3 accepted - journal of virologyjvi.asm.org/content/early/2008/02/06/jvi.02568-07.full.pdf · 3...
TRANSCRIPT

1
SINGLE DOSE PROTECTION AGAINST P. berghei BY SIMIAN ADENOVIRAL 1
VECTOR USING HUMAN CMV PROMOTER CONTAINING INTRON A 2
3
Sridhar, S.*,†, Reyes-Sandoval, A.† ,Draper,S.J.†, Moore, A.C.†, Gilbert, S.C.†, Gao, 4
G.P. ¶, Wilson, J.M.¶ and Hill, A.V.S.† 5
6
† The Wellcome Trust Centre for Human Genetics, University of Oxford, Roosevelt 7
Drive, Oxford, OX3 7BN, U.K., 8
¶Department of Pathology and Laboratory Medicine, School of Medicine, University 9
of Pennsylvania, 2000 Translational Research Program 125S, 31st Street 10
Philadelphia, PA 19104-3403, U.S.A. 11
. 12
Running title: Simian adenoviral vector with novel promoter as a malaria vaccine. 13
14
* CORRESPONDING AUTHOR: 15
The Wellcome Trust Centre for Human Genetics, University of Oxford, Roosevelt 16
Drive, Oxford, OX3 7BN. U.K. 17
e-mail: [email protected]. Phone: +441865287592; fax: +441865287686 18
19
Abstract Word Count : 244 20
21
Manuscript Word Count: 6801 22
23
ACCEPTED
Copyright © 2008, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Virol. doi:10.1128/JVI.02568-07 JVI Accepts, published online ahead of print on 6 February 2008
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

2
ABSTRACT 1
Human adenovirus serotype 5 (AdH5) vectored vaccines elicit strong immune 2
responses to the encoded antigen and have been used in various disease models. 3
We designed AdH5 vectors expressing antigen under the control of a Human 4
Cytomegalovirus (HCMV) immediate-early promoter containing its Intron A 5
sequence. The transcriptional levels of antigen and immune responses to antigen in 6
vectors with the HCMV promoter with Intron A sequence (LP) was greater than 7
those of AdH5 vectors using HCMV promoter sequence without Intron A (SP). We 8
compared E1E3-deleted AdH5 adenoviral vector, which affords more space for 9
insertion of foreign sequence and showed them to be as immunogenic as E1-10
deleted AdH5 vector. Neutralizing antibodies to AdH5 limit the efficacy of vaccines 11
based on AdH5 serotype and simian adenoviral vectors offer an attractive option to 12
overcome this. We constructed E1E3-deleted human and simian adenoviral vectors 13
encoding a pre-erythrocytic malarial antigen, P. berghei circumsporozoite protein 14
(Pb.CSP). We compared the immunogenicity and efficacy of AdC6, a recombinant 15
simian adenovirus serotype 6, in a murine malaria model to AdH5 and pox viral 16
vectors, MVA and FP9. AdC6 induced sterile protection in 90% of mice in contrast to 17
AdH5 (25%) and pox viral vectors MVA and FP9 (0%) from a single dose. 18
Adenoviral vectors maintained potent CD8+ T cell responses for a longer period 19
after immunization compared to pox viral vectors and mainly induced an effector 20
memory phenotype of cells. Significantly, AdC6 was able to maintain protection in 21
the presence of pre-existing immunity to AdH5. 22
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

3
INTRODUCTION 1
Malaria remains one the of the world’s most important public health problems. It is 2
estimated that 300-500 million clinical episodes occur every year with about a 3
million deaths, primarily in sub-Saharan Africa among children under the age of five 4
years and in pregnant women (54). In spite of current control measures, prevalence 5
as well as resurgence of malaria continues to be evident worldwide mainly due to 6
drug-resistant parasites and insecticide-resistant vectors. The development of a 7
malaria vaccine could control both the transmission and burden of disease. Sterile 8
protection against malaria parasites can be obtained in animals and also in humans 9
by repeated immunization with radiation-attenuated sporozoites (γ-spz) (20, 40).This 10
protective immunity is dependent on CD8+ T cells and IFN-γ production that targets 11
the intra-hepatic schizont stage of the parasite (50). Two candidate 12
vaccines,RTS,S/AS02 and FP9-METRAP/MVA-METRAP, that have completed 13
Phase IIb clinical trials in African children target the pre-erythrocytic stage of the 14
parasite’s life cycle. RTS, S/AS02 is a protein/adjuvant vaccine that encodes most of 15
the circumsporozoite protein (CSP) of Plasmodium falciparum. CSP is the major 16
antigen on the surface of the sporozoite and an important target of the pre-17
erythrocytic immune response (33). This vaccine showed 34% overall efficacy in a 18
phase IIb field efficacy trial conducted in adults in The Gambia (13). In a trial 19
conducted among 1-4 year old children in Mozambique, the vaccine was much more 20
immunogenic than in tropical Africa and was able to decrease clinical malaria 21
incidence rates by 28.9% in one of the two cohorts studied and maintained this 22
efficacy for 18 months (1, 2). Pox viral vectors, MVA and FP9 encoding P.falciparum 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

4
TRAP antigen with a multi-epitope string (ME-TRAP) have been used in a prime-1
boost regime and tested in malaria naive and semi-immune volunteers. Following a 2
sporozoite challenge in naive individuals, the FP9-MVA regime conferred sterile 3
protection in some volunteers and delayed development of parasitemia in others 4
(57). Trials conducted in Gambian adults (38) and Kenyan children (10) showed this 5
vaccine regime to be immunogenic, but it did confer any protective efficacy in a 6
Phase IIb study in Kenyan children in an area of holoendemic transmission, where 7
immunogenicity was unexpectedly low (9). 8
Adenoviral vectors of the human serotype 5 (AdH5) are promising vaccine vector 9
candidates that induce a potent cellular and humoral immune response and have 10
demonstrated protective efficacy in different disease models (19, 47). This vaccine 11
vector has been previously tested and shown efficacy in mouse malaria models (28, 12
45). Most adenoviral vectors currently used as vaccine carriers have a deletion in 13
the E1 region of their adenoviral genome and employ the HCMV immediate early 14
promoter to drive antigen expression. The deletion of the E1 region of the adenoviral 15
genome inhibits replication and increases space for foreign DNA insertion. To 16
incorporate larger foreign DNA sequences, adenoviral vectors with E1 and E3 17
regions deleted have been developed. However, few studies have compared the 18
effect of different deletions on the immunogenicity induced by the adenoviral vaccine 19
vector. To improve the immune response induced to the encoded antigen, studies 20
have attempted to optimize the promoter used to drive antigen expression. The 21
commonly used HCMV immediate early promoter is one of the strongest promoters 22
known to date and consists of the enhancer/promoter sequence of the gene. DNA 23
vaccines constructed with the HCMV promoter along with its Intron A sequence have 24
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

5
been shown to be immunogenic in animal and human studies (36, 48, 49) but few 1
studies have compared the effect of addition of full length Intron A to the HCMV 2
promoter sequence. 3
A major limitation of AdH5 in humans is the high prevalence of neutralizing 4
antibodies (55) to the virus caused by frequent childhood infections. To circumvent 5
interference by pre-existing immunity, rare adenovirus serotypes that do not circulate 6
in human populations have been developed as vaccine vectors. One strategy is to 7
use human adenovirus serotypes that have low prevalence in human populations 8
such as Ad35 and Ad11 (8, 34). Unfortunately, these vaccine vectors induce weaker 9
responses than AdH5. A different approach is the use of chimpanzee adenoviral 10
vectors that do not circulate in human populations. Studies have demonstrated the 11
immunogenicity and efficacy of simian adenoviral vectors in disease models of HIV, 12
Ebola and SARS (32, 44, 63). 13
14
Our study describes the improved design of a HCMV promoter to drive antigen 15
expression from an adenoviral vector and further compares the immune responses 16
induced by an E1 region deleted vector to an E1E3 region deleted adenoviral vector. 17
We describe for the first time the use of a simian adenoviral vector AdC6 as a pre-18
erythrocytic malaria vaccine in a mouse malaria model. We show that AdC6 can 19
induce very strong CD8+ T cell responses compared to AdH5 and provide high 20
levels of sterile protection to a potent sporozoite challenge. We show that in 21
conditions of pre-existing immunity to AdH5 which abrogated protection induced by 22
AdH5 vector, AdC6 was still able to maintain protection. 23
24
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

6
1
MATERIALS AND METHODS 2
Cell Lines 3
Human Embryonic Kidney (HEK) 293 cells were propagated in DMEM 4
supplemented with glutamine, antibiotic and 10% FCS (Sigma). 5
Viral vectors 6
A shuttle vector pENTR4.LP was constructed by inserting the enhancer/promoter 7
sequence with exon 1 and intron A sequence of HCMV Immediate-early gene (nt 8
163-2091 GenBank accession number M60321), a multiple cloning site and bovine 9
growth hormone poly A sequence from pSG2 plasmid(36) between the 10
recombination sequences in pENTR4 (Invitrogen Life Technologies, Inc). 11
Plasmodium yoelii merozoite surface protein 142 (MSP-142) and Mycobacterium 12
tuberculosis 85A gene were cloned into the multiple cloning site of either pENTR4 or 13
pENTR4.LP shuttle vectors. This was then recombined into E1 and E3 deleted AdH5 14
adenoviral genome vectors from the Virapower adenoviral expression System 15
(Invitrogen Life Technologies, Inc). pENTR4.LP was recombined with pAd/PL-DEST 16
to generate AdH5 adenoviral genome vectors containing the HCMV promoter (LP), 17
antigen and BHG Poly A sequence from pENTR4.LP. pENTR4 vectors containing 18
antigen were recombined with pAd/CMV/V5/DEST generating AdH5 adenoviral 19
genome vectors containing antigen driven by the HCMV IE promoter (SP) in 20
pAd/CMV/V5/DEST. The difference in the two promoters is shown in Fig.1A. For 21
viral rescue, the vector was transfected into HEK 293 cells, expanded and purified 22
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

7
using Adenopure purification column (Puresyn Inc.) or CsCl gradient centrifugation. 1
The number of viral particles(vp) was determined by spectrophotmetry as described 2
previously(35). PFU were determined by plaque assay and ranged between 1PFU 3
per 20 – 500 vp. E1-deleted AdH5 adenoviral vector and the E1E3-deleted simian 4
adenoviral vector (AdC6) was constructed and propagated as described previously 5
(44). Construction of the AdH5 (28), MVA and FP9 (3) viral vectors expressing 6
Plasmodium berghei CSP under the control of HCMV/IE promoter containing Intron 7
A(LP) in the case of AdH5, and the vaccinia virus p7.5 promoter for MVA and FP9, 8
has been described earlier. 9
10
Reverse Transcriptase quantitative Real Time PCR 11
293A cells were infected at a MOI of 50 with adenoviral vectors expressing antigen 12
85A or MSP-142 under the control of either the Short Promoter (SP) or the Long 13
Promoter (LP). Cells were harvested 48h post –infection, centrifuged and used to 14
extract mRNA. mRNA was extracted using QIAamp RNA Blood Mini Kit (Qiagen) 15
following the manufacturer’s instructions. RNA was then reverse transcribed into 16
cDNA using the Omniscript Reverse Transcriptase Kit (Qiagen) (containing 17
Omniscript Reverse Transcriptase enzyme with its buffer, dNTP mix) and oligo-dT 18
primer. Real Time PCR on cDNA was performed on the Rotorgene 3000(Corbett 19
Research, Sydney, Australia) machine using the Quantitect SybrGreen PCR kit 20
(Qiagen). The Sybr mix contained HotStar Taq DNA polymerase, SybrGreen I dye, 21
optimized SybrGreen buffer, 5mM MgCl2 and dNTP mix including dUTP. Forward 22
primer MSP-142 SS2 (5’-CATGGATGGTATGGATTTATTAGGTG-3’) and reverse 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

