metformin, an antidiabetic agent, suppresses the...
TRANSCRIPT

Metformin, an Antidiabetic Agent, Suppresses the Productionof Tumor Necrosis Factor and Tissue Factor by Inhibiting EarlyGrowth Response Factor-1 Expression in Human Monocytesin Vitro
Masatoku Arai, Mitsuhiro Uchiba, Hidefumi Komura, Yuichiro Mizuochi, Naoaki Harada,and Kenji OkajimaDepartments of Translational Medical Science Research (M.A., N.H., K.O.) and Anesthesiology and Medical Crisis Management(H.K., Y.M.), Nagoya City University Graduate School of Medical Sciences, Nagoya, Japan; and Department of BloodTransfusion and Cell Therapy, Kumamoto University Hospital, Kumamoto, Japan (M.U.)
Received December 24, 2009; accepted March 31, 2010
ABSTRACTMetformin, an antidiabetic agent, has been shown to reduceatherothrombotic disease in diabetic patients independent ofantihyperglycemic effect. Recent studies have demonstratedthat metformin attenuates the proinflammatory responses inhuman vascular wall cells and macrophages. However, thedetailed molecular mechanisms underlying these therapeuticeffects remain unclear. In the present study, we investigatedthe effects of metformin on tumor necrosis factor (TNF) pro-duction and tissue factor (TF) expression in isolated humanmonocytes stimulated with lipopolysaccharide (LPS) or oxi-dized low-density lipoprotein (oxLDL). Metformin significantlyinhibited both TNF production and TF expression in isolated
human monocytes stimulated with LPS or oxLDL. Metforminalso significantly inhibited TNF and TF mRNA in humanmonocytes stimulated with LPS. Although metformin did notinhibit the activation of either nuclear factor-�B or activatorprotein-1, it inhibited the expression of early growth re-sponse factor-1 (Egr-1) and phosphorylation of extracellularsignal-regulated protein kinase (ERK) 1/2 in monocytes stim-ulated with LPS or oxLDL. These results suggest that met-formin may attenuate the inflammatory responses, at least inpart, by suppressing the production of both TNF and TFthrough the inhibition of the ERK1/2-Egr-1 pathway in humanmonocytes.
Metformin is one of the most commonly used antidiabeticagents. It improves insulin sensitivity in patients with type 2diabetes (Stumvoll et al., 1995). In addition, metformin re-duces the risk of atherothrombotic disease accompanied withdiabetes independent of its antihyperglycemic effect (UKProspective Diabetes Study Group, 1998).
It is now well accepted that atherosclerosis is not merely alipid disorder, but also an inflammatory disease (Ross, 1999;Libby, 2002). Inflammatory cells, such as monocytes/macro-phages, play a critical role in the initiation, progression, andcomplication of atherosclerotic lesions (Ross, 1999; Libby,2002).
Various cytokines contribute to pathogenesis of inflamma-
tion observed in atherosclerosis (Kleemann et al., 2008).Among these cytokines, tumor necrosis factor (TNF) plays animportant role. TNF is shown to be present in atheroscleroticlesions but not in normal vessels in humans (Tipping andHancock, 1993). TNF is associated with the progression ofatherosclerotic lesions by regulating the functions of vascularwall cells to promote adhesion, migration, growth, and acti-vation of leukocytes (Young et al., 2002), thus leading to thefurther accumulation of mononuclear phagocytes and ampli-fication of inflammation (Ross, 1999; Libby, 2002). TNF isalso associated with complications of atherosclerosis by in-creasing the production of matrix metalloproteinases contrib-uting to the vulnerability of the fibrous cap (Young et al.,2002).
The activation of coagulation also contributes to the patho-genesis of atherosclerosis (Ross, 1999; Libby, 2002). Tissuefactor (TF) is an important factor initiating blood coagulation
Article, publication date, and citation information can be found athttp://jpet.aspetjournals.org.
doi:10.1124/jpet.109.164970.
