILVA, J.R.A.; ASSUNÇÃO, I.P.; MELO, P.M.; FEIJÓ, F.M.; MATOS, K.S.;
LIMA, G.S.A.; ANDRADE, G.S.; BESERRA JR, J.E.A. Colletotrichum species causing
anthracnose on lima bean in Brazil. Tropical Plant Pathology. Vol. 43, Issue 1, pp 78–84.
2018.
SREENIVASAPRASAD, S., MILLS, P. R.; MEEHAN, B. M.; BROWN, A. E. Phylogeny
and systematics of 18 Colletotrichum species based on ribosomal DNA spacer sequences. Genome 39: 498-512. 1996.
STERLING,A.; RODRÍGUEZ, C.; BETANCURT, B.; MAZORRA, A.; DUSSAN, I.;
GALINDO, L.; HERNANDEZ, J.; PLAZA, C. P.F.; GAMBOA, A. C. D.; MARTÍNEZ, O.
Capitulo 4. Evaluación del desempeño y comportamiento fitosanitario de genotipos élites de
H. brasiliensis de origen franco en Campo Clonal a Pequeña Escala CCPE. In: Sterling A,
Rodriguez C (eds). Nuevos clones de caucho natural para la Amazonia colombiana:
énfasis en la resistencia al mal suramericano de las hojas (Microcyclus ulei). Instituto
Amazónico de Investigaciones Científicas – SINCHI. pp. 85-138. 2011.

126
SUTTON, B. C. The genus Glomerella and its anamorph Colletotrichum. In: BAILEY, J. A.;
JEGER, M. J. (Eds.). Colletotrichum: biology, pathology and control. Wallingford: CAB
international, 1992. p. 1-26.
THAMBUGALA, T.A.D.P.; DESHAPPRIYA ,N. The role of Colletotrichum species on the
Colletotrichum leaf disease of Hevea brasiliensis a preliminary study. Journal of the
National Science Foundation Sri Lanka 37:135-138. 2009.
TOZZE JUNIOR, H. J. Caracterização e identificação de espécies Colletotrichum
associadas à antracnose do pimentão (Capsicum annuum) no Brasil. 2007. 81 f.
Dissertação (Mestrado em Agronomia) – Escola Superior de Agricultura ―Luiz de Queiroz‖,
Universidade de São Paulo, Piracicaba, 2007.
TOZZE JÚNIOR, H. J.; MELLO, B. A.; MASSOLA-JÚNIOR, N. S. Caracterização
morfológica e fisiológica de isolados de Colletotrichum sp. causadores de antracnose em
solanáceas. Summa Phytopathologica, Botucatu, v. 32, n. 1, p. 77-79, 2006.
TOZZE JÚNIOR, H.J.; FISCHER, I.H.; CÂMARA, M.P.S.; MASSOLA JÚNIOR, N.S. First
report of Colletotrichum boninense infecting yellow passion fruit (Passiflora edulis f.
flavicarpa) in Brazil. Australasian Plant Disease Notes, Orange, v.5, p.70-72, 2010.
TRINDADE, D.R.; FURTADO, E.L. Doenças da seringueira [ Hevea brasilensis (Willd.
ExAdr. De Juss.) Müell. Arg.]. Em: Manual de Fitopatologia (Kimati, H., Amorin, L.,
Bergamin Filho, A., Camargo, LEA e Rezende, JAM, eds.). 2º Vol. Agronômica Ceres Ltda.,
São Paulo, 628-641. 1997.
VALVERDE, S.R. DA SILVA MIRANDA, M.A.;FONSECA FALEIRO, P.H. Aspectos
econômicos, mercadológicos, e gerenciais da heveicultura. In: Seringueira. EPAMIG Zona
da Mata, Viçosa, MG. 2014.
VIÉGAS, A.P. Índice de fungos da América do Sul. Campinas, 1961. 921 .pp
VIRGENS FILHO, A. de C. Explotação de seringais. Informe Agropecuário, v. 28, n. 237,
p. 105 - 119, 2007.
VIRGENS FILHO, A. de C. Organização e exploração do seringal. In: ALVARENGA,
A.P.de.; CARMO, C.A.F.S.do (coord.). Seringueira. 2º ed. rev. e atual. Viçosa, MG:
EPAMIG, 2014.
WEIR, B.S.; JOHNSTON, P.R.; DAMM, U. The Colletotrichum gloeosporioides species
complex. Studies Mycology. 73, 115–180. 2012.
WILLIAMS, J.G.; KUBELIK, A.R.; LIVAK, K.J.; RAFALSKI JA, T. S.V. DNA
polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic
Acids Research 18: 6531-6535. 1990.

