vicia sativa l. aggregate (fabaceae) in the flora of former ussr
TRANSCRIPT

Genetic Resources and Crop Evolution 44: 199–209, 1997. 199c 1997 Kluwer Academic Publishers. Printed in the Netherlands.
Vicia sativa L. aggregate (Fabaceae) in the flora of former USSR
Elena K. PotokinaDepartment of Plant Introduction and Taxonomy, N.I. Vavilov Institute of Plant Industry, Bolshaya MorskayaStreet, 42, 190000 St. Petersburg, Russia
Received 4 December 1995; accepted 10 July 1996
Key words: geographical distribution, natural populations, taxonomy, variability, Vicia sativa agg.
Summary
Morphological differences and ecogeographical characteristics of five species from Vicia sativa agg. in the areaof former USSR were investigated. The key to V. sativa agg. species and subspecies as well as maps of theirareals are presented. A complementary study of polymorphism at the phenotypical level of 58 local weedy andwild populations from different geographical regions was conducted to determine areas with higher variability ofmorphological characters used for intraspecific taxonomy and cultivar identification. The regions of greatest interestfor curators and collectors of V. sativa agg. germplasm are some regions of Transcaucasus (Nagornyi Karabakh,Talysh range, Lenkoran valley) and South of Crimea.
Introduction
Vicia sativa agg., an important fodder legume crop,combines a group of autogamous closely related taxa.This group is taxonomically complicated since mor-phological distinctions between taxa in the V. sativaagg. are often not well expressed, the species occu-py more-or-less similar habitats, and most of them aredistributed sympatrically.
Some authors recognized eight separate species inthis aggregate (Fedchenko, 1948; Mettin & Hanelt,1964; Hanelt & Mettin, 1966; Tzvelev, 1987) viz. : V.sativa L., V. angustifolia Reichard, V. cordata Wulf.ex Hoppe, V. macrocarpa (Moris) Bertol., V. amphi-carpa Dorth., V. incisa Bieb., V. pilosa Bieb., V. sege-talis Thuill. Other taxonomists regard as only a singlespecies V. sativa L. sensu lato with certain subspeciesand varieties (Ball, 1968; Davis & Plitmann, 1970;Stankevich, 1978; Zohary & Plitmann, 1979; Maxted,1995). The latter point of view is based on the reasonthat the diagnostic features for some species prove inpractice to be ‘overlapping’ characters. In this con-nection there are transitional forms, and taxa representmore peaks in a continuous variation than clear-cutsectors.
The group is no less problematic cytologically.Three different chromosome numbers 2n= 10,2n= 12and 2n = 14 were reported for this aggregate. Met-tin & Hanelt (1964) and Yamamoto (1973) statedthe relationship between the morphological charactersand karyotypes among species of V. sativa agg. Theremarkable intrataxon karyotypic variation wasdescribed by Hollings & Stace (1974). Ladizinsky(1978) found a given karyotype in several taxa andseveral karyotypes in the same taxon.
Spontaneous hybridizations and recombinationsbetween the different karyotypes as well as sponta-neous chromosomal reconstructions into V. sativa com-plex have been reported by many authors (Mettin &Hanelt, 1973; Holling & Stace, 1974; Zohary & Plit-man, 1979).
Apart from complicated taxonomical relations withother species of the aggregate, Vicia sativa L. sensustricto (common vetch) is distinguished by the greatamount of phenotypic variability in populations ofweedy, escapees and cultivated forms. Many attemptshave been made to describe this variability by taxo-nomical categories. So the recent intraspecific classi-fication of V. sativa subsp. sativa proposed by Repyev& Stankevich (1989) is based on the variability of seedcoat pattern and flower colour and describes seven vari-

200
eties and 12 forms. All of them can occur in the localpopulations of V. sativa, which are common in fallowfields, roadsides and also sometimes grow on untilledlands. Knowledge of the geographical distribution ofvarieties and forms of V. sativa agg. in the area of theformer USSR is needed to set up strategies for sam-pling Vicia germplasm.
Besides, there are many qualitative characters likecolour of cotyledons and hilum colour of seeds, whichusually are not used for taxonomy,but are important forcultivar identification. Information on the variabilityof these hereditary characteristic in populations fromdifferent geographical regions can also be useful forcurators and collectors.
The purpose of the present work was to study andclassify the diversity of Vicia sativa agg. in the floraof the former USSR on both an inter- and intraspecificlevels. To this end we conducted a taxonomic studyof V. sativa agg. forms as well as an analysis of thepolymorphism of weedy and wild populations of V.sativa agg. taxa.