ABBREVIATIONS: TNF, tumor necrosis factor; AMPK, AMP-activated protein kinase; AP-1, activator protein-1; Egr-1, early growth responsefactor-1; ELISA, enzyme-linked immunosorbent assay; ERK, extracellular signal-regulated protein kinase; JNK, c-Jun N-terminal kinase; LPS,lipopolysaccharide; NF-�B, nuclear factor-�B; oxLDL, oxidized low-density lipoprotein; TF, tissue factor; RT-PCR, reverse transcription-poly-merase chain reaction; I��, inhibitor �B.
0022-3565/10/3341-206–213$20.00THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS Vol. 334, No. 1Copyright © 2010 by The American Society for Pharmacology and Experimental Therapeutics 164970/3596086JPET 334:206–213, 2010 Printed in U.S.A.
206
at ASPE
T Journals on July 6, 2018
jpet.aspetjournals.orgD
ownloaded from

(Edgington et al., 1991), which is expressed in atheroscleroticlesions in humans (Tipping et al., 1989; Wilcox et al., 1989).Current evidence suggests that the exposure of TF-positivemonocyte-derived macrophages to blood on plaque disruptionmay trigger both intravascular thrombosis and a suddenprogression of atherosclerotic lesions (Libby, 2002).
Although various causes contributing to the pathogenesisof inflammation observed in atherosclerosis have been iden-tified, oxidized low-density lipoprotein (oxLDL) in hypercho-lesterolemia and lipopolysaccharide (LPS) from infectiousmicroorganisms are recognized as potentially importantsources of chronic inflammation in the development of ath-erosclerotic lesions (Ross, 1999; Libby, 2002).
In the present study, we examined whether metformininhibits TNF production and TF expression in human mono-cytes stimulated with LPS or oxLDL and attempted to iden-tify the underlying molecular mechanisms.
Materials and MethodsMaterials. Metformin (1,1-dimethylbiguanide) was a generous
gift from Nippon Shinyaku Co., Ltd. (Kyoto, Japan). LPS (Esche-richia coli, serotype 055:B5) was purchased from Sigma-Aldrich (St.Louis, MO). oxLDL was purchased from Intracel Resources, LLC(Frederick, MD). Polyclonal rabbit antibodies against inhibitor �B(I��) �, p38, phosphorylated p38 (Thr180/Tyr182), c-Jun N-terminalkinase (JNK), phosphorylated JNK (Thr183/Tyr185), extracellularsignal-regulated protein kinase (ERK) 1/2, phosphorylated ERK1/2(Thr202/Tyr204), early growth response factor-1 (Egr-1), andcaspase-3 were purchased from Cell Signaling Technology (Danvers,MA). Antibodies against PU.1 and �-actin were obtained from SantaCruz Biotechnology, Inc. (Santa Cruz, CA). All reagents used were ofanalytical grade.
Monocyte Preparation and Incubation. Human peripheralblood mononuclear cells were isolated from buffy coats provided bythe local Central Institute of Blood Transfusion or obtained fromhealthy volunteer blood donors by Ficoll-Hypaque density gradi-ent centrifugation on Lymphoprep (Axis-Shield poC AS, Oslo,Norway) and then were further fractionated as described previ-ously (Uchiba et al., 1997). This study was performed with theapproval of the ethics committee of the Nagoya City UniversityGraduate School of Medical Sciences, and blood donors enrolledfor the study gave informed consent in accordance with the Dec-laration of Helsinki. To minimize any possible interindividualvariation among blood donors, each experiment was conducted byusing monocytes from two to four buffy coats and repeated threetimes by using independent mononuclear cell preparations fromdifferent donors. The mononuclear cells in plastic dishes withRPMI medium 1640 (Invitrogen, Carlsbad, CA) supplementedwith 1% calf serum (HyClone Laboratories, Logan, UT) were in-cubated for 16 h at 37°C in a humidified 5% CO2 incubator.Lymphocytes were then removed from the adherent monocytes byrepeated rinsing with serum-free RPMI medium 1640. The result-ing cell preparations were �90% monocytes, as determined byMay-Giemsa staining. Cell viability was �95%, as determined bya trypan blue dye exclusion test. Monocytes thus obtained wereadjusted to an appropriate volume and cultured in RPMI medium1640 supplemented with 1% calf serum at 37°C in a humidified 5%CO2 incubator. Various concentrations of metformin were addedto cells at 2 h before stimulation with LPS (100 ng/ml) or oxLDL(8 �g/ml). After further incubation under the same culture condi-tions for the required period, the cell suspensions were centri-fuged. The resulting supernatant fractions were used to deter-mine the levels of TNF, and the sedimented mononuclear pelletswere used for TF activity measurements.
Measurement of TNF Level. Human monocytes (1 � 106 cells/assay) were stimulated with LPS for 6 h or oxLDL for 12 h in thepresence or absence of metformin. The concentrations of TNF inculture media were determined by using an enzyme-linked immu-nosorbent assay (ELISA) kit for human TNF (BioSource Interna-tional, Camarillo, CA).
Measurement of TF Activity. Human monocytes (1 � 106 cells/assay) were stimulated with either LPS or oxLDL for 6 h in thepresence or absence of metformin. The TF activities on monocyteswere measured as described previously (Molor-Erdene et al., 2005).In brief, human monocytes (1 � 106 cells) were washed twice by aphosphate-buffered saline and then stored at �80°C until TF activitywas measured. After thawing, the cell pellets were sonicated for 30 sto scrap the monocyte pellets and then dissolved in 100 �l of clottingbuffer (12 mM sodium acetate, 7 mM diethylbarbitate, and 130 mMsodium chloride). Fifty microliters of the resuspended cells wasmixed with 50 �l of citrated plasma, and the clotting times weremeasured after recalcification with 50 �l of 20 mM CaCl2 solution at37°C. The TF equivalents were determined by using a standardcurve obtained from rabbit brain thromboplastin (Neoplastin Plus;Roche Diagnostic, Mannheim, Germany).
Detection of Specific Binding of p65, p50, and c-Fos to DNAby Enzyme-Linked Immunosorbent Assay. Human monocytes(1 � 107 cells/assay) were stimulated with either LPS or oxLDL for1 h in the presence or absence of metformin. Nuclear extracts wereprepared as described previously (Yuksel et al., 2003). The specificbinding of p65, p50, and c-Fos to their DNA consensus oligonucleo-tides was evaluated in nuclear extracts by using ELISA-based assaykits (Trans AM; Active Motif Inc., Carlsbad, CA) as described previ-ously (Molor-Erdene et al., 2005).
RNA Isolation and Quantitative mRNA Analysis. RT-PCRassays were used to assess TNF and TF mRNA levels in culturedhuman monocytes. Total RNA was extracted from cultured humanmonocytes by using TRIzol reagent (Invitrogen) according to themanufacturer’s instruction. This procedure yielded 5 to 10 �g oftotal RNA from 5 � 105 cells of cultured human monocytes. RNAsamples were diluted in RNase-free water and stored at �80°Cuntil they were used. Real-time PCR was performed by using theABI PRISM 7700 Sequence Detection System, TaqMan One-StepRT-PCR Master Mix Reagents Kit, and commercially availablepredesigned, gene-specific primers and FAM-labeled probe setsfor quantitative gene expression (TaqMan Gene Expression As-says; human TNF code Hs00174128_m1, human TF codeHs00175225_ml, and human glyceraldehyde 3-phosphate dehy-drogenase code Hs99999905_m1; Applied Biosystems, Foster City,CA). All probes used in these experiments spanned an exon–intronboundary. TNF, TF, and glyceraldehyde 3-phosphate dehydroge-nase mRNA were quantified by parallel estimation. The thermalcycler conditions were 30-min hold at 48°C and 10-min hold at95°C, followed by 40 cycles of 15 s at 95°C and 1 min at 60°C.
Western Blot Analysis. Human monocytes (2 � 106 cells/assay)were stimulated with LPS or oxLDL for various times in the presenceor absence of metformin (10 �M). Whole-cell lysates were collected asdescribed previously (Komura et al., 2008). Samples containingequal amounts of protein were separated by using sodium dodecylsulfate-polyacrylamide gel electrophoresis, and after electrophoresis,the proteins were transferred onto a polyvinylidene difluoride mem-brane (Bio-Rad Laboratories, Hercules, CA). The membranes wereincubated with appropriate antibodies at 4°C overnight and thensubsequently with horseradish peroxidase-conjugated secondary an-tibody for 1 h at room temperature. Specific proteins were visualizedby using an enhanced chemiluminescence system (GE Healthcare,Little Chalfont, Buckinghamshire, UK). The densitometric quantifi-cation of the bands was performed by using National Institutes ofHealth Image software (version 1.61).
Cell Viability. Isolated monocytes were stimulated with LPS oroxLDL in the presence or absence of metformin. The number ofmonocytes was counted at 6 h after stimulation with LPS or 12 h
Inhibition of Egr-1 Expression by Metformin 207
at ASPE
T Journals on July 6, 2018
jpet.aspetjournals.orgD
ownloaded from
after stimulation with oxLDL. Cell viability was evaluated by usinga trypan blue dye exclusion test (Uchiba et al., 1997).
Assessment of Apoptosis. Isolated monocytes were stimulatedwith LPS for 6 h or oxLDL for 12 h in the presence or absence ofmetformin (10 �M). Assessing the apoptosis of monocytes wasperformed by a Western blot analysis probed with anticaspase-3antibody.
Statistical Analysis. The values are distributed in parametricmanner and expressed as means S.D. of one experiment represen-tative of three separate experiments that gave similar results. Dif-ferences in TNF production, TF activity, and mRNA levels wereassessed by using analysis of variance followed by Scheffe’s post hoctest. In densitometric analysis data of Western blotting, the differ-ence between baseline values (time 0) and subsequent values foreach experiment was compared by using analysis of variance fol-lowed by Scheffe’s post hoc test, and the differences between thevalues with and without metformin treatment at each time pointwere compared by unpaired t test. These analyses were carried outwith StatView 5.0 software (SAS Institute, Cary, NC). Statisticalsignificance was defined as a level of p 0.05.
ResultsEffect of Metformin on the Production of TNF and
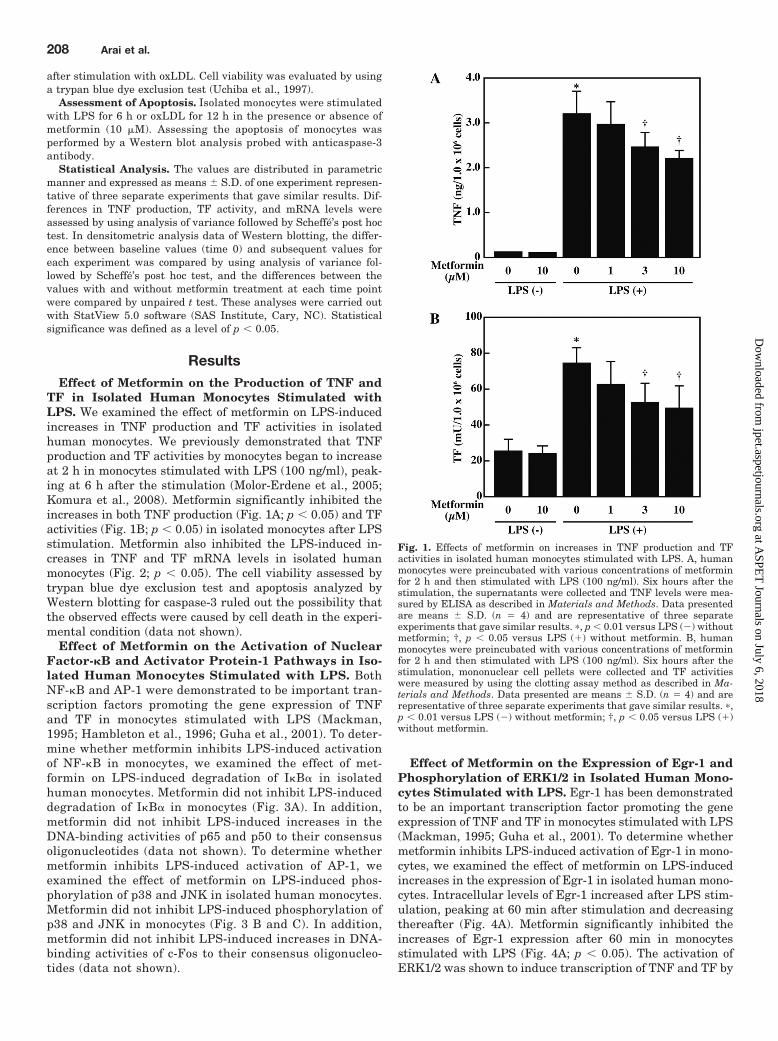
TF in Isolated Human Monocytes Stimulated withLPS. We examined the effect of metformin on LPS-inducedincreases in TNF production and TF activities in isolatedhuman monocytes. We previously demonstrated that TNFproduction and TF activities by monocytes began to increaseat 2 h in monocytes stimulated with LPS (100 ng/ml), peak-ing at 6 h after the stimulation (Molor-Erdene et al., 2005;Komura et al., 2008). Metformin significantly inhibited theincreases in both TNF production (Fig. 1A; p 0.05) and TFactivities (Fig. 1B; p 0.05) in isolated monocytes after LPSstimulation. Metformin also inhibited the LPS-induced in-creases in TNF and TF mRNA levels in isolated humanmonocytes (Fig. 2; p 0.05). The cell viability assessed bytrypan blue dye exclusion test and apoptosis analyzed byWestern blotting for caspase-3 ruled out the possibility thatthe observed effects were caused by cell death in the experi-mental condition (data not shown).
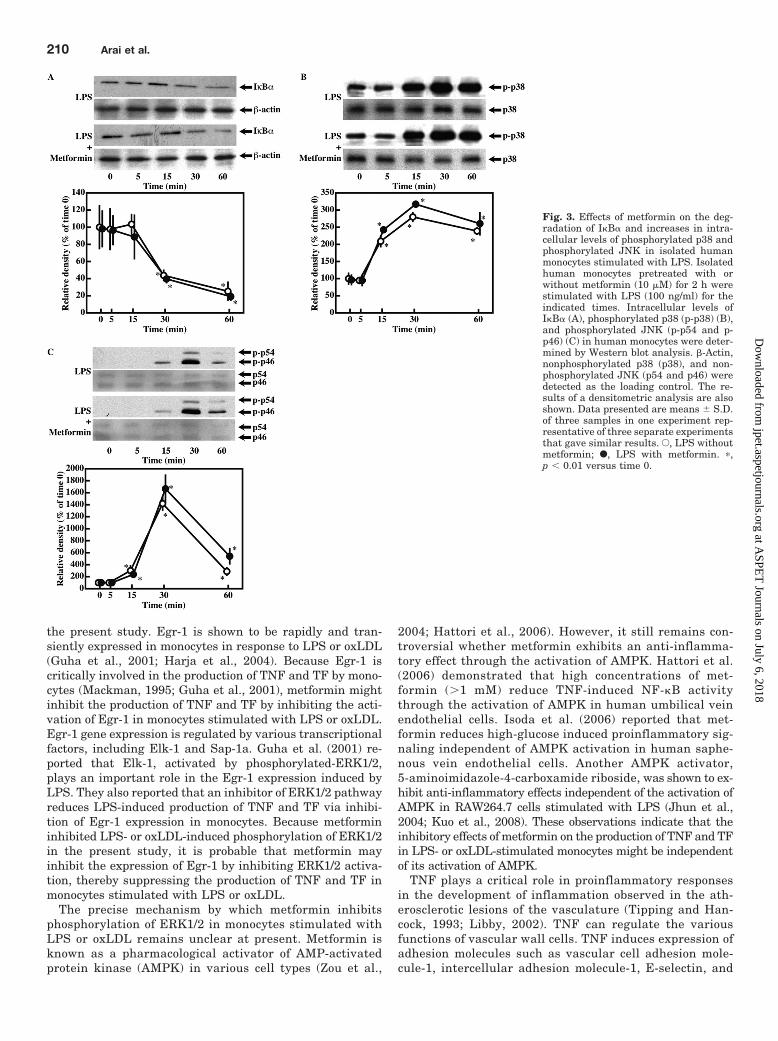
Effect of Metformin on the Activation of NuclearFactor-�B and Activator Protein-1 Pathways in Iso-lated Human Monocytes Stimulated with LPS. BothNF-�B and AP-1 were demonstrated to be important tran-scription factors promoting the gene expression of TNFand TF in monocytes stimulated with LPS (Mackman,1995; Hambleton et al., 1996; Guha et al., 2001). To deter-mine whether metformin inhibits LPS-induced activationof NF-�B in monocytes, we examined the effect of met-formin on LPS-induced degradation of I�B� in isolatedhuman monocytes. Metformin did not inhibit LPS-induceddegradation of I�B� in monocytes (Fig. 3A). In addition,metformin did not inhibit LPS-induced increases in theDNA-binding activities of p65 and p50 to their consensusoligonucleotides (data not shown). To determine whethermetformin inhibits LPS-induced activation of AP-1, weexamined the effect of metformin on LPS-induced phos-phorylation of p38 and JNK in isolated human monocytes.Metformin did not inhibit LPS-induced phosphorylation ofp38 and JNK in monocytes (Fig. 3 B and C). In addition,metformin did not inhibit LPS-induced increases in DNA-binding activities of c-Fos to their consensus oligonucleo-tides (data not shown).
Effect of Metformin on the Expression of Egr-1 andPhosphorylation of ERK1/2 in Isolated Human Mono-cytes Stimulated with LPS. Egr-1 has been demonstratedto be an important transcription factor promoting the geneexpression of TNF and TF in monocytes stimulated with LPS(Mackman, 1995; Guha et al., 2001). To determine whethermetformin inhibits LPS-induced activation of Egr-1 in mono-cytes, we examined the effect of metformin on LPS-inducedincreases in the expression of Egr-1 in isolated human mono-cytes. Intracellular levels of Egr-1 increased after LPS stim-ulation, peaking at 60 min after stimulation and decreasingthereafter (Fig. 4A). Metformin significantly inhibited theincreases of Egr-1 expression after 60 min in monocytesstimulated with LPS (Fig. 4A; p 0.05). The activation ofERK1/2 was shown to induce transcription of TNF and TF by
Fig. 1. Effects of metformin on increases in TNF production and TFactivities in isolated human monocytes stimulated with LPS. A, humanmonocytes were preincubated with various concentrations of metforminfor 2 h and then stimulated with LPS (100 ng/ml). Six hours after thestimulation, the supernatants were collected and TNF levels were mea-sured by ELISA as described in Materials and Methods. Data presentedare means S.D. (n � 4) and are representative of three separateexperiments that gave similar results. �, p 0.01 versus LPS (�) withoutmetformin; †, p 0.05 versus LPS (�) without metformin. B, humanmonocytes were preincubated with various concentrations of metforminfor 2 h and then stimulated with LPS (100 ng/ml). Six hours after thestimulation, mononuclear cell pellets were collected and TF activitieswere measured by using the clotting assay method as described in Ma-terials and Methods. Data presented are means S.D. (n � 4) and arerepresentative of three separate experiments that gave similar results. �,p 0.01 versus LPS (�) without metformin; †, p 0.05 versus LPS (�)without metformin.
208 Arai et al.
at ASPE
T Journals on July 6, 2018
jpet.aspetjournals.orgD
ownloaded from

increasing the expression of Egr-1 (Guha et al., 2001). Todetermine whether metformin inhibits LPS-induced activa-tion of ERK1/2, thereby suppressing Egr-1 expression, weexamined the effect of metformin on LPS-induced phosphor-ylation of ERK1/2 in isolated human monocytes. Intracellu-lar levels of phosphorylated ERK1/2 increased after LPSstimulation, peaking at 30 min and decreasing thereafter(Fig. 4B). Metformin significantly inhibited LPS-inducedphosphorylation of ERK1/2 in monocytes at 30 min afterstimulation (Fig. 4B; p 0.05).
Effect of Metformin on the Production of TNF andTF in Isolated Human Monocytes Stimulated withoxLDL. oxLDL has been reported to induce TNF productionand TF activity in isolated human monocytes in vitro(Jovinge et al., 1996; Petit et al., 1999). The effect of met-formin on oxLDL-induced increases in TNF production andTF activities was examined in isolated human monocytes.Metformin (10 �M) significantly inhibited the increases inboth TNF production (Fig. 5A; p 0.05) and TF activities
(Fig. 5B; p 0.05) in isolated monocytes at 12 and 6 h afteroxLDL (8 �g/ml) stimulation, respectively. The cell viabilityassessed by a trypan blue dye exclusion test and apoptosisanalyzed by Western blotting for caspase-3 ruled out thepossibility that the observed effects were caused by cell deathin the experimental conditions (data not shown).
Effect of Metformin on the Activation of NF-�B,AP-1, and ERK1/2-Egr-1 Pathways in Isolated HumanMonocytes Stimulated with oxLDL. We examinedwhether metformin inhibits oxLDL-induced activation ofNF-�B and AP-1 in monocytes. Metformin did not inhibitdegradation of I�B�, phosphorylation of p38 and JNK, orDNA-binding activities of p65, p50, and c-Fos in isolatedhuman monocytes stimulated with oxLDL (data not shown).
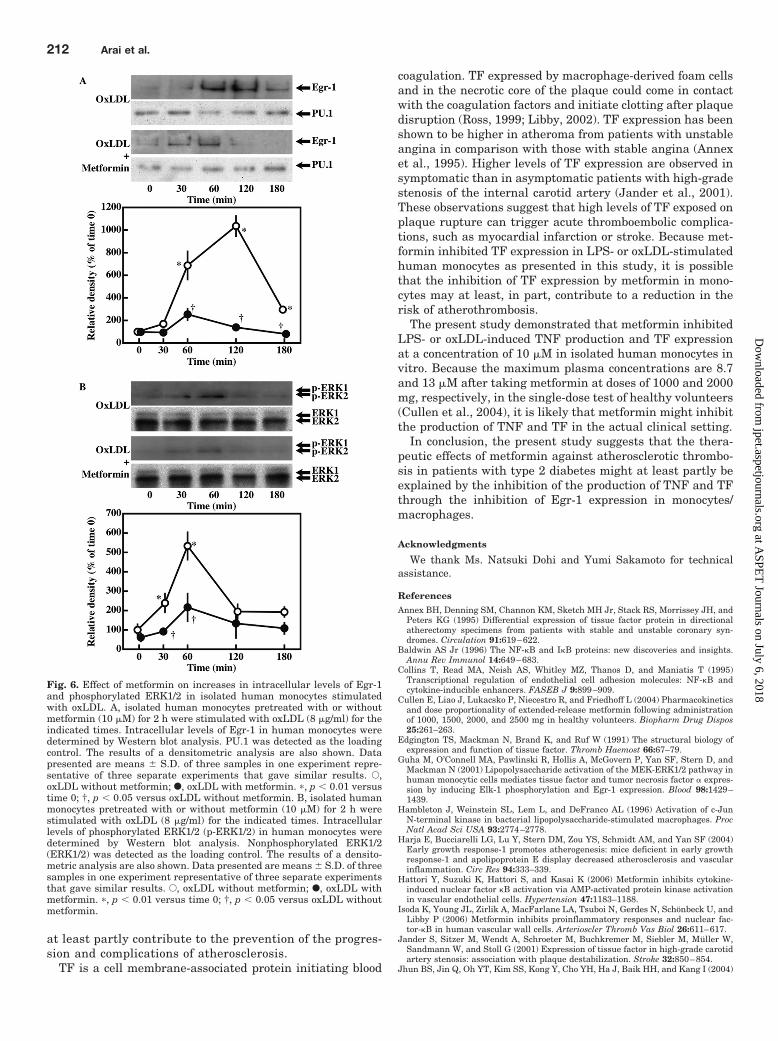
To determine whether metformin inhibits oxLDL-in-duced activation of Egr-1 in monocytes, