adaptation of barley roots low oxygen supply and its ... · adaptation of barley roots to low...
TRANSCRIPT

Plant Physiol. (1969) 44, 1233-1240
Adaptation of Barley Roots to Low Oxygen Supply and its Relationto Potassium and Sodium Uptake
M. G. Pitman'Mineral Nutrition Laboratory, USDA, ARS, SWCRD, Beltsville, Maryland
Received March 10, 1969.
Abstract. The uptake of Na4 and K4 by barley seedlings grown on aerated or non-aeratedsolutions was studied. Plants growing in culture solution took up K+ with high selectivitywhether the solution was aerated or not. Roots of plants grown on aerated CaSO4 and trans-ferred to a solution of KCI and NaCl had a lower preference for K+ than roots of plants grownon non-aerated CaS04. Both kinds of low-salt roots were much less able to discriminatebetween K4 and Na+ than high-salt roots grown on a culture solution. The different levels ofK4 selectivity are suggested to be related to H4 release from *the tissue.
Roots of barley seedlings can become adapted tolow oxygen in their immediate environment by de-velopment of an extensive air path that facilitatesgas exchange with the air via the intercellular spacesof the shoot. Consequently, the seedlings are ableto grow in solutions whether aerated or not (1,23,24).
Physiological differences have been observed be-tween plants adapted to different levels of oxygensupply. Van der Heide et al. (23) found that thesugar level in barley roots growing on one-fourthHoagland solution was higher when nitrogen wasbubbled through the solution than when air was used.In addition, the rate of K4 and Cl- uptake (relativeto dry weight) was about 20 % higher in the 'nitro-gen' plants.
A distinction needs to be made between kinds ofbarley root preparations used to study salt uptake,that is, between low- and high-salt roots. Prepara-tions commonly used to study rates of uptake aregrown from germination on dilute CaSO4 and arelow-salt roots. In this case K4 is restricted to thatoriginally in the seed and so is at a low level in theplant. Roots of seedlings grown on a culture solu-tion are useful for studies of tracer exchange asthere is little or no net salt uptake and the cells arein flux equilibrium with the solution. In generalhigh salt roots show more preference for K4 than dolow salt roots (13, 18). Low-salt roots can reachhigh-salt status as a result of salt uptake and thenalso show a higher preference for K4 vs. Na+, asmeasured by tracer uptake. Measurement of the
On leave from School of Biology, University ofSydney, Sydney, Australia.
level of K+ and Na+ in such roots when they reachhigh salt status need not show much discrimination.These levels will have been determined mainly bythe initial uptake which was of low selectivity. Forthis reason the history of the roots needs to betaken into account when judging the selectivity forK4 vs. Na4.
The purpose of this paper is to examine theeffects that adaptations to aeration, or lack of it,have on K+ and Na+ uptake into roots of barleyseedlings. The majority of workers in this fieldhave used roots grown on aerated solutions; in myown work I have used roots of plants grown onnon-aerated solutions. The comparisons given hereprovide a basis for relating results for these differentkinds of roots.
It is shown that uptake into plants growing in anutrient or salt solution is strongly preferential forK4 whether aerated or not. However, low-salt rootsgrown on aerated CaSO4 solution are much lessselective for KE vs. Na+ than roots grown on non-aerated CaSO4. The differences in K4 selectivityare suggested to be related to H4 release induced bysalt uptake.
Materials
Seeds of Hordeum vulgare cv. Compana, weresoaked in aerated water for about 8 hr then germi-nated on cheesecloth over stainless steel gauze abovean aerated solution of 0.5 mm CaSO4. When theroots were about 1 cm long, and reached into thesolution from the gauze, the pots of seedlings weredivided into 2 sets; one was aerated and in the otheraeration was discontinued. Roots from these 2 setswill be referred to as AER and N-AER, respectively.
1233 www.plantphysiol.orgon March 21, 2019 - Published by Downloaded from
Copyright © 1969 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
The seedlinigs were used when 5 days old. Atthis stage, the AER roots were about 11 cmii long and450 It diameter; N-AER roots were about 6 cm longand 600 tu in diamiieter. Microscopic examinationshowed that this differeence in root diameter wcas dueto greater cell widtlh and nlot to an increase in cellnumber. The cell lengtlh in AER roots was abotit20 % greater tlhain in N-AER roots. As a result ofdifferences in cell size, the surface area of corticalcells per gram of roots was greater in AER roots(1400 cm2/g) than in N-AER roots (1000 cm112/g)and the volume of intercellular spaces was less. TheN-AER roots floated on solution and were intenselywhite due to internal scattering of light; the AERroots were slightly denser than water and translucent.Some properties of AER and N-AER roots arelisted in table I.
The basal (upper) 3 to 4 cim of AER roots and2 to 3 cm of N-AER roots were niot used in experi-ments. It was found that uptake by variouts partsof the rest of the root was about the same.
For analIysis of salt content, roots were ashedat 4500 and dissolved in a mixtutre containinig 0.1 AHNO3 + 10 % v/v acetic acid. Concentrations ofK+, Na+, Ca2 , and Mg2+ were determinied by atomiiicabsorption spectroscopy; Cl- was miieasured by coIn-lombic titration.
Table 1. Comipiarison of Propnrties of Barley RootsGrowo1. ont A4eratcd or A on-Acrated
CaSO4 SollutionAThe difference in root diameter is due to differences
in cell width, not cell niumber. Of the amino acidsl)resent, the maini difference w-as in alanine level. Totalnitrogen levels, and respiration rates wx-ere the same forAER and N-AER roots. N'-AER roots contained muclhless Ca2+ than AER roots. All quantities per g arerelative to 1 g fresh weight of root.
Tracer 86Rb and 22Na uptake was measured bycouintilln, y-radiationi. Samples wTere rinsed for 60sec in ice-water after removal from the labeledsolution, blotted and weighed.
Selectivity of K' is (lefinied as
TK-Naiuptake of K+ / uptake of Na+K+ in solution Na+ in solution
Uptake may be net tuptake or tracer uptake. WhenSK.'Na is greater than 1.0, there is a preference forpotassium, and when less thLan 1.0, a preference forsodium.
Results
Uptake by High-Salt Roots. High SK,Na seemsto be a genieral property of highi.-salt barley rootsgrown onI a culture solutioni. Fig. 1 shows thatuptake by roots of non-aerated plaants in the range10 to 100 mM (K+ +- Na+) has a uniformly highpreference for 1K- with SK.NIt about 8.5 (14). Tablel1 compares the content of seedlings grown onaerated and non-aerated cuilture solutionis containing10 iMi (K- -4 Na ). For both kinds of root, 5KNawas relatively- hiiglh but tihere was a slightly higherl)reference for K in aerated roots, possibly due tothe more efficienit stirring. Uptake to the shoot wasthe same in eaclh case, anid it appears that aeration,Mr lack of it. had little effect on salt uptake.
Uptake I;ito L(o}wt Salt Roots. Wheni roots ofl)lants grown ill (lilute CaSO, (low-salt roots) arel)ut into salt solution, SK'N,, calculated from rates ofuptake decreases with increasing concentration (Fig.1'). Thlus selectivity of uptake by low-salt roots
Plants grown on
Length (cm)Dianmeter (L)No. of cortical cells/sectionSurface area of cortical cells
(cm2/g)Dry weight (nmg/g)K+ (/Leq/g)Na "Ca2+Mg2+Total N (umoles/g,)uISoluble N 2
Alanine 2
Total sugarGlucoseRespiration (,umoles 0,/g.hr)Salt respiration (10 m.a)
(,umoles O,/ghr)Potential difference (mV),"
CaSO4AER N-AER
11.0 6.0450 600250 ± 20 220 ± 20
1400 1000
50 - 70 50- 7018 182.0 2.07.0 3.15.2 3.4
180 19031 340.5 6.0
90 13030 4814.0 13.53.5 3.5
-110 -80
10 1
8SK6N
4
2
20 40 60 80 100Cl mM
FIG. 1. Comparison of SN, No in high- and low-saltroots. Uptake to high-salt roots (0) calculated frompreviously published data (14) was at a uniformly highselectivity. Uptake to low-salt roots, ( 0, open star)becamiie much less selective at higher conicentrations.(Open star), calculated froml data of Rains and Epstein19) and (20). (j>), fromi table TIT.
- 0
* 0. pHigh-salt roots 0
Low-salt rootsI--
4 4---4_--4 --~
Assayed by J. E. Leggett2 Amino acid separation by Miss M. E Enigelhaupt.3 In collaboration with Prof. N. Higinbotham.
1234
www.plantphysiol.orgon March 21, 2019 - Published by Downloaded from Copyright © 1969 American Society of Plant Biologists. All rights reserved.

PITMAN-AERATION AND K, Na4 UPTAKE BY BARLEY ROOTS
Table II. Content of Seedlings Grown for 6 Days AfterGermiiination on a Full Nutrient Containiing
2.5 1m1 K+ antd 7.5 m7tm Na+Composition of nutrient solution: Ca24 = 6, Mg24 =
4, K+ = 2.5, Na+ = 7.5, NH+4 = 0.4, N0-3 = 16,S042- =4, H2PO4; 0.4 (all mN); Fe3+ as Fe-EDTA,trace elements.
Fresh wt K+ Na+ Ca24 Mg2+ SK,Na
mig/plant pueq/g fresh wtRootsAerated 125 90 28 2.2 8.2 9.6Non-aerated 121 74 36 2.4 7.7 6.2
ShootsAerated 249 134 15 9.5 15 7 27Non-Aerated 228 139 15.5 6.1 12.4 27
from a 10 mm (K4 -4- Na4) solution can be verymuch less than that of roots grown from germinationin a 10 mM (K4 + Na+) solution, and hence athigh-salt status (16.,18).