8
primer MSP-142 SS2 (5’-TCCGTACTTTCCGCATTTTGA-3’) were used to amplify a 1
fragment of 237 base pairs in the MSP-1 42 antigen. Forward primer 85AU SS (5’-2
GCG CCG TCA CGG GTA TGT- 3’) and reverse primer 85AL SS (5’- GCA CCA 3
CCA CTT TGGAAT TG-3’) were used to amplify a fragment of 196 base pairs in the 4
85A antigen. A 243 base pairs fragment from the region of the adenoviral gene 5
AdE4orf1 was also amplified from each sample using the forward primer AdE4orf F1 6
(5’- GGTAGCGCGGGTCTCTGTCTCA-3’) and reverse primer AdE4orf R1 (5’-7
GGGGGCGCCTGGATGCT-3’). Real-time PCR amplification was performed in 25 µl 8
volume containing Quantitect Mastermix Kit (Qiagen), 100 nM each primer and 1µl 9
cDNA. The thermal cycling conditions were 95 °C for 15 min, 40 cycles of 95 °C for 10
10 seconds, annealing at55 °C for 15 seconds and extension at 72° C. At least two 11
“no-template-control” and plasmid encoding the antigen as positive control was used 12
in every PCR run. Standard curves of quantified and diluted PCR product for each 13
primer pair were generated and used in different experiments to calculate the cDNA 14
amount from each sample. A standard curve was rejected if the correlation 15
coefficient of the trend line was < 0.95. The numbers of copies of the RT-PCR 16
standards were calculated and used to compare antigen mRNA expression levels. 17
To normalize differences in RNA quality between the individual samples and in RNA 18
loading for the RT-PCR procedure, the antigen mRNA expression level for a 19
particular sample was defined as its antigen gene expression level divided by its 20
Ad.E4orf1gene expression level. 21
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

9
Mice and immunizations 1
Female BALB/c mice 4 to 6 week of age were purchased from the Biomedical 2
Services Unit at the John Radcliffe Hospital, Oxford with all animals used in 3
accordance with the terms of the U.K. Home Office Animals Act Project License. 4
Immunizations were performed intradermally, which has previously been shown to 5
elicit better immunogenicity when compared to other routes e.g. sub-cutaneous, 6
intramuscular (28). MVA.PbCSP or FP9.Pb.CSP were administered at a dose of 7
1x107 pfu, and all adenoviruses at a dose of 1x1010 or 1x109 viral particles (vp). All 8
vectors were resuspended in endotoxin-free PBS prior to immunization. 9
Peptides 10
The Pb9 (SYIPSAEKI) peptide(28) that carries the immunodominant H-2Kd-restricted 11
epitope of P.berghei CSP, the p11(EWYDQSGLSVVMPVGGQSSF) and p15-12
(TFLTSELPGW LQANRHVKPT) peptides of 85A carrying CD8 and CD4 epitopes 13
respectively(22), have been previously described. T cell epitopes in MSP-142 are 14
mainly contained in the MSP-133 region. CD8+ T cell epitopes within MSP-133 were 15
predicted from known H-2 Class I binding motifs using the epitope prediction section 16
of the SYFPEITHI website 17
(http://www.syfpeithi.de/Scripts/MHCServer.dll/EpitopePrediction.htm) developed at 18
the University of Tübingen. The six strongest candidates for BALB/c (H-2d) mice 19
(157, 206, 178, 28, 60, and 120) were selected. Epitope B80 is the strongest 20
predicted epitope for C57BL/6 mice, but was also a weaker candidate for BALB/c 21
mice. Cm15 and Cm21 are known T cell 20mer “protective” epitopes within MSP-133 22
from BALB/c mice, recognized by CD4+ effector T cell lines capable of delaying 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

10
growth of blood-stage P. yoelii YM (59). Table 1 gives the sequence and predicted 1
strength of MSP-142 epitopes. 2
Ex vivo IFN-γ ELISPOT 3
ACK-treated PBMCs or spleens were cultured for 18-20 hours on IPVH-membrane 4
plates (Millipore) with the peptide at a final concentration of 1µg/ml. ELISPOT was 5
performed as previously described (37) . 6
Intracellular Cytokine Staining 7
ACK-treated splenocytes were incubated for 5 hours in presence of 1µg/ml Pb9 and 8
2µl/ml Golgi-Plug (BD). Intracellular cytokine staining (ICS) was performed with BD 9
cytofix/cytoperm plus kit according to the manufacturer’s instructions. Splenocytes 10
were stained with a suitable combination of fluorochrome-conjugated antibodies, 11
specific for CD8 (clone 53-6.7, eBioscience), IFN-γ (clone XMG1.2, eBioscience), 12
CD27 (clone LG.7F9, eBioscience), CD43 (clone 1B11, BD/Pharmingen), CD127 13
(clone A7R34, eBioscience), IL-2 (clone JES6-5H4, eBioscience), TNFα (clone MP6-14
XT22,eBioscience) ,mouse isotype controls IGg2a (eBR2a, eBioscience), 15
CD16/CD32 Fcγ III/II Receptor (2.4G2, BD/Pharmingen), anti-Granzyme B (clone 16
GB12, Caltag), IgG1 isotype control (Caltag). When CD62L (clone MEL-14, 17
eBioscience) was used, stimulated cells were incubated with TAPI-2 peptide 18
(Peptides International, USA) at a final concentration of 250 µM to prevent CD62L 19
shedding from the cell surface. Flow cytometric analyses were performed using a 20
FACSCanto (BD Biosciences) and data were analyzed with Flow Jo (Tree Star) 21
software. 22
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

11
Evaluation of antigen-specific CD8+ T-cell response by flow cytometry 1
The frequency of IFN-γ + CD8+ T cells was calculated by subtracting the values from 2
the unstimulated control, which never exceeded 0.1% in any of the experiments. For 3
the phenotypic makers investigated, each marker was compared to an isotype 4
control. 5
Parasite challenge 6
Plasmodium berghei (ANKA strain clone 234) sporozoites (spz) were isolated from 7
salivary glands of female Anopheles stephensi mosquitoes. Parasites were 8
resuspended in RPMI-1640 medium with each mouse receiving a total of 1,000 spz 9
via the i.v. route. Blood samples were taken daily from day 5 to 20; smears were 10
stained with Giemsa and screened for the presence of blood stage parasites. 11
Survival was defined as complete absence of parasites in blood. 12
Statistical analysis 13
Statistical significance of flow cytometry samples were analyzed with either a one or 14
two-way ANOVA and a Bonferroni post-test. All statistic tests were performed using 15
GraphPad Prism version 4.03 for Windows, GraphPad Software, San Diego 16
California, USA, www.graphpad.com. 17
18
19
20
21
22
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

12
RESULTS 1
3.1 Effect of Intron A on the strength of HCMV IE promoter 2
We compared antigen transcript levels driven by the HCMV IE promoter with Intron 3
A (LP) or the promoter without Intron A (SP) (Fig.1A) by reverse transcriptase RT-4
PCR. Cell cultures of HEK 293 cells were infected at a MOI of 50 with human 5
adenovirus serotype 5 (AdH5) expressing either MSP1-42 antigen or 85A antigen 6
under the control of the CMV I/E promoter containing Intron A (Ad.H5.MSP1-42.LP, 7
Ad.H5.85A.LP) or the CMV IE promoter without Intron A (Ad.H5.MSP1-42.SP, 8
Ad.H5.85A.SP). Cells were harvested 24 hours later and quantitative real time 9
reverse transcriptase PCR was undertaken for the specific antigen and AdE4orf1 10
gene to quantify gene mRNA levels. Antigen mRNA levels were normalized to 11
Ad.E4orf1 mRNA and compared. For both antigens, the HCMV IE promoter with 12
Intron A (LP) directed a higher level of transcription than the promoter without Intron 13
A. Ad.MSP1-42 LP had 15 fold higher levels of gene expression than Ad.MSP1-14
42SP (Fig.1B) while Ad.85ALP showed 2 fold higher expression than Ad.85ASP 15
(Fig.1C). We further tested whether this difference in transcriptional activity of the 16
antigen resulted in a more immunogenic vaccine. Balb/c mice were immunized with 17
1010 vp of AdH5.MSP-142 .SP, AdH5.MSP-142 .LP, AdH5.85A.LP or AdH5.85A.SP. 18
Immune responses in the spleen were measured by ex-vivo IFN-γ ELISPOT 14 days 19
after immunization. The immune responses were stronger for all defined epitopes in 20
MSP-142 and were significantly stronger to defined CD8 epitopes in MSP-142 21
(Fig.1D) in vaccine constructs driving antigen expression from the HCMV promoter 22
with the Intron A compared to constructs using the HCMV promoter without Intron A 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

13
to drive antigen expression. Although no difference in the immune response to 1
epitopes in 85A was seen with AdH5.85A.LP compared to AdH5.85A.SP (Fig.1E) 2
when immunizing mice with 1010 vp lowering the immunizing dose to 109 vp showed 3
a significant difference in the immune response to the CD8 epitope in 85A (Fig.1F). 4
The data suggests that antigen under the control of the HCMV promoter with Intron 5
A (LP) is expressed at higher levels and results in stronger immune responses 6
compared to the HCMV promoter without Intron A. . 7
8
3.2 Comparison of E1 region deleted or E1E3 region deleted adenoviral 9
vectors 10
Most adenoviral vectors currently in use have the E1 region of adenovirus deleted 11
which makes the virus replication-incompetent in mammalian cells. The E3 region of 12
the adenovirus expresses a number of proteins that are involved in a complex 13
interplay functioning mainly to subvert the host immune system. We compared the 14
immunogenicity of an AdH5 vector with only E1 region deleted to one that had E1 15
and E3 regions deleted. Balb/c mice were immunized with 1010 vp of E1-deleted 16
AdH5.85A.LP and E1E3-deleted AdH5.85A.LP and immune responses were 17
measured in the spleen two weeks after immunization. While, there is a trend 18
towards the E1E3 deleted vector inducing stronger immune responses to antigen 19
85A epitopes, these responses were not significantly different from that induced by 20
E1 deleted AdH5.85A.LP (Fig.2). The advantage of using E1E3-deleted vectors is 21
the ability to incorporate larger sequences of foreign DNA and potentially diminished 22
anti-vector immunity (52). While E1-deleted vectors can accommodate a 5 kb foreign 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

