universidade estadual de santa cruz programa...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DE SANTA CRUZ PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUÇÃO VEGETAL
FRANCINNE RIBEIRO ALMEIDA
CULTIVO DE Jatropha curcas L. EM CASA DE VEGETAÇÃO PARA ENSAIO DE DEFICIÊNCIA HÍDRICA: INFLUÊNCIA DO VOLUME DE SOLO
ILHÉUS – BAHIA – BRASIL FEVEREIRO de 2018

FRANCINNE RIBEIRO ALMEIDA
CULTIVO DE Jatropha curcas L. EM CASA DE VEGETAÇÃO PARA ENSAIO DE DEFICIÊNCIA HÍDRICA: INFLUÊNCIA DO VOLUME DE SOLO
ILHÉUS – BAHIA – BRASIL
FEVEREIRO de 2018
Dissertação apresentada à Universidade Estadual de Santa Cruz, como parte das exigências para obtenção do título de Mestre em Produção vegetal. Área de Concentração: Cultivos em ambiente tropical úmido. Orientador: Prof. Fábio Pinto Gomes Coorintadores: Prof. Ândrea Carla Dalmolin e Prof. Marcelo Schramm Mielke

FRANCINNE RIBEIRO ALMEIDA
CULTIVO DE Jatropha curcas L. EM CASA DE VEGETAÇÃO PARA
ENSAIO DE DEFICIÊNCIA HÍDRICA: INFLUÊNCIA DO VOLUME DE SOLO
Ilhéus, 23 de Fevereiro de 2018
______________________________________ Fábio Pinto Gomes – DS
UESC/DCB (Orientador)
______________________________________ Tessio Araujo de Santana
IFBAIANO/Campus de Teixeira de Freitas
______________________________________ Rogério Ferreira Ribas
UFRB/CCAAB
Dissertação apresentada à Universidade
Estadual de Santa Cruz, como parte das
exigências para obtenção do título de Mestre
em Produção Vegetal.
Área de Concentração: Cultivos em ambiente
tropical úmido

iv
DEDICATÓRIA
Á toda família, amigos e companheiro, principalmente aos meus pais:
Alexsandra Ribeiro (in memorian) e Joeliton Gomes Almeida que mesmo
distantes, estavam comigo ao longo dessa trajetória acadêmica.

1
AGRADECIMENTOS
Agradeço primeiramente a Santíssima Trindade e à Nossa Senhora pela
força e dedicação que me concedeste para conduzir o experimento.
À minha família que mesmo estando fisicamente longe, foram presentes
de uma forma ou de outra, no percorrer desse caminho.
Ao orientador Fábio Gomes pelos ensinamentos transmitidos, pela
orientação e incentivos que foram de grande importância durante o mestrado.
À professora Ândrea Carla Dalmolin, pelo auxílio em fisiologia vegetal e
pela contribuição que tornaram esse trabalho viável.
À equipe Jatropha formada por professores, orientador, coorientadores,
doutorandos e alunos de iniciação científica pela valiosa ajuda no experimento,
principalmente nas medições de área foliar e potencial hídrico. Além de todos os
conhecimentos que foram compartilhados.
Ao Júnior que mesmo entre altos e baixos é meu companheiro nessa
jornada desde a graduação.
Aos integrantes do laboratório de Fisiologia Vegetal, que prestativamente
sempre me auxiliaram quando precisei.
Ao professor Maurício Antonio Coelho Filho e a Mabel Ribeiro Sousa por
toda receptividade e auxílio nas análises morfológicas de raízes no WinRHizo na
EMBRAPA mandioca e fruticultura/ Cruz das Almas-BA.
Ao Dr. Bruno G. Laviola (EMBRAPA – Agroenergia) pelos comentários e
pelo fornecimento de sementes de J. curcas para os ensaios.
À Zezinho e seu Adelino da casa de vegetação e viveiro pela prestativa
ajuda durante o experimento.
À Fundação de Amparo à Pesquisa da Bahia (FAPESB) pela concessão
da bolsa, à Universidade Estadual de Santa Cruz (UESC), ao Conselho Nacional
de Desenvolvimento Científico e Tecnológico (CNPq) que permitiram a
concretização desse trabalho.

2
RESUMO
Devido à importância de Jatropha curcas L. no contexto da crescente busca por
energias alternativas, associada aos efeitos da deficiência hídrica na produção
vegetal, neste trabalho propõe-se estudar os aspectos fisiológicos e
morfológicos agregados à oferta e à demanda hídrica, condições de cultivo, e
suas variações genotípicas mantidas sob déficit hídrico moderado. Os objetivos
desse estudo foram: a) determinar a associação existente entre suprimento
hídrico, volume de solo, crescimento e a biomassa seca dos genótipos CNPAE
517 e 557 e b) Verificar alterações nessa associação em plantas submetidas à
deficiência hídrica moderada no solo em diferentes genótipos da espécie em
estudo. O experimento foi conduzido entre abril de 2017 e julho de 2017, em
casa de vegetação (14°47'00" S, 39°02'00" W). Os tratamentos consistiram em
dois regimes hídricos, plantas controle (100% da CC), plantas submetidas à
deficiência hídrica moderada (75% da CC), cinco volumes de solo (2,4, 4, 8,2,
16,4 e 33 dm³) e dois genótipos. O experimento foi conduzido em um
delineamento inteiramente casualizado, em esquema fatorial (2 genótipos x 2
regimes hídricos x 5 volumes de solo) com 12 repetições, totalizando 240
plantas. Aos 30 dias após à imposição dos tratamentos (DAIT), houve coleta
destrutiva de 50% das plantas, permanecendo 120 em casa de vegetação até
os 60 DAIT. Os resultados foram submetidos à análise de variância (ANOVA) e,
quando indicado, as comparações de médias foram realizadas pelo Teste de F
a 5% de probabilidade, pela diferença mínima significativa (também a 5%) e por
regressões. Verificou-se que houve efeito significativo dos genótipos, dos
tratamentos hídricos e/ou de volumes de solo em todas as variáveis avaliadas
(relações hídricas, trocas gasosas foliares, variáveis de crescimento e descrição
morfológica de raízes). As trocas gasosas foram avaliadas aos 30 e 60 DAIT, e
pôde-se observar que no genótipo 557, com deficiência hídrica no solo, a
redução da condutância estomática ao vapor de água (gs) foi maior comparado
ao CNPAE 517. Houve efeito significativo dos tratamentos sobre a taxa

3
fotossintética, sendo que o genótipo CNPAE 557 apresentou valores inferiores
de fotossíntese em comparação ao CNPAE 517. No genótipo CNPAE 557, sob
deficiência hídrica no solo, foi observado maiores reduções na taxa de
crescimento relativo de altura, aos 30 DAIT, área foliar total e comprimento
radicular, ao passo que, no genótipo CNPAE 517 verificou-se menores reduções
na taxa de crescimento relativo de diâmetro do caule, aos 30 DAIT, biomassa
radicular e relação raiz/parte aérea. As variáveis de crescimento tais como:
Altura da planta, área foliar e biomassa total, também foram reduzidas
significativamente nas plantas submetidas aos menores volumes de solo do que,
nas plantas cultivadas em maiores volumes. A maioria das variáveis
morfológicas de raízes, avaliadas aos 60 DAIT, apresentaram maiores médias
em plantas submetidas à deficiência hídrica moderada, do que, nas plantas
controles, indicando que o investimento em raízes, sobretudo finas, é uma
estratégia de tolerância à seca para esta espécie. O cultivo de J. curcas L. em
volumes de solo menores que 16,4 dm³ por 60 dias resultou em menor
crescimento das plantas em termos de biomassa e de trocas gasosas foliares. A
limitação ao crescimento, provocada pelo menor volume de solo, foi observada
primeiro em plantas controle, mascarando os efeitos da deficiência hídrica
moderada em experimentos com esse intuito. Nos dois genótipos estudados,
diferentes padrões de crescimento e de trocas gasosas em volumes de solo
diversos foram observados. Sob disponibilidade hídrica moderada e nos
menores volumes de solo, maior comprimento, área de raízes e classes de
diâmetro radicular foram detectados.
Palavras-chave: Trocas gasosas, fotossíntese, crescimento, volume de raiz,
épocas de coleta
ALMEIDA, Francinne Ribeiro, M.Sc., Universidade Estadual de Santa Cruz,
fevereiro de 2018. Cultivo de Jatropha curcas L. em casa de vegetação para
ensaio de deficiência hídrica: influência do volume de solo. Orientador:
Fábio Pinto Gomes. Coorientadores: Ândrea Carla Dalmolin e
Marcelo Schramm Mielke.

4
ABSTRACT
Due to the importance of Jatropha curcas L. in the context of the growing search
for alternative energies, associated with the effects of water deficiency on plant
production, this work proposes to study the physiological and morphological
aspects aggregated to water supply and demand, growing conditions, and their
genotypic variations maintained under moderate water deficit. The objectives of
this study are: a) to determine the association between water supply, soil volume
effect, growth and dry biomass of genotypes CNPAE 517 and 557; b) To verify
changes in this association in plants submitted to moderate water deficiency in
the soil in different genotypes of the species under study. The experiment was
conducted between april 2017 and july 2017, under greenhouse conditions (14 °
47'00 "S, 39 ° 02'00" W). The treatments consisted of two water regimes, control
plants (100% of FC), plants submitted to moderate water deficiency (75% of FC)
and five soil volumes (2,4, 4, 8,2, 16,4 and 33 dm³). Stomatal conductance (gs)
evaluations were performed using a portable system of photosynthesis
measurements, the water status of the plants was evaluated through
measurements of leaf water potential (Ψw). The characteristics and morphology
of the roots at 60 days after the imposition of the treatment were evaluated
through the WinRhizo software. The experiment was conducted in a completely
randomized design, in a factorial scheme. The results were submitted to analysis
of variance (ANOVA) and, when indicated, the means comparisons were
performed by the F test at 5% of probability and by the least significant difference
(also at 5%). There was a significant effect of genotypes and water treatments
and soil volume in almost all evaluated variables. The reduction of stomatal
conductance to water vapor with soil water deficiency was slower in the CNPAE
557. There was a significant effect of the treatments on the photosynthetic rate,
where the genotype CNPAE 557 showed the lowest photosynthesis. In the
genotype CNPAE 557 greater reductions in soil water deficit were observed in
the relative growth rate of height, leaf area and root length, while in the genotype
CNPAE 517 there were smaller reductions in the relative growth rate of stem
diameter, root biomass and root / shoot ratio. Growth variables such as plant

5
height, leaf area and total biomass were also significantly reduced in plants with
lower soil volumes than in plants grown in higher volumes. Almost all root
morphological variables evaluated at 60 days after treatment imposition showed
higher mean values in plants submitted to the deficiency, allowing that in the
control plants, indicating a drought tolerance strategy for this species. The soil
volume, under the conditions of the present experiment, significantly influenced
the growth, biomass partition and foliar gas exchange of the plants of J. curcas.
The different soil volumes studied, under the conditions tested, have different
effects under full irrigation or under moderate water deficiency. The two
genotypes present different growth patterns and gas changes in several soil
volumes and under limiting water availability. As plants grown in lower volumes
of soil and water supply, in comparisons such as control plants, volume, root area
and diameter classes.
Keywords: Gaseous exchanges, photosynthesis, growth, root volume,
harvesting times
ALMEIDA, Francinne Ribeiro, M.Sc., Santa Cruz State University, February, 2018. Cultivation of Jatropha curcas L. in a greenhouse to test for water deficiency: influence of soil volume. Advisor: Fábio Pinto Gomes. Coadvisers: Ândrea Carla Dalmolin and Marcelo Schramm Mielke.

6
LISTA DE SÍMBOLOS E ABREVIAÇÕES
A Taxa de fotossíntese líquida (µmol m-2 s-1)
AF Área foliar total
BR Biomassa Radicular
C Plantas controles
CC Capacidade de campo (%)
CR Comprimento Radicular
DAIT Dias após a imposição do tratamento
DH Plantas submetidas à deficiência hídrica no solo
gs Condutância estomática ao vapor de água (mol H2O m-2 s-1)
RFA Radiação fotossinteticamente ativa (μmol fotons m-2 s-1)
R/PA Razão Raiz/Parte Área
T Temperatura (°C)
TCRa Taxa de crescimento relativo em altura
TCRd Taxa de crescimento relativo em diâmetro
VR Volume Radicular
UR Umidade Relativa (%)
US Umidade do Solo (%)
Ψw Potencial hídrico foliar (MPa)

7
SUMÁRIO
RESUMO.............................................................................................................2
ABSTRACT..........................................................................................................4
1.INTRODUÇÃO..................................................................................................8
2.REVISÃO DE LITERATURA.............................................................................9
2.1 SITUAÇÃO ENERGÉTICA NO MUNDO.............................................9
2.2 PINHÃO MANSO (Jatropha curcas L.).............................................11
2.3 DEFICIÊNCIA HÍDRICA ...................................................................13
2.4 EFEITO DO VOLUME DE SOLO......................................................14
3.MATERIAL E MÉTODOS...............................................................................16
3.1 MATERIAL BIOLÓGICO E CONDIÇÕES DE CULTIVO..................16
3.2 CARACTERIZAÇÃO MICROCLIMÁTICA.........................................18
3.3 TROCAS GASOSAS FOLIARES......................................................19
3.4 RELAÇÕES HÍDRICAS.....................................................................19
3.5 AVALIAÇÃO DE CRESCIMENTO ..................................................19
3.6 DESCRIÇÃO MORFOLÓGICA DE RAÍZES.....................................20
3.7 DELINEAMENTO EXPERIMENTAL E ANÁLISE ESTATÍSTICA.....20
4. RESULTADOS...............................................................................................21
4.1 RELAÇÕES HÍDRICAS ...................................................................21
4.2 TROCAS GASOSAS FOLIARES......................................................23
4.3 VARIÁVEIS DE CRESCIMENTO......................................................26
4.4 DESCRIÇÃO MORFOLÓGICA DE RAÍZES.....................................35
5. DISCUSSÃO..................................................................................................45
6.CONCLUSÕES ..............................................................................................48
7.REFERÊNCIAS..............................................................................................50

8
1. INTRODUÇÃO
A produção de culturas para a utilização como fonte energética impacta a
qualidade da água (pH, teor de nutrientes, sais, metais pesados) bem como a
quantidade a ser consumida, sendo que este consumo varia em função desde o
sistema de produção das matérias-primas até a conversão destas em
combustíveis. O grau de impacto dependerá de variáveis como a região,
condições climáticas, cadeia de fornecimento, tipo da matéria-prima, métodos de
produção, eficiência de conversão e o produto final requerido (UNEP, 2010).
Nota-se que a produção em larga escala dos cultivos energéticos pode aumentar
a evapotranspiração e, em alguns países, potencializar uma situação de
estresse hídrico (YANG et al., 2009).
O pinhão manso (Jatropha curcas L.), pertencente à família Euforbiaceae, é
uma espécie oleaginosa, bastante promissora para produção de biodiesel, e
embora tenha sido caracterizada como uma espécie tolerante à seca, tem
demonstrado respostas negativas ao déficit hídrico, apesar de sobreviver e
mostrar boa capacidade de recuperação (SAPETA et al., 2013).
A espécie é conhecida e cultivada no continente americano desde a época
pré-colombiana e esta disseminada em todas as regiões tropicais e até em
algumas áreas temperadas, com um bom desenvolvimento em regiões de solos
férteis e bem estruturados, clima quente e úmido (ACHTEN et al., 2008, contudo
trata-se de uma espécie em domesticação (ACHTEN et al., 2010). A viabilidade
de cultivo ainda depende do desenvolvimento de tecnologia agronômica e do
cultivo para obtenção de altas produtividades nas diferentes regiões do Brasil
(FREITAS et al., 2011; LAVIOLA et al., 2012).
O déficit hídrico modifica diversos processos fisiológicos das plantas, dentre
eles, destacam-se o fechamento estomático, redução do limbo foliar, do
crescimento e da biomassa seca, geralmente aumentando a resistência
estomática ao vapor de água, reduzindo a transpiração e, consequentemente, o
suprimento de CO2 para a realização do processo de fotossíntese, bem como
alterando a produção de ácido abscísico, abscisão foliar e as características
osmóticas das plantas (GOMES; PRADO, 2007).

9
Outro parâmetro igualmente importante, é o estudo do volume dos vasos
utilizados no cultivo de plantas, pois, o tempo de permanência podem interferir
no desenvolvimento vegetal, especialmente quando ocorre limitação física no
crescimento radicular, promovendo alterações na capacidade de absorção de
água, nos compostos sintetizados e seu transporte para as regiões de
crescimento da parte aérea. Carneiro et al. (2007), relatou a influência do
recipiente utilizado na formação de mudas, salientando a importância de se
conhecer seus efeitos, sendo necessário estudar as dimensões e os volumes
dos vasos. Uma vez que o sub ou superdimensionamento pode implicar em
restrições radiculares, o que afeta diretamente o metabolismo vegetal e impede
o pleno crescimento e desenvolvimento das plantas, além de onerar a atividade.
Diante das hipóteses de que (a) o volume do solo afeta o crescimento,
partição de biomassa e trocas gasosas de J. curcas; (b) os efeitos do volume de
solo são diferentes sob irrigação plena ou sob deficiência hídrica e (c) os dois
genótipos possuem diferentes padrões de crescimento e trocas gasosas quando
cultivados em recipientes com diferentes volumes de solo e sob disponibilidade
hídrica limitante, os objetivos desse estudo foram: 1) Verificar a existência de
uma associação entre o suprimento hídrico, efeito de volume de solo,
crescimento e a biomassa dos genótipos CNPAE 517 e CNPAE 557, e 2)
Descrever quais os tipos de alterações ocorrem nessa associação em plantas
submetidas à deficiência hídrica moderada no solo nos diferentes genótipos da
espécie em estudo.
2. REVISÃO DE LITERATURA
2.1 SITUAÇÃO ENERGÉTICA NO MUNDO
Atualmente uma grande preocupação tem surgido com relação às novas
fontes de energia que possam estar suprindo a demanda energética mundial e
preservando o meio ambiente, de forma a reduzir a emissão de gases do efeito
estufa liberados na utilização de combustíveis fósseis para geração de energia.