Materials and methods
The taxonomic conclusions reached in this study weremainly the result of morphological studies involv-ing herbarium specimens, live populations of VIRgermplasm collections and natural populations of allthe V. sativa agg. taxa concerned.
The materials from most important herbaria of theformer USSR were used: Komarov Institut of Botany,St. Petersburg (LE); Vavilov Institut of Plant Industry,St. Petersburg (WIR); St. Petersburg State Universi-ty (LECB); Moscow State University (MW); MoscowBotanical Garden (MHA); Botanical Institutes ofAcademy of Science of Azerbaijan (BAK), Turkme-nia (ASH), Tadjikistan (TAD), Uzbekistan (TASH).
The curators of the following herbaria kindly assist-ed with loans of specimens: Krylov Herbarium ofTomsk State University, Russia (TK); Institute ofZoology and Botany, Tartu, Estonia (TAA); Botani-cal Institute of the Bulgarien Academy of Sciences,Sofia, Bulgaria (SOM); Institut fur SystematischeBotanik und Pflanzengeographie der Martin-Luther-Universitat, Halle, Germany (HAL); Fielding Herbar-ium, Department of Botany, Oxford, Great Britain(OXF); Instituto Botanico de Barcelona, Spain (BC);Instituto ‘Antonio Jose Cavanilles’, Jardin Botanico,Madrid, Spain (MA); Institut de Botanique, Universite
de Montpellier, France (MPU); Department of Botany,University of Uppsala, Sweden (S).
Maps of geographical distribution of V. sativa agg.taxa were composed using herbarium specimens exam-ined. For rare species with limited distribution thedetailed information regarding locality, ecology andcollectors names are indicated in Appendices 1–3.
Live populations of 247 accessions Vicia sativa agg.taxa from the VIR world germplasm collection, rep-resenting different varieties and forms from differentgeographical origins, were studied at the VIR Experi-mental Station (St. Petersburg province) in 1987–1990.
The herbarium specimens and seed samples from58 local weedy and natural populations of V. sativaagg. were collected during field trips in 1987–1991to Caucasus (Azerbaijan, Russia), Crimea and Kerchpeninsula (Ukraina), Central Asia (Turkmenia, Tad-jikistan, Uzbekistan) and the North European part ofRussia (Arkhangelsk, Vologda, St. Petersburg, Nov-gorod provinces).
Populations phenetics
There are several approaches to describing the diversityof real natural populations. We followed one of them,which has been called population phenetics.
Population phenetics is an extension of genet-ic principles and approaches to species and formsof which genetic study is difficult or unfeasible(Yablokov, 1986). The subject of phenetics is sub-specific variability. The latter is in the final analysisreduced to discrete alternative markers of the popula-tion genotypic composition, i.e. phenes. The methodsof phenetics are qualitative and quantitative investiga-tions of phenes in populations and other con-specificand interspecific units. The two main trends in phe-netic research are: (a) a study of structure and dynam-ics of separate phene pools, and (b) a study of thespatial distribution of phene pools (phenogeography).The term ‘phene’ was introduced by W. Johannsen(1909) along with the terms gene, genotype, pheno-type, allele to denote a genetically determined char-acter of the phenotype. A definition of the subject,objectives and methods of population phenetics wasproposed by Timofeeff-Ressovsky et al. (1977).
A theoretical basis for this approach has been laidby the rule of homologous series in inherited variabili-ty formulated by N.I. Vavilov in 1920. Vavilov appliedgenetic criteria in his analysis of individual phenotypesin natural populations. The key point in this approachwas the recognition of genotype markers within the

201
Table 1. Phene of colour and seed coat pattern detected in local populationsof V. sativa and V. angustifolia
Phene Phene Taxon for V. sativa
number description described
1 Reticulation with black var. typica Beck
points trough all the seed
2 Reticulation without var. variabilis Ted.
points
3 Separate large spots var. maculata Burnot.
without reticulation
4 Seed coat pattern absent var. melanosperma Reichenb.
colour of seed black
5 Seed coat pattern absent var. erythrosperma Reichenb.
colour of seed brown
6 Seed coat pattern absent var. immaculata Ted.
colour of seed green
7 Seed coat pattern absent var. leucosperma Ser.
colour of seed white
overall variation in the character. Such are invariably,qualitative, discrete, alternative traits, like the greenand yellow colour of pea seeds in Mendel’s experi-ments.