14
sequence, E1E3-deleted adenoviral vectors allow for the insertion of up to 7.5 kb of 1
foreign DNA and therefore we used E1E3 deleted adenoviral vectors for further 2
studies. 3
3.3 Evaluation of antigen specific CD8+ T cell responses 4
We constructed E1E3-deleted adenoviral vectors expressing the Plasmodium 5
berghei circumsporozoite protein (Pb.CSP) under the control of HCMV promoter with 6
Intron A. We evaluated the immunogenicity and immune kinetics of recombinant 7
Human adenovirus serotype 5 (AdH5), Simian adenovirus serotype 6(AdC6) and 8
pox viral vectors Modified Vaccinia Ankara (MVA) and Fowlpox (FP9) expressing 9
Pb.CSP by measuring the IFN-γ response to previously defined CD8+ specific T cell 10
epitope Pb9. Of all the vectors, AdC6 was the most immunogenic vaccine and FP9 11
was the least immunogenic (Fig. 3A). The adenoviral vectors were more 12
immunogenic than the pox viral vectors at all time points measured except for one 13
week after immunization when the immune response induced by MVA.PbCSP was 14
the strongest. The kinetics of the immune response varied between the pox viral and 15
adenoviral vectors. While pox viral vectors MVA and FP produced an immune 16
response which peaked one week after vaccination and contracted rapidly by 14 17
days, the immune response induced by the adenoviral vectors peaked at 2 weeks 18
after immunization and contracted more slowly (Fig.3A). Between the adenoviral 19
vectors, AdC6 was significantly more immunogenic and was able to maintain a 20
higher frequency of CD8+IFN-γ + up to at least 140 days after immunization. 21
Observing the difference in the immune kinetics induced by different viral vectors, 22
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

15
we further extended our study to the difference between viral vectors in their 1
development of effector and memory phenotype of CD8+IFN-γ + cell populations. 2
3
3.4 Development of Effector CD8+ T cell response. 4
The acquisition of effector functions was determined by the expression of phenotypic 5
markers - CD43 and Granzyme B. CD43 expression has previously been shown to 6
be upregulated during the effector phase of the CD8 response. Effector cells 7
undertake their cytolytic functions through effector molecules like Granzyme B (GrB) 8
and TNF-α which are involved in direct CD8+ T cell mediated killing of infected cells. 9
In general, adenoviral vectors induced a significantly higher percentage of CD8+IFN-10
γ+CD43high cells over the entire course of the immune response when compared to 11
the poxviral vectors. Irrespective of the vector used, the percentage of CD8+IFN-12
γ+CD43high cells decreases with time indicating a transition to the memory phase of 13
the immune response, but the kinetics of this transition varied between poxviral and 14
adenoviral vectors. At the peak of the immune response induced by pox viral vectors 15
(day 7), 50% of CD8+IFN-γ+ cells were CD43high but as early as two weeks post-16
vaccination this frequency dropped to 20%, suggesting a rapid transition towards the 17
memory phase (Fig.3B). In comparison, mice immunized with the adenoviral vectors 18
still retained a high percentage of CD8+IFN-γ+ CD43high cells even until 56 days post-19
prime (40% for AdC6 and 35% for AdH5) compared to the pox viral vectors (Fig.3B) 20
and it took 140 days after immunization for the levels of CD8+IFN-γ+ CD43high cells to 21
drop to about 20%. CD8 T cells induced by both the adenoviral vectors expressed 22
greater levels of GrB than the pox viral vectors at day 14 indicating a rapid loss of 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

16
cytolytic molecules following pox viral vaccination. However, by day 28 the 1
adenoviral vector-induced cells also had low levels of GrB expression 2
(Fig.3C).These results demonstrate that the adenoviruses are able to maintain an 3
effector response with preservation of cytolytic molecules for longer periods of time 4
compared to the pox viral vectors. 5
6
3.5 Development of memory CD8+ T cells. 7
One of the main objectives of any vaccination regime is the generation of memory 8
CD8+ T cells that are capable of persisting in vivo and expanding rapidly upon 9
encounter with pathogens thus affording protection. It has been proposed that the 10
development of intra-hepatic effector and central memory phenotype of cells is 11
crucial to protection against malaria sporozoite challenge (11). Different molecules 12
have been suggested to correspond to different sub-types of memory cells (7). In 13
this study we chose to differentiate Effector (TE), Effector Memory (TEM) and Central 14
Memory (TCM) cells on the basis of phenotypic markers CD127, CD62L, IL2 and 15
CD27 to determine whether individual vectors induced different memory populations. 16
During the first 14 days post immunization, CD8+ IFN-γ+ cells generated by either 17
FP9 or MVA displayed a CD62L-, CD127+ or a CD62L-CD127- phenotype indicating 18
mainly a TEM or a TE phenotype, respectively. During the same period, the majority 19
of cells generated by adenoviral vectors were TE cells with a CD127-CD62L- 20
phenotype (Fig. 4A). By 56 days post immunization, the majority of CD8+IFN-γ+ cells 21
induced by pox viral vectors were of a CD62L+CD127+ phenotype and 40% of 22
antigen-specific cells were producing IL2 indicating development of TCM cells. On 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

17
the other hand, by day 56 post-prime the majority of CD8+IFN-γ+ cells induced by 1
adenoviral vectors displayed a TEM phenotype (CD62L-, CD127+ and low 2
percentage of IL-2 producing cells) and only 8% of CD8+IFN-γ+ cells showed a TCM 3
phenotype (Fig. 4A,B). Interestingly, while AdH5 was able to generate 46% of 4
CD8+IFN-γ+ cells with a TCM phenotype by day 140, the majority of cells induced by 5
AdC6 still remained of a TEM phenotype, suggesting the delayed development of 6
central memory cells following AdC6 immunization (Fig. 4A). 7
CD27 is a surface marker that is down regulated following repeated antigenic 8
stimulation by CD70. Initially the levels of CD8+IFN-γ+ CD27+ cells were close to 9
40% in mice vaccinated with adenoviral vectors but by day 28 post-prime this 10
frequency dropped to 20% and only began to return to its original levels by day 110. 11
This is in stark contrast to pox viral vectors which showed a steady increase in the 12
percentage of CD8+CD27+ cells over time. Since CD8+ CD27- cells are maintained 13
in response to persistent antigenic stimulation (4), this may suggest that prolonged 14
antigen stimulation was occurring in response to the adenoviral immunization. 15
Antigen stimulation is thought to hinder the development of central memory cells 16
while maintaining cells in an effector state and interestingly, this rise in CD27+ cells 17
corresponds to the beginning of development of cells with a TCM phenotype. In 18
summary, these results demonstrate that vaccination with adenoviral vectors using 19
the Pb.CSP insert maintains a predominantly TEM response (as evidenced by a 20
CD62L-, CD127+, IL-2low phenotype) up to 8 weeks after immunization and delays 21
the development of central memory cells (as evidenced by CD62L+CD127+, IL2high 22
phenotype) as compared to poxviral vectors. 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

18
1
3.6 Efficacy of Adenoviral vectors against sporozoite challenge 2
To test the efficacy of these vectors, mice immunized with viral vectors were 3
challenged with 1000 sporozoites of P.berghei. Mice were challenged at 7 days, 14 4
days and 56 days after immunization to investigate efficacy data at the peak of the 5
effector response and during the memory phase. Both AdC6 and AdH5 afforded 6
significant protection (p=0.0001 and p=0.01) when mice were challenged 14 days 7
after immunization. AdC6 protected 90% of mice compared to only 25% by AdH5 8
while both pox viral vectors failed to offer any sterile protection (Table 2). Mice were 9
also challenged 7 days after immunization corresponding to the peak of the immune 10
response for pox viral vectors but neither MVA nor FP9 conferred protection at this 11
time point. To investigate durability of protection mice were challenged 8 weeks after 12
immunization and only C6 afforded a statistically significant degree of 13
protection(p=0.015) and was able to protect 36% of the challenged mice. 14
15
3.7 Effect of pre-existing immunity on immunogenicity and efficacy of 16
adenoviral vectors. 17
45% of humans have neutralizing antibodies to human adenovirus serotype 5 (55) 18
and limit the efficacy of AdH5 vaccine vectors (63). Simian adenoviruses offer a 19
promising strategy to circumvent this problem. To test the efficacy of AdH5 and 20
AdC6 vectors in the presence of pre-existing immunity, we immunized mice with 21
AdH5 or AdC6 expressing an unrelated antigen (85A) and 6 weeks later immunized 22
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

19
them with vectors containing Pb.CSP. Titers of neutralizing antibodies to the AdHu5 1
and AdC6 vector in the sera of mice were tested before immunization with vectors 2
containing Pb.CSP. The titers were found to be 1:320 – 1:400, which is higher than 3
titers commonly found in human adults. In the presence of pre-existing immunity to 4
AdH5, immunization with AdH5.PbCSP led to a significant decrease in the immune 5
response to Pb9 (p<0.01) (Fig.5A) and abrogation of protection (Fig.5B) as 6
compared to AdH5.PbCSP in the absence of pre-existing immunity while 7
AdC6.PbCSP afforded 66% protection and significantly better immune responses 8
compared to AdH5.PbCSP (p<0.001) in the presence of pre-existing immunity to 9
AdH5. In mice with pre-existing immunity to AdC6, immunogenicity of AdC6.PbCSP 10
was significantly reduced (p<0.01) while the immune response induced by 11
AdH5.PbCSP was not reduced as compared to the immune response induced by 12
the respective vaccines in the absence of pre-existing immunity to AdC6. This 13
decrease in immunogenicity led to a decrease in protective efficacy as AdC6 was 14
only able to protect 33% of mice in the presence of pre-existing immunity to 15
AdC6.while AdH5.PbCSP provided 33% protection (Fig.5A, B). These results further 16
emphasize the problem of pre-existing immunity in the context of adenoviral 17
vaccines and demonstrate the efficacy of AdC6 vaccine in the presence of pre-18
existing immunity to human adenoviral serotype 5. 19
DISCUSSION 20
Replication-deficient adenoviral vectors have been used as a vaccine strategy to 21
deliver antigen in different disease models. A variable influencing the capacity for 22
adenoviral vectors to be successful as vaccines could be an ability to generate high 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

20
level expression of the antigen for induction of a strong immune response. The gene 1
encoding the immediate-early 72-kDa protein of human cytomegalovirus (HCMV) is 2
transcribed by one of the strongest enhancer/promoters currently known (14) and is 3
able to induce high levels of transgene expression in a wide variety of cells types 4
after both transient and stable transfections(14, 26). Therefore, it is currently the 5
most widely used viral promoter used in vaccine and gene therapy applications. 6
However, the expression levels induced in vivo are often suboptimal for therapeutic 7
applications. While the strength of the CMV I/E promoter lies in its enhancer region 8
which contains binding sites for transcription factors of the NF-κB, cAMP responsive 9
binding protein and the nuclear factor-1 families, the Intron A region of the HCMV 10
promoter has been investigated for its regulatory role on the enhancer region and 11
shown to enhance gene expression in a transient transfection system (18). The 12
enhancing effect of Intron A on HCMV promoter directed transgene expression led 13
to the addition of the Intron A sequence to the HCMV enhancer/promoter sequence 14
for controlling antigen expression in DNA vaccines (42, 53) and viral vectors (28). 15
The Intron A sequence used in the DNA vaccine constructs was a partial intron 16
sequence. Here we demonstrate that addition instead of the full Intron A sequence 17
to the HCMV promoter to drive antigen expression in adenoviral vectors can 18
significantly improve immunogenicity to the antigen compared to similar HCMV 19
promoter without the Intron A sequence. Adenoviral vectors expressing Plasmodium 20
yoelii MSP-142 antigen and Mycobacterium tuberculosis 85A antigen from HCMV 21
promoters either with or without the Intron A sequence were compared. These 22
antigens containing defined murine CD4 and CD8 epitope were chosen for vaccine 23
constructs to allow the measurement of CD4 and CD8 specific immune responses. 24
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