In general SI(,Na was higher for N-AER thanfor AER roots, as seen in table III and Fig. 2.
Table III. Comiiparison of Selectivity of Uptake of K+antd Na+ b,y AER and N-AER Roots From
Solutions of Different ConcentrationsUptake was measured over the first 4 hr in salt
solution. N+ and Na+ concentrations were equal andpH was maintained at 5.3.
Solution Uptake of (K+ + Na+) SK,NaAER N-AER AER N-AER
Cl- ,ueq/g10 mM 64 45 0.4 1.61 mM 31 23.5 0.3 0.750.2 mm 22 24 1.75 7.0so42-10 mN 35.5 29 0.07 0.70
0 20 10 20Hous Hovrs
FIG. 2. Time course of net K+ and 'a+ uptake andH+ loss from roots in [2.5 mnt KCI + 7.5 mM NaCl+ 0.5 mM CaSO ] at 230. ST. Na in AER roots dur-ingf the first 4 hr (0.3) was much less than that inN-AER roots (18). H- release was less rapid fromN-AER roots than AER roots and stopped after about8 hr.
Table III shows that this difference between N-AERand AER roots was found at concentrations whereMeclhanism I shotild be dominant (0.2 mM Cl-) andalso when MIechanism II was well developed (10 mMCl-). Other experiments have shown that this effectwould be fouind up to at least 50 mm Cl-. Table IIIalso shows that the difference occurs in 10 mNSO42- solutions though SIKNa was lower than in a
similar Cl- solution. The pH of the solution was
maintained at 5.3 0.2 by titration with a mixtureof KOH and NaOH of the same K4 and Na4 con-
centrations as the uptake solutions.Uptakes determined in a number of experiments
are combined in table IV. The solution containedthe usual level of 2.5 mM KCI + 7.5 mM NaCl +0.5 mM CaSO4. As in table II, these results showthat more Cl- and more cation (K+ and Na+) weretaken up by AER than by N-AER roots. However,the rates of uptake into AER and N-AER rootsexpressed on the basis of cell surface are the same.
It appears that rates of transport across the cellsurface in each kind of root are the same, and that
Table IV. Uptake Into AER and N-AER Roots on Basis of Root Fresh WVeight and Surface Area of CellsThe greater uptake into AER roots appears to be due to the larger surface area of cells in these roots.
K' Na+ Cl- (K4 ± Na4) SK, NaAER roots (7) ,ueqig
4 hr 6 ±2 49 ±2 40 ±1.5 55 +3 0.377 hr 11 ±1 67 +3 62 ±2 78 ±3 0.50
N-AER roots (5)4 hr 18.5 ±1 20.5 ±3 26.5 ±0.5 39 ±3 2.77 hr 29 ±4 27 ±4 44 ±3 56 +5 3.2
AeqllOOO Cm12 cell suirfaceAER roots
4 hr 4.3 35 28.5 397 hr 8 48 44 56
N-AER roots4 hr 18.5 20.5 26.5 397 hr 29 27 44 56
1235
N-AER
50
Na-
o0 --0 .
/ K
H'Ioss
www.plantphysiol.orgon March 21, 2019 - Published by Downloaded from Copyright © 1969 American Society of Plant Biologists. All rights reserved.
PLANT PfIYSIOLOGYlarger uptake to AER roots is simply due to thesmaller cells in these roots. It is suggested thereforethat the difference in SKINa between the 2 kinds ofroots is not due to an effect on rate of total salttransport across the cell membranes, but to a separateaction on K+ and Na+ transport. As high-salt rootsshow high SK,Na whether grown on aerated solutionsor not, the difference between N-AER and AERroots can be thought of conveniently as a greaterreduction of SK,Na in AER roots than in N-AERroots.
H+ Release and Selectivity Changes. Fig. 2showed there was a rapid loss of H+ when low-saltroots were put into 10 mm salt solution. This H"loss continued at a nearly constant rate for about4 hr but after 6 hr in solution the rate of lossdecreased. This change in rate of H+ release takesplace at about the same time as an increase in SK,Na.For example in 1 experiment with AER roots up-takes of K+ and Na+ were 6.9 and 23 tteq/g between? and 4 hr, and 5.9 and 6.5 ueq/g between 4 and 8.1r, giving SK,Na of 0.9 and 2.7 respectively. Similarchanges in SK,Na were reported previously for non-aerated roots (16, 18).
Tracer 86Rb Uptake. The initial rates of 86Rbuptake by AER and N-AER roots were very nearlythe same and about 8.0 ,xeq per g per hr (Fig. 3).About 20 min after the start of uptake there was areduction in rate of uptake, which was more markedfor AER than for N-AER roots. Between 20 minand 1 hr the rates of uptake were 3.4 jeq per g per hrinto AER roots and 6.7 ,ueq per g per hr into N-AERroots. Consequently over longer periods there wasgreater 86Rb uptake into N-AER roots.