We analysed the polymorphism of populations forV. sativa agg. taxa, which is quite widely distributedthroughout the investigated area: V. sativa, V. angus-tifolia and V. segetalis. Fifty-eight local native andweedy populations were investigated using the meth-ods and approaches of population phenetics. Variantsof the following characteristic were chosen as phenes:hilum colour (black and white), endosperm colour(orange, yellow, pink) and seven variants of seed coatpattern (Table 1). All of them are included in the Inter-national Comecon List of Descriptors for the speciesVicia sativa L. (1985), except for the pink colour ofcotyledons, which has been found only in the pop-ulations of V. angustifolia. The distribution of thesephenes and their frequency in local populations wereanalysed.
Results
Taxonomy of Vicia sativa agg.
The proposed key for identification of V. sativa agg.taxa includes two species that are not presented inthe flora of the former USSR: V. macrocarpa (Moris)Bertol. and V. cordata Wulf. ex Hoppe. The morpho-
logical description of these species is based on a studyof foreign herbaria and on the examination of live pop-ulations grown from seeds obtained from the VIR col-lection. V. cordata has earlier been reported from theCaucasus (Fedchenko, 1948; Grossheim, 1952), butthe revision of herbarium specimens and natural pop-ulation observations in Caucasus, Crimea and Cen-tral Asia did not confirm this fact. In the main speci-mens with heart-shaped leaflets identified as V. cordatashould be related to V. angustifolia subsp. segetalis onthe basis of pod and seed features.
Key to Vicia sativa agg. species and subspecies
1. At least some leaflets incised, corolla blue : : : : : :
: : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : V. incisa— All leaflets entire, corolla purple : : : : : : : : : : : : : : 2
2. Corolla 23–28 mm long; legume 50–80 mm long,10–15 mm wide. Mature legumes (blackening)with rough lignified walls, on their surface a netof veins is observed; immature legumes with thickfleshy walls : : : : : : : : : : : : : : : : : : : : :V. macrocarpa
— Corolla 9–24 mm long; legumes 20–80 mm long,4–8 mm wide : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : 3
3. Legumes densy hairy, hairs 0.7–1.0 mm long;plants 5–30 cm height, pubescent : : : : : : : : : : : : : 4
— Legumes either naked or with hairs not morethan 0.3 mm long; plants 25–90 cm height,glabrous : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : 5

202
Fig
ure
1.D
istr
ibut
ion
ofth
etw
osu
bspe
cies
ofV.
sativ
ain
the
form
erU
SSR
.�
V.sa
tiva
subs
p.sa
tiva;
+V.
sativ
asu
bsp.
linea
rfol
ia.

203
4. Corolla 17–24 mm, double length of calyx; calyxwith few hairs along veins located; plants formingunderground stems with flowers and legumes : : : :
: : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : V. amphicarpa— Corolla 9–12 mm long, slightly longer or approx-
imately equal to calyx; calyx with numerous longwhite hairs; plants without underground stems : : :
: : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : V. pilosa
5. Flowers 13–24 mm long; legumes yellow or brown,with short hairs; legumes distinctly laterally flat-tened in cross-section and distinct constrictionsbetween seeds : : : : : : : : : : : : : : : : : : : : : : : : V. sativa
6. Legumes 60–80 mm long,7–9 mm wide, with 8–12seeds; leaflets elliptic or obovate : : : subsp. sativa
— Legumes 45–55 mm long, 5–7 mm wide, with 5–7seeds; leaflets linear or cuneate : : : : : : : : : : : : : : : : :
: : : : : : : : : : : : : : : : : : : : : : : : : : : : subsp. linearifolia— Flowers 13–18 mm long; legumes glabrous, black
or brown; legumes rounded in cross-section, con-strictions between seeds are absent : : : : : : : : : : : : 7
7. Seeds number in legume 5–8; mature legume light-or dark-brown; seeds 4–5 mm in diam : : :V. cordata
— Seeds number in legume 9–12(14); mature legumeblack; seeds 2–3 mm in diam : : : : : V. angustifolia
8. Leaves with 3–4 pairs of leaflets; flowers usuallyone-coloured, reddish purple : : : : : : : : : : : : : : : : : :
: : : : : : : : : : : : : : : : : : : : : : : : : : : : subsp. angustifolia— Leaves with (4)5–7 pairs of leaflets; flowers
bicoloured: standard face pale lilac, wings purple: : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : subsp. segetalis
Vicia sativa L. s.str.subsp. sativa–V. sativa L. 1753, Sp.P1.: 736.