10
A produção de biocombustíveis em larga escala configura uma alternativa para
geração de renda e emprego em muitas áreas rurais em todo o mundo. Em vista
do enorme mercado de combustíveis para transporte, os benefícios podem ser
realmente expressivos, indo desde menor dependência do petróleo até a
exportação de biocombustíveis, passando pelos ganhos ambientais da redução
da emissão de gases do efeito estufa e do aumento do sequestro de carbono
atmosférico em biomassa, sobretudo no caso do uso de espécies perenes
(CREMONEZ et al., 2015).
As crescentes preocupações sobre a diminuição da disponibilidade dos
combustíveis fósseis principalmente oriundos do petróleo, juntamente com os
problemas ambientais resultantes da sua exploração, produção e utilização, têm
estimulado o interesse pelos combustíveis renováveis a fim de aumentar a
necessidade global de reduzir a emissão de gases de efeito de estufa. Os
biocombustíveis que são globalmente considerados fontes de energia
sustentáveis e ecológicas, com papel importante na diversificação dos
suprimentos energéticos para atender às crescentes necessidades energéticas
do mundo e diminuir a dependência de combustíveis fósseis, o que tem se
tornando uma alternativa de combustível menos poluente (JAT et al., 2015).
Além dos efeitos ambientais, as culturas estudadas tais como a mamona, o
dendê, a soja, o pinhão manso, o girassol, a canola, o amendoim, o algodão e
outras oleaginosas podem ser produzidas de forma sustentável, trazendo
desenvolvimento regional e inclusão social por meio da agricultura familiar.
Nesse sentido, como incentivo ao pequeno produtor, o governo instituiu impostos
diferenciados de acordo com cada matéria-prima, sendo reservado ao Norte-
Nordeste o maior desconto por matéria-prima produzida (JESUS e SILVA, 2013).
Entre as inúmeras fontes de óleo vegetal para a fabricação de biodiesel,
Jatropha curcas, uma espécie perene de porte arbóreo-arbustivo, pertencente a
família Euphorbiaceae, tem sido apontada como uma das mais promissoras
(EDRISI et al., 2015). Essa espécie apresenta-se amplamente distribuída em
regiões tropicais, onde tem mostrado crescimento satisfatório em solos
pedregosos e de baixa umidade. O óleo extraído das sementes (que pode chegar
a 50% em alguns genótipos) tem sido utilizado como purgativo além de inúmeros
outros usos medicinais, na fabricação de tintas e sabões e como lubrificante e
combustível para motores a diesel. Além disso, J. curcas apresenta alto potencial

11
industrial, sendo o resíduo da extração do óleo rico em proteína (60-65%),
podendo ser transformado em um excelente alimento para aves, ruminantes e
peixes (CONTRAN et al., 2013). Importante ressaltar, ainda, uma outra
vantagem de J. Curcas quando comparada à outras espécies, que é a não
competição com outras culturas produtoras de óleos comestíveis (CONTRAN et
al., 2013).
2.2 PINHÃO MANSO (Jatropha curcas L.)
O pinhão manso (Jatropha curcas L.), pertencente à família Euforbiaceae,
é uma espécie conhecida e cultivada no continente americano desde a época
pré-colombiana e esta disseminada em todas as regiões tropicais e até em
algumas áreas temperadas, com um bom desenvolvimento em regiões de solos
férteis e bem estruturados, clima quente e úmido (ACHTEN et al., 2008, contudo
trata-se de uma espécie em domesticação (ACHTEN et al., 2010). A viabilidade
de cultivo ainda depende do desenvolvimento de tecnologia agronômica e do
cultivo para obtenção de altas produtividades nas diferentes regiões do Brasil
(LAVIOLA et al., 2012).
Essa espécie possui seu centro de origem na América central,
principalmente no México (MAGHULY et al., 2015), mas atualmente é distribuída
e cultivada em todos os trópicos e está sendo considerada uma opção agrícola
para as regiões áridas e semiáridas, por ser uma espécie exigente em luz e com
características de tolerância à seca (ACHTEN et al., 2010).
Jatropha curcas caracteriza-se como uma pequena árvore ou um grande
arbusto que atinge de 5 à 10 m de altura com uma expectativa de vida de até
50 anos, podendo florescer em várias zonas climáticas, com precipitação anual
entre 250 e 1200 mm (ACHTEN et al., 2008). O pinhão manso é um arbusto de
crescimento rápido que em condições favoráveis, atingindo geralmente, uma
altura entre 8 a 10 metros, as plantas são monóicas com flores unissexuais, a
floração ocorre durante a estação chuvosa sendo que em regiões úmidas ocorre
ao longo do ano, levando de três a quatro anos para atingir a idade produtiva,

12
possuindo uma colheita a cada seis meses que se estende por 40 anos (ACHEN
et al., 2008; KUMAR; SUMAR, 2008).
As plantas geralmente formam uma raiz central (pivotante), quatro raízes
laterais e muitas raízes secundárias. São decíduas, perdendo as folhas na
estação de seca. As folhas são dispostas alternadamente sobre o caule, lobadas
superficialmente e variam de 6 a 15 cm de comprimento e largura; o tamanho e
a forma das folhas podem diferir de uma variedade para outra. Tal como com
outros membros desta família, os tecidos vasculares dos caules e ramos contêm
látex branco. Os ramos e hastes são ocos e a madeira macia é de pouco valor
(KUMAR et al., 2016; MARTIN et al., 2010).
Os frutos são elipsoidais, verdes e carnudos, ficando amarelos e depois
castanhos à medida que envelhecem. As frutas tornam-se maduras e prontas
para a colheita cerca de 90 dias após a floração. Cada fruto contém duas ou três
sementes pretas, com cerca de 2 cm x 1 cm de tamanho. Suas sementes
possuem elevada quantidade de óleo (teor de óleo: 27 a 40%, 250 g de óleo por
kg de sementes) produz no mínimo, duas toneladas de óleo por hectare, sendo
uma das plantas oleaginosas mais promissoras para produção do biodiesel. Para
melhores rendimentos de óleo, as sementes devem ser colhidas na maturidade
quando a cor dos frutos mudarem de verde para amarelo-marrom (ACHEN et al.,
2008; AHAMED, 2016; MARTIN et al., 2010).
Todas as partes da planta de J. curcas podem ser utilizadas para as mais
variadas aplicações como adubo, combustível, inseticida, cerca viva em algumas
pequenas propriedades. Na medicina tradicional, o látex pode atuar como
cicatrizante em feridas externas, além de suas folhas e a amêndoa apresentarem
propriedade purgativa, porém a ingestão de grandes quantidades pode levar à
óbito (ARAÚJO, 2008).
É uma espécie oleaginosa, bastante promissora para produção de
biodiesel, e embora tenha sido caracterizada como uma espécie tolerante à
seca, tem demonstrado respostas negativas ao déficit hídrico, apesar de
sobreviver e mostrar boa capacidade de recuperação (FINI et al., 2013; SAPETA
et al., 2013)

13
2.3 DEFICIÊNCIA HÍDRICA
Dentre os diversos fatores abióticos limitantes ao desenvolvimento das
culturas, a água é considerada como o fator mais restritivo. Em situações de
déficit hídrico, as plantas têm seus processos metabólicos alterados. Após a
estagnação do crescimento da parte aérea devido ao estresse, ocorre uma
consequente diminuição na demanda de fotoassimilados, que podem ser
alocados na parte subterrânea, favorecendo o crescimento de raízes na busca
de locais mais úmidos do solo (TAIZ; ZEIGER, 2009).
A deficiência hídrica modifica diversos processos fisiológicos das plantas,
geralmente aumentando a resistência estomática ao vapor de água, reduzindo a
transpiração e, consequentemente, o suprimento de CO2 para a realização do
processo de fotossíntese, bem como alterando a produção de ácido abscísico,
abscisão foliar e as características osmóticas das plantas (GOMES; PRADO,
2007), retardando o seu crescimento e desenvolvimento, característica negativa
para qualquer cultivo econômico (De OLIVEIRA, 2013).
A deficiência hídrica definida como um desequilíbrio entre a
disponibilidade de água no solo e a demanda por evaporação que pode
naturalmente ocorrer no campo, provocam nas plantas uma diminuição da
acumulação de carbono e do número de células, as paredes celulares dos
tecidos foliares tornam-se menos extensíveis, reduzindo assim, a taxa de
expansão dos tecidos, têm efeito da redução da área foliar na interceptação de
luz (menor ganho de carbono), restrição da condutância estomática e redução
da biomassa. Quando os estômatos se fecham parcialmente, diminuindo assim
a transpiração, o potencial hídrico foliar (a variável física que representa o grau
de hidratação tecidual) aumenta, este mecanismo permite que as folhas
mantenham seu status de água em uma escala estreita. As raízes têm um papel
de sinalização neste processo através da circulação de mensagens hidráulicas
e químicas para as folhas quando elas começam a sofrer um estresse por
esgotamento da água (TARDIEU et al., 2011).
As respostas das plantas aos efeitos provocados pelo déficit hídrico se
dão por meio de diversos processos adaptativos, podendo apresentar, por

14
exemplo, fechamento dos estômatos em horários com alta taxa de evaporação
para manutenção do equilíbrio hídrico celular (PIMENTEL, 2004).
A deficiência hídrica é um dos fatores limitantes da produção vegetal, pois
afeta as relações hídricas nas plantas e ocorre em grandes extensões de áreas
cultiváveis (PASSIOURA, 2007).
Devido aos altos custos da irrigação e a necessidade de cultivar em
ambientes cada vez mais áridos, torna-se necessária à utilização de genótipos
mais adaptados à seca. Um mecanismo que faz a planta utilizar a água com o
máximo de eficiência é a redução da condutância estomática, diminuindo a taxa
transpiratória. Porém, ao selecionar espécies tolerantes à seca com tais
características, essas manterão a limitação de seu crescimento, devido ao
fechamento dos estômatos, que reduzirá a captação de CO2, composto
necessário para reação da fotossíntese (SAMBATTI; CAYLOR, 2007).
Apesar de resultados mostrarem um atraso no crescimento do pinhão
manso submetidos à deficiência hídrica, Fini et al. (2013) concluíram que J.
curcas pode sobreviver a períodos de seca de intensidade moderada.
Entretanto, os efeitos na rentabilidade econômica nesse tipo de cultivo ainda não
são completamente elucidados.
Estudos relacionados às respostas de genótipos, provenientes de
diferentes regiões geográficas e climáticas, à deficiência hídrica estão em
constante processos experimentais, sendo necessário ainda, uma maior
caracterização dos genótipos obtidos. Dessa forma, avaliar o crescimento e
desenvolvimento de diferentes genótipos torna-se importante para o sucesso de
um programa de melhoramento para esta oleaginosa.
2.4 EFEITO DO VOLUME DE SOLO
As plantas são frequentemente expostas à deficiência de água e à
reidratação em ciclos no solo, e é uma das principais causas de perda e redução
nos rendimentos das culturas (CHEN et al., 2015). Em uma escala evolutiva à
exposição de diferentes tipos ambientais, o fator tempo, também levou as
plantas a desenvolver algumas adaptações complexas para evitar ou tolerar

15
mudanças periódicas ou esporádicas na disponibilidade de água e nutrientes no
solo (DOBRA et al., 2010). As limitações no teor de água do solo podem ser
percebidas pelas plantas, física e quimicamente, levando a mudanças
fisiológicas, morfológicas, anatômicas, bioquímicas e moleculares,
fundamentais para aumentar a sobrevivência destas, sob condições
estressantes (XU et al., 2010).
O volume e tipo de solo utilizado no cultivo de plantas influenciam no
transporte de água das raízes às folhas, como suprimento hídrico, sendo de
fundamental importância para a produção das culturas. Estudando o sistema
radicular, Zonta et. al. (2006) afirma que somente a redução no comprimento de
raízes não deve ser vista como uma diminuição no crescimento radicular, pois
em condições de limitação de crescimento em profundidade, ocorre maior
proliferação de raízes laterais finas e aumento da superfície radicular especifica,
como estratégia de sobrevivência.
O conteúdo e o potencial de água no solo, as condições de temperatura,
aeração, resistência mecânica do solo à penetração e a exploração do sistema
radicular, além da disponibilidade de nutrientes e água influenciam no
desenvolvimento radicular (SÁ; SANTOS JÚNIOR, 2005).
Variáveis de crescimento tais como: altura, diâmetro de caule, número de
folhas, volume radicular, dentre outras, são fortemente afetadas em função do
volume de solo contido no recipiente utilizado para o crescimento das plantas.
Souza; Freire e Sampaio, (2009) reportaram que mudas de J. curcas produzidas
em volumes de 2,1 e 3,9 dm³ de volume de solo, apresentaram maior
crescimento em altura e no diâmetro do caule seis meses após o plantio nas
condições de Porto Velho, RO, quando comparados à menores volumes de solo.
O uso de pequenos volumes de solo para fins de pesquisa pode
apresentar desvantagens, que estão mais relacionadas a restrições
biológicas. Um pequeno volume implica quase invariavelmente resulta, numa
redução na disponibilidade de água e nutrientes para a planta. Além da reduzida
disponibilidade de recursos, pode ocorrer também, o impedimento físico do
crescimento das raízes. Muitas espécies produzem facilmente raízes de mais de
1 m de comprimento (JACKSON et al. 1996), mesmo em idade relativamente
jovem (FUSSEDER 1987), excedendo assim as dimensões da maioria dos
recipientes utilizados. Plantas grandes em pequenos vasos podem ter uma

16
grande fração de raízes 'encapsuladas', com todo tipo de conseqüências
secundárias (HEROLD e MCNEIL, 1979).
Esforços consideráveis têm sido dirigidos para o entendimento das
associações existentes entre as condutâncias (estomática e mesofílica) do CO2
e o suprimento hídrico para sustentar a demanda via condutividade hidráulica
das raízes para as folhas (GRIFFITHS et al., 2013). Assim, os estudos sobre
alocação e distribuição hídrica, bem como as relações entre essas
características durante o desenvolvimento de raízes sob condições estressantes
no solo devem ser incluídos nas pesquisas sobre hidráulica e relações hídricas
em plantas.
3. MATERIAL E MÉTODOS
3.1 MATERIAL BIOLÓGICO E CONDIÇÕES DE CULTIVO
O experimento foi conduzido em casa de vegetação, no Campus da
Universidade Estadual de Santa Cruz, município de Ilhéus, BA, Brasil (14°47'00"
S, 39°02'00" W). De acordo com a classificação climática de Köppen, o clima
local é do tipo Af, com temperaturas médias anuais de 22 a 25°C. O período
experimental foi de 30 de abril de 2017 até 28 de julho de 2017. Foram utilizadas
sementes de dois genótipos de Jatropha curcas L., (CNPAE 517 e CNPAE 557),
oriundos do banco de germoplasma da Embrapa – Agroenergia / DF.
As sementes foram higienizadas com solução de hipoclorito de sódio a
2% e tratadas com fungicida Carbamax 500 sc de acordo com a recomendação
do fabricante. Em seguida, foram colocadas para germinar em vasos (cinco
sementes por vaso), contendo 2,4, 4, 8,2, 16,4 e 33 dm³ de solo com classe
textual médio arenoso, (Tabela 1), dimensão do solo especificada na tabela 2
e densidade do solo de 1,22 g dm-³, determinado pelo método da proveta
descrita por Brasil (1949) e EMBRAPA (1979).
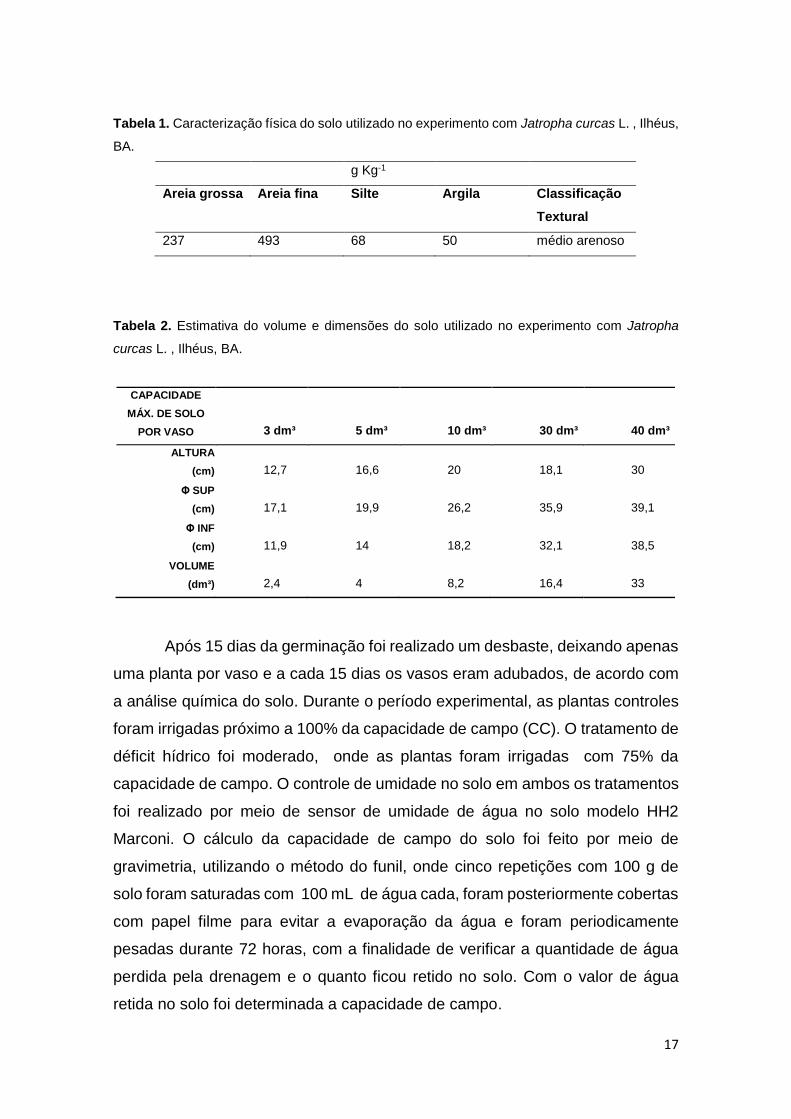
17
Tabela 1. Caracterização física do solo utilizado no experimento com Jatropha curcas L. , Ilhéus,
BA.
Tabela 2. Estimativa do volume e dimensões do solo utilizado no experimento com Jatropha
curcas L. , Ilhéus, BA.
Após 15 dias da germinação foi realizado um desbaste, deixando apenas
uma planta por vaso e a cada 15 dias os vasos eram adubados, de acordo com
a análise química do solo. Durante o período experimental, as plantas controles
foram irrigadas próximo a 100% da capacidade de campo (CC). O tratamento de
déficit hídrico foi moderado, onde as plantas foram irrigadas com 75% da
capacidade de campo. O controle de umidade no solo em ambos os tratamentos
foi realizado por meio de sensor de umidade de água no solo modelo HH2
Marconi. O cálculo da capacidade de campo do solo foi feito por meio de
gravimetria, utilizando o método do funil, onde cinco repetições com 100 g de
solo foram saturadas com 100 mL de água cada, foram posteriormente cobertas
com papel filme para evitar a evaporação da água e foram periodicamente
pesadas durante 72 horas, com a finalidade de verificar a quantidade de água
perdida pela drenagem e o quanto ficou retido no solo. Com o valor de água
retida no solo foi determinada a capacidade de campo.
g Kg-1
Areia grossa Areia fina Silte Argila Classificação
Textural
237 493 68 50 médio arenoso
CAPACIDADE
MÁX. DE SOLO
POR VASO 3 dm³ 5 dm³ 10 dm³ 30 dm³ 40 dm³
ALTURA
(cm) 12,7 16,6 20 18,1 30
Φ SUP
(cm) 17,1 19,9 26,2 35,9 39,1
Φ INF
(cm) 11,9 14 18,2 32,1 38,5
VOLUME
(dm³) 2,4 4 8,2 16,4 33