The shoulder at 20 to 30 min is thought torepresent a change in K+ fluxes and not to be the
12 -Rb uptake/ ~~0
10 N-AER,'
8 /oueq/gg
6 AER
4 :20
0AER 2:
1 2 3 4Hours
FIG. 3. Uptake of 86Rb by AER (0)) and N-AER(*) roots from 2.5 mm KC1 + 7.5 mini NaCl + 0.5mmr CaSO . The initial uptake to both sets of rootsat 230 was the same (8.0 ueq/g*hr). The free spaceuptake at 20 (open star) was 0.3 ,eq/g.
result of isotopic equilibration of the cytoplasm. Thecontenit of K+ in the cytoplasm of these roots cannotbe estimated with mnuch certainty, but tracer effluxmeasurements show it is not more than 1 yeq/g ofroot. The values of fluxes estimated here give amaximum time for lhalf-exchange of tracer in thecytoplasm of about 5 min and the real value is likelyto be very much less. The time for half-exchangeof the free space is about 2 min (115). Within 10
in, then, the cytoplasm specific activity will haverisen sufficiently for tracer uptake to be no longer*soO, but
* SO - o s., or,
poc *ocv so- ¢co0
vc *s
(kcs + c^
where the subscripts o, c. and v show direction offluxes (O) or location of specific activity (s) [cf.Cram (2)].
This view is supported by comparison of uptakeof 86Rb into AER roots (a) put from CaSO. tolabeled salt solution and (b) put from CaSO4 tounlabeled salt solution for 1 hr,. then transferred tolabeled solution. In the period 0 to 30 min uptakeinto (a) was 3.3 ,ueq/g and into (b) 1.9 jueq/g.From 30 to 60 min, uptake into (a) was 1.8 andinto (b) 1.9 ,ueq/g.
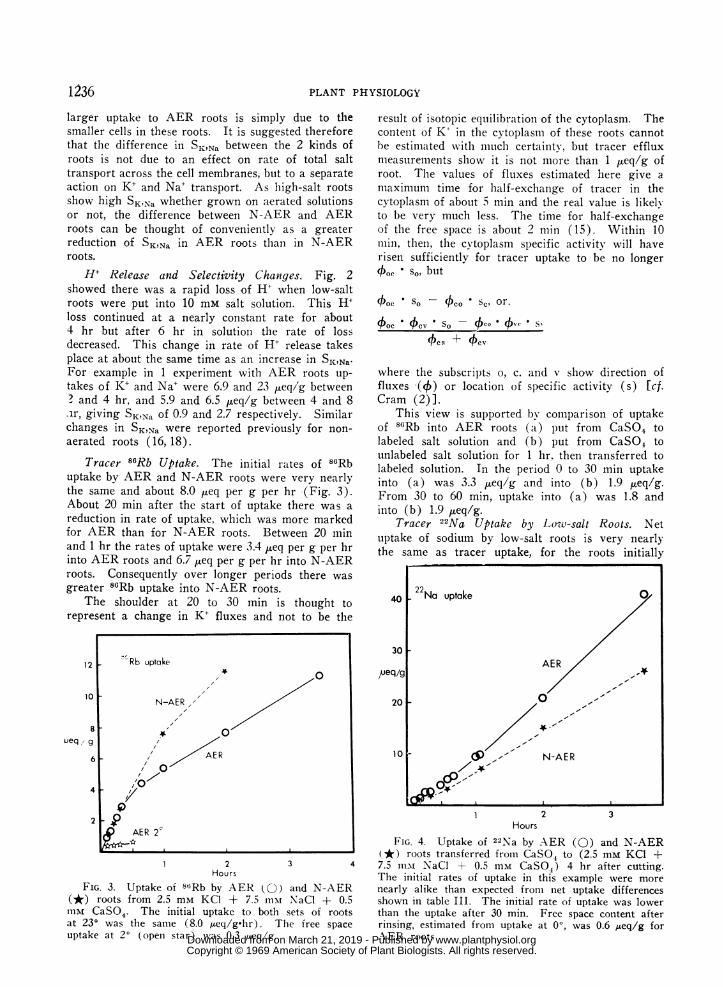
Tracer 22lNa Uptake by Low-salt Roots. Netuptake of sodium by low-salt roots is very nearlythe same as tracer uptake, for the roots initially
30
peq/g
10
2Hours
3
FIG. 4. Uptake of 22Na by AER (0) and N-AER(*) roots transferred from CaSO4 to (2.5 mM KCI +7.5 nim NaCl + 0.5 mm CaSO,) 4 hr after cutting.The initial rates of uptake in this example were morenearly alike than expected from net uptake differencesshown in table III. The initial rate of uptake was lowerthan the uptake after 30 min. Free space content afterrinsing, estimated from uptake at 00, was 0.6 Aeq/g forAER roots
.22Na uptake40 [
AER
201-
N-AER
1236
I
www.plantphysiol.orgon March 21, 2019 - Published by Downloaded from Copyright © 1969 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
pueq /
1 2Hours
3 4
FIG. 5. Comparison of tracer (open symbols) andniet K- uptake (closed symbols) to AER roots whichhad been kept in 0.5 mmi CaSO4 (a) for 15 min 0, 0,
(b) for 4 hr (open star), * after cutting. Freshly cutroots show initial loss of K+ followed by uptake at thesame rate as the tracer. Tracer uptake was the same forboth sets of roots and in good agreement with that inFig. 3. Sodium uptake was (a) 48 jueq/g; (b) 42.5,ueq/g, and chloride uptake (a) 388 ,eq/g, (b) 38.9,eq/g. The pH was maintained at 53.3.