Combines mesophyllic forms, widely cultivated and as weedsof legume and cereal crops in the European part of Russia. In Cen-tral Asia, Siberia and Far East is an adventitious plant (Figure 1).Flowering: June–August. 2n = 12.subsp. linearifolia Stankev. 1978, Bull. VIR. 81:6–V. cosentini Guss.1828, Fl.Sic.Prodr. 2:426.–V. sativa var. cosentini (Guss.) Arcang.1882, Comp.Fl.It. ed.1:201; Mettin & Hanelt, 1964, Kulturpflanze,12:212.
It differs from the previous subspecies in xeromorphic habit, lin-ear cuneate leaflets, narrower and shorter pods. Distributed in theNorth of Crimea, at the Caucasus, in Central Asia as weeds ingramineous crops, along road sides, in weedy locations (Figure 1).In Turkmenia (Kara-Kala) was found in natural coenoses (mountainslopes, stream banks). Fl.: April–June. 2n = 12; 14 (Stankevich1978).
Vicia angustifolia Reichardsubsp. angustifolia.–V. angustifolia Reichard, 1778, Fl.Moeno-Francof. 2:44; L. 1759 Amoen.Acad. 4:105, nom.nud.; Hoppe inSturm, 1812, Deutschl. Fl. 1, 32: tab.498.–V. sativa var. nigra L.1763, Sp.Pl., ed. 2:1037.
It is distributed in the Northern part of European Russia, theBaltic Sea countries and Beylorussia (Figure 2) and grows in drymeadows, along road sides, as weeds in cereal crops, preferringloose sandy substrates, F1.: June–September. 2n = 12.
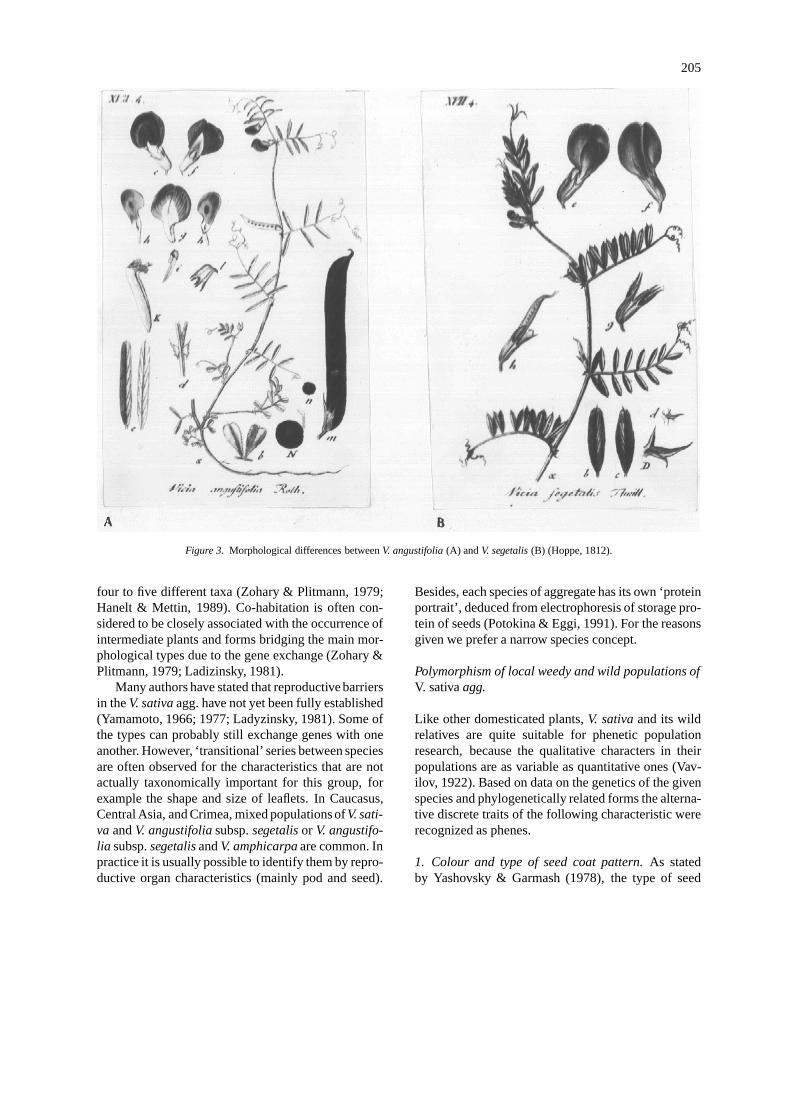
Herbarium specimens originated from Caucasus, Crimea andCentral Asia and identified as V. angustifolia should be related to V.angustifolia subsp. segetalis. The illustrations proposed by D. Hoppe(1812) quite clearly show the morphological differences between V.angustifolia and V. segetalis (Figure 3).subsp. segetalis (Thuill.)Gaud. 1829, Fl.Helvet. 4:511.–V. segetalisThuill. 1799, F1.Paris, ed.2: 367; Hoppe in Sturm, 1812, Deutschl.F1, 32: tab.499.–V. angustifolia subsp. segetalis(Thiull.)Arcang.1822, Comp. Fl.Ital.:202; Mettin & Hanelt, 1964, Kulturpflanze,12:207.