18
O controle de pragas no experimento foi realizado de forma integrada,
com aplicação do inseticida sistêmico Actara 250 wg, a concentração de acordo
com o ministério da agricultura pecuária e abastecimento (MAPA) e com o uso
de armadilhas adesivas.
O registro microclimático foi realizado utilizando uma estação
metereológica HOBOWare Pro Series (Onset, USA), durante todo o período
experimental.
3.2 CARACTERIZAÇÃO MICROCLIMÁTICA
A intensidade da radiação fotossinteticamente ativa (RFA) no interior da
casa de vegetação, acompanhada de 06:00 às 18:00 h, durante todo o período
de condução do experimento (60 DAIT) variou entre 2,8 e 13 mol de fótons m-
2 dia-1, com uma média de 10,6 mol de fótons m-2 dia-1 (Tabelas 3).
Tabela 3. Radiação fotossinteticamente ativa (RFA - mol fótons m-2 dia-1), temperatura do ar (T
- °C), umidade relativa do ar (UR - %), e US (W%) em plantas controle (C) e sob deficiência
hídrica moderada no solo (DH) dentro da casa de vegetação durante o período experimental (0-
30 DAIT e 30-60 DAIT). Os valores correspondem à média (± erro padrão da média), máximo e
o mínimo diário.
MÉDIA
30 DAIT
MÍNIMO MÉDIA
60 DAIT
MÍNIMO MÁXIMO MÁXIMO
RFA 9,8±0,04 11,4 3,3 10,6±0,04 13 2,8
T 20,8±0,02 31,6 18 23,8±0,03 32 16
UR% 79,8±0,2 98,5 57,3 87,4±0,3 98,7 57,3
WDH% 24,5±0,6 26,0 19,0 24,5±0,6 26,0 19,0
WC% 35±0,5 38,0 32,0 35±0,5 38,0 32,0
A temperatura média diária do ar variou de 16,3 a 31,7 °C com uma média
de 23,8 °C, (± 0,03 ºC).

19
Os picos dos valores de umidade relativa do ar foram 57,3% (mínimo) a
98,7% (máximo), atingindo valores médios de 87,4%, (± 0,3).Com relação à
umidade do solo.
Os maiores valores de umidade no solo foram obtidos no início do
tratamento, e posteriormente, em plantas controles, que se mantiveram
constantes. Plantas submetidas à DH apresentaram as menores umidades do
solo durante o período experimental.
3.3 TROCAS GASOSAS FOLIARES
As trocas gasosas foram avaliadas mensalmente (Aos 30 e 60 DAIT) em
folhas completamente maduras, entre 8:00 e 12:00 horas da manhã, com um
sistema portátil de medição de fotossíntese (IRGA Infrared Gas Analizer) modelo
Li-6400 (Li-Cor, Biosciences Inc., Nebraska, EUA). Durante as avaliações, a
Radiação foi de 1.000 mol fótons m-2 s-1, e (Ca) ambiente de CO2 de ± 390 µmol
mol- 1 e a temperatura do bloco foi fixada em 28 °C. A condutância estomática ao
vapor de água (gs) e a taxa fotossintética (A) foram medidas usando os valores
da variação de CO2 e da umidade no interior da câmara, ambos avaliados pelo
analisador de gases por infravermelho do sistema portátil de fotossíntese.
3.4 RELAÇÕES HÍDRICAS
O estado hídrico das plantas foi medido in situ por meio do potencial
hídrico foliar (Ψam), avaliado aos 30 e aos 60 dias após a imposição do
tratamento, utilizando uma câmara de pressão modelo 1000 (PMS Instrument
Company, EUA), no período de antemanhã.
3.5 AVALIAÇÃO DE CRESCIMENTO
O crescimento das plantas foi quantificado mensalmente por meio de
medidas de altura e diâmetro do coleto. Ao final de cada coleta (aos 30 e 60
DAIT), (figura 1), realizou-se a medida da área foliar de cada planta. A altura foi

20
medida usando uma fita métrica graduada, considerando-se a distância entre o
nível do solo e o ápice da planta. O diâmetro do coleto foi determinado com o
auxílio de um paquímetro. Para calcular a taxa de crescimento relativo para
altura e diâmetro foi utilizada a fórmula descrita por Hunt, (1989).
A área foliar das plantas foi medida ao final do experimento, em cada
coleta, utilizando o medidor de área foliar LI-3100 (Li- Cor, inc. Lincoln, Nebraska,
USA).
Ao final das épocas de coleta, as plantas foram coletadas para a
determinação da biomassa, particionadas em raiz, caule e folhas, e, logo após,
secas em estufa de ventilação forçada de ar (65°C ± 5°), até atingirem massa
constante. A partir da biomassa dos diferentes órgãos, foi determinada a razão
raiz/parte aérea (R/PA), obtida pela razão entre a biomassa seca de raiz e a
biomassa seca do caule e das folhas.
3.6 DESCRIÇÃO MORFOLÓGICA DE RAÍZES
As amostras de raízes coletadas aos 60 DAIT (quatro repetições), foram
armazenadas de acordo com Bohm (2012) até a análise. O comprimento da raiz,
volume, densidade, diâmetro, comprimento por diâmetro, classes e área de
superfície, foram determinadas usando o software WinRhizo (WR, versão 2013d,
Regent Instrument, Quebec, Canadá).
3.7 DELINEAMENTO EXPERIMENTAL E ANÁLISE ESTATÍSTICA
O experimento foi conduzido em delineamento inteiramente casualizado
(DIC), em um esquema fatorial formado por dois regimes hídricos x dois
genótipos de J. curcas, x cinco volumes de solo x seis repetições para as
plantas controles x seis repetições para o tratamento com deficiência hídrica
moderada e duas épocas de coleta perfazendo um total de 240 plantas (Figuras
1 e 2). Os resultados foram submetidos à análise de variância (ANOVA) e,
quando indicado, as comparações de médias foram realizadas pelo Teste de F
a 5% de probabilidade e pela diferença mínima significativa (também a 5%).

21
Regressões foram feitas sempre que houve diferença significativa entre
os tratamentos.
Figura 1. Visão do experimento com Jatropha curcas L. utilizando genótipos CNPAE 517 e 557,
nos volumes 2,4, 4, 8,2, 16,4 e 33 dm³ de solo, sob plena irrigação 100% CC e deficiência hídrica
moderada 75% CC, após 30 DAIT.(Letra A) e próximo aos 60 DAIT (Letra B).
aos 60 DAIT.
4. RESULTADOS
4.1 RELAÇÕES HÍDRICAS
Aos 30 e aos 60 DAIT, verificou-se diferenças significativas (p<0,05) no
potencial hídrico foliar antemanhã, entre os volumes de solo, com valores
variando entre -0,2 (2,4 dm³) e -0,6 (2,4 dm³) MPa no genótipo CNPAE 557, nas
diferentes épocas de coleta. Já o genótipo CNPAE 517, apresentou diferença
significativa entre volumes de solo, apenas aos 30 DAIT, com valores que
variaram entre -0,3 e -0,5 MPa. O ΨWam não diferiu entre tratamento hídrico, e
nem entre genótipos. Em geral, as plantas submetidas à DH apresentaram
valores mais negativos do que os das plantas controle, sendo que, nas plantas
do genótipo 517 o valor mais negativo foi de -0,5 Mpa, em volume de 4 dm³ de
Fontes: Arquivo Pessoal B
Fonte: Arquivo Pessoal
A B
Fonte: Arquivo Pessoal
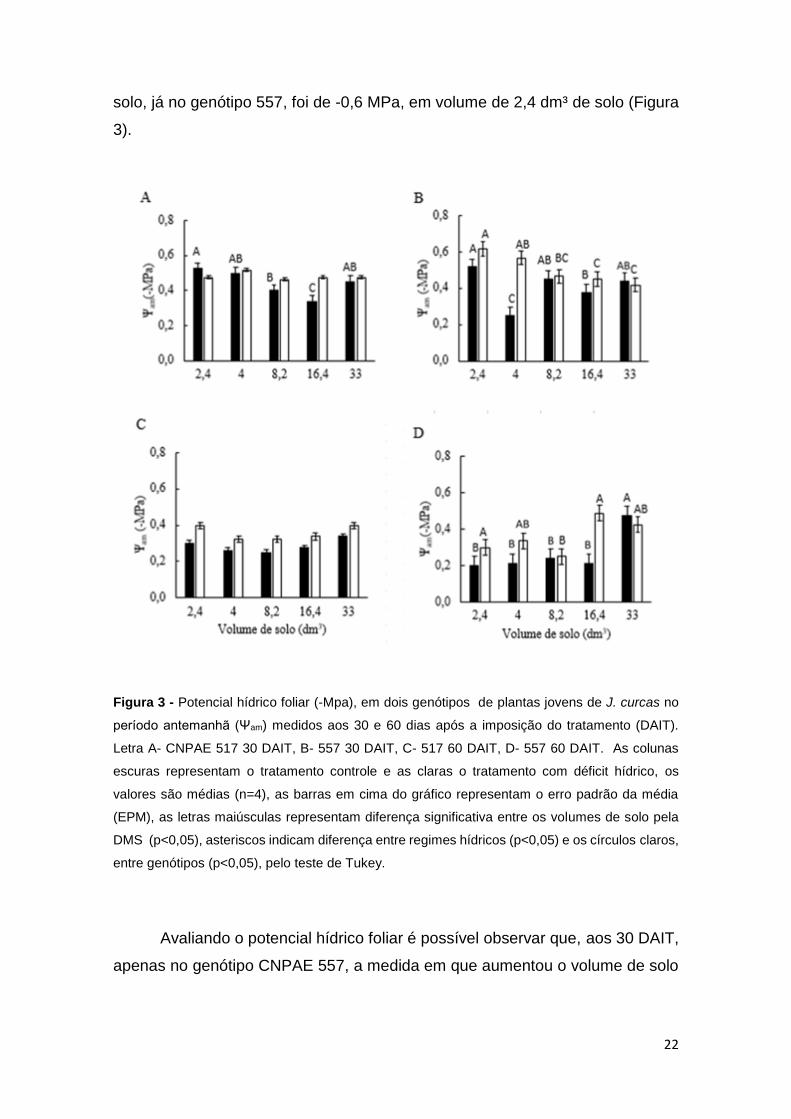
22
solo, já no genótipo 557, foi de -0,6 MPa, em volume de 2,4 dm³ de solo (Figura
3).
Figura 3 - Potencial hídrico foliar (-Mpa), em dois genótipos de plantas jovens de J. curcas no
período antemanhã (Ψam) medidos aos 30 e 60 dias após a imposição do tratamento (DAIT).
Letra A- CNPAE 517 30 DAIT, B- 557 30 DAIT, C- 517 60 DAIT, D- 557 60 DAIT. As colunas
escuras representam o tratamento controle e as claras o tratamento com déficit hídrico, os
valores são médias (n=4), as barras em cima do gráfico representam o erro padrão da média
(EPM), as letras maiúsculas representam diferença significativa entre os volumes de solo pela
DMS (p<0,05), asteriscos indicam diferença entre regimes hídricos (p<0,05) e os círculos claros,
entre genótipos (p<0,05), pelo teste de Tukey.
Avaliando o potencial hídrico foliar é possível observar que, aos 30 DAIT,
apenas no genótipo CNPAE 557, a medida em que aumentou o volume de solo
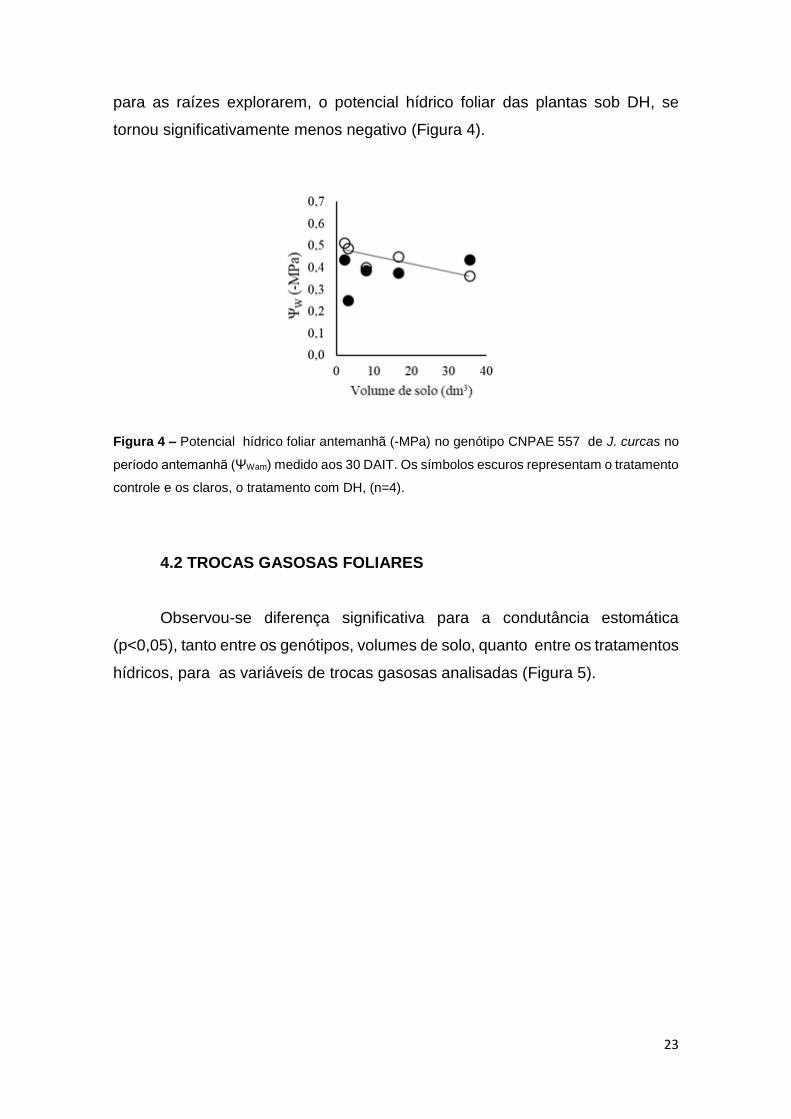
23
para as raízes explorarem, o potencial hídrico foliar das plantas sob DH, se
tornou significativamente menos negativo (Figura 4).
Figura 4 – Potencial hídrico foliar antemanhã (-MPa) no genótipo CNPAE 557 de J. curcas no
período antemanhã (ΨWam) medido aos 30 DAIT. Os símbolos escuros representam o tratamento
controle e os claros, o tratamento com DH, (n=4).
4.2 TROCAS GASOSAS FOLIARES
Observou-se diferença significativa para a condutância estomática
(p<0,05), tanto entre os genótipos, volumes de solo, quanto entre os tratamentos
hídricos, para as variáveis de trocas gasosas analisadas (Figura 5).
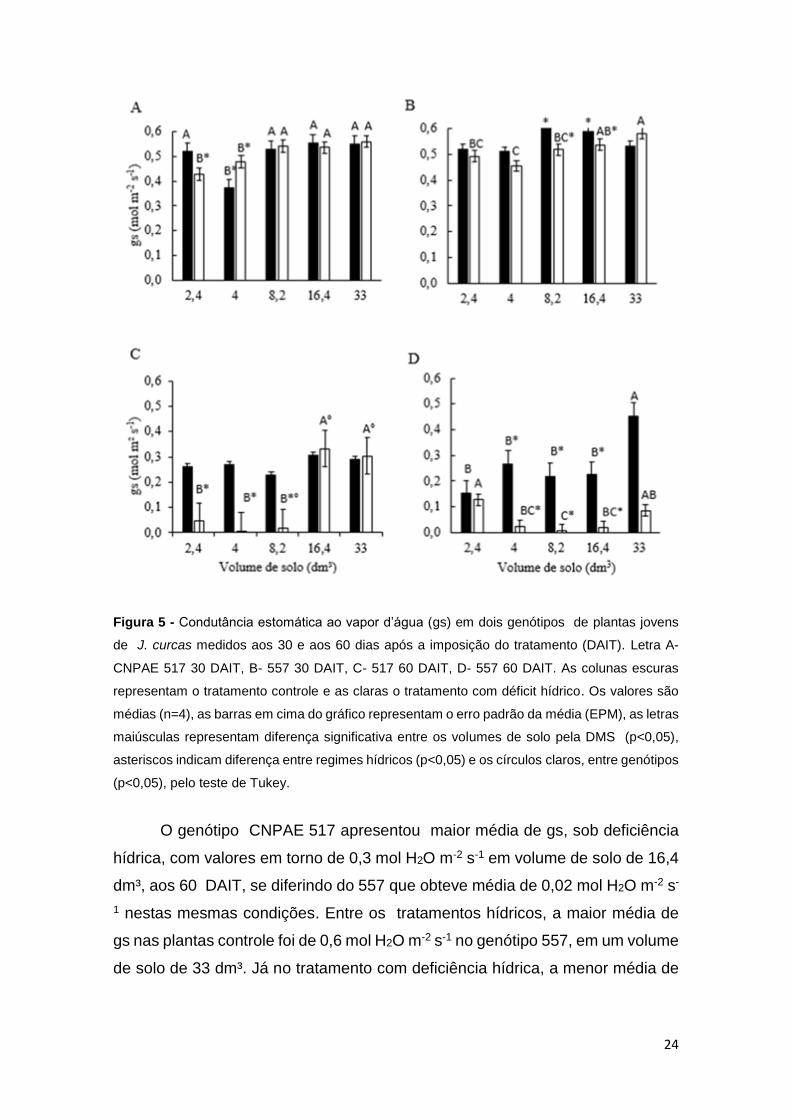
24
Figura 5 - Condutância estomática ao vapor d’água (gs) em dois genótipos de plantas jovens
de J. curcas medidos aos 30 e aos 60 dias após a imposição do tratamento (DAIT). Letra A-
CNPAE 517 30 DAIT, B- 557 30 DAIT, C- 517 60 DAIT, D- 557 60 DAIT. As colunas escuras
representam o tratamento controle e as claras o tratamento com déficit hídrico. Os valores são
médias (n=4), as barras em cima do gráfico representam o erro padrão da média (EPM), as letras
maiúsculas representam diferença significativa entre os volumes de solo pela DMS (p<0,05),
asteriscos indicam diferença entre regimes hídricos (p<0,05) e os círculos claros, entre genótipos
(p<0,05), pelo teste de Tukey.
O genótipo CNPAE 517 apresentou maior média de gs, sob deficiência
hídrica, com valores em torno de 0,3 mol H2O m-2 s-1 em volume de solo de 16,4
dm³, aos 60 DAIT, se diferindo do 557 que obteve média de 0,02 mol H2O m-2 s-
1 nestas mesmas condições. Entre os tratamentos hídricos, a maior média de
gs nas plantas controle foi de 0,6 mol H2O m-2 s-1 no genótipo 557, em um volume
de solo de 33 dm³. Já no tratamento com deficiência hídrica, a menor média de
25
gs foi em torno de 0,007 mol H2O m-2 s-1 no 517 aos 60 DAIT, em um volume de
solo de 4 dm³.
No tratamento com DH, as reduções nos valores de gs tanto para o
genótipo CNPAE 517, quanto o CNPAE 557, ocorreu de forma mais intensa nos
menores volumes de solo (2,4, 4 e 8,2 dm³) aos 60 DAIT, onde a gs decresceu
85%, 97% e 91% no genótipo 517 em relação as plantas controles, já no genótipo
557, essa diminuição, foi de 20%, 91% e 96% respectivamente.
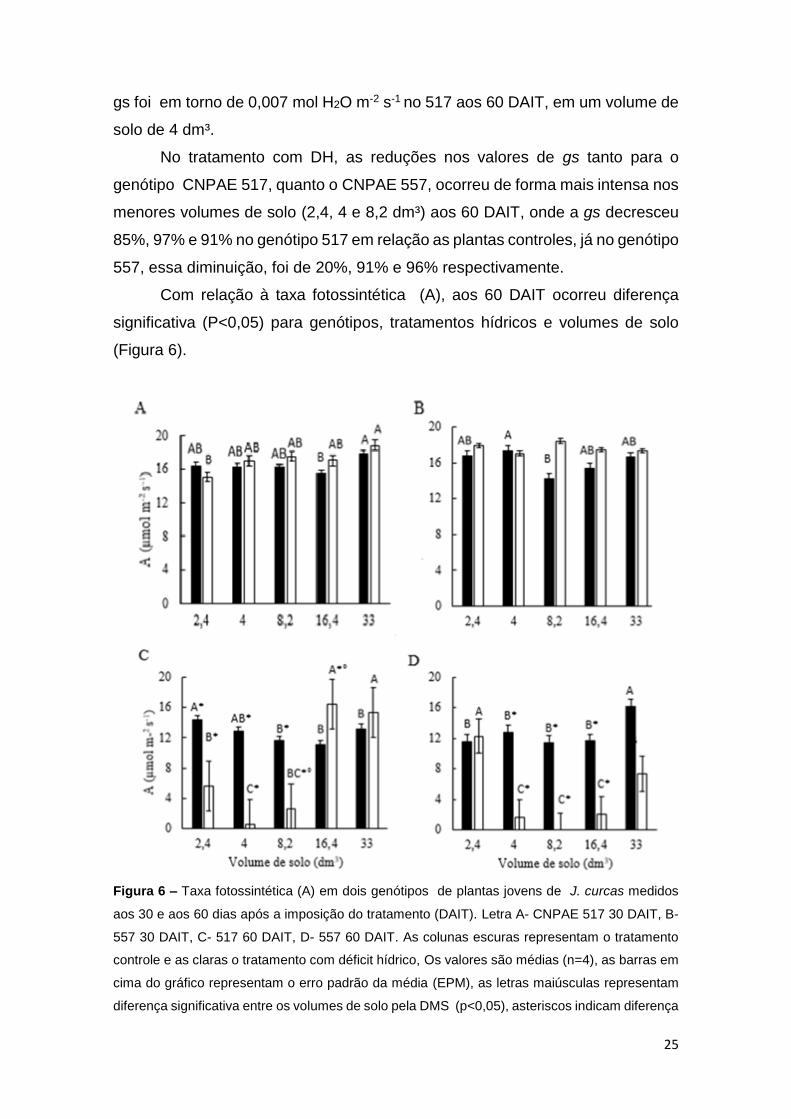
Com relação à taxa fotossintética (A), aos 60 DAIT ocorreu diferença
significativa (P<0,05) para genótipos, tratamentos hídricos e volumes de solo
(Figura 6).
Figura 6 – Taxa fotossintética (A) em dois genótipos de plantas jovens de J. curcas medidos
aos 30 e aos 60 dias após a imposição do tratamento (DAIT). Letra A- CNPAE 517 30 DAIT, B-
557 30 DAIT, C- 517 60 DAIT, D- 557 60 DAIT. As colunas escuras representam o tratamento
controle e as claras o tratamento com déficit hídrico, Os valores são médias (n=4), as barras em
cima do gráfico representam o erro padrão da média (EPM), as letras maiúsculas representam
diferença significativa entre os volumes de solo pela DMS (p<0,05), asteriscos indicam diferença