contain only 2 ueq/g of sodium. Initial rates ofuptake of sodium into AER roots are greater thaninto N-AER roots, as would be expected from theresults in table III (Fig. 4). The rate of uptakeinto the roots increased by about 1.0 to 1.5 ,ueq per gper hr at about the same time as there was a decreasein 86Rb uptake (Fig. 4).
Pre-treatment of Excised Roots and K+ Uptake.Comparisons of K+ and Na+ uptake were complicatedby an effect of pre-treatment, particularly when usingAER roots. Roots kept in 0.5 mm CaSO4 for 2 to4 hr after cutting took up more K+ than either rootsof whole plants or excised roots used after a shortrinse in CaSO4 solution. For example uptakes ofK+, Na+ and Cl- in 4 hr from the standard solutionby roots of whole plants were 4.7, 48.5, and 42,ueq/g. Uptakes by roots kept in 0.5 mM CaSO4 for4 hr after cutting were 10.6, 41, and 43 FLeq/grespectively. Total salt uptake was the same butdifferent proportions of K+ and Na+ were accumu-lated. Fig. 5 shows that the difference in net K'uptake was largely due to efflux of K+ from theroots. Tracer uptake was the same in both cases.
This K+ efflux appeared to be from all the cellsof the cortex and not from the cut end of the stele.Measurements were made of K+ lost from the cutends and K+ uptake over the whole root surface.Using freshly cut roots in a solution of 0.1 mMKC1 + 10.0 mm NaCl + 0.5 mM CaSO4 there wasa loss of 0.4 ,ueq/g from the cut end and an uptakeof 1.1 /Leq/g-hr. When roots were kept for 4 hr inCaSO4 after cutting, loss from the cut end was thesame but uptake along the root was 4.0 iaeq per gper hr.
Table V. Effect of pH and Ca2+ on Uptake of Na+, K+,and 86Rb Over a Period of 4 Hr
The salt solution contained 25 mm KCl + 7.5 mMNaCl + 0.5 mM CaSO4 (except when Ca2+ was absent).Roots were kept in aerated 0.5 mm CaSO4 after cutting.
UptakepH Root Na+ K+ 86Rb
pH varied ,ueq perg per4hr4.3 AER 46 5.8 + 0.3 10.4 + 0.45.5 41 106 1.0 9.6 0.84.3 N-AER 34 9.4 1.0 16.0 0.45.5 34 16.0 0.6 15.2 0.4pH 5.5 Ca2+ variedCheck AER 52 8.4No Ca2+ 52 0.8Check N-AER 29 14.0No Ca2+ 28 13.0
Effect of pH and Ca2+ Level on K+ Uptake.Measurements of uptake over the first 4 hr in saltsolution are convenient for study of the effect ofHW and Ca2+ on K+ uptake. Roots rinsed in CaSO4for 4 hr were used.
At low pH, uptake into both AER and N-AERroots was less selective than at high pH. Table Vshows that the lower K+ uptake at low pHl was dueto greater K+ efflux. Tracer influx was not affectedby pH. Absence of Ca21 from the solution severelvreduced K+ uptake into AER roots, but uptake intoN-AER roots was the same as in the control. Theactions of pH, low Ca°+, and pretreatment were tosome extent additive. Using freshly cut roots in a
solution without Ca2' and at pH 4.6, there was aloss of 13.5 j,eq/g K+ from the tissue (Fig. 6).
Total salt uptake into both N-AER and AERroots was about 10 % less at pH 4.3 than at pH 5.5.Lack of Ca2+ did not affect uptake of Cl- into N-AERroots, but reduced Cl- uptake into AER roots byabout 20 %.
Relation to Other Published Data on K+ and Na+Uptake. Table VI compares results for K+ and Na+uptake from several authors. The differences inSK,Na within the same variety can be explainedadequately by the differences described in this paperbetween roots grown on aerated and non-aeratedCaSO4 solution, or by differences in the method usedto measure uptake. Uptake over the first few hris less selective than that over 24 hr (Fig. 2) andSKI,Na measured by tracer uptake can be larger thanthat measured by net uptake, particularly if there isefflux of K+.
In general, differences in K+ and Na+ uptakebetween varieties of barley do not seem to be aslarge as the changes in selectivity due to pretreat-ment or method of measuring uptake. Greenway(5) has also found that selectivity of uptake by anumber of varieties was very nearly the same despitedifferences in salt tolerance.