It is widely distributed in central and southern regions of theEuropean part of Russia, in the Crimea, at the Caucasus and CentralAsia (Figure 2). In Siberia and in the Far East occurs as adventitiousplant. Preferring the same habitat as the latter subspecies. F1.: May–September. 2n = 12.
Vicia amphicarpa Dorth.1789 Journ.phys. 35–131.–V. sativa subsp. amphicarpa(Boiss.)Aschers.et Graebn., 1909, Syn.Mitteleurop.F1. 6, 2:974.–V. pilosa Bieb. var. amphicarpa Krjuk. 1964, Bot.Journ.of USSR.49.8:1208.
It is distributed in the Crimea, Kerch peninsula and Transcauca-sus (Figure 4, Appendix 1). and inhabits in dry meadows, mountainslopes. In the Crimea often grows in common phytocoenoses withV. angustifolia subsp.segetalis. In this case V. amphicarpa can berecognized by strong pubescence of all the plant parts including podand underground shoot formation. F1.: April–June. 2n = 14.
Vicia pilosa Bieb.1808 Fl.Taur.-Cauc. 2:161; Hanelt & Mettin, 1966. Kulturpflanze.13:138.–V. sativa subsp. pilosa (Bieb.)Zohary & Plitmann, 1979,Plant Syst.Evol. 131, 12:146.
It is distributed in the southern Crimea and Transcaucasus (Fig-ure 4, Appendix 2). Grows in stony slopes. The specimen fromTarkhankut peninsula (Crimea) (Krjukova, 1964) described as V.pilosa var.amphicarpa Kruik. should be related to V. amphicarpa.F1.: April–June. 2n = 14.
Vicia incisa Bieb.1819 Fl.Taur.-Cauc. 3:471.–V. sativa subsp. incisa (Bieb.)Arcang.1882, Comp.F1.Ital., ed.2:201.
A species with a narrow endemic areal (south of France, Greece,the Crimea) (Figure 4, Appendix 3). In the Crimea it occurs in moistshady canyons, oak-forest margins. Fl.: May–July. 2n = 14.
Debates on the taxonomic status of some taxa of V. sati-va agg. arise due to the their not well expressed mor-phological distinctions. The cause of this is a parallelhomologous variability of V. sativa agg. taxa (Davis &Plitmann, 1970). Some of them are probably hybridderivatives, the result of hybridization between therelated former well-defined taxa, adapted to more-or-less primary habitats. At the present time the weedytypes of taxa colonize distributed sites and are not iso-lated ecologically, so one field may be inhabited by

204
Fig
ure
2.D
istr
ibut
ion
ofth
etw
osu
bspe
cies
ofV.
angu
stifo
liain
the
form
erU
SSR
.�
V.an
gust
ifolia
subs
p.se
geta
lis;
+V.
angu
stifo
liasu
bsp.
angu
stifo
lia.
205
Figure 3. Morphological differences between V. angustifolia (A) and V. segetalis (B) (Hoppe, 1812).
four to five different taxa (Zohary & Plitmann, 1979;Hanelt & Mettin, 1989). Co-habitation is often con-sidered to be closely associated with the occurrence ofintermediate plants and forms bridging the main mor-phological types due to the gene exchange (Zohary &Plitmann, 1979; Ladizinsky, 1981).
Many authors have stated that reproductive barriersin the V. sativa agg. have not yet been fully established(Yamamoto, 1966; 1977; Ladyzinsky, 1981). Some ofthe types can probably still exchange genes with oneanother. However, ‘transitional’ series between speciesare often observed for the characteristics that are notactually taxonomically important for this group, forexample the shape and size of leaflets. In Caucasus,Central Asia, and Crimea, mixed populations of V. sati-va and V. angustifolia subsp. segetalis or V. angustifo-lia subsp. segetalis and V. amphicarpa are common. Inpractice it is usually possible to identify them by repro-ductive organ characteristics (mainly pod and seed).
Besides, each species of aggregate has its own ‘proteinportrait’, deduced from electrophoresis of storage pro-tein of seeds (Potokina & Eggi, 1991). For the reasonsgiven we prefer a narrow species concept.