26
entre regimes hídricos (p<0,05) e os círculos claros, entre genótipos (p<0,05), pelo teste de
Tukey.
A diferença entre os genótipos foi observada apenas no tratamento com
DH, nos volumes 8,2 e 16,4 dm³ de solo, sendo que o genótipo CNPAE 517 foi
o que obteve maior média, com valor de 16 µmol m-2 s-1, em um volume de 16,4
dm³, enquanto que no 557, a média foi de 2 µmol m-2 s-1.
Entre tratamentos, nas plantas controle, verificou-se a maior média de A,
que foi de 16 µmol m-2 s-1, no genótipo 557, em um volume de 33 dm³ de solo,
e no tratamento com DH a maior média também ficou em torno de 16 µmol m-2
s-1, porém, no genótipo 517, em um volume de 16,4 dm³ de solo.
A diferença significativa nos valores de A entre os tratamentos hídricos
(p<0,05) ocorreu aos 60 DAIT para os dois genótipos, sendo que as plantas
submetidas à DH, do 517, sofreu um maior estresse em um volume de 4 dm³,
diminuindo em 95% em comparação com as controle. Já as plantas submetidas
à DH, do genótipo 557, apresentou a maior redução de PN em um volume de
8,2 dm³ de solo, com 100% a menos, quando comparadas com as plantas
controles.
Para os diferentes volumes de solo, desde os 30 DAIT, já foi possível
observar efeito significativo (p<0,05).
4.3 VARIÁVEIS DE CRESCIMENTO
Das figuras 7 a 24 são apresentadas as variáveis de crescimento (taxa de
crescimento relativo em altura e diâmetro, área foliar total, biomassa de raiz,
razão raiz/parte aérea, crescimento e densidade de comprimento radicular,
volume de raiz, área de superfície de raiz, diâmetro médio, e classes de
diâmetro de raízes), comparando os efeitos dos genótipos, regimes hídricos e
volumes de solo aos 30 e aos 60 DAIT.
Observou-se que o efeito do volume de solo foi significativo (P<0,05) para
todas as variáveis de crescimento analisadas em ambas as épocas de avaliação.
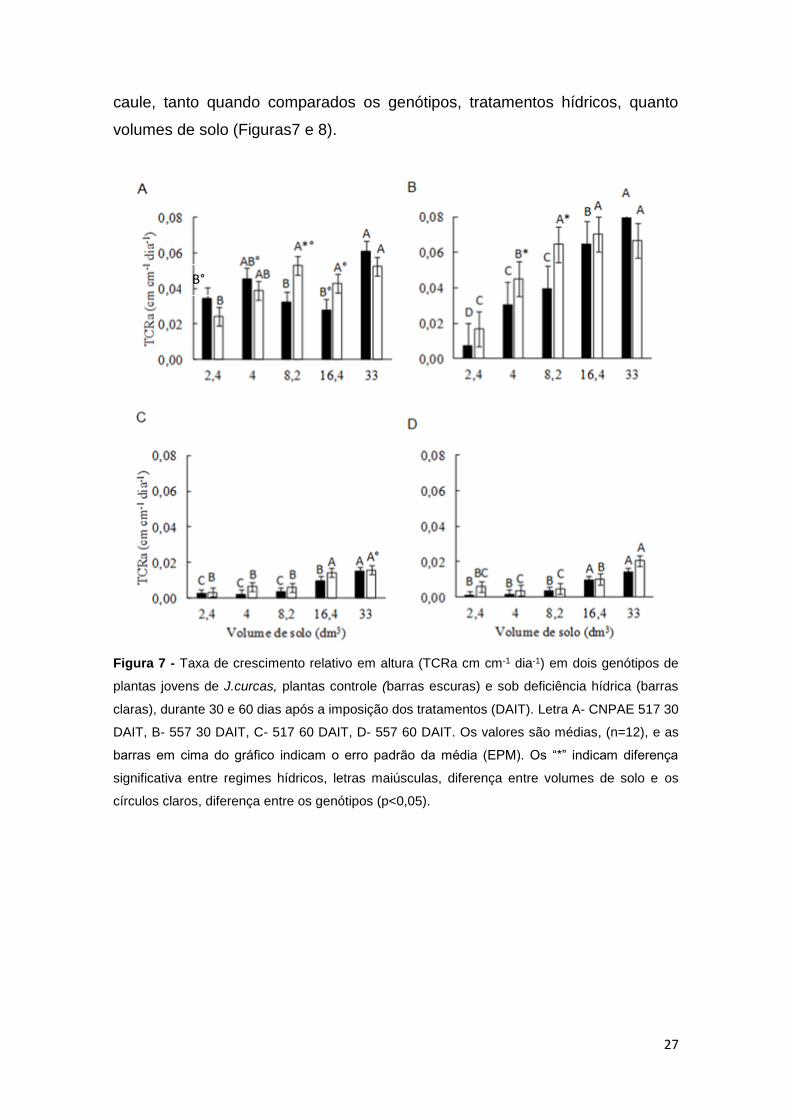
Aos 30 DAIT, já foi possível observar diferenças significativas (p<0,05),
para as variáveis de taxa de crescimento relativo de altura e de diâmetro do
27
caule, tanto quando comparados os genótipos, tratamentos hídricos, quanto
volumes de solo (Figuras7 e 8).
Figura 7 - Taxa de crescimento relativo em altura (TCRa cm cm-1 dia-1) em dois genótipos de
plantas jovens de J.curcas, plantas controle (barras escuras) e sob deficiência hídrica (barras
claras), durante 30 e 60 dias após a imposição dos tratamentos (DAIT). Letra A- CNPAE 517 30
DAIT, B- 557 30 DAIT, C- 517 60 DAIT, D- 557 60 DAIT. Os valores são médias, (n=12), e as
barras em cima do gráfico indicam o erro padrão da média (EPM). Os “*” indicam diferença
significativa entre regimes hídricos, letras maiúsculas, diferença entre volumes de solo e os
círculos claros, diferença entre os genótipos (p<0,05).
B°
28
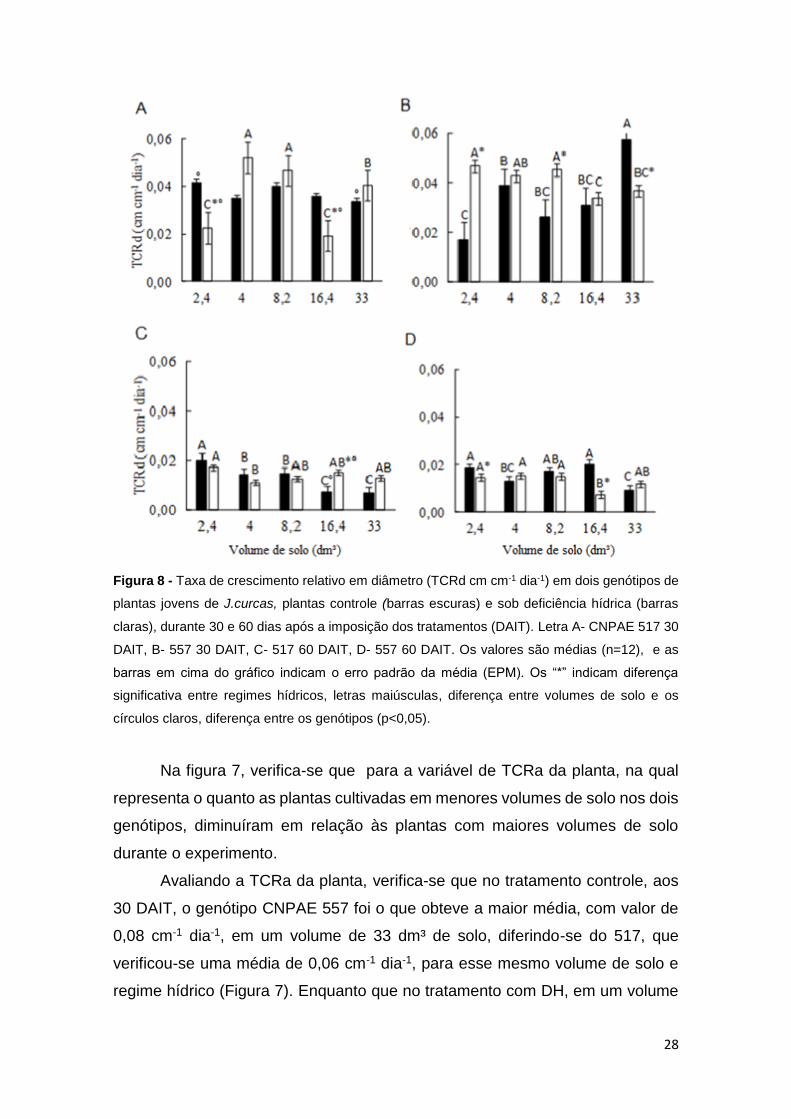
Figura 8 - Taxa de crescimento relativo em diâmetro (TCRd cm cm-1 dia-1) em dois genótipos de
plantas jovens de J.curcas, plantas controle (barras escuras) e sob deficiência hídrica (barras
claras), durante 30 e 60 dias após a imposição dos tratamentos (DAIT). Letra A- CNPAE 517 30
DAIT, B- 557 30 DAIT, C- 517 60 DAIT, D- 557 60 DAIT. Os valores são médias (n=12), e as
barras em cima do gráfico indicam o erro padrão da média (EPM). Os “*” indicam diferença
significativa entre regimes hídricos, letras maiúsculas, diferença entre volumes de solo e os
círculos claros, diferença entre os genótipos (p<0,05).
Na figura 7, verifica-se que para a variável de TCRa da planta, na qual
representa o quanto as plantas cultivadas em menores volumes de solo nos dois
genótipos, diminuíram em relação às plantas com maiores volumes de solo
durante o experimento.
Avaliando a TCRa da planta, verifica-se que no tratamento controle, aos
30 DAIT, o genótipo CNPAE 557 foi o que obteve a maior média, com valor de
0,08 cm-1 dia-1, em um volume de 33 dm³ de solo, diferindo-se do 517, que
verificou-se uma média de 0,06 cm-1 dia-1, para esse mesmo volume de solo e
regime hídrico (Figura 7). Enquanto que no tratamento com DH, em um volume