21
HCMV promoter with Intron A sequence showed higher levels of mRNA expression 1
in vitro for both antigens compared to HCMV promoter without Intron A. 2
Furthermore, AdH5 vectors with HCMV promoter with Intron A are capable of 3
inducing higher levels of IFN-γ producing antigen specific CD8 and CD4 T cells than 4
vectors with HCMV promoter without the Intron A sequence. Immune responses to 5
epitopes of MSP-142 antigen were significantly improved by the addition of Intron A 6
to the promoter while peptide specific responses to antigen 85A did not showed any 7
difference between vaccine constructs containing promoters with and without Intron 8
A. However, lowering the immunizing dose of the Ad.85A vaccines unmasked the 9
enhanced immunogenicity of the vaccine with HCMV promoter containing Intron A 10
compare to that without Intron A (Fig.1F). The difference in mRNA transcription 11
levels between AdH5.85A.LP and AdH5.85A.SP are not as significant as those with 12
AdH5.MSP-142 vectors and this may explain the lack of difference in immune 13
responses between the two AdH5.85A vaccines. In addition, we also suggest that a 14
high immunizing dose of AdH5.85A provides a level of immunogenicity that cannot 15
be significantly enhanced by the addition of the Intron A to the promoter. Our 16
findings suggest that the HCMV promoter with Intron A may be preferentially 17
beneficial in vaccines or systems with low levels of antigen expression like MSP-142 18
in which even a high immunizing dose is unable to compensate for low expression 19
levels of antigen from HCMV promoter without Intron A. While the differences 20
between the two promoter constructs (Fig.1A) include a larger regulatory sequence, 21
an exon sequence and the full length Intron A sequence we suggest that the 22
difference in levels of expression and immunogenicity are most likely linked to the 23
effect of Intron A on enhancing antigen expression. Similar use of introns in plasmids 24
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

22
was shown to increase levels of expression in various systems (16, 31). Intron A of 1
HCMV promoter contains a Nuclear factor 1 binding site that may act as a 2
downstream transcriptional activator, however it is more likely that the positive 3
effects of Intron A arise from a combination of mRNA stabilization and increased 4
transcriptional activity of Intron A (18). 5
Packaging capability of the adenoviral capsid is an important consideration when 6
constructing adenoviral vectors (12, 56). Foreign sequences of up to 1.8 kb can be 7
inserted into the adenoviral genome without necessitating deletions. The deletion of 8
the E1 region renders the viral construct replication-incompetent and allows an extra 9
3.5 kb of foreign DNA to be added. The E3 region of the adenovirus genome while 10
dispensable for growth of the virus encodes a series of proteins that function as 11
immuno-modulatory agents (23) and has been deleted in vaccine vectors (62) to 12
increase the cloning capacity of the adenoviral vector to 7.5kb. We demonstrate that 13
there is no significant difference in the immune response to antigen delivered by an 14
E1-deleted or an E1E3-deleted adenoviral vector (Fig.2) after a single dose of the 15
vaccine. Our promoter of choice, the HCMV promoter with Intron A (LP) is 1.9 kb 16
long and its use in an E1-deleted adenoviral vector would permit the insertion of an 17
antigen not greater than 3.4 kb in size. Our results demonstrate that an E1E3-18
deleted vector is as potent as an E1-deleted vector and the E1E3-deleted vector 19
with the LP to drive antigen would allow an antigen of up to 5.6 kb to be inserted. 20
The use of an E1E3-deleted adenoviral vector with the HCMV promoter containing 21
Intron A combines a potent promoter and enough free space in the adenoviral 22
genome to insert large antigens. Further studies comparing the HCMV promoter 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

23
containing full length Intron A with HCMV promoter with shortened versions of Intron 1
A need to be undertaken. 2
A malaria vaccine against the pre-erythrocytic stage of the parasite would be able to 3
control infection thereby preventing any symptoms of the disease. While sterile 4
protection induced by strong IFN-γ+CD8+ T cells has been achieved using γ-5
irradiated sporozoites, it would be very difficult to deploy such a vaccine system 6
which required liquid nitrogen cryopreservation, in a worldwide immunization 7
program. Thus, the search for an efficacious and immunogenic vaccine delivery 8
system that can be deployed in humans continues. The circumsporozoite protein is 9
the most abundant antigen expressed on the surface of the sporozoites and has 10
been the main target antigen in the development of pre-erythrocytic malaria 11
vaccines. Immunization of mice with DNA vaccines (27) or recombinant viruses (45, 12
57) has previously been shown to elicit CD8+ T cell responses against epitopes in 13
CSP. 14
Adenoviral vectors are potent inducers of a cellular immune response and have 15
conferred protection against a malaria sporozoite challenge when used either as a 16
single dose (P.yoelii) (45) or in prime-boost regimens in combination with pox viral 17
vectors (P.berghei) (28). However, in the only previous study of adenoviral vectors in 18
the P.berghei model of malaria, protection by homologous prime-boost using AdH5 19
was minimal (28). Human serotype 5 adenovirus is a ubiquitous virus causing 20
recurrent asymptomatic infection and subsequently resulting in over one-third of the 21
adult human population carrying high levels of neutralizing antibodies to AdH5. This 22
antibody mediated pre-existing immunity has been shown to reduce the efficacy of 23
H5 adenoviral vaccine vectors in animal models (17). To circumvent this issue, 24
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

24
chimpanzee adenovirus serotypes which do not circulate in human population have 1
been developed as vaccine vectors. Previous studies have shown the ability of such 2
recombinant simian adenoviral vectors to elicit potent B and CD8+ T-cell-mediated 3
immune responses in mice and primate models of rabies (60), SARS (63) and HIV 4
(44). This paper reports the use of a simian adenoviral vector using the HCMV 5
promoter with Intron A (LP) to drive expression of P. berghei CSP and compares the 6
immunogenicity and efficacy of this novel simian adenoviral vector AdC6, human 7
adenoviral vector AdH5 to MVA and FP9, pox viral vectors that have been widely 8
used in clinical trials. 9
10
First, we investigated the immunogenicity and kinetics of the antigen specific CD8+ 11
response following single immunization with the viral vectors. The simian adenovirus 12
vector AdC6 was the most immunogenic vaccine, eliciting the strongest peptide 13
specific CD8+IFN-γ T cell response. The response elicited by the adenoviral vectors 14
peaked around 2 weeks in contrast to the pox viral vectors which peaked by day 7. 15
While the response to pox viral vectors contracted rapidly after day 7, the pb9 16
specific response by adenoviral vectors contracted more slowly with a high 17
frequency CD8+IFN-γ+ cells remaining even 56 days after immunization with AdC6 18
(Fig.3A). Characterization of the immune response using phenotypic markers for 19
effector and memory CD8T cells showed that adenoviral vectors were able to elicit a 20
much more potent effector response as measured by the levels of CD43 and 21
Granzyme B expression than the pox viral vectors. The effector response induced by 22
adenoviral vectors was also maintained for a longer period of time compared to the 23
pox viral vectors (Fig.3B, C). We further analyzed the phenotype of memory T cells 24
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

25
generated in response to the different viral vectors. Based on the expression of 1
CD127 and CD62L, antigen specific CD8+ T cells can be classified into Effector T 2
cells TE (CD127-CD62L-), Effector Memory TEM (CD127+CD62L-) and Central 3
memory T cells TCM (CD127+CD62L+) with each phenotype having different 4
functional characteristics (5). At the peak of the immune response with adenoviral 5
vectors (day 14 post-prime) about 60% of cells were CD8+CD127-CD62L- TE cells. 6
By 56 days post-prime the majority of antigen specific CD8+ cells in mice immunized 7
with pox viral vectors were of the CD127+CD62L+ TCM phenotype, while adenoviral 8
vectors mainly induced CD8+CD127+CD62L- TEM cells at the same time point. TCM 9
cells constituted the majority of antigen specific CD8+ T cells only 140 days after 10
immunization with AdH5 and only 22% of cells following AdC6 immunization. The 11
development of memory T cells is thought to depend on antigen exposure, type of 12
antigen and persistence of antigen (6). To further understand the delayed 13
development of TCM following adenoviral immunization, we analyzed the expression 14
of CD27 on antigen specific cells. CD27 is a member of the TNFR family and in mice 15
repeated antigenic stimulation leads to the loss of CD27 from CD8+ T cell surface 16
and these CD8+CD27- cells express high levels of cytotoxic effector molecules like 17
Granzyme B and perforin (4). Following immunization with adenoviral vectors 20% of 18
antigen specific CD8+IFN-γ+CD27+ cells are maintained until 56 days post-prime. 19
This is different to pox viral vectors which induce a high percentage of CD27+ cells 20
as early as 14 days after immunization. Our results demonstrating the delayed 21
development of TCM cells, maintenance of CD43high and TEM cells in combination with 22
the low frequency of CD27+ cells suggests that adenoviral vectors may cause 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

26
prolonged expression of antigen thereby delaying contraction of the immune 1
response and subsequently maintaining an effector response for an extended time. 2
This feature of adenoviral vectors make them especially attractive as vaccine 3
vectors for disease states that require high levels of effector response for protection. 4
Studies conducted in various disease models have underlined the importance of TEM 5
and TCM cells in conferring protection(58). It has also been shown that sporozoite-6
induced protection against P.berghei sporozoite challenge was effected mainly by 7
intra-hepatic TEM cells and not TCM cells (11). In this study, both AdH5 and AdC6 8
conferred protection (25% and 90% respectively) to a stringent P.berghei challenge 9
2 weeks after immunization. It is interesting to note the striking difference in 10
protective efficacy between the two adenoviral vectors when challenged 14 days 11
after immunization. At this time point there is no difference the percentage of antigen 12
specific TE and TEM cells (Fig.3 and Fig.4) or the percentage of CD8+IFN-γ+ cells 13
secreting effector cytokines such as Granzyme B and TNF-α (unpublished data). It is 14
therefore likely that this difference in the protective efficacy is a result of the 15
enhanced immunogenicity of AdC6 compared to AdH5. However, pre-erythroycytic 16
stage protection is also mediated through factors other than IFN-γ, like Nitric Oxide, 17
IL-12 or Natural Killer cells(29) and it is probable that AdC6 and AdH5 may differ in 18
their induction of these factors as well. When the challenge was performed at 8 19
weeks after immunization, AdC6 was able to protect 36% of the mice while AdH5 20
showed only 14% protection. Neither pox viral vector was able to protect against 21
sporozoite challenge, even when the challenge was undertaken at the peak of the 22
effector response (day7) for these vectors. It is encouraging to note that AdC6 was 23
able to protect 36% of mice challenged 8 weeks after immunization. Long-lasting 24
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