127
APÊNDICE A - OCORRÊNCIA DE OÍDIO EM MUDAS DE SERINGUEIRA NO
ESTADO DA BAHIA, BRASIL

128
9. OCORRÊNCIA DE OÍDIO EM MUDAS DE SERINGUEIRA NO ESTADO DA
BAHIA, BRASIL
Nota científica submetida à revista Agrotrópica
Tacila Ribeiro Santos1, José Luiz Bezerra
1, Adonias de Castro Virgens Filho
2, Lívia Fernanda
Lavrador Toniasso3
¹Universidade Estadual de Santa Cruz (UESC) - Programa de Pós-Graduação em Produção
Vegetal – PPGPV. Rod. Jorge Amado Km 16, 45662-900, Ilhéus, Bahia, Brasil;
²CEPLAC/CEPEC - Seção de Diversificação, km 22, Rod. Ilhéus/Itabuna. Caixa postal 07,
45600-970, Itabuna, Bahia, Brasil; 3Plantações Michelin da Bahia Ltda – Departamento de
Pesquisa e Desenvolvimento (P&D) Rod. Ituberá-Camamu, Km 05, Igrapiúna, Bahia, Brasil.
[email protected]; [email protected]
RESUMO
Oidium heveae é relatado infectando mudas de seringueira coletadas nas Plantações Michelin
da Bahia, município de Igrapiúna, Bahia. Este é o primeiro relato do patógeno sobre folhas de
Hevea brasiliensis no Estado da Bahia.
Palavras-chave: Oidium heveae, Hevea brasiliensis, parasita biotrófico obrigatório.
ABSTRACT
Oidium heveae is reported infecting rubber tree seedlings collected at Michelin Plantations of
Bahia, Igrapiúna, Bahia. This is the first report of the pathogen on leaves of Hevea
brasiliensis in the State of Bahia.
Key words: Oidium heveae, Hevea brasiliensis, obligate biotrophic parasite.
Oídio é uma doença foliar de grande importância para a cultura da seringueira (Hevea
brasiliensis Müll.Arg.), a qual é causada por Oidium hevea Steinm., parasita biotrófico
obrigatório, reportado principalmente em países asiáticos como a Malásia, Sri Lanka e
Indonésia (BURCHILL, 1978). No Brasil, a doença tem sido relatada nos estados de São
Paulo, e também no Espírito Santo (FURTADO; SILVEIRA, 1993).

129
Os folíolos infectados com o patógeno perdem o brilho natural, apresentando um
micélio esbranquiçado nas faces abaxial e adaxial, podendo ocorrer enrugamento e posterior
queda dos mesmos. Em condições favoráveis, o fungo pode esporular e formar lesões de
coloração marrom avermelhada (GASPAROTTO et al., 2012), podendo ser observado em
diversos órgãos vegetais como meristemas, ramos jovens, flores, frutos em formação (AUER,
2001).
Em outubro de 2017, na casa telada das Plantações Michelin da Bahia Ltda, município
de Igrapiúna, Bahia, foram coletadas amostras de folhas de seringueira apresentando sintomas
e sinais de oídio.
No laboratório de Diversidade de Fungos do Centro de Pesquisas do Cacau
(Ceplac/Cepec), o material foi examinado ao microscópio estereoscópico MOTIC SMZ -168
para observação topográfica das colônias. A caracterização morfológica foi feita utilizando-se
um microscópio LEICA DM500 acoplado a uma câmara digital SONY Cyber-shot DSC-
WX30. As mensurações das estruturas foram efetuadas por meio de micrômetro ocular
devidamente calibrado. Para observação do micélio íntegro, utilizou-se a técnica de Callan e
Carris (2004), substituindo o acetato de celulose por esmalte de unha incolor de secagem
rápida (Risqué Technology), seguindo-se de montagem em PVLG (álcool polivinílico + ácido
lático + glicerol) (HOSAGOUDAR; RIJU, 2013).
O fungo estudado possui micélio esbranquiçado com hifas ramificadas, septadas, de
parede fina e conidióforos retos produzindo cadeias basípetas de dois a sete conídios.
Conídios hialinos, elípticos, lisos, medindo de 23-42 x 16-20 µm (Figura 1). As características
morfológicas descritas correspondem às da espécie Oidium heveae, cujo morfo sexual é
desconhecido.
Material examinado: BRASIL. BAHIA: Igrapiúna, Plantações Michelin da Bahia, sobre
folhas de Hevea brasiliensis 24.10.2017, T.R. Santos e J.L. Bezerra (CEPEC 2497).
Comentários: Apesar da doença ser conhecida no Brasil, não havia, até o momento, registro
oficial de O. heveae no estado da Bahia.

130
FIGURA 1. Oidium heveae. A. Muda de seringueira com sintomas da doença. B. Folíolo com
micélio de oídio. C, D. Conidióforo e conídios. E. Conídios em cadeia. Barra = 37 µm.
REFERÊNCIAS BIBLIOGRÁFICAS
AUER, C. G. 2001. Oídios de espécies florestais. In: STADNIK, M. J.; RIVERA, M.C.
(Ed.). Oídios. Jaguariúna: Embrapa Meio Ambiente, 484p.
CALLAN, B.E.; CARRIS, L.M. 2014. Fungi on living plant substrata, including fruits. In:
MUELLER, G.M.; BILLS, G.F.; FOSTER, M.S (eds), Biodiversity of Fungi–Inventory and
Monitoring Methods. California: Elsevier, 777p.
FURTADO, E.L.; SILVEIRA, A.P. 1993. Nova ocorrência de oídio da seringueira no Estado
de São Paulo. São Paulo: Instituto Biológico, (Comunicado Técnico, 3), 3p.
GASPAROTTO, L.; FERREIRA, F. Α.; SANTOS, A. F.dos.; PEREIRA, J. C. R.;
FURTADO, E. L. Doenças das Folhas. In: GASPAROTTO, L.; PEREIRA, J.C.R (Ed. Tec.).
A B
C D E

131
Doenças da seringueira no Brasil. 2. ed. rev. e atual. EMBRAPA: Brasília, DF, Brasil, 2012.
cap. 3, p. 35-176.
HOSAGOUDAR, V.B.; RIJU, M.C. 2013. Foliicolous fungi of Silent Valley National Park. .
Journal of Threatened Taxa (India) 5: 3701–3788