Polymorphism of local weedy and wild populations ofV. sativa agg.
Like other domesticated plants, V. sativa and its wildrelatives are quite suitable for phenetic populationresearch, because the qualitative characters in theirpopulations are as variable as quantitative ones (Vav-ilov, 1922). Based on data on the genetics of the givenspecies and phylogenetically related forms the alterna-tive discrete traits of the following characteristic wererecognized as phenes.
1. Colour and type of seed coat pattern. As statedby Yashovsky & Garmash (1978), the type of seed

206
Figure 4. Distribution of V. amphicarpa, V. pilosa and V. incisa in Caucasus and Crimea. See Appendix 1, 2, 3.
coat pattern as a complicated qualitative character iscontrolled by no fewer then 10 genes, activating orsupressing the pigment synthesis, Tedin & Witt (1900)described nine varieties of common vetch by theirtypes of seed coat pattern. This classification has beenused with success for cultivar identification. Sevenphenes of colour and type of seed coat pattern occurin the local populations of V. sativa and V. angustifo-lia in Transaucasus, North Caucasus and Central Asia(Table 1). The average frequency of the phenes in eachregion is shown in Figure 5.The region with the highestdiversity of this characteristic for V. sativa was assignedto southern Azerbaijan and Nagornyi Karabakh (Tran-scaucasus region) (Figure 5, Graphs A and B). Morethan six variants of seed coat pattern can be found inthe local population of this area. In Central Asia coun-
tries (Turkmenia, Tadjikistan) only two phenes weredetected and the diversity seemed to be lower (Fig-ure 5, Graphs D and E). The centre of diversity ofthis characteristic for V. angustifolia subsp. segetalisappeared to be located in the Crimea rather than inTranscaucasus (Figure 2, Graph F). Weedy and wildpopulations of V. angustifolia subsp. angustifolia wereinvestigated in the Northern part of European Russia.The variability of colour and type of seed coat pat-tern in this subspecies was considered to be lower thanthat in V. augustifolia subsp. segetalis. The most pre-dominant phene 1 accounted for approx. 97% in thepopulations and only three phenes were detected.
2. Cotyledon colour of mature seeds. This trait hasbeen used by Vavilov (1920) to analyse natural inher-

207
Figure 5. Frequency of phenes of seed colour and coat pattern in populations of V. sativa and V. angustifolia in each region: A – Azerbaijan; B– Nagornyi Karabakh; C – North Caucasus; D – Turkmenia; E – Tadjikistan; F – Crimea. 1–7: No. of phenes.
ited variability of vetch, pea, lentils and other plantsof Leguminosae. Colour of cotyledons may be usedwith success for vetch cultivar identification (Redki-na, 1970). Three phenes of this character were foundin local populations of V. sativa and V. angustifolia:orange, yellow, pink. In contrast to the trait discussedabove the frequency of cotyledon colour in V. sati-va agg. populations serves as a marker of species andsubspecies. Plants with orange coloured cotyledons arepredominant in the populations of V. sativa and are rarein V. angustifolia (Figure 6). The frequency of yellowand pink cotyledons in populations of V. angustifoliashows a clear geographical cline (Figure 6). The factis that the local populations of V. angustifolia in theNorthern part of the European Russia was identifiedas V. angustifolia subsp. angustifolia. The populations
from Caucasus, Crimea and Central Asia should berelated to V. angustifolia susbsp. segetalis. The popu-lations of V. angustifolia subsp. angustifolia are char-acterized by the high frequency of plants with pinkcoloured cotyledons (75–91%). The plants with yel-low cotyledons are predominant in populations of V.angustifolia subsp. segetalis (89%).
3. Colour of seed hilum. According to Bould & Grofton(1987), this character is controlled by one locus. Thewhite coloured hilum is dominant. The average fre-quency of black hilum in the populations investigat-ed was not above 6%. The frequency distribution ofhilum colour phenes depends neither on the taxonomi-cal belonging of population nor its geographical local-ity.

208
Figure 6. Frequency of occurrence (relative size of the circles andsquares) of three cotyledon colour phenes in populations of V. sativaand V. angustifolia. Phenes: 1 – yellow; 2 – orange; 3 – pink.
Conclusions
The results of our study of V. sativa agg. in the area offormer USSR may be summarized as following:
V. sativa agg. is presented in the flora of formerUSSR by five morphologically separated taxa. Two ofthese (V. sativa sensu stricto and V. angustifolia) maybe classified into subspecies.