27
sterile protection with a single vaccine dose has been achieved only with whole 1
parasite vaccine strategies, either by irradiation or by genetic attenuation. The 2
deployment of these whole parasite vaccine strategies worldwide would be 3
logistically very problematic, so a subunit vaccine that confers durable protection 4
after a single dose is a promising finding. Sterile protection against rodent malaria 5
using adenoviruses has been achieved in prime-boost regimes involving AdH5-6
vaccinia virus (P.yoelii) (15) or AdH5-MVA (P.berghei) (28). It has been previously 7
shown that a single dose of AdH5 encoding P.yoelii CSP resulted in 93% reduction 8
in liver stage development of the parasite (45), however the dose used in our study 9
is 10 times lower than the dose of adenovirus used in this particular study. Similar 10
efficacy with single dose of adenovirus has not been demonstrated in P.berghei 11
model of malaria. In humans, the liver stage of P.falciparum lasts for 6-7 days as 12
compared to P.berghei (2 -3 days). While effector and TEM cells that respond rapidly 13
to infection may provide sterile protection following P.berghei infection, the longer 14
hepatic stage of P.falciparum may allow time for TCM cells that can proliferate and 15
divide into effector cells to play a role in affording protection. The protective efficacy 16
of adenoviral vectors against the liver-stage of rodent malaria may be ascribed to 17
their ability to induce high frequency of effector and TEM cells which provide 18
immediate protection. The drop in protection over time with adenoviral vectors 19
parallels a drop in the levels of effector and TEM cells. This supports the notion that 20
immediate protection (conferred by effector cells) requires persisting antigen (64). 21
On the other hand, in P.falciparum infection, the ability of adenoviral vectors to delay 22
development of potentially protective TCM cells might make such vaccines provide 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

28
long-lasting protection especially if adenoviral vaccines are used in prime-boost 1
regimes that induce a protective threshold level of TCM cells. 2
Our results demonstrating AdC6 to be more immunogenic and efficacious than 3
AdH5 is consistent with an earlier report evaluating AdC6 as a HIV vaccine in a 4
vaccinia virus challenge model (43). It is as yet unclear to as why simian 5
adenoviruses are more immunogenic than human adenovirus serotype 5. The cell 6
entry receptor for AdC6 is yet to be identified, but its hexon and fiber sequences 7
share 90% homology with AdC68(46) which uses CAR to enter cells(21). It is likely 8
that AdC6 uses CAR to gain entry into cells although other non-CAR mediated entry 9
into cells cannot be ruled out. This mechanism of cellular entry could impact the 10
manner of activation of the innate immune system and downstream activation of the 11
adaptive cellular response. Type I IFN can induce IFN-γ secretion from CD8 T 12
cells(39) and the activation of naïve CD8 T cells is dependent on co-stimulatory 13
signals expressed from mature dendritic cells (DCs). It has been reported that 14
simian adenoviruses induce more type I IFN and also affect a more rapid and 15
effective maturation of DCs(30). We speculate that one of the reasons for the 16
difference in immunogenicity between AdC6 and AdH5 might be related to their 17
differential induction of type I interferon and activation of DCs. 18
A significant percentage of humans carry neutralizing antibodies to the AdH5 and 19
pre-existing immunity to AdH5 has been shown to reduce the efficacy of AdH5 viral 20
vectors (63). To avoid interference by pre-existing antibodies to H5, alternative 21
human serotypes (41) and simian adenoviral vectors have been developed and 22
testing efficacy in the presence of pre-existing immunity is important to establish the 23
potential of the vaccine for use in humans. We induced the development of strong 24
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

29
pre-existing immunity by immunizing mice with AdH5.85A 6 weeks before 1
immunization with a malaria vaccine. We show that pre-existing immunity to AdH5 2
decreased immune responses and completely abrogated protection induced by 3
AdH5.PbCSP vaccination. In contrast, there was no significant drop in 4
immunogenicity of the AdC6.PbCSP vaccine which afforded 66% protection in the 5
setting of pre-existing immunity to AdH5. These results are consistent with similar 6
reports that have demonstrated the abrogation of protective efficacy of AdH5 7
vaccines in the presence of pre-existing immunity to AdH5 (44, 63). Our result 8
showing a drop in protection afforded by AdC6 vaccine in the presence of pre-9
existing immunity to AdH5 is similar to findings in studies using AdC7 (63) and 10
AdC68 (44) vaccine in SARS and HIV model respectively and it is suggested that 11
this phenomenon might be explained by cross-reactive CD8+ T cells that kill 12
transduced APC’s. A recent study demonstrated between 8-18% prevalence of 13
neutralizing antibodies in parts of sub-Saharan Africa to AdC6 (61). A malaria 14
vaccine is most required in this region and despite this low prevalence of neutralizing 15
antibodies to AdC6 we were interested in assessing the effect of our adenoviral 16
vaccines in the presence of pre-existing immunity to AdC6. We induced the 17
development of pre-existing immunity to C6 by immunizing with C6.85A 6 weeks 18
before immunization with a malaria vaccine. In the presence of pre-existing immunity 19
to C6, immunogenicity of AdC6.PbCSP was significantly decreased and 20
consequently protection afforded by AdC6.PbCSP dropped to 33%. Immunogenicity 21
of AdH5.PbCSP did not decrease significantly in the presence of pre-existing 22
immunity to AdC6 and 33% of mice were protected. We observed that similar levels 23
of protection were offered by AdC6 and AdH5 despite differences in the levels of 24
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

30
peptide specific IFN-γ in the blood measured by ELISPOT. This lack of correlation 1
between IFN-γ SFCs and protection has been previously observed in humans (24, 2
25) and in mice (51) and may be indicative of other correlates of protection. While 3
this is the first study to report the effect of pre-existing immunity to simian adenovirus 4
on efficacy of a simian adenoviral vaccine, further work is needed to confirm whether 5
this reduction of efficacy is also caused by neutralizing antibodies. 6
In conclusion, we describe an optimized adenoviral vaccine vector system that 7
amalgamates a novel improved HCMV promoter in E1E3-deleted adenoviral vector 8
to increase antigen transcription and subsequently enhance the immune response. 9
We transfer this optimized adenoviral vector design to the simian adenoviral vector 10
AdC6 to elicit potent CD8+ T cell responses against a pre-erythrocytic antigen of 11
rodent malaria. In contrast to rare human adenovirus serotypes, such as AdHu35 12
(34), the immunogenicity and efficacy of AdC6 simian vector is greater than AdH5 13
and two pox viral vectors, FP9 and MVA. We also demonstrate that the 14
adenoviruses were able to induce and maintain a high frequency of effector CD8+ T 15
cells for a longer period of time than pox viral vectors. Unlike the pox viral vectors 16
which induced TCM cells early after immunization, the adenoviral vectors delay 17
development of TCM cells and maintain a TEM type of memory response for a 18
prolonged duration after immunization. In addition, AdC6.PbCSP induced significant 19
levels of sterile and long-lasting protection in the absence and presence of 20
preexisting immunity to AdH5. Our data demonstrates for the first time that a single 21
dose of a subunit vaccine is able to elicit protection to P. berghei and highlights the 22
potential of AdC6 as a viral vector for further clinical consideration. 23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

31
FIGURES AND TABLE LEGENDS 1
2
Table 1. Sequence of MSP-142 epitopes 3
The amino acid (αα) sequences of the predicted and published epitopes from 4
P.yoelii strain YM MSP-1-42 are listed. Epitopes were predicted using the epitope 5
prediction section of the SYFPEITHI website 6
(http://www.syfpeithi.de/Scripts/MHCServer.dll/EpitopePrediction.htm) developed at 7
the University of Tübingen. 8
9
Figure 1. Comparison of the different promoters. 10
AdH5 adenoviral vector was constructed expressing antigens MSP-142 and 85A 11
under the control of HCMV promoters either with or without an Exon A and Intron A 12
sequence. Transcriptional activity and immunogenicity of each construct was 13
investigated and compared (A) Schematic diagram of the promoter elements 14
used in the adenoviral vectors. SP is a HCMV promoter in pAd/CMV/V5/DEST 15
containing the enhancer/promoter sequence and a short regulatory sequence while 16
LP is the HCMV promoter containing enhancer/promoter sequence along with the 17
exon A and Intron A sequence. (B – C)Comparison of mRNA expression levels 18
between promoters. 293 cells were infected at an MOI of 50 with AdH5.85A.SP, 19
AdH5.85A.LP, MSP-142 .SP or AdH5.MSP-142 .LP and RNA extracted from the 20
cells 48 hours after infection. Reverse-Transcriptase Real-time PCR was undertaken 21
to compare mRNA levels of antigen transcribed by the LP or SP. Antigen mRNA 22
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

32
levels were normalized to AdE4orf1 levels and expressed as a ratio of antigen levels 1
to AdE4orf1 levels. Panel (B) and (C) show the data for MSP-142 and 85A antigens 2
respectively. Statistical analysis was performed using unpaired t-test (GraphPad 3
Prism). **, p < 0.01. The results shown are representative of two independent 4
experiments. (D-F) Effect of promoter on immunogenicity of adenoviral vector. 5
Female Balb/c mice (3/group) were immunized with 1010 vp of AdH5.85A.SP or 6
AdH5.85A.LP (E), 1010 vp of AdH5.MSP-142 .SP or AdH5.MSP-142 .LP (D) and 109 7
vp of AdH5.85A.SP or AdH5.85A.LP (F) intra-dermally (i.d) in the ear. 14 days later, 8
splenocytes were assayed for IFN-γ secretion by ex-vivo ELISPOT. Columns 9
represent mean number of IFN-γ SFC per million splenocytes + SEM. Statistical 10
analysis was performed using a nonparametric two-way ANOVA (GraphPad Prism). 11
*, p<0.05, **, p<0.01 compared to mice immunized with vectors containing SP. 12
13
14
Figure 2. Comparison of E1-deleted and E1E3- deleted adenoviral vectors. 15
Balb/c mice (3/group) were immunized with 1010 vp of E1-deleted AdH5.85A.LP or 16
E1E3-deleted AdH5.85A.LP i.d. in the ear. 14 days later, splenocytes were assayed 17
for IFN-γ secretion by ex-vivo ELISPOT. Columns represent mean number of IFN-γ 18
SFC per million splenocytes + SEM. 19
20
21
22
23
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