1086Rb and net K+uptake |
8
g~~~~~~~~~~~~~
24 __-0a
2*.
1237
I
www.plantphysiol.orgon March 21, 2019 - Published by Downloaded from Copyright © 1969 American Society of Plant Biologists. All rights reserved.

Table Vl. Co0miparison of Published Results of K+ andNa+ Uptake Fronm a Solu(tiont of 5 WvM KCI + 5 I?lM NaCI+ 0.5 Wi1M CaSO4
The pH was between 5 and 5.7. Temp 250 except Rains and Epstein's data at 300.
BarleyAuthor Ref variety SIC, Nn Type of measurement
Grown on aerated CaSO4Jackson and Stieff 8 Trebi 0.2 Net uptake in 3 hr
9 'II98 Compana 0.5 Initial net uptakeThis paper Compana 0.4 Net uptake in 4 hrHiatt I Compana 1.1 Net uptake in 24 hrRains and Epstein 18 Arivat 1.4 Initial tracer uptake
Grown on non-aerated CaSO4Pitman et al. 16 Cape 1.3 Tracer uptake in 2 hrPitman (unpublished ) Bolivia 1.1 Tracer uptake in 3 hr
Bolivia 2.2 Net uptake in 24 hrrhis paper Compana 1.6 Net uptake in 4 hr
1 Personal communication.
Discussion
Growing barley seedlings on CaSO, solution withaeration produced roots that differed in morphologyand physiology fronm roots grown without aeration.Roots grown without aeration had wider cells, largerair spaces and were shorter than those grown withaeration. There were also differences in sugar leveland relative abundance of the different amino acids
0
10 /S,,,010~~K + (a v
,ueq /9'g
/ o'~~~
v(b) -
'0
10 * ," (c)
*__ *
2 4 6 8 10 12Hours
FIG. 6. Changes in K+ level in AER roots aftertransfer to 2.5 mm KCl + 7.5 mM NaCl. (a) Solutionalso containing 0 5 mM CaSO4 and pH adjusted to 5.3,roots in CaSO1 for 30 min after cutting. (b) Solutionas (a) but pH not adjusted and roots used within 15min of cutting. The pH fell to 4.6. (c) No CaSO4added to the solution, otherwise as (b). Note theinitial loss of K- and recovery as in Fig. 5.
(table I). The higher level of alanine in non-aeratedroots has a parallel in the water fern Marsilea, forGaudet (4) found that the water form of this ferncontained very much more alanine (and ammoniaand glutamine) than the land form.
When these low-salt roots were put into saltsolutions containing K+ and Na' they showed a largedifference in selectivity for K-. Roots grown with-out aeration (N-AER) had much larger SK,Na thanroots grown witlh aeration (AER) (Fig. 2). Thesedifferences were found for a wide range of total(K+ + Na+) concentratioins. Differences betweenthe roots were also found in total salt uptake, but thelarger uptake by roots grown with aeration can beexplained by the larger sturface area of cells pergram of those roots compared with N-AER roots(table IV). The surface area cotuld not accouint forthe difference in SK,NI in the 2 kinds of root.
Tracer measurements showed that there was achange in Na+ and K+ influxes into both AER andN-AER roots about 20 to 30 miil after putting theroots into a 10 mM salt solution. The differencein SKINa seen in long term uptake into these roots(Fig. 2) was partly due to larger Na+ and smallerK+ influxes into AER roots, but also to a largerK+ efflux from AER than from N-AER roots. Thedevelopment of KI efflux was also responsible forthe low SK,Na for roots in solutions of low pH (cf.table V).
Despite the differences in uptake into AER andN-AER roots, both root preparations belhaved aslow-salt roots, having a large net uptake, and slhow-ing less ability to distinguish between K+ and Na+than higlh-salt roots. Fig. 1 and table II slhowedthat SKINa for low salt roots was muclh less than forroots grown in a culture solution. Similar differ-ences between roots of low- and high-salt status lhavealso been found for tracer influx (18).
Comparison of SK'Na and HE release shbows thatlow SK,5.Ia occurs when 1If release is hiiglh. Thus
1238 PLANT PHYSIOLOGY
www.plantphysiol.orgon March 21, 2019 - Published by Downloaded from Copyright © 1969 American Society of Plant Biologists. All rights reserved.

1239PITMAN-AERATION AND K, Na- UPTAKE BY BARLEY ROOTS
Fig. 2 showed that rate of H+ release decreasedduring salt accumulation by AER roots; at the sametime SK,Na increased. Again SK,Na was much lowerand H+ loss more rapid from AER than N-AERroots. As another example table III showed thatSKNNa in 0.2 mM solution was higher than in 10 mM:H' release (not reported) was much less from rootsin the low concentration. In 10 mN sulphate solu-tion SK,Na was low. H' release into SO42- was8 ,ueq/g hr compared with 4 Leq/g hr into C1- solu-tion, possibly due to organic acid formation by rootsin the S042- solution (6).