29
de solo de 2,4 dm³ o CNPAE 557 foi o genótipo que apresentou a menor média
observada para a TCRa com um valor de 0,016 cm-1 dia-1, já o 517 apresentou
0,024 cm-1 dia-1 de média, para este mesmo volume de solo e regime hídrico.
Aos 60 DAIT, os valores de ambos os genótipos, nos diferentes volumes de solo
e tratamentos hídricos, reduziram proporcionalmente, mantendo a maior TCRa
em plantas cultivadas em volume de solo de 33 dm³.
O genótipo CNPAE 557 foi o que apresentou a maior taxa de redução na
variável da TCRa com 91,25% a menos, em plantas controles, em um volume
de 2,4 dm³ de solo, em relação as plantas cultivadas em um volume de 33 dm³
de solo, aos 30 DAIT.
Na variável TCRd, para o tratamento controle, aos 30 DAIT, o CNPAE 557
foi o genótipo em que se observou a menor média, apresentando 0,017 cm cm-
1 dia-1 no volume de 2,4 dm³ de solo, em contrapartida, no 517, a maior média
foi de 0,033 cm cm-1 dia-1, no volume de 33 dm³ de solo. No tratamento com DH,
o genótipo com a menor média observada foi no CNPAE 517, com um valor de
0,019 cm cm-1 dia-1, já no 557, a menor média foi de 0,033 cm cm-1 dia-1, ambas,
no volume de 16,4 dm³ de solo.
Aos 60 DAIT, houve também, uma redução proporcional nas taxas de
TCRd para os diferentes volumes de solo, porém, ao contrário da TCRa, na
TCRd as maiores médias foram encontradas nos menores volumes de solo.
Quando comparados os tratamentos hídricos, aos 30 DAIT os valores da TCRa
da planta apresentou diferença (p<0,05) nos volumes de 8,2 dm³ de solo em
ambos genótipos, e também no de 4 dm³ de solo no 557, sendo que, aos 60
DAIT essa diferença deixou de existir. Já para a variável TCRd do caule o
CNPAE 517 e o 557 apresentaram diferenças significativas entre os tratamentos,
em alguns dos volumes de solo avaliados, tanto aos 30 como aos 60 DAIT
(Figuras 7 e 8).
Ainda com relação ao TCRd, verificou-se que, semelhantemente a TCRa,
o CNPAE 557 foi o genótipo que mais decresceu, com taxa de 70% de redução,
no volume de 2,4 dm³ em relação as plantas cultivadas em volume de 33 dm³
de solo, em plantas controle, aos 30 DAIT. Em oposição, ao avaliar as plantas
sob DH, o genótipo 517, foi o que apresentou um maior decréscimo, com 63,5%
de redução nas plantas cultivadas em um volume de 16,4 dm³ de solo, quando
30
comparadas as plantas cultivadas em 4 dm³ volume de solo, para este mesmo
regime hídrico e período de avaliação (Figura 8).
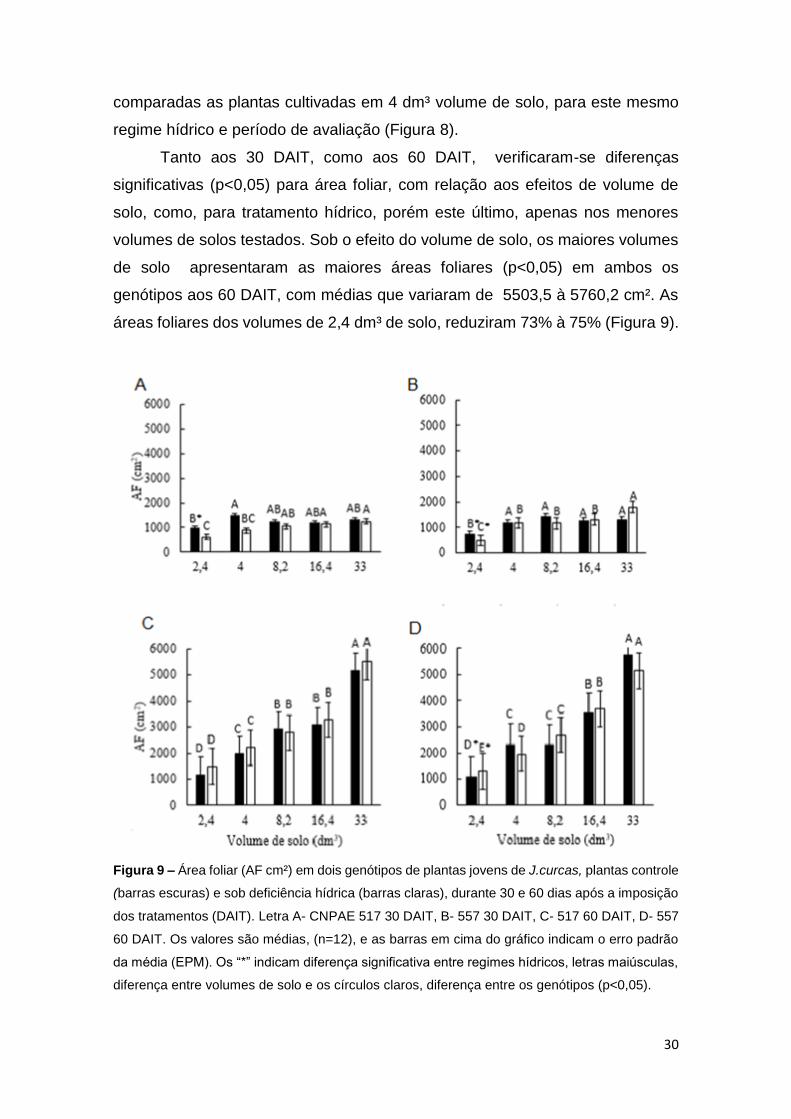
Tanto aos 30 DAIT, como aos 60 DAIT, verificaram-se diferenças
significativas (p<0,05) para área foliar, com relação aos efeitos de volume de
solo, como, para tratamento hídrico, porém este último, apenas nos menores
volumes de solos testados. Sob o efeito do volume de solo, os maiores volumes
de solo apresentaram as maiores áreas foliares (p<0,05) em ambos os
genótipos aos 60 DAIT, com médias que variaram de 5503,5 à 5760,2 cm². As
áreas foliares dos volumes de 2,4 dm³ de solo, reduziram 73% à 75% (Figura 9).
Figura 9 – Área foliar (AF cm²) em dois genótipos de plantas jovens de J.curcas, plantas controle
(barras escuras) e sob deficiência hídrica (barras claras), durante 30 e 60 dias após a imposição
dos tratamentos (DAIT). Letra A- CNPAE 517 30 DAIT, B- 557 30 DAIT, C- 517 60 DAIT, D- 557
60 DAIT. Os valores são médias, (n=12), e as barras em cima do gráfico indicam o erro padrão
da média (EPM). Os “*” indicam diferença significativa entre regimes hídricos, letras maiúsculas,
diferença entre volumes de solo e os círculos claros, diferença entre os genótipos (p<0,05).
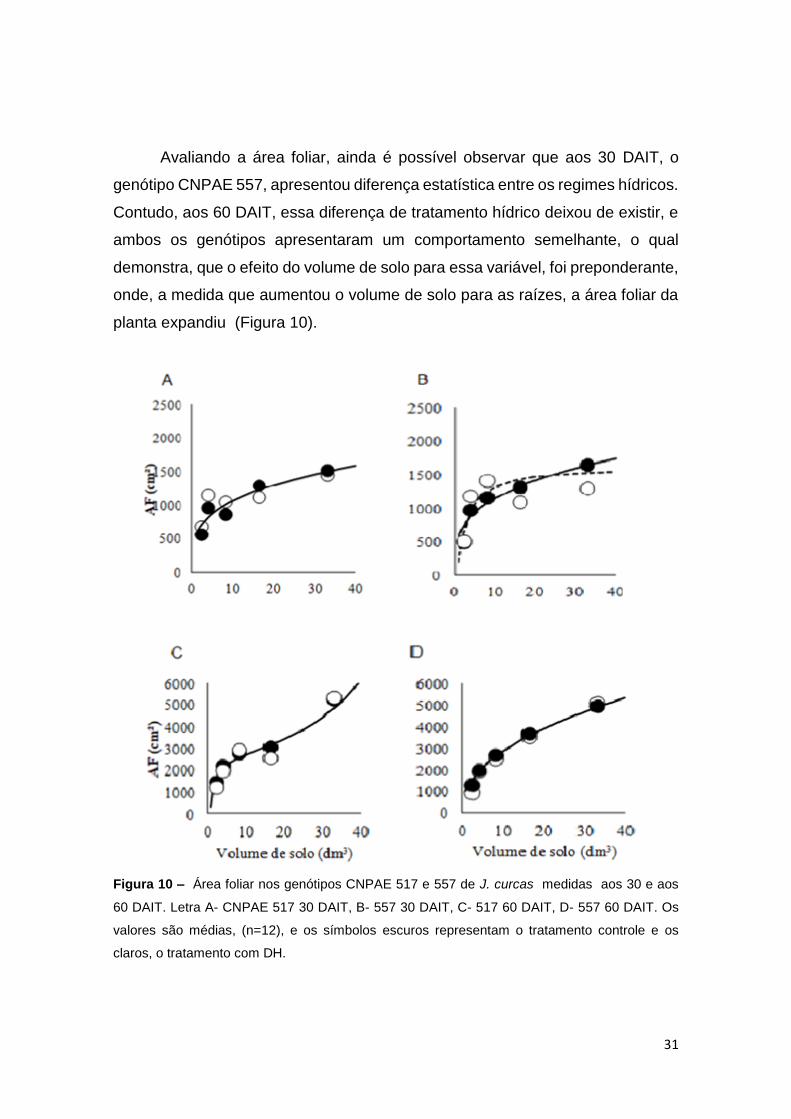
31
Avaliando a área foliar, ainda é possível observar que aos 30 DAIT, o
genótipo CNPAE 557, apresentou diferença estatística entre os regimes hídricos.
Contudo, aos 60 DAIT, essa diferença de tratamento hídrico deixou de existir, e
ambos os genótipos apresentaram um comportamento semelhante, o qual
demonstra, que o efeito do volume de solo para essa variável, foi preponderante,
onde, a medida que aumentou o volume de solo para as raízes, a área foliar da
planta expandiu (Figura 10).
Figura 10 – Área foliar nos genótipos CNPAE 517 e 557 de J. curcas medidas aos 30 e aos
60 DAIT. Letra A- CNPAE 517 30 DAIT, B- 557 30 DAIT, C- 517 60 DAIT, D- 557 60 DAIT. Os
valores são médias, (n=12), e os símbolos escuros representam o tratamento controle e os
claros, o tratamento com DH.
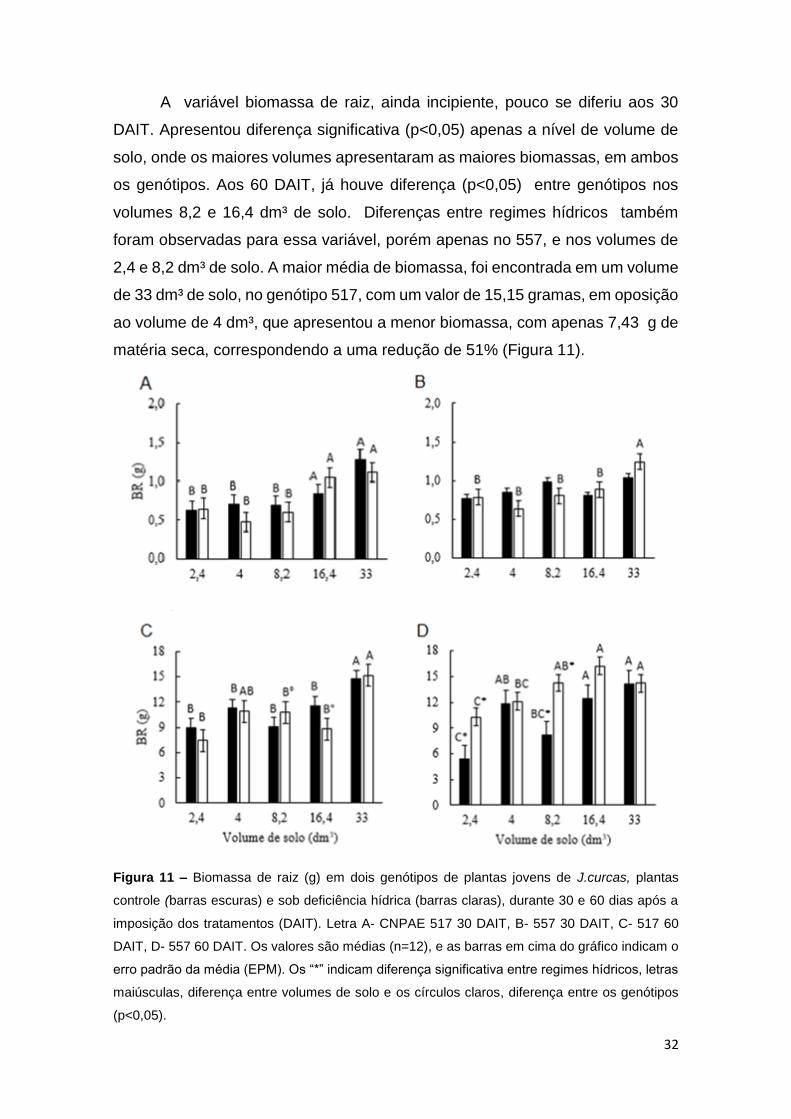
32
A variável biomassa de raiz, ainda incipiente, pouco se diferiu aos 30
DAIT. Apresentou diferença significativa (p<0,05) apenas a nível de volume de
solo, onde os maiores volumes apresentaram as maiores biomassas, em ambos
os genótipos. Aos 60 DAIT, já houve diferença (p<0,05) entre genótipos nos
volumes 8,2 e 16,4 dm³ de solo. Diferenças entre regimes hídricos também
foram observadas para essa variável, porém apenas no 557, e nos volumes de
2,4 e 8,2 dm³ de solo. A maior média de biomassa, foi encontrada em um volume
de 33 dm³ de solo, no genótipo 517, com um valor de 15,15 gramas, em oposição
ao volume de 4 dm³, que apresentou a menor biomassa, com apenas 7,43 g de
matéria seca, correspondendo a uma redução de 51% (Figura 11).
Figura 11 – Biomassa de raiz (g) em dois genótipos de plantas jovens de J.curcas, plantas
controle (barras escuras) e sob deficiência hídrica (barras claras), durante 30 e 60 dias após a
imposição dos tratamentos (DAIT). Letra A- CNPAE 517 30 DAIT, B- 557 30 DAIT, C- 517 60
DAIT, D- 557 60 DAIT. Os valores são médias (n=12), e as barras em cima do gráfico indicam o
erro padrão da média (EPM). Os “*” indicam diferença significativa entre regimes hídricos, letras
maiúsculas, diferença entre volumes de solo e os círculos claros, diferença entre os genótipos
(p<0,05).
a
33
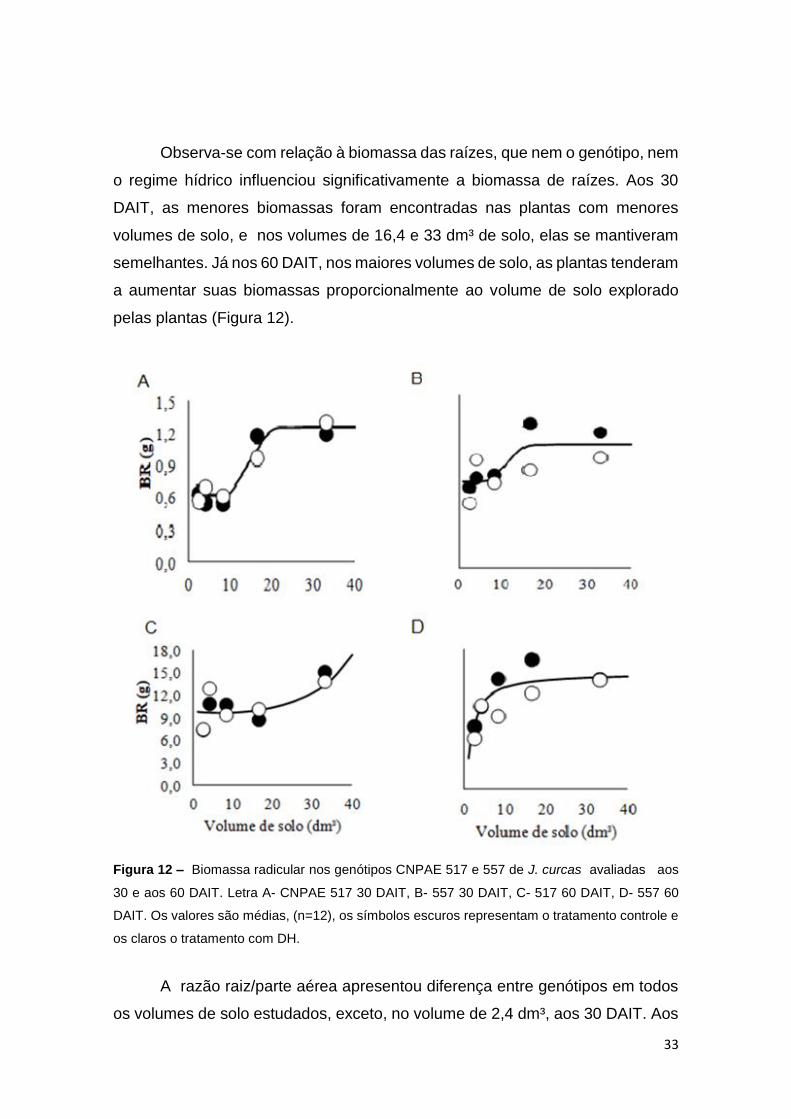
Observa-se com relação à biomassa das raízes, que nem o genótipo, nem
o regime hídrico influenciou significativamente a biomassa de raízes. Aos 30
DAIT, as menores biomassas foram encontradas nas plantas com menores
volumes de solo, e nos volumes de 16,4 e 33 dm³ de solo, elas se mantiveram
semelhantes. Já nos 60 DAIT, nos maiores volumes de solo, as plantas tenderam
a aumentar suas biomassas proporcionalmente ao volume de solo explorado
pelas plantas (Figura 12).
Figura 12 – Biomassa radicular nos genótipos CNPAE 517 e 557 de J. curcas avaliadas aos
30 e aos 60 DAIT. Letra A- CNPAE 517 30 DAIT, B- 557 30 DAIT, C- 517 60 DAIT, D- 557 60
DAIT. Os valores são médias, (n=12), os símbolos escuros representam o tratamento controle e
os claros o tratamento com DH.
A razão raiz/parte aérea apresentou diferença entre genótipos em todos
os volumes de solo estudados, exceto, no volume de 2,4 dm³, aos 30 DAIT. Aos
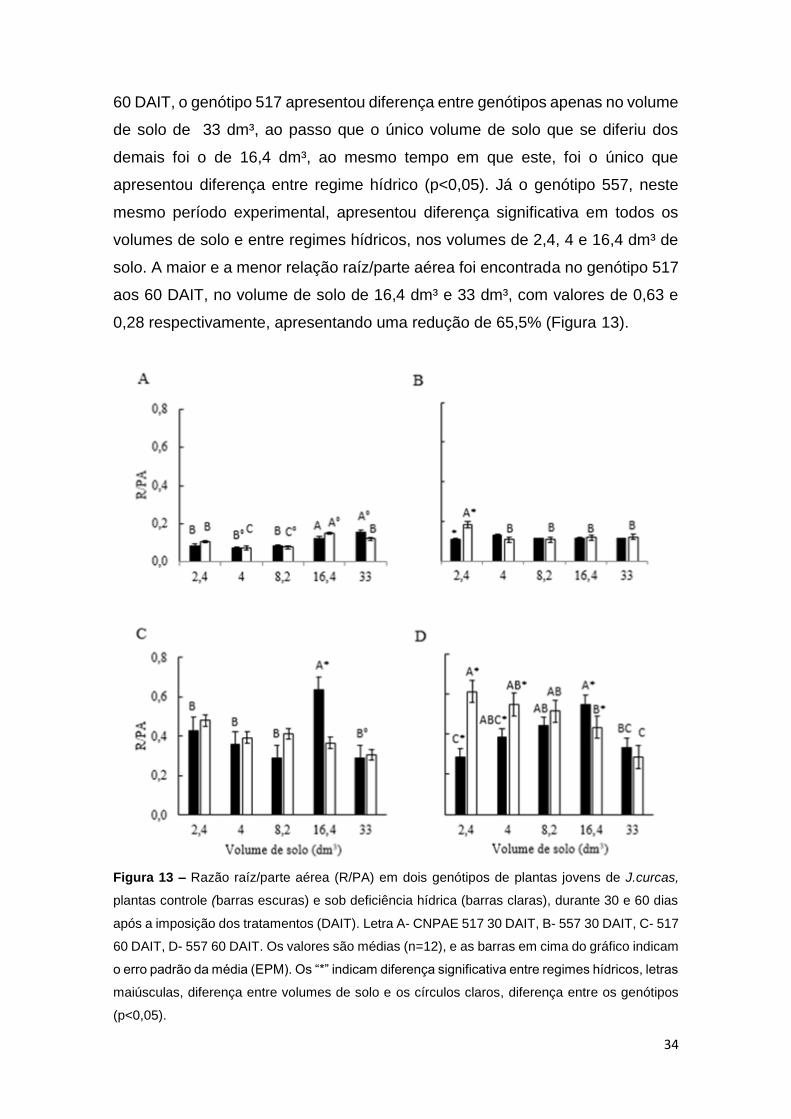
34
60 DAIT, o genótipo 517 apresentou diferença entre genótipos apenas no volume
de solo de 33 dm³, ao passo que o único volume de solo que se diferiu dos
demais foi o de 16,4 dm³, ao mesmo tempo em que este, foi o único que
apresentou diferença entre regime hídrico (p<0,05). Já o genótipo 557, neste
mesmo período experimental, apresentou diferença significativa em todos os
volumes de solo e entre regimes hídricos, nos volumes de 2,4, 4 e 16,4 dm³ de
solo. A maior e a menor relação raíz/parte aérea foi encontrada no genótipo 517
aos 60 DAIT, no volume de solo de 16,4 dm³ e 33 dm³, com valores de 0,63 e
0,28 respectivamente, apresentando uma redução de 65,5% (Figura 13).
Figura 13 – Razão raíz/parte aérea (R/PA) em dois genótipos de plantas jovens de J.curcas,