33
1
Figure 3. Immunogenicity and effector T cell development following immunization 2
with viral vectors expressing Pb.CSP. 3
(A) Immunogenicity and immune kinetics. Female Balb/c mice (3 mice /group) were 4
immunized with AdH5.PbCSP,AdC6.PbCSP (1010 vp) or MVA.PbCSP and FP9.PbCSP 5
(107 pfu) i.d. Spleens were harvested at different time points post-immunization and 6
antigen specific responses were assayed by intracellular cytokine staining for IFN-γ 7
(CD8+T cells) following stimulation of splenocytes with Pb9. Data are mean + SEM for 8
each group. (B) Phenotyping of Effector cell population. Data shows the percentage of 9
CD8+IFN-γ+ CD43high cells. Columns represent mean + SEM for each group. (C) 10
Cytolytic function of antigen specific cells. CD8+IFN-γ+ Granzyme B co-expression 11
from representative mice at indicated days post-prime. Histogram shows GrB expression 12
(white background) after staining with anti-human GrB, compared to an isotype control 13
(gray background). The number corresponds to MFI of the positive sample (black solid 14
line). 15
16
Figure 4. Phenotyping of memory CD8+ T cell following viral vector 17
immunization. 18
Mice were immunized as described in Figure 2. (A) Long term evolution of CD62L 19
and CD127 on antigen specific cells following immunization. Splenocytes were 20
co-stained for CD8, IFN-γ, CD62L and CD127. Representative stainings are shown 21
for three mice per group. (B) Development of IL2 producing cells. Data shows the 22
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

34
percentage of CD8+IFN-γ+ IL2+ cells. Columns represent mean + SEM for each 1
group. (C) CD27 expression. Data shows the percentage of CD8+IFN-γ+ CD27+ 2
cells. Columns represent mean + SEM for each group. 3
Table 2. Efficacy of viral vector vaccines to a sporozoite challenge 4
Balb/c mice were immunized with adenoviral (1010 vp) or pox viral (107 pfu) vectors 5
and challenged with 1000 P.berghei sporozoites i.v. at different time points. The pre-6
patent period is the time until the appearance of blood stage parasite in Giemsa-7
stained blood smears. Statistical analysis was performed using Kaplan-Meyer 8
survival analysis to calculate the p value (GraphPad Prism). 9
10
11
Figure 5. Immunogenicity and efficacy of adenoviral vectors in the presence of 12
pre-existing immunity 13
Pre-existing immunity was induced by immunizing groups of 6 Balb/c mice with 14
5X109 vp of AdH5.85A or C6.85A. 6 weeks later mice were immunized with 15
adenoviral vectors expressing PbCSP at a dose of 1010 vp per mouse. (A) Effect of 16
pre-existing immunity on immunogenicity of viral vectors. 12 days after 17
immunization blood was collected to analyse IFN-γ secretion by ex-vivo ELISPOT. 18
Data shown represents each individual mouse response expressed as IFN-γ SFC 19
per million PBMCs and the line represents mean response per group. Statistical 20
analysis was performed using nonparametric two-way ANOVA with adjustment for 21
multiple comparisons (GraphPad Prism). Statistical differences are indicated as: **, 22
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

35
p<0.05, ***, p<0.001. (B) Protective efficacy in presence of pre-existing 1
immunity. 14 days post immunization with adenoviral vectors coding for Pb.CSP, 2
mice were challenged with 1000 P.berghei sporozoites i.v. The bars represent the 3
percentage of mice that were protected in each group. All the naïve mice were 4
infected by day 6. 5
6
7
REFERENCES 8
9
10
1. Alonso, P. L., J. Sacarlal, J. J. Aponte, A. Leach, E. Macete, P. Aide, B. Sigauque, J. 11
Milman, I. Mandomando, Q. Bassat, C. Guinovart, M. Espasa, S. Corachan, M. 12
Lievens, M. M. Navia, M. C. Dubois, C. Menendez, F. Dubovsky, J. Cohen, R. 13 Thompson, and W. R. Ballou. 2005. Duration of protection with RTS,S/AS02A malaria 14
vaccine in prevention of Plasmodium falciparum disease in Mozambican children: single-15
blind extended follow-up of a randomised controlled trial. Lancet 366:2012-8. 16
2. Alonso, P. L., J. Sacarlal, J. J. Aponte, A. Leach, E. Macete, J. Milman, I. 17
Mandomando, B. Spiessens, C. Guinovart, M. Espasa, Q. Bassat, P. Aide, O. Ofori-18
Anyinam, M. M. Navia, S. Corachan, M. Ceuppens, M. C. Dubois, M. A. Demoitie, 19
F. Dubovsky, C. Menendez, N. Tornieporth, W. R. Ballou, R. Thompson, and J. 20 Cohen. 2004. Efficacy of the RTS,S/AS02A vaccine against Plasmodium falciparum 21
infection and disease in young African children: randomised controlled trial. Lancet 22
364:1411-20. 23
3. Anderson, R. J., C. M. Hannan, S. C. Gilbert, S. M. Laidlaw, E. G. Sheu, S. Korten, 24
R. Sinden, G. A. Butcher, M. A. Skinner, and A. V. Hill. 2004. Enhanced CD8+ T cell 25
immune responses and protection elicited against Plasmodium berghei malaria by prime 26
boost immunization regimens using a novel attenuated fowlpox virus. J Immunol 27
172:3094-100. 28
4. Baars, P. A., S. Sierro, R. Arens, K. Tesselaar, B. Hooibrink, P. Klenerman, and R. 29
A. van Lier. 2005. Properties of murine (CD8+)CD27- T cells. Eur J Immunol 35:3131-30
41. 31
5. Bachmann, M. F., P. Wolint, K. Schwarz, P. Jager, and A. Oxenius. 2005. Functional 32
properties and lineage relationship of CD8+ T cell subsets identified by expression of IL-33
7 receptor alpha and CD62L. J Immunol 175:4686-96. 34
6. Badovinac, V. P., and J. T. Harty. 2006. Programming, demarcating, and manipulating 35
CD8+ T-cell memory. Immunol Rev 211:67-80. 36
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

36
7. Badovinac, V. P., K. A. Messingham, A. Jabbari, J. S. Haring, and J. T. Harty. 2005. 1
Accelerated CD8+ T-cell memory and prime-boost response after dendritic-cell 2
vaccination. Nat Med 11:748-56. 3
8. Barouch, D. H., M. G. Pau, J. H. Custers, W. Koudstaal, S. Kostense, M. J. 4
Havenga, D. M. Truitt, S. M. Sumida, M. G. Kishko, J. C. Arthur, B. Korioth-5
Schmitz, M. H. Newberg, D. A. Gorgone, M. A. Lifton, D. L. Panicali, G. J. Nabel, 6 N. L. Letvin, and J. Goudsmit. 2004. Immunogenicity of recombinant adenovirus 7
serotype 35 vaccine in the presence of pre-existing anti-Ad5 immunity. J Immunol 8
172:6290-7. 9
9. Bejon, P., J. Mwacharo, O. Kai, T. Mwangi, P. Milligan, S. Todryk, S. Keating, T. 10
Lang, B. Lowe, C. Gikonyo, C. Molyneux, G. Fegan, S. C. Gilbert, N. Peshu, K. 11 Marsh, and A. V. Hill. 2006. A Phase 2b Randomised Trial of the Candidate Malaria 12
Vaccines FP9 ME-TRAP and MVA ME-TRAP among Children in Kenya. PLoS Clin 13
Trials 1:e29. 14
10. Bejon, P., J. Mwacharo, O. K. Kai, S. Todryk, S. Keating, T. Lang, S. C. Gilbert, N. 15
Peshu, K. Marsh, and A. V. Hill. 2006. Immunogenicity of the candidate malaria 16
vaccines FP9 and modified vaccinia virus Ankara encoding the pre-erythrocytic antigen 17
ME-TRAP in 1-6 year old children in a malaria endemic area. Vaccine 24:4709-15. 18
11. Berenzon, D., R. J. Schwenk, L. Letellier, M. Guebre-Xabier, J. Williams, and U. 19
Krzych. 2003. Protracted protection to Plasmodium berghei malaria is linked to 20
functionally and phenotypically heterogeneous liver memory CD8+ T cells. J Immunol 21
171:2024-34. 22
12. Bett, A. J., L. Prevec, and F. L. Graham. 1993. Packaging capacity and stability of 23
human adenovirus type 5 vectors. J Virol 67:5911-21. 24
13. Bojang, K. A., P. J. Milligan, M. Pinder, L. Vigneron, A. Alloueche, K. E. Kester, W. 25
R. Ballou, D. J. Conway, W. H. Reece, P. Gothard, L. Yamuah, M. Delchambre, G. 26
Voss, B. M. Greenwood, A. Hill, K. P. McAdam, N. Tornieporth, J. D. Cohen, and 27 T. Doherty. 2001. Efficacy of RTS,S/AS02 malaria vaccine against Plasmodium 28
falciparum infection in semi-immune adult men in The Gambia: a randomised trial. 29
Lancet 358:1927-34. 30
14. Boshart, M., F. Weber, G. Jahn, K. Dorsch-Hasler, B. Fleckenstein, and W. 31
Schaffner. 1985. A very strong enhancer is located upstream of an immediate early gene 32
of human cytomegalovirus. Cell 41:521-30. 33
15. Bruna-Romero, O., G. Gonzalez-Aseguinolaza, J. C. Hafalla, M. Tsuji, and R. S. 34
Nussenzweig. 2001. Complete, long-lasting protection against malaria of mice primed 35
and boosted with two distinct viral vectors expressing the same plasmodial antigen. Proc 36
Natl Acad Sci U S A 98:11491-6. 37
16. Buchman, A. R., and P. Berg. 1988. Comparison of intron-dependent and intron-38
independent gene expression. Mol Cell Biol 8:4395-405. 39
17. Casimiro, D. R., A. Tang, L. Chen, T. M. Fu, R. K. Evans, M. E. Davies, D. C. 40
Freed, W. Hurni, J. M. Aste-Amezaga, L. Guan, R. Long, L. Huang, V. Harris, D. 41
K. Nawrocki, H. Mach, R. D. Troutman, L. A. Isopi, K. K. Murthy, K. Rice, K. A. 42 Wilson, D. B. Volkin, E. A. Emini, and J. W. Shiver. 2003. Vaccine-induced immunity 43
in baboons by using DNA and replication-incompetent adenovirus type 5 vectors 44
expressing a human immunodeficiency virus type 1 gag gene. J Virol 77:7663-8. 45
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