V. cordata, which earlier has been reported fromthe Crimea and Caucasus in many cases was confusedwith related taxa. According to our data, this speciesof Mediterranean distribution is not present in the floraof the former USSR.
V. pilosa var. amphicarpa, which has beendescribed in 1963 from Tarkhankut peninsula (theCrimea) should be related to V. amphicarpa. These twospecies may be distinguished by their calyx pubescenceand size of flowers.
Though there is some overlap in their morphologi-cal characteristic, most of the specimens of V. angus-tifolia subsp. angustifolia and V. angustifolia subsp.
segetalis are distinguishable by the number of leafletsand colour of flowers. They also have some differencein their geographical distribution and phenetical struc-ture of populations.
All variants of colour and coat pattern of seeds forV. sativa described as ‘varieties’ were also detected inpopulations of V. angustifolia.
The centre of diversity in V. sativa as indicated bythe phenetic population research was considered to beTranscaucasus area (Nagornyi Karabakh,Talysh range,Lenkoran valley). The most variable populations of V.angustifolia were found in the South of Crimea.
References
Ball, P.W., 1968. Vicia L. In: Flora Europaea, 2:129–134. Cam-bridge.
Bould, A. & G. Grofton, 1987. Variability in the expression ofhilum colour in field bean varieties in relation to seed certificationstandards. Seed Sci and Technol 15, 3:11–17.
Davis, P.H. & U. Plitman, 1970. Vicia L. In: P.H. Davis (Ed.), Floraof Turkey 3, p.p. 274–325, University Press, Edinburgh.
Fedchenko, B.F., 1848. Vicia L. In:(Flora of USSR) 13:460–465,Moscow.
Grossgeim, A.A., 1952. Flora of Caucasus. Vol. 5, 453 pp. Moscow–Leningrad.
Hanelt, P. & D. Mettin, 1966. Cytosystematische Untersuchungen inder Artengruppe um Vicia sativa L. II. Kulturpflanze 14:137–161.
Hanelt, P. & D. Mettin, 1989. Biosystematics of the genus Vicia L.(Leguminosae). Annu Rev Ecol Syst 20:199–223.
Hollings, E. & C.A. Stace, 1974. Karyotype variation and evolutionin the Vicia sativa aggregate. New Phytol. 73:195–208.
Hoppe, D., 1812. Vicia L. In: J. Sturm. (Ed.) Deutschlands Flora,3:10–11, Nurnberg.
International Comecon List of Descriptors for the speciesVicia sativaL., 1985. Leningrad, 59 pp.
Johannsen, W., 1909. Elements der exacten Erblicheitlsehre. Fischer,Jena.
Krjukova, I.V., 1964. Amfikarpija u Vicia pilosa M.B. Bot. Zurn.49:1207–1209.
Ladizinsky, G., 1978. Chromosomal polymorphism in wild popula-tions of Vicia sativa L. Caryologia 31:233–241.
Ladizinsky, G., 1981. Consequences of hybridization in Vicia sativaaggregate. Heredity 47:431–438.
Maxted, N. 1995. An ecogeographical study ofVicia subgenus Vicia.Systematics and Ecogeographic Studies on Crop Genepools. 8.,184 pp.
Mettin, D. & P. Hanelt, 1964. Cytosystematische Untersuchungen inder Artengruppe um Vicia sativa L. Kulturpflanze 12:163–225.
Mettin, D. & P. Hanelt, 1973. Uber Speziationsvorgange in derGattung Vicia L. Kulturpflanze 21:25–54.
Potokina, E.K. & E.E. Eggi, 1991. Electrophoresis of seed storageprotein in the solution of the problems on Vicia sativa agg. tax-onomy. Bull of Applied Botany, Genetics and Plant Breeding139:112–123.
Redkina, Z.V., 1970. Razrabotka metodov opredelenija podlinnostisemjan vik i gorokha. Avtorefx. Dis. Leningrad, 28 pp.

209
Repyev, S.I. & A.K. Stankevich, 1989. Klassifikacija vozdelyvae-mych form viki posevnoj (Vicia sativa L.). Bulleten VIR, 190:7–11.
Stankevich, A.K., 1978. K sistematike politipnogo vida Vicia sativaL.s.l. Bulleten VIR, 81:3–11.
Tedin, H. & H. Witt, 1900. Botanisk-kemisk Untersokning af 77po 2 undantog nor, nyk vickerformer, upprdgna vid SverigesUtsaderforening po Svalof. Malvo, 60 pp.
Timofeeff-Ressovsky, N.V., A.V. Yablokov & N.V. Glotov, 1977.Grundris der Populationslehre. Jena, 278 pp.