plantas controle (barras escuras) e sob deficiência hídrica (barras claras), durante 30 e 60 dias
após a imposição dos tratamentos (DAIT). Letra A- CNPAE 517 30 DAIT, B- 557 30 DAIT, C- 517
60 DAIT, D- 557 60 DAIT. Os valores são médias (n=12), e as barras em cima do gráfico indicam
o erro padrão da média (EPM). Os “*” indicam diferença significativa entre regimes hídricos, letras
maiúsculas, diferença entre volumes de solo e os círculos claros, diferença entre os genótipos
(p<0,05).
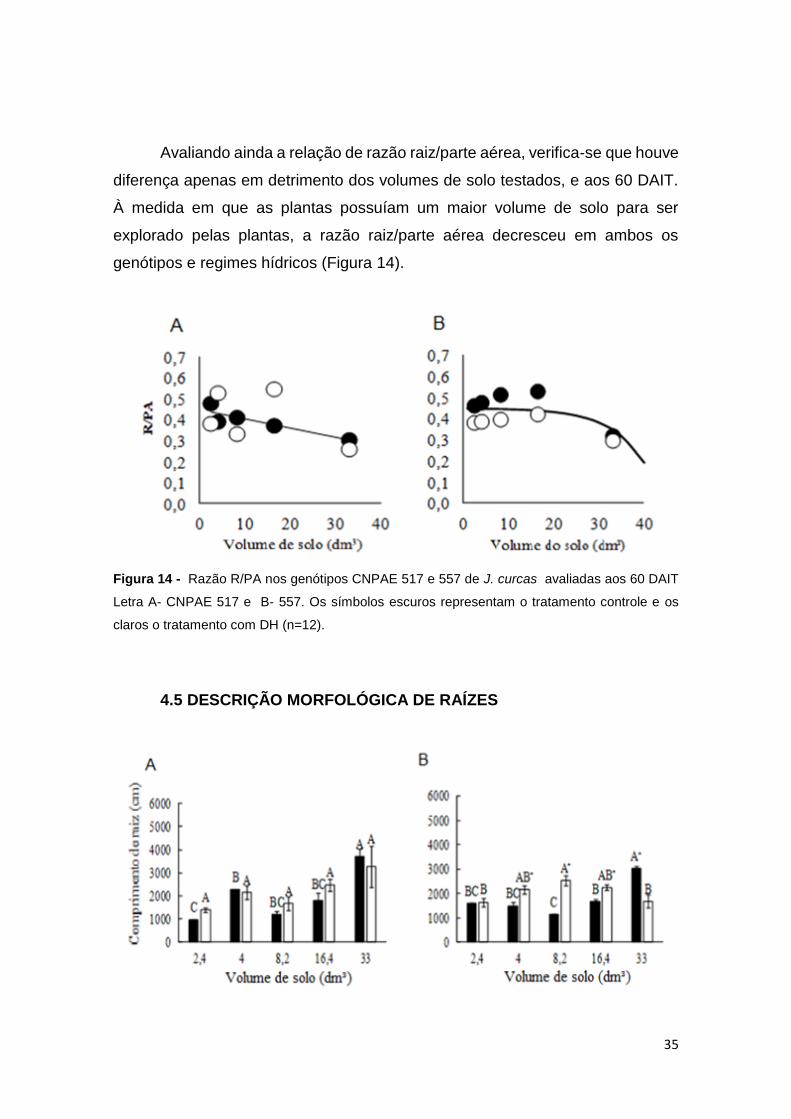
35
Avaliando ainda a relação de razão raiz/parte aérea, verifica-se que houve
diferença apenas em detrimento dos volumes de solo testados, e aos 60 DAIT.
À medida em que as plantas possuíam um maior volume de solo para ser
explorado pelas plantas, a razão raiz/parte aérea decresceu em ambos os
genótipos e regimes hídricos (Figura 14).
Figura 14 - Razão R/PA nos genótipos CNPAE 517 e 557 de J. curcas avaliadas aos 60 DAIT
Letra A- CNPAE 517 e B- 557. Os símbolos escuros representam o tratamento controle e os
claros o tratamento com DH (n=12).
4.5 DESCRIÇÃO MORFOLÓGICA DE RAÍZES
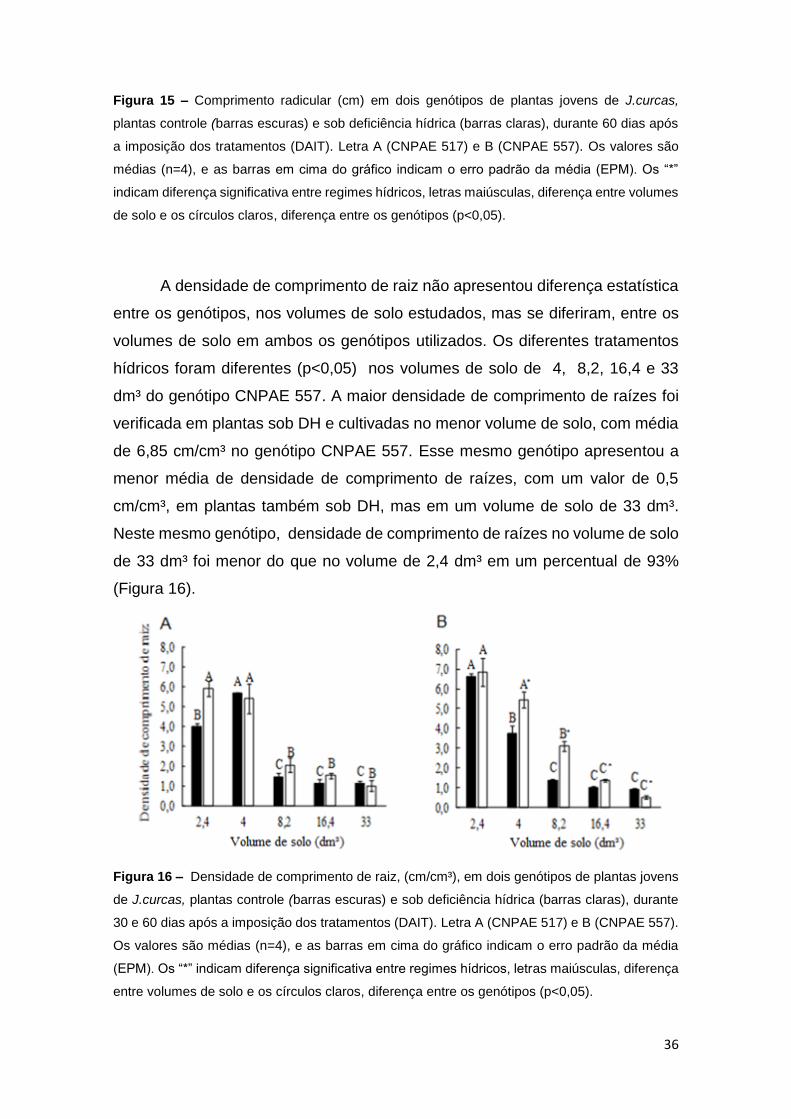
36
Figura 15 – Comprimento radicular (cm) em dois genótipos de plantas jovens de J.curcas,
plantas controle (barras escuras) e sob deficiência hídrica (barras claras), durante 60 dias após
a imposição dos tratamentos (DAIT). Letra A (CNPAE 517) e B (CNPAE 557). Os valores são
médias (n=4), e as barras em cima do gráfico indicam o erro padrão da média (EPM). Os “*”
indicam diferença significativa entre regimes hídricos, letras maiúsculas, diferença entre volumes
de solo e os círculos claros, diferença entre os genótipos (p<0,05).
A densidade de comprimento de raiz não apresentou diferença estatística
entre os genótipos, nos volumes de solo estudados, mas se diferiram, entre os
volumes de solo em ambos os genótipos utilizados. Os diferentes tratamentos
hídricos foram diferentes (p<0,05) nos volumes de solo de 4, 8,2, 16,4 e 33
dm³ do genótipo CNPAE 557. A maior densidade de comprimento de raízes foi
verificada em plantas sob DH e cultivadas no menor volume de solo, com média
de 6,85 cm/cm³ no genótipo CNPAE 557. Esse mesmo genótipo apresentou a
menor média de densidade de comprimento de raízes, com um valor de 0,5
cm/cm³, em plantas também sob DH, mas em um volume de solo de 33 dm³.
Neste mesmo genótipo, densidade de comprimento de raízes no volume de solo
de 33 dm³ foi menor do que no volume de 2,4 dm³ em um percentual de 93%
(Figura 16).
Figura 16 – Densidade de comprimento de raiz, (cm/cm³), em dois genótipos de plantas jovens
de J.curcas, plantas controle (barras escuras) e sob deficiência hídrica (barras claras), durante
30 e 60 dias após a imposição dos tratamentos (DAIT). Letra A (CNPAE 517) e B (CNPAE 557).
Os valores são médias (n=4), e as barras em cima do gráfico indicam o erro padrão da média
(EPM). Os “*” indicam diferença significativa entre regimes hídricos, letras maiúsculas, diferença
entre volumes de solo e os círculos claros, diferença entre os genótipos (p<0,05).
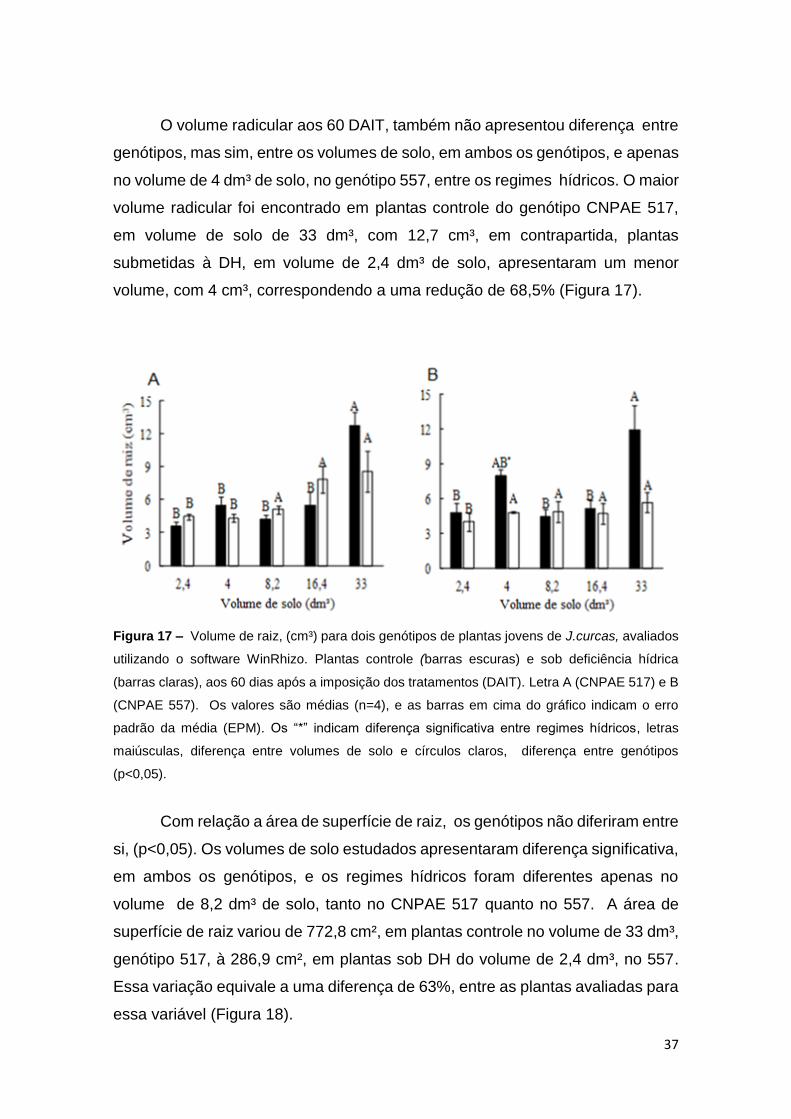
37
O volume radicular aos 60 DAIT, também não apresentou diferença entre
genótipos, mas sim, entre os volumes de solo, em ambos os genótipos, e apenas
no volume de 4 dm³ de solo, no genótipo 557, entre os regimes hídricos. O maior
volume radicular foi encontrado em plantas controle do genótipo CNPAE 517,
em volume de solo de 33 dm³, com 12,7 cm³, em contrapartida, plantas
submetidas à DH, em volume de 2,4 dm³ de solo, apresentaram um menor
volume, com 4 cm³, correspondendo a uma redução de 68,5% (Figura 17).
Figura 17 – Volume de raiz, (cm³) para dois genótipos de plantas jovens de J.curcas, avaliados
utilizando o software WinRhizo. Plantas controle (barras escuras) e sob deficiência hídrica
(barras claras), aos 60 dias após a imposição dos tratamentos (DAIT). Letra A (CNPAE 517) e B
(CNPAE 557). Os valores são médias (n=4), e as barras em cima do gráfico indicam o erro
padrão da média (EPM). Os “*” indicam diferença significativa entre regimes hídricos, letras
maiúsculas, diferença entre volumes de solo e círculos claros, diferença entre genótipos
(p<0,05).
Com relação a área de superfície de raiz, os genótipos não diferiram entre
si, (p<0,05). Os volumes de solo estudados apresentaram diferença significativa,
em ambos os genótipos, e os regimes hídricos foram diferentes apenas no
volume de 8,2 dm³ de solo, tanto no CNPAE 517 quanto no 557. A área de
superfície de raiz variou de 772,8 cm², em plantas controle no volume de 33 dm³,
genótipo 517, à 286,9 cm², em plantas sob DH do volume de 2,4 dm³, no 557.
Essa variação equivale a uma diferença de 63%, entre as plantas avaliadas para
essa variável (Figura 18).
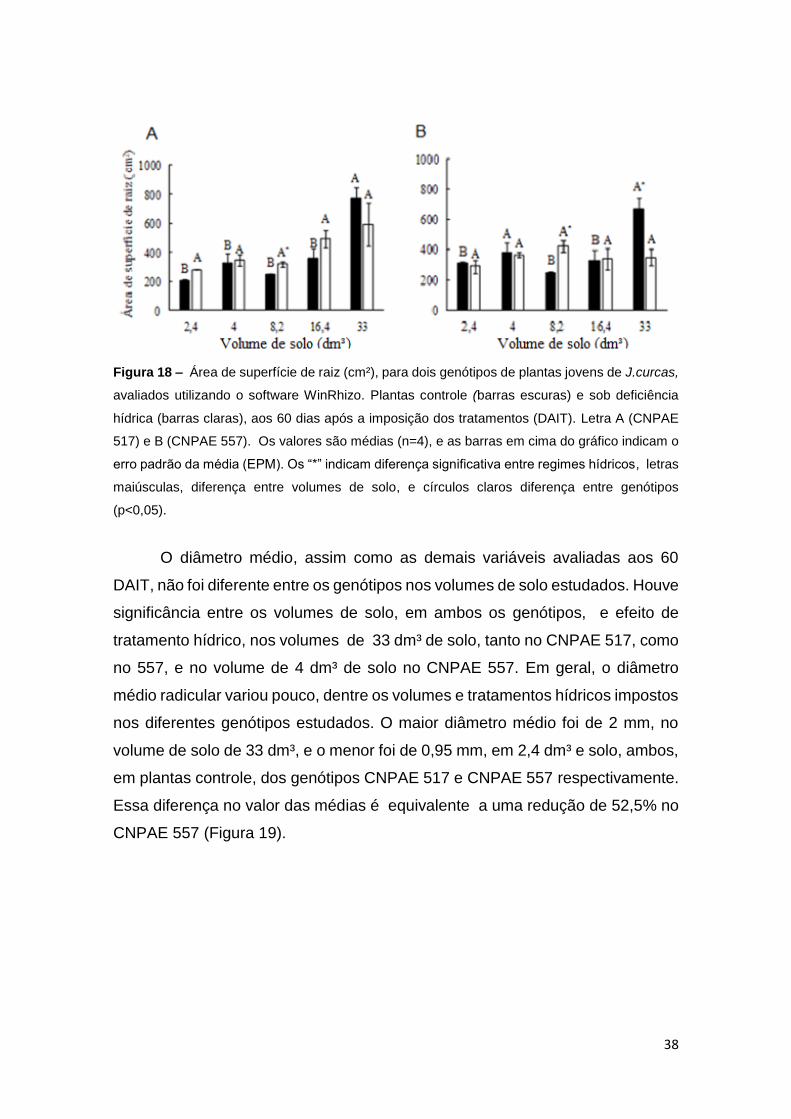
38
Figura 18 – Área de superfície de raiz (cm²), para dois genótipos de plantas jovens de J.curcas,
avaliados utilizando o software WinRhizo. Plantas controle (barras escuras) e sob deficiência
hídrica (barras claras), aos 60 dias após a imposição dos tratamentos (DAIT). Letra A (CNPAE
517) e B (CNPAE 557). Os valores são médias (n=4), e as barras em cima do gráfico indicam o
erro padrão da média (EPM). Os “*” indicam diferença significativa entre regimes hídricos, letras
maiúsculas, diferença entre volumes de solo, e círculos claros diferença entre genótipos
(p<0,05).
O diâmetro médio, assim como as demais variáveis avaliadas aos 60
DAIT, não foi diferente entre os genótipos nos volumes de solo estudados. Houve
significância entre os volumes de solo, em ambos os genótipos, e efeito de
tratamento hídrico, nos volumes de 33 dm³ de solo, tanto no CNPAE 517, como
no 557, e no volume de 4 dm³ de solo no CNPAE 557. Em geral, o diâmetro
médio radicular variou pouco, dentre os volumes e tratamentos hídricos impostos
nos diferentes genótipos estudados. O maior diâmetro médio foi de 2 mm, no
volume de solo de 33 dm³, e o menor foi de 0,95 mm, em 2,4 dm³ e solo, ambos,
em plantas controle, dos genótipos CNPAE 517 e CNPAE 557 respectivamente.
Essa diferença no valor das médias é equivalente a uma redução de 52,5% no
CNPAE 557 (Figura 19).
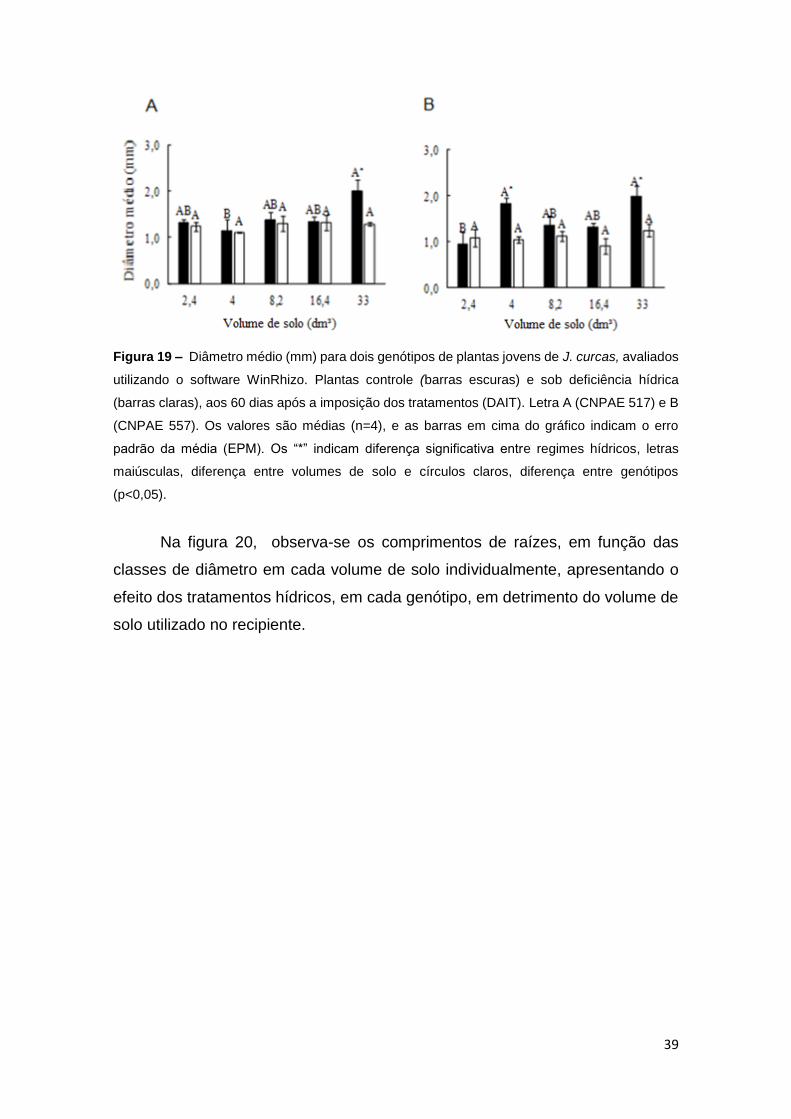
39
Figura 19 – Diâmetro médio (mm) para dois genótipos de plantas jovens de J. curcas, avaliados
utilizando o software WinRhizo. Plantas controle (barras escuras) e sob deficiência hídrica
(barras claras), aos 60 dias após a imposição dos tratamentos (DAIT). Letra A (CNPAE 517) e B
(CNPAE 557). Os valores são médias (n=4), e as barras em cima do gráfico indicam o erro
padrão da média (EPM). Os “*” indicam diferença significativa entre regimes hídricos, letras
maiúsculas, diferença entre volumes de solo e círculos claros, diferença entre genótipos
(p<0,05).
Na figura 20, observa-se os comprimentos de raízes, em função das
classes de diâmetro em cada volume de solo individualmente, apresentando o
efeito dos tratamentos hídricos, em cada genótipo, em detrimento do volume de
solo utilizado no recipiente.
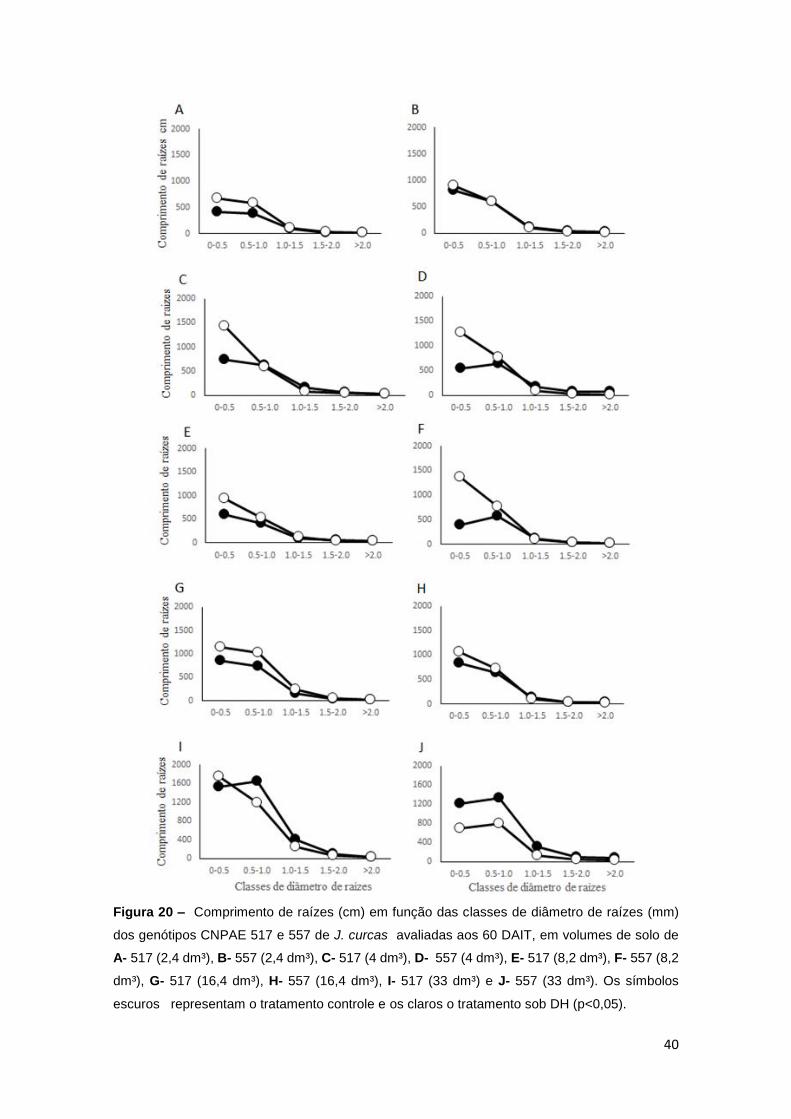
40
Figura 20 – Comprimento de raízes (cm) em função das classes de diâmetro de raízes (mm)
dos genótipos CNPAE 517 e 557 de J. curcas avaliadas aos 60 DAIT, em volumes de solo de
A- 517 (2,4 dm³), B- 557 (2,4 dm³), C- 517 (4 dm³), D- 557 (4 dm³), E- 517 (8,2 dm³), F- 557 (8,2
dm³), G- 517 (16,4 dm³), H- 557 (16,4 dm³), I- 517 (33 dm³) e J- 557 (33 dm³). Os símbolos
escuros representam o tratamento controle e os claros o tratamento sob DH (p<0,05).