37
18. Chapman, B. S., R. M. Thayer, K. A. Vincent, and N. L. Haigwood. 1991. Effect of 1
intron A from human cytomegalovirus (Towne) immediate-early gene on heterologous 2
expression in mammalian cells. Nucleic Acids Res 19:3979-86. 3
19. Chiuchiolo, M. J., J. L. Boyer, A. Krause, S. Senina, N. R. Hackett, and R. G. 4
Crystal. 2006. Protective immunity against respiratory tract challenge with Yersinia 5
pestis in mice immunized with an adenovirus-based vaccine vector expressing V antigen. 6
J Infect Dis 194:1249-57. 7
20. Clyde, D. F., H. Most, V. C. McCarthy, and J. P. Vanderberg. 1973. Immunization of 8
man against sporozite-induced falciparum malaria. Am J Med Sci 266:169-77. 9
21. Cohen, C. J., Z. Q. Xiang, G. P. Gao, H. C. Ertl, J. M. Wilson, and J. M. Bergelson. 10
2002. Chimpanzee adenovirus CV-68 adapted as a gene delivery vector interacts with the 11
coxsackievirus and adenovirus receptor. J Gen Virol 83:151-5. 12
22. Denis, O., A. Tanghe, K. Palfliet, F. Jurion, T. P. van den Berg, A. Vanonckelen, J. 13
Ooms, E. Saman, J. B. Ulmer, J. Content, and K. Huygen. 1998. Vaccination with 14
plasmid DNA encoding mycobacterial antigen 85A stimulates a CD4+ and CD8+ T-cell 15
epitopic repertoire broader than that stimulated by Mycobacterium tuberculosis H37Rv 16
infection. Infect Immun 66:1527-33. 17
23. Fessler, S. P., F. Delgado-Lopez, and M. S. Horwitz. 2004. Mechanisms of E3 18
modulation of immune and inflammatory responses. Curr Top Microbiol Immunol 19
273:113-35. 20
24. Flanagan, K. L., E. A. Lee, M. B. Gravenor, W. H. Reece, B. C. Urban, T. Doherty, 21
K. A. Bojang, M. Pinder, A. V. Hill, and M. Plebanski. 2001. Unique T cell effector 22
functions elicited by Plasmodium falciparum epitopes in malaria-exposed Africans tested 23
by three T cell assays. J Immunol 167:4729-37. 24
25. Flanagan, K. L., T. Mwangi, M. Plebanski, K. Odhiambo, A. Ross, E. Sheu, M. 25
Kortok, B. Lowe, K. Marsh, and A. V. Hill. 2003. Ex vivo interferon-gamma immune 26
response to thrombospondin-related adhesive protein in coastal Kenyans: longevity and 27
risk of Plasmodium falciparum infection. Am J Trop Med Hyg 68:421-30. 28
26. Foecking, M. K., and H. Hofstetter. 1986. Powerful and versatile enhancer-promoter 29
unit for mammalian expression vectors. Gene 45:101-5. 30
27. Gilbert, S. C., M. Plebanski, S. J. Harris, C. E. Allsopp, R. Thomas, G. T. Layton, 31
and A. V. Hill. 1997. A protein particle vaccine containing multiple malaria epitopes. 32
Nat Biotechnol 15:1280-4. 33
28. Gilbert, S. C., J. Schneider, C. M. Hannan, J. T. Hu, M. Plebanski, R. Sinden, and 34
A. V. Hill. 2002. Enhanced CD8 T cell immunogenicity and protective efficacy in a 35
mouse malaria model using a recombinant adenoviral vaccine in heterologous prime-36
boost immunisation regimes. Vaccine 20:1039-45. 37
29. Good, M. F., and D. L. Doolan. 1999. Immune effector mechanisms in malaria. Curr 38
Opin Immunol 11:412-9. 39
30. Hensley, S. E., W. Giles-Davis, K. C. McCoy, W. Weninger, and H. C. Ertl. 2005. 40
Dendritic cell maturation, but not CD8+ T cell induction, is dependent on type I IFN 41
signaling during vaccination with adenovirus vectors. J Immunol 175:6032-41. 42
31. Huang, M. T., and C. M. Gorman. 1990. Intervening sequences increase efficiency of 43
RNA 3' processing and accumulation of cytoplasmic RNA. Nucleic Acids Res 18:937-44
47. 45
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

38
32. Kobinger, G. P., H. Feldmann, Y. Zhi, G. Schumer, G. Gao, F. Feldmann, S. Jones, 1
and J. M. Wilson. 2006. Chimpanzee adenovirus vaccine protects against Zaire Ebola 2
virus. Virology 346:394-401. 3
33. Kumar, K. A., G. Sano, S. Boscardin, R. S. Nussenzweig, M. C. Nussenzweig, F. 4
Zavala, and V. Nussenzweig. 2006. The circumsporozoite protein is an 5
immunodominant protective antigen in irradiated sporozoites. Nature 444:937-40. 6
34. Lemckert, A. A., S. M. Sumida, L. Holterman, R. Vogels, D. M. Truitt, D. M. Lynch, 7
A. Nanda, B. A. Ewald, D. A. Gorgone, M. A. Lifton, J. Goudsmit, M. J. Havenga, 8 and D. H. Barouch. 2005. Immunogenicity of heterologous prime-boost regimens 9
involving recombinant adenovirus serotype 11 (Ad11) and Ad35 vaccine vectors in the 10
presence of anti-ad5 immunity. J Virol 79:9694-701. 11
35. Maizel, J. V., Jr., D. O. White, and M. D. Scharff. 1968. The polypeptides of 12
adenovirus. I. Evidence for multiple protein components in the virion and a comparison 13
of types 2, 7A, and 12. Virology 36:115-25. 14
36. McConkey, S. J., W. H. Reece, V. S. Moorthy, D. Webster, S. Dunachie, G. Butcher, 15
J. M. Vuola, T. J. Blanchard, P. Gothard, K. Watkins, C. M. Hannan, S. Everaere, 16
K. Brown, K. E. Kester, J. Cummings, J. Williams, D. G. Heppner, A. Pathan, K. 17
Flanagan, N. Arulanantham, M. T. Roberts, M. Roy, G. L. Smith, J. Schneider, T. 18 Peto, R. E. Sinden, S. C. Gilbert, and A. V. Hill. 2003. Enhanced T-cell 19
immunogenicity of plasmid DNA vaccines boosted by recombinant modified vaccinia 20
virus Ankara in humans. Nat Med 9:729-35. 21
37. Moore, A. C., A. Gallimore, S. J. Draper, K. R. Watkins, S. C. Gilbert, and A. V. 22
Hill. 2005. Anti-CD25 antibody enhancement of vaccine-induced immunogenicity: 23
increased durable cellular immunity with reduced immunodominance. J Immunol 24
175:7264-73. 25
38. Moorthy, V. S., E. B. Imoukhuede, S. Keating, M. Pinder, D. Webster, M. A. 26
Skinner, S. C. Gilbert, G. Walraven, and A. V. Hill. 2004. Phase 1 evaluation of 3 27
highly immunogenic prime-boost regimens, including a 12-month reboosting vaccination, 28
for malaria vaccination in Gambian men. J Infect Dis 189:2213-9. 29
39. Nguyen, K. B., W. T. Watford, R. Salomon, S. R. Hofmann, G. C. Pien, A. 30
Morinobu, M. Gadina, J. J. O'Shea, and C. A. Biron. 2002. Critical role for STAT4 31
activation by type 1 interferons in the interferon-gamma response to viral infection. 32
Science 297:2063-6. 33
40. Nussenzweig, R. S., J. Vanderberg, H. Most, and C. Orton. 1967. Protective immunity 34
produced by the injection of x-irradiated sporozoites of plasmodium berghei. Nature 35
216:160-2. 36
41. Ophorst, O. J., K. Radosevic, M. J. Havenga, M. G. Pau, L. Holterman, B. 37
Berkhout, J. Goudsmit, and M. Tsuji. 2006. Immunogenicity and protection of a 38
recombinant human adenovirus serotype 35-based malaria vaccine against Plasmodium 39
yoelii in mice. Infect Immun 74:313-20. 40
42. Otten, G. R., M. Schaefer, B. Doe, H. Liu, I. Srivastava, J. Megede, J. Kazzaz, Y. 41
Lian, M. Singh, M. Ugozzoli, D. Montefiori, M. Lewis, D. A. Driver, T. Dubensky, J. 42 M. Polo, J. Donnelly, D. T. O'Hagan, S. Barnett, and J. B. Ulmer. 2005. Enhanced 43
potency of plasmid DNA microparticle human immunodeficiency virus vaccines in 44
rhesus macaques by using a priming-boosting regimen with recombinant proteins. J Virol 45
79:8189-200. 46
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

39
43. Pinto, A. R., J. C. Fitzgerald, G. P. Gao, J. M. Wilson, and H. C. Ertl. 2004. 1
Induction of CD8+ T cells to an HIV-1 antigen upon oral immunization of mice with a 2
simian E1-deleted adenoviral vector. Vaccine 22:697-703. 3
44. Reyes-Sandoval, A., J. C. Fitzgerald, R. Grant, S. Roy, Z. Q. Xiang, Y. Li, G. P. 4
Gao, J. M. Wilson, and H. C. Ertl. 2004. Human immunodeficiency virus type 1-5
specific immune responses in primates upon sequential immunization with adenoviral 6
vaccine carriers of human and simian serotypes. J Virol 78:7392-9. 7
45. Rodrigues, E. G., F. Zavala, D. Eichinger, J. M. Wilson, and M. Tsuji. 1997. Single 8
immunizing dose of recombinant adenovirus efficiently induces CD8+ T cell-mediated 9
protective immunity against malaria. J Immunol 158:1268-74. 10
46. Roy, S., G. Gao, D. S. Clawson, L. H. Vandenberghe, S. F. Farina, and J. M. Wilson. 11
2004. Complete nucleotide sequences and genome organization of four chimpanzee 12
adenoviruses. Virology 324:361-72. 13
47. Santra, S., M. S. Seaman, L. Xu, D. H. Barouch, C. I. Lord, M. A. Lifton, D. A. 14
Gorgone, K. R. Beaudry, K. Svehla, B. Welcher, B. K. Chakrabarti, Y. Huang, Z. Y. 15 Yang, J. R. Mascola, G. J. Nabel, and N. L. Letvin. 2005. Replication-defective 16
adenovirus serotype 5 vectors elicit durable cellular and humoral immune responses in 17
nonhuman primates. J Virol 79:6516-22. 18
48. Schneider, J., S. C. Gilbert, T. J. Blanchard, T. Hanke, K. J. Robson, C. M. Hannan, 19
M. Becker, R. Sinden, G. L. Smith, and A. V. Hill. 1998. Enhanced immunogenicity 20
for CD8+ T cell induction and complete protective efficacy of malaria DNA vaccination 21
by boosting with modified vaccinia virus Ankara. Nat Med 4:397-402. 22
49. Schneider, J., J. A. Langermans, S. C. Gilbert, T. J. Blanchard, S. Twigg, S. Naitza, 23
C. M. Hannan, M. Aidoo, A. Crisanti, K. J. Robson, G. L. Smith, A. V. Hill, and A. 24 W. Thomas. 2001. A prime-boost immunisation regimen using DNA followed by 25
recombinant modified vaccinia virus Ankara induces strong cellular immune responses 26
against the Plasmodium falciparum TRAP antigen in chimpanzees. Vaccine 19:4595-27
602. 28
50. Schofield, L., J. Villaquiran, A. Ferreira, H. Schellekens, R. Nussenzweig, and V. 29
Nussenzweig. 1987. Gamma interferon, CD8+ T cells and antibodies required for 30
immunity to malaria sporozoites. Nature 330:664-6. 31
51. Sedegah, M., T. R. Jones, M. Kaur, R. Hedstrom, P. Hobart, J. A. Tine, and S. L. 32
Hoffman. 1998. Boosting with recombinant vaccinia increases immunogenicity and 33
protective efficacy of malaria DNA vaccine. Proc Natl Acad Sci U S A 95:7648-53. 34
52. Sekaly, R. P. 2008. The failed HIV Merck vaccine study: a step back or a launching 35
point for future vaccine development? J Exp Med. 36
53. Skinner, M. A., B. M. Buddle, D. N. Wedlock, D. Keen, G. W. de Lisle, R. E. Tascon, 37
J. C. Ferraz, D. B. Lowrie, P. J. Cockle, H. M. Vordermeier, and R. G. Hewinson. 38 2003. A DNA prime-Mycobacterium bovis BCG boost vaccination strategy for cattle 39
induces protection against bovine tuberculosis. Infect Immun 71:4901-7. 40
54. Snow, R. W., C. A. Guerra, A. M. Noor, H. Y. Myint, and S. I. Hay. 2005. The global 41
distribution of clinical episodes of Plasmodium falciparum malaria. Nature 434:214-7. 42
55. Tatsis, N., and H. C. Ertl. 2004. Adenoviruses as vaccine vectors. Mol Ther 10:616-29. 43
56. Tatsis, N., L. Tesema, E. R. Robinson, W. Giles-Davis, K. McCoy, G. P. Gao, J. M. 44
Wilson, and H. C. Ertl. 2006. Chimpanzee-origin adenovirus vectors as vaccine carriers. 45
Gene Ther 13:421-9. 46
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