Hydrogen ions released from the cell membranemust diffuse from the site of formation to the ex-ternal solution. Due to the diffusion resistance,high rates of H+ release should lead to large localconcentrations of H+ ions. As the cell wall has theproperties of a Donnan system of negative chargeswith pKa of about 2.8 (15), high H+ concentrationswould reduce local density of fixed changes and soreduce Ca2+ concentrations at the cell wall/membraneinterface. Both TI+ and Ca2+ ions have effects onmembrane permeability and/or ion transport.
Low pH and/or low Ca2+ lead to increased per-meability of the cell membranes. This effect is shownby the increased K' efflux at pH 4.3 compared withpH 5.5 (table V) and the K+ loss from roots insolutions lacking Ca2+ (Fig. 6). There are othersamples of this action in the literature (e.g. 3,9, 12,22) and the interaction of Ca2' and H+ to increasemembrane permeability has been discussed by Mar-schner and Mengel (10,11).
Active uptake of salt does not seem to be affectedby low pH to the same extent as cell permeability.In table V uptake of tracer was the same at pH 4.3and 5.5, and total salt uptake was only 10 % lower atpH 4.3 than at 5.5. Jackson and Edwards (7) alsofound that variation in pH in the range 4 to 7 hadlittle effect on K+ and Cl- influxes from KCI solu-tions. Similarly Rains et al. (21) studying uptakeof K+ at low concentrations (Mechanism I uptake)found that 80Rb influx at pH 4.1 was only 10 %lower than at pH'5.7.
Welch and Epstein (25) have shown recentlythat Mechanism I is responsible for almost all theK+ uptake from solutions containing large propor-tiolns of Na' (sucli as the ones used in these experi-ments). They found that K+ uptake from 0.1 mMKCl and 10 nim NaCl was very nearly the same asthat from 0.1 mm KCI and from 5.0 mm KCI +10.0 mm NaCl. This Mechanism I process is thesame, I think, as the active K+ influx shown to bepresent in higlh salt roots (17), and located at theplasmalemma. Mechanism I appears to hiave a valueof about 4 to 6 Aeq/ghir at 250 while total salt uptakefrom a 10 mM Cl- solution is about 10 to 12 peqper g per hr.
To account for clhanges in selectivity as low saltroots accumulated salt, I suggested (18) that changes
in selectivity reflected changes in cytoplasmic K+ andNa+. The combination of active K+ uptake (Mecha-nism I) and effect of H+ ions on cell permeabilityshows one way that this change in selectivitv couldbe induced.
In high-salt roots a high value of K+/Na+ in thecytoplasm can be maintained by the Mechanism Iinflux of K+ and possibly by an Na+ active effluxtoo (17). The effectiveness of this system dependson the "leak" of Na' into the cell past the pumps;if the cell permneability to Na' is low then K+/Na"will be high in the cytoplasm. Increasing cell per-meability by the action of H+ ions would increaseNa+ influx and so reduce the ratio of K' to Na+ inthe cytoplasm. Selectivity would be further reducedif H+ loss from the cell is an active secretion whichtakes place instead of Na+ efflux in the low-saltcondition. Substantiation of this hypothesis reqtuiresmeaFurements of permeability of the cell and of Na+influx at different stages of uptake. It is doubtfulhow much reliance could he placed on these kinds ofmeasurements with present techniques. Nonetheless,this hypothesis allows behavior of low- and high-saltroots to be explained on the same model, and hasthe experimental support of the relationship betweenH+ and SK,Na described above.
This comparison of uptake into roots grown onaerated and non-aerated solutions has the advantagethat the plant material is genetically the same thoughmodified by environment. Apart from the usefulnessof this comparison for salt uptake study, it is agood example of the way plant roots may becomeadapted morphologically and physiologically to localdifferences in environment. Plants growing in soilcan experience fluctuations in their environment. Assynthesis (or activation) of enzymes is a relativelyrapid process, adaptational responses of cell metabo-lism could occur within 12 hr of an environmentalchange. A study of adaptations of cell metabolismto, for example, low water availability, high saltlevels, or flooding should be a necessary part of thestudy of plant nutrition.
Acknowledgments
This work was carried our during tenure of a Post-doctoral Research Association, National Research Coun-cil, National Academy of Sciences, at United StatesDepartment of Agriculture, Beltsville, Maryland.
I am grateful to Dr. Sterling B. Hendricks, Dr. JamesE. Leggett, and Mrs. Patricia C. Jackson for helpfuldiscussion, and to Miss M. E. Engelhaupt for analysisof amino acids in barley roots.
Literature Cited
1. BRYANT, A. E. 1934. Comparison of anatomicaland histological differences between roots of barleyseedlings grown in aerated and in non-aeratedsolutions. Plant Physiol. 9: 389-91.
www.plantphysiol.orgon March 21, 2019 - Published by Downloaded from Copyright © 1969 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
2. CRAM, W. J. 1968. Compartmentation and ex-change of chloride in carrot root tissue. Biochim.Biophys. Acta 136: 339-53.