41
Em ambos os genótipos, houve efeito significativo dos regimes hídricos
impostos, em todos os volumes de solo estudados (Figuras 21 à 24). Os
tratamentos hídricos se diferiram, principalmente, nas menores classes de
diâmetro de raízes, e nos volumes de 2,4 a 16,4 dm³ de solo, onde as plantas
submetidas à DH tenderam a aumentar o comprimento de suas raízes (cm), em
classes de diâmetro do menor tamanho ao tamanho intermediário, em relação
às plantas controles. Apenas no maior volume de solo, 33 dm³, não foi verificado
esse comportamento.
Figura 21. Raízes de plantas jovens de J. curcas em condição de DH aos 30 DAIT. A- 2,4 dm³,
B- 4 dm³, C- 8,2 dm³, D- 16,4 dm³ e E- 33 dm³.

42
Figura 22. Raízes de plantas jovens de J. curcas sob condições controle aos 30 DAIT. A- 2,4
dm³, B- 4 dm³, C- 8,2 dm³, D- 16,4 dm³ e E- 33 dm³.

43
Figura 23. Raízes de plantas jovens de J. curcas em condições de DH aos 60 DAIT. A- 2,4
dm³, B- 4 dm³, C- 8,2 dm³, D- 16,4 dm³ e E- 33 dm³.
B
44
Figura 24. Raízes de plantas jovens de J. curcas em condições controle aos 60 DAIT. A- 2,4
dm³, B- 4 dm³, C- 8,2 dm³, D- 16,4 dm³ e E- 33 dm³.

45
5. DISCUSSÃO
Como verificado, o CNPAE 517 não apresentou diferença (p<0,05) aos
60 DAIT entre os volumes de solo, mostrando-se possuir uma maior regulação
para o potencial hídrico foliar. Este resultado é diferente do obtido por Tiwari et
al. (2013), onde estresse hídrico causou uma redução no potencial hídrico foliar
em plantas jovens de J. curcas cultivadas por 60 dias, com um potencial que
variou de -0,49 MPa nas plantas controles à -1,95 MPa nas plantas sob
deficiência hídrica.
Moura et al. (2016), estudando relações hídricas em J. curcas,
observaram reduções semelhantes no Ψw ante manhã, quando submetidas à
deficiência hídrica de 40% da CC do solo e 60 dias de tratamento, em vasos de
11 kg de solo. Segundo Chaves et al., (2009), plantas submetidas ao déficit
hídrico, ocorre uma redução de Ψw que irá levar a um maior fechamento
estomático, a fim de reduzir a perda de água pela da transpiração. Com a menor
condutância estomática, a resistência ao fluxo de CO2 para as folhas aumenta,
consequentemente a fotossíntese diminui.
Durante o período experimental, foi possível observar que dentre as
variáveis de trocas gasosas foliares nos dois genótipos de J. curcas aqui
estudados, a diminuição da condutância estomática gs é o primeiro sinal de
resposta da planta quando submetida à deficiência hídrica (Figura 5). Essa
redução de gs ocorre devido ao fechamento estomático, para evitar maiores
perdas de água pela transpiração.
A grande diminuição na condutância estomática observada também,
inclusive nas plantas controle, no presente experimento, pode ter sido
proveniente da transição do período climático a qual as plantas foram expostas,
quando foram cultivadas.
Diferente dos resultados obtidos no presente estudo, De SANTANA et al.
(2015), estudando três acessos de J. curcas (CNPAE 126, CNPAE 137 e CNPAE
139), observaram diferença significativa (p <0,05) entre os tratamentos de
regime hídrico para as variáveis de trocas gasosas foliares após 55 DAIT. Esses
autores relataram que não houve diferença estatística entre os acessos