40
57. Webster, D. P., S. Dunachie, J. M. Vuola, T. Berthoud, S. Keating, S. M. Laidlaw, S. 1
J. McConkey, I. Poulton, L. Andrews, R. F. Andersen, P. Bejon, G. Butcher, R. 2 Sinden, M. A. Skinner, S. C. Gilbert, and A. V. Hill. 2005. Enhanced T cell-mediated 3
protection against malaria in human challenges by using the recombinant poxviruses FP9 4
and modified vaccinia virus Ankara. Proc Natl Acad Sci U S A 102:4836-41. 5
58. Wherry, E. J., V. Teichgraber, T. C. Becker, D. Masopust, S. M. Kaech, R. Antia, U. 6
H. von Andrian, and R. Ahmed. 2003. Lineage relationship and protective immunity of 7
memory CD8 T cell subsets. Nat Immunol 4:225-34. 8
59. Wipasa, J., C. Hirunpetcharat, Y. Mahakunkijcharoen, H. Xu, S. Elliott, and M. F. 9
Good. 2002. Identification of T cell epitopes on the 33-kDa fragment of Plasmodium 10
yoelii merozoite surface protein 1 and their antibody-independent protective role in 11
immunity to blood stage malaria. J Immunol 169:944-51. 12
60. Xiang, Z., G. Gao, A. Reyes-Sandoval, C. J. Cohen, Y. Li, J. M. Bergelson, J. M. 13
Wilson, and H. C. Ertl. 2002. Novel, chimpanzee serotype 68-based adenoviral vaccine 14
carrier for induction of antibodies to a transgene product. J Virol 76:2667-75. 15
61. Xiang, Z., Y. Li, A. Cun, W. Yang, S. Ellenberg, W. M. Switzer, M. L. Kalish, and 16
H. C. Ertl. 2006. Chimpanzee adenovirus antibodies in humans, sub-Saharan Africa. 17
Emerg Infect Dis 12:1596-9. 18
62. Xiang, Z. Q., Y. Yang, J. M. Wilson, and H. C. Ertl. 1996. A replication-defective 19
human adenovirus recombinant serves as a highly efficacious vaccine carrier. Virology 20
219:220-7. 21
63. Zhi, Y., J. Figueredo, G. P. Kobinger, H. Hagan, R. Calcedo, J. R. Miller, G. Gao, 22
and J. M. Wilson. 2006. Efficacy of severe acute respiratory syndrome vaccine based on 23
a nonhuman primate adenovirus in the presence of immunity against human adenovirus. 24
Hum Gene Ther 17:500-6. 25
64. Zinkernagel, R. M., M. F. Bachmann, T. M. Kundig, S. Oehen, H. Pirchet, and H. 26
Hengartner. 1996. On immunological memory. Annu Rev Immunol 14:333-67. 27
28
29 ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
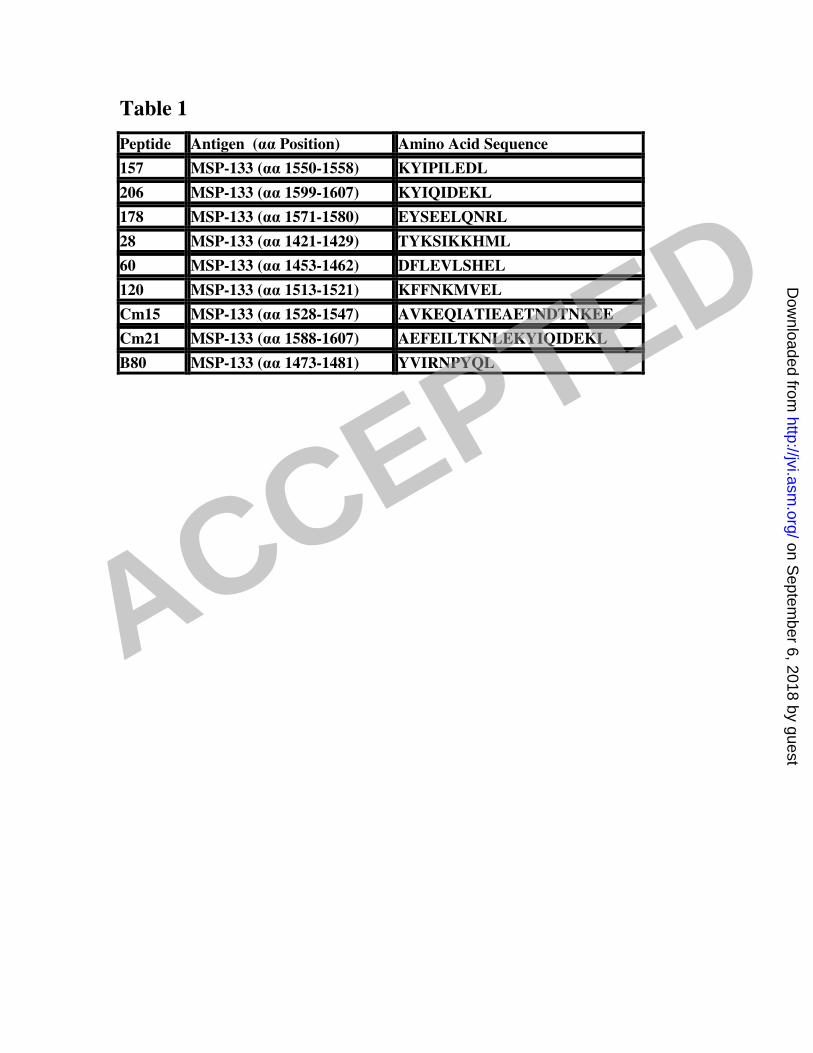
Table 1
Peptide Antigen (αα Position) Amino Acid Sequence
157 MSP-133 (αα 1550-1558) KYIPILEDL
206 MSP-133 (αα 1599-1607) KYIQIDEKL
178 MSP-133 (αα 1571-1580) EYSEELQNRL
28 MSP-133 (αα 1421-1429) TYKSIKKHML
60 MSP-133 (αα 1453-1462) DFLEVLSHEL
120 MSP-133 (αα 1513-1521) KFFNKMVEL
Cm15 MSP-133 (αα 1528-1547) AVKEQIATIEAETNDTNKEE
Cm21 MSP-133 (αα 1588-1607) AEFEILTKNLEKYIQIDEKL
B80 MSP-133 (αα 1473-1481) YVIRNPYQL
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from



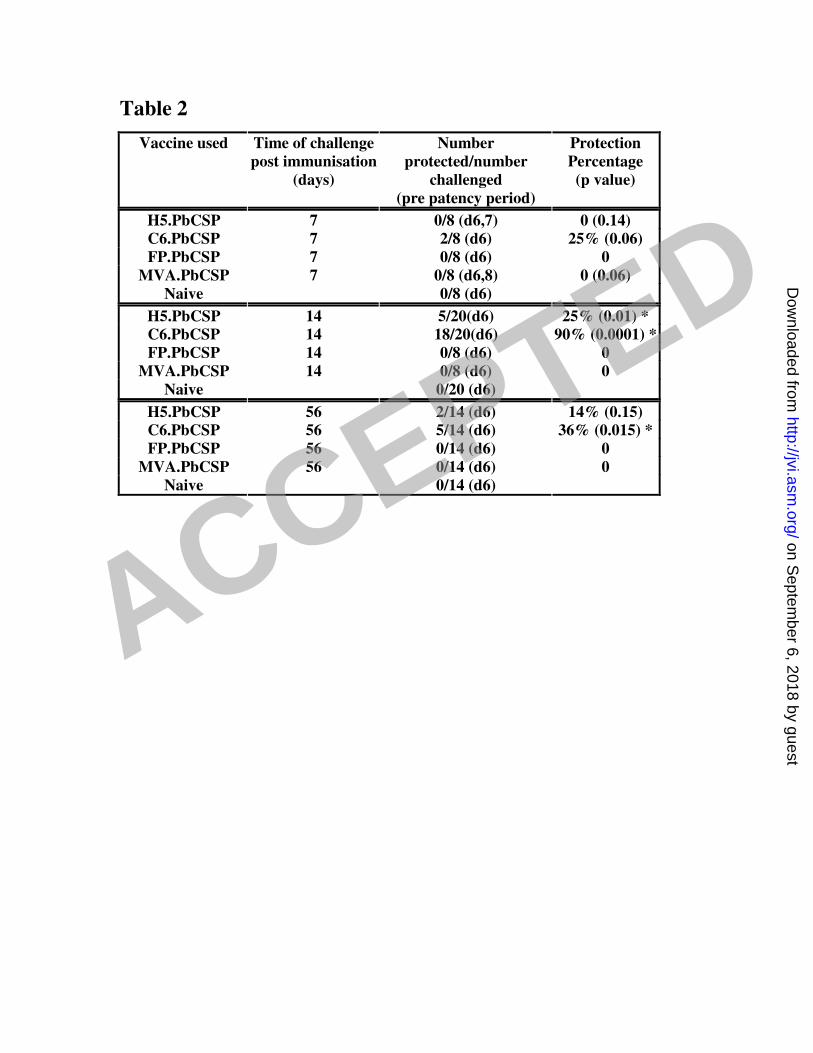
Table 2
Vaccine used Time of challenge
post immunisation
(days)
Number
protected/number
challenged
(pre patency period)
Protection
Percentage
(p value)
H5.PbCSP 7 0/8 (d6,7) 0 (0.14) C6.PbCSP 7 2/8 (d6) 25% (0.06) FP.PbCSP 7 0/8 (d6) 0
MVA.PbCSP 7 0/8 (d6,8) 0 (0.06) Naive 0/8 (d6)
H5.PbCSP 14 5/20(d6) 25% (0.01) * C6.PbCSP 14 18/20(d6) 90% (0.0001) * FP.PbCSP 14 0/8 (d6) 0
MVA.PbCSP 14 0/8 (d6) 0 Naive 0/20 (d6)
H5.PbCSP 56 2/14 (d6) 14% (0.15) C6.PbCSP 56 5/14 (d6) 36% (0.015) * FP.PbCSP 56 0/14 (d6) 0
MVA.PbCSP 56 0/14 (d6) 0 Naive 0/14 (d6)
ACCEPTED
on Septem
ber 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