3. EPSTEIN, E. 1961. The essential role of calciumin selective cation transport by plant cells. PlantPhysiol. 36: 437-44.
4. GAUDET, J. J. 1968. The correlation of physio-logical differences and various leaf forms of anaquatic plant. Physiol. Plantarum 21: 594-601.
5. GREENWAv, H. 1962. Plant response to salinesubstrates. I. Growth and ion uptake of severalvarieties of Hordeumii during and after sodiumchloride treatment. Australian J. Biol. Sci. 15:16-38.
(. HIATT, A. J. 1967. Relationship of cell sap toorganic acid change during ion uptake. PlantPhysiol. 42: 294-98.
7. JACKSON, P. C. AND D. G. EDWARDS. 1966. Cationeffects oIn chloride fluxes and accumulation levelsin barley roots. J. Gen. Physiol. 50: 225-41.
8. JACKSON, P. C. AND K. J. STIEFF. 1965. Equili-brium and ion exchange characteristics of potassiumand sodium accumulation by barley roots. J. Gen.Physiol. 48: 601-16.
9. LEGGETT, J. E. AND W. A. GILBERT. 1967. Locali-zation of the Ca-mediated apparent ion selectivityin the cross-sectional volume of soybean roots.Plant Physiol. 42: 1658-64.
10. MARSCHNER, H. 1964. Einfluss von Calcium aufdie Natrium Aufnahme und die Kalium Abgabeisolierter Gerstenwurzeln. Z. Pflanzenern5hr.Dueng. Bodenk. 107: 20-32.
11. MARSCHNER, H. AND K. MENGEL. 1966. Der Ein-fluss von Ca- und H-Ionen bei unterschiedlichenStoffwechselbedingungen auf die Membraneperme-abilitat junger Gerstenwurzeln. Z. Pflanzener-nahr. Dueng. Bodenk. 112: 3949.
12. PITMAN, M. G. 1964. The effect of divalent ca-tion on the uptake of salt by beetroot tissue. J.Exptl. Botany 15: 444 56.
13. PITMAN, M. G. 1965. Sodium and potassium up-take by seedlings of Hordeumn vdlgare. AustralianJ. Biol. Sci. 18: 10-24.
14. PITMAN. M. G. 1965. Tranispiration and the se-
lective uptake of potassium by barley seedlings(Hordeum zvlgare cv. Bolivia). Australian J.Biol. Sci. 18: 987-98.
15. PITMAN, M. G. 1965. Ion exchange and diffusionin roots of Hordeutm vulgare. Australian J. Biol.Sci. 18: 541-46.
16. PITMAN, M. G. 1967. Conflicting measurements ofsodium and potassium uptake by barley roots.Nature 216: 1343-44.
17. PITMAN, M. G. AND H. W. SADDLER. 1967. Activesodium and potassium transport in cells of barleyroots. Proc. Natl. Acad. Sci. 57: 44-52.
18. PITMAN, M. G., A. C. COURTICE, AND B. LEL 1968.Comparison of potassium and sodium uptake bybarley roots at high and low salt status. Aus-tralian J. Biol. Sci. 21: 871-81.
19. RAINS, D. W. AND E. EPSTEIN. 1967. Sodiumabsorption by barley roots: Role of the dualmechanisms of alkali cation transport. Plant Phy-siol. 42: 314-18.
20. RAINs, D. W. AND E. EPSTEIN. 1967. Sodiumabsorption by barley roots: Its mediation by mech-anism of alkali cation transport. Plant Physiol.42: 319-23.
21. RAINS, D. W., W. E. SCHMID., AND E. EPSTEIN.1964. Absorption of cations by roots. Effects ofhydrogen ions and essential role of calcium. PlantPhysiol. 39: 274-78.
22. SIEGEL, S. M. AND 0. DALY. 1966. Regulation ofbetacyanin efflux from beet root by poly-L-lysine,Ca-ion, and other substances. Plant Physiol. 41:1429-34.
23. VAN DER HEIDE, H., B. M. DE BOER-BOLT, AND M.H. VAN RAALTE. 1963. The effect of a low oxy-gen content of the medium on the roots of barleyseedlings. Acta Botan. Neerl. 12: 231-47.
24. VLAMIS, J. AND A. R. DAVIS. 1944. Effects ofoxygen tension on certain physiological responsesof rice, barley, and tomato. Plant Physiol. 19:33-51.
25. WELCH, R. M. AND E. EPSTEIN. 1968. The dualmechanisms of alkali cation absorption by plantcells; their parallel operation across the plas-Inalemma. Proc. Nat]. Acad. Sci. 61: 447-53.
1240
www.plantphysiol.orgon March 21, 2019 - Published by Downloaded from Copyright © 1969 American Society of Plant Biologists. All rights reserved.