46
estudados por eles. Após este período, a gs diminuiu à medida que o tempo de
exposição à deficiência hídrica aumentou. Os valores médios de gs foram de
0,35 e 0,03 mol de H2O m-2 s-1 nas plantas controles e sob DH, respectivamente,
valores diferentes dos encontrados no presente estudo. Essa discrepância entre
esses resultados pode ser devida dentre outros fatores, o tipo de solo usado, o
efeito dos diferentes volumes de solo testados neste trabalho (50 dm³), além do
tempo de exposição à deficiência hídrica (De SANTANA et al. 2015).
Com relação a variável taxa fotossintética, os autores Sousa et al. (2012),
utilizando o tratamento de 50% da CC, encontraram reduções em torno de 70%
da fotossíntese em relação ao controle. Os diferentes resultados encontrados
por esses autores, em relação aos obtidos no presente estudo, podem ser
associados às condições edafoclimáticas de cada região, o volume de solo
utilizado, o déficit de pressão de vapor e a temperatura, assim como, o estado
nutricional e os fatores genéticos que também podem influenciar nas
características fisiológicas da planta.
Para as variáveis de crescimento TCRa, TCRd e AF, quando submetidas
aos maiores volumes de solo e disponibilidade hídrica, observou-se um aumento
nas médias de plantas de J. curcas nos diferentes genótipos, comparadas às
plantas sob DH. Para a TCRa da planta, o decréscimo nas plantas submetidas
à DH variou entre 60 e 80% em comparação com as plantas controles nos
diferentes volumes de solo e genótipos, enquanto que a TCRd do caule sofreu
uma menor variação, apresentando valores de redução de 64 à 42%. Essa
menor redução na TCRd, pode ser decorrente do caule suculento que plantas
de J. curcas apresentam, como estratégia de sobrevivência sob DH. Outro
motivo para justificar esses valores na TCRd pode ser explicado, porque as
plantas controle em períodos mais quentes do dia, transpiram mais do que às
submetidas à DH, que fecham os estômatos para evitar a perda de água.
Verma et al. (2012), observaram ao final de 60 dias do experimento, com
diferentes níveis de água no solo (75, 50 e 25% da CC), que as variáveis altura,
diâmetro, número de folhas, expansão da área foliar e biomassa, são afetadas
pelo aumento do déficit hídrico em comparação com às plantas controles (100%
da CC). Com 25% da capacidade de campo, a altura da planta e o diâmetro do
coleto decresceu 39 e 40%. Também diminuiu o número de folhas, expansão da
área foliar e área foliar específica, com 44% 55% e 40% respectivamente,

47
juntamente com a perda de produção de biomassa total de 33% em relação às
plantas controles.
Em mamona, oleaginosa pertencente à mesma família botância de J.
Curcas, o volume de solo contido no recipiente, assim como no presente
trabalho, também influenciou as variáveis área foliar, biomassa aérea e das
raízes (LIMA et al., 2006).
Em um estudo realizado por Díaz-López et al. (2012), esses autores
observaram que a razão raiz/parte aérea, em J. curcas aumentou com o
acréscimo do deficit hídrico do solo, já que, enquanto a massa seca de raiz só
diferenciou do controle no momento em que houve corte total da irrigação, a
massa seca foliar diminuiu significativamente (p<0,05) a partir da imposição de
deficiência hídrica de 50% da capacidade de campo, sendo que esses autores
testaram 100, 75, 50, 25 e 0% da capacidade de campo no solo.
Cavatte et al. (2008), realizando estudos com café, consideram a
capacidade de manutenção da hidratação celular como uma característica dos
genótipos mais adaptados à restrição de água, quando comparados aos
genótipos intolerantes ao estresse hídrico. Semelhante aos resultados obtidos
por esses autores, no presente estudo, observou-se que os genótipos
apresentaram influência significativa sobre a manutenção hídrica, em algumas
variáveis fisiológicas aqui estudadas, todavia, os tratamentos hídricos e
sobretudo, os volumes de solo utilizados no cultivo das plantas, foram
responsáveis pelas alterações de água nas células, conduzindo as respostas
das plantas, aos estresses abióticos simultâneos nelas impostos.
Sob condições ótimas ou estressantes, o crescimento vegetal pode ser afetado por
vários fatores, incluindo a anatomia, a arquitetura e a condutividade hidráulica do sistema
radicular (MARTÍNEZ-ALCÂNTARA et al., 2013). No presente estudo, sugere-se que a
manutenção do crescimento radicular foi uma estratégia das plantas para se manterem em
condições de déficit de água no solo. Evidências anteriores constataram aumentos significativos
na biomassa da raiz de Citros submetidos ao estresse por seca (PEDROSO et al., 2014).
Corroborando aos autores, nossos resultados demonstraram que a seca pode aumentar a
quantidade de raízes finas, bem como, o comprimento total da raiz, volume, área e densidade
do comprimento (Figuras 20 à 24).
Após 60 dias da imposição de DH, houve um aumento no comprimento
de raízes das plantas estressadas quando comparadas às plantas controle,

48
consequente à isso, o comprimento de diferentes classes de diâmetro foi
observado em maiores proporções nas plantas cultivadas em menores volumes
de solo e sob disponibilidade hídrica moderada.
De acordo com Taiz e Zeiger, (2009) as plantas sob estresse utilizam esse
mecanismo de aumentar o comprimento de suas raízes para explorar um maior
volume de solo e evitar a desidratação (Figura 15). Essa reação é conhecida
como um tipo de estratégia de “tolerância” à seca para deficiência hídrica
moderada, quando as plantas fecham os estômatos e aumentam o volume do
sistema radicular.
Segundo Correia e Nogueira (2004) um menor teor de umidade disponível
no solo para as plantas causa uma redução na alocação de biomassa das folhas
e caules, aumentando a raíz.
O aumento significativo de raízes finas (diâmetro <0,5 mm) em plantas
sob DH, pode ter contribuído para manter o crescimento, uma vez que as raízes
finas são a principal via de absorção da água e nutrientes dissolvidos (JACKSON
et al., 2000). Plantas naturalmente sob condições de campo apresentam um
grande número de raízes finas (diâmetro <1 mm), que rapidamente se renovam
entre um e dez dias, e elas dependem da frequência da chuva para crescer (DA
SILVA; KUMMEROW, 1999). Além disso, o crescimento das plantas pode sofrer
interferência da dinâmica de crescimento e renovação das raízes, entre outros
fatores, como a forma de propagação, que podem reduzir a translocação de água
e nutrientes (MARTÍNEZ-BALLESTA et al., 2010; ZOBEL et al., 2007).
6. CONCLUSÕES
O cultivo de J. curcas em volumes de solo menores que 16,4 dm³ por 60
dias resultou em menor crescimento das plantas em termos de biomassa e
trocas gasosas foliares.
A limitação ao crescimento das plantas, provocada pelo menor volume de
solo, foi observada primeiro nas plantas irrigadas (controle), o que mascarou os
efeitos da deficiência hídrica em experimentos com esse intuito.

49
Os dois genótipos estudados possuem diferentes padrões de crescimento
e de trocas gasosas em volumes de solo diversos e sob disponibilidade hídrica
moderada.
Maior comprimento, área e classes de diâmetro de raízes foram
observados nas plantas cultivadas em menores volumes de solo e sob
deficiência hídrica, quando comparadas aos demais tratamentos.

50
7. REFERÊNCIAS
ACHTEN, W.M.J. et al. Jatropha bio-diesel production and use. Biomass and
Bioenergy v. 32, p. 1063–1084, 2008.
ACHTEN, W.M.J. et al. Biomass production and allocation in Jatropha curcas L.
seedlings under different levels of drought stress. Biomass and Bioenergy, p.
1-10, 2010.
ARAÚJO, L. G e SOUSA, K. C. I. Pinhão manso para produção de biodiesel.
Revista Anhangüera, v.9 n.1. p. 95-119, jan./dez. 2008.
BASSO, S. M. S. Caracterização morfológica e fixação biológica de
nitrogênio de espécies de Adesmia DC. e Lótus L. 268f. Tese (Doutorado) –
Universidade Federal do Rio Grande do Sul, Porto Alegre, 1999.
BOHM, W., 2012. Methods of studying root systems. Springer Science &
Business Media.
CARNEIRO, J. G. A.; BARROSO, D. G.; SOARES, L. M. S. Crescimento de
mudas em raiz nua de Pinus taeda L., sob cinco espaçamentos no viveiro e seu
desempenho no campo. Revista Brasileira Agrociência, v.13, p.305-310, 2007.
CASTANHEIRA, E.G. et al. Environmental sustainability of biodiesel in Brazil.
Energy Policy, v. 65, p.680-691, 2014.
CHEN, D. et al. Genotypic variation in growth and physiological response to
drought stress and re-watering reveals the critical role of recovery in drought
adaptation in maize seedlings. Front. Plant Sci, v. 6, p. 1-15.
http://dx.doi.org/10.3389/fpls.2015.01241.

51
CONTRAN, N. et al. State-of-the-art of the Jatropha curcas productive chain:
From sowing to biodiesel and by-products. Industrial Crops and Products, vol.
42, p. 202– 215, 2013.
CREMONEZ, P.A.. Current scenario and prospects of use of liquid biofuels in
South America. Renewable and Sustainable Energy Reviews, vol. 43, p. 352–
362, 2015.
DA SILVA, W. S., KUMMEROW, J. Fine-root growth and longevity in a cacao
(Theobroma cacao L.) plantation. Agrotrópica, v. 10, p. 31-34, 1999.
De OLIVEIRA, P. S. Trocas gasosas foliares e produção de biomassa em
plantas jovens de Jatropha curcas L. submetidas ao déficit hídrico no solo.
Dissertação de mestrado. Ilhéus: UESC, 2013.
De SANTANA, T.A. et al. Water use efficiency and consumption in different
Brazilian genotypes of Jatropha curcas L. subjected to soil water deficit. Biomass
& Bioenergy, v. 75, p. 119-125, 2015.
DÍAZ-LÓPEZ, L. et al. Jatropha curcas seedlings show a water conservation
strategy under drought conditions based on decreasing leaf growth and stomatal
conductance. Agricultural Water Management, v. 105, p.48–56, 2012.
DOBRA, J. Comparison of hormonal responses to heat, drought and combined
stress in tobacco plants with elevated proline content. J. Plant Physiol, v. 167,
p. 1360-1370, 2010.
EDRISI, A. A. Jatropha curcas L.: A crucified plant waiting for resurgence.
Renewable and Sustainable Energy Reviews, v. 41, p. 855–862, 2015.
FINI, A. et al. Water relations, growth, and leaf gas exchange as affected by water
stress in Jatropha curcas. Journal of Arid Environments, v. 89, n. 2, p. 1-29,
2013.

52
FREITAS, R.G. et al. Genetic evaluation of Jatropha curcas: an important oilseed
for biodiesel production. Genetics and Molecular Research 10 (3): 1490-1498,
2011.
FUSSEDER A. A longevidade e atividade da raiz primária do milho. Plant and
Soil, vol. 101, p. 257-265, 1987.
GOMES, F.P.; PRADO, C.H.B.A. Ecophysiology of coconut palm under water
stress. Brazilian Journal of Plant Physiology v. 19, p. 377-391, 2007.
GRIFFITHS, H., WELLER, G., TOY, L.F.M., DENNIS, R.J. You’re so vein: bundle
sheath physiology, phylogeny and evolution in C3 and C4 plants. Plant, Cell and
Environment, 36, 249-261, 2013.
HEROLD A, MCNEIL P. H. Restauração da fotossíntese em plantas de tabaco
encapsuladas em vasos. Journal of Experimental Botany vol. 30, p. 1187-
1194, 1979.
JACKSON, R. B., SPERRY, J. S., DAWSON, T. E. Root Water Uptake and
Transport: Using Physiological Predictions, vol. 5, 2000.
JACKSON R. B. et al. Uma análise global das distribuições de raízes para
biomassas terrestres. Oecologia, vol. 108, p. 389-411, 1996.
JAT, L. K. et al. Jatropha curcas for biofuel: a hope of future renewable energy.
Poll researche. V. 34, n. 4, p. 795-799, 2015.
JESUS, M. M. G., Z. M. SILVA. A isenção tributária do selo combustível social
como estimuladora do Programa Nacional de Produção e Uso do Biodiesel
(PNPB). Direito E-Nergia. Ano 5 – v.8, Ago-Dez, 2013.
KING, A.J. et al. Potential of Jatropha curcas as a source of renewable oil and
animal feed. Journal of Experimental Botany 60: 2897-2905, 2009.

53
KUMAR, A.; SHARMA, S. An evaluation of multipurpose oil seed crop for
industrial uses (Jatropha curcas L.): a review. Industrial crops and products,
v. 28, n. 1, p. 1-10, 2008.
KUMAR, P.; SRIVASTAVA, V. C.; JHA, M. K. Jatropha curcas phytotomy and
applications: Development as a potential biofuel plant through biotechnological
advancements. Renewable and Sustainable Energy Reviews, v. 59, p. 818-
838, 2016.
LAVIOLA, B.G. et al. Estimates of genetic parameters for Physic nut traits based
in the germplasm two years evaluation. Ciência Rural, v. 42, p. 429-435, 2012.
LIMA, R. L. S. et al. Volume de recipiente e composição de substrato para
produção de mudas de mamoneira. Ciência Agrotécnica, Lavras, v. 30, n. 3, p.
480-486, 2006.
MAGHULY, F. et al. Geographic origin is not supported by the genetic variability
found in a large living collection of Jatropha curcas with accessions from three
continents. Biotechnology journal, v. 10, n. 4, p. 536-551, 2015.
MARTIN, I. et al. Jatropha: A Smallholder Bioenergy Crop-The Potential for Pro-
Poor Development. Integrated Crop Management. Vol. 8. Experimental
Agriculture, v. 46, n. 4, p. 567, 2010.
MARTÍNEZ-ALCÂNTARA, B. et al. Relationship between hydraulic conductance
and citrus dwarfing by the Flying Dragon rootstock (Poncirus trifoliata L. Raft var.
monstruosa). Trees - Struct. Funct. v. 27, p. 629-638, 2013.
MARTÍNEZ-BALLESTA, M. C., et al. Physiological aspects of rootstock-scion
interactions. Sci. Hortic. Amst, v. 127, p. 112-118, 2010.
MOURA, A. R.; NOGUEIRA, R. J. M. C.; SILVA, J. A. A.; LIMA, T. V. Relações
hídricas e solutos orgânicos em plantas jovens de Jatropha curcas L. sob

54
diferentes regimes hídricos. Ciência Florestal, Santa Maria, v. 26, n. 2, p. 345-
354, abr.-jun., 2016.
MURPHY, M.R.C, JORDAN, G.J., BRODRIBB, T.J. Acclimation to humidity
modifies the link between leaf size and the density of veins and stomata. Plant,
Cell and Environment, 37, 124-131, 2014.
NORTH, C.A. 1956. Technique for measuring structural features of plant
epidermis using cellulose acetate films. Nature 178: 1186–1187.
PANDEY, V.C. et al. Jatropha curcas: A potential biofuel plant for sustainable
environmental development. Renewable and Sustainable Energy Reviews 16:
2870– 2883, 2012.
PASSIOURA, J. The drought environment: physical, biological and agricultural
perspectives. Journal of Experimental Botany v. 58, n. 2, p. 113-117, 2007.
PEDROSO, F. K. J. V. Drought tolerance in citrus trees is enhanced by rootstock-
dependent changes n root growth and carbohydrate availability. Environ. Exp.
Bot. v. 101, p. 26-35, 2014.
PIMENTEL, C. A relação da planta com a água. Seropédica, Rio de Janeiro
edur. 19p, 2004.
PINHEIRO, H. A. et al. Drought tolerance is associated with rooting depth and
stomatal control of water use in clones of Coffea canephora. Annals of Botany,
v. 96, p. 101-108, 2005.
SÁ, M. A. C.; SANTOS JUNIOR, J. D. G. Compactação do solo:
consequências para o crescimento vegetal. Planaltina: Embrapa Cerrados,
2005. 26 p. (Embrapa Cerrados. Documentos, 136).
SAMBATTI B.M.; CAYLOR, M.M. When is breeding for drought tolerance optimal
if drought is random? New Phytologist. n.175, p. 70-80, 2007.

55
SAPETA, H. et. al. Drought stress response in Jatropha curcas: Growth and
physiology. Environmental and Experimental Botany, v. 85, p. 76-84, 2013.
SOUSA, A. E. C. et al. Trocas gasosas e conteúdo de carboidratos e compostos
nitrogenados em pinhao-manso irrigado com águas residuária e salina.
Pesquisa agropecuária brasileira, v.47, n.10, p.1428-1435, 2012.
TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 4.ed. Porto Alegre: Artmed, 2009.
819p.
TARDIEU, F.; GRANIER, C.; MULLER, B. Water deficit and growth. Co-
ordinating processes without an orchestrator?. Current opinion in plant
biology, v. 14, n. 3, p. 283-289, 2011.
TIWARI, N., PUROHIT, M., SHARMA, G., NAUTIYAL, A. R. Changes in Morpho-
Physiology of Jatropha curcas grown under different water regimes. Nature and
Science, vol. 11, p. 76-83, 2013.
UNEP – United Nations Environment Programme. Division of Technology,
Industry and Economics. Bioenergy issue paper series, 2010. Disponível em
http://www.unep.fr/en/info/publications.htm. Acesso em 19 de Abril de 2015.
VERMA, K. K. et al. Developing a mathematical model for variation of
physiological responses of Jatropha curcas leaves depending on leaf positions
under soil flooding. Acta Physiologiae Plantarum, v. 34, n. 4, p. 1435-1443,
2012.
YANG, H. et al. Land and water requirements of biofuel and implications for food
supply and the environment in China. Energy policy, v. 37, p. 1876-1885, 2009.
XU, Z., ZHOU, G., SHIMIZU, H. Plant responses to drought and rewatering. Plant
Signal. Behav, vol. 5, p. 649-654, 2010.

56
ZOBEL, R. W.; KINRAIDE, T. B.; BALIGAR, V. C. Fine root diameters can change
in response to changes in nutrient concentrations. Plant Soil, v. 297, p. 243-254,
2007.
ZONTA. E. et al. O sistema radicular e suas interações com o ambiente
edáfico. In: FERNANDES, M. S. (Ed.). Nutrição mineral de plantas. Viçosa:
Sociedade Brasileira de Ciência do Solo, 2006. p. 7-52.