Tzvelev, N.N., 1987. Vicia L. In: (Flora of the European Part ofUSSR). 6:127–147, Leningrad.
Vavilov, N.I., 1922. The law of homologous series in variation. J.Genetics 12:47–89.
Yablokov, A.V., 1986. Population biology. Advances in Scienceand Technology in the USSR. Biology Series. Mir PublishersMoscow, 303 pp.
Yamamoto, K., 1966. Studies on the hybrids among the Vicia sativaL. and its related species. Mem Fac Agr Kagawa Univ 21:1–104.
Yamamoto, K., 1973. Karyotaxonomical studies on Vicia L. I. Onthe karyotype and character of some annual species of Vicia. JpnJ Genetics 48, No.5:315–327.
Yamamoto, K., 1977. On the intraspecific hybrid between Viciaamphicarpa and V. sativa. Tech Bull Fac Agr Kagawa Univ28:23–31.
Yashovsky, I.V. & E.S. Garmash, 1978. Character of inheritance ofseed colour in common vetch (Vicia sativa L.) hybrids. Genetics(USSR) 24(8):1423–1431.
Zohary, D. & U. Plitman, 1979. Chromosome polymorphism,hybridization and colonization in the Vicia sativa group(Fabaceae). Plant Syst & Evol 31:143–156.
Appendix 1
Current distribution of V. amphicarpa in former USSR.See Figure 4.
Caucasus: Nagornyi Karabakh, Zangelan reg.,Pirchevan sett., alt. 400 m, stone slopes, 7 May 1948,Vassilyev (LE); Daghestan, distr.Kurinski, Bilidzhi,1902, Alexeenko (LE); Dadhestan, Derbent, in her-bosis inter paliureta, 1902, Alexeenko (LE under thename V. cordata).
Crimea: Koktebel, 1903, Golde (LE); Koktebel,1907, Junge (LE); Simeiz, 1903, Golde (LE); Sudakreg., Planerskoye sett., Biyuk-Enishar range, cobbleslopes, 26 April 1988, Shatko & Volkovskaya (MHA);Sudak reg., Biyuk-Enyshar range, seeside slope, 9 May1986, Belyanina (MHA); Sudak reg., Biyuk-Enishar
range, near Planerskoye sett., slopes, 21 May 1987,Smirnova & Shatko (MHA); Kerch peninsula, AzovSea coast, Kultepe sett., 1908, Klopotov (LE, under thename V. angustifolia f. pusilla); Kerch pen., Leninskreg., Zolotoye sett., dry steppe slopes, 12 May 1988,Shatko & Volkova (MHA); Kerch pen., Arabatskyi Bayof Azov Sea, Kamenskoye sett., dry steppe slopes, 27June 1988, Potokina (VIR); Tarkhankut pen.,Olenevkasett, old fallow, 24 June 1963, Kryukova (LE, underthe name V. pilosa var. amphicarpa).
Appendix 2
Current distribution of V. pilosa in former USSR. SeeFigure 4.
Caucasus: Azerbaijan, Dzherbrail reg., betweenGuzlyak and Dash-kesan sett., alt 400 m, stony slopes,28 April 1948, Vassilyev (LE); Nagornyi Karabakh,Gadrut reg., Dash-Bashi sett., alt. 650 m, stone slopes,2 May 1948, Grossgheim (LE); Daghestan, Derbent,in decliv steppaceis, 17 May 1902, Alexeenko (LE).
Crimea: Baydarskaya valley, Orlinoye sett., rockyslopes, 1984, Tzvelev, Geltman & Medvedeva (LE);Sudak reg., NE of Planerskoye, Biyuk-Enyshar range,seeside slopes, 10 May 1986, Belyanina (MHA);Sudak reg., Planerskoye sett., slopes, 14 May 1978,Shvedchikova (MW); Sudak, stone slopes, 29 April1977, Shvedchickova (MW); Laspi valley, Juniperusforest, 19 May 1986, Belyanina & Shatko (MHA);Sudak reg., Karadag, Kok-Kaya range, seeside slopes,26 May 1984, Shako (MHA).
Appendix 3
Current distribution of V. incisa in former USSR. SeeFigure 4.
Crimea: Sudak reg., Perchem range, oak-forest, 1May 1978, Shvedchikova (MW); Yalta reg., Ayu-Dagmountain, oak-forest, 13 June 1988, Potokina (VIR);Agu-Dag, 1962, Chitkina (LE); Nikita, 1922, Stankov(LE); Massandra, 1901, Transhel (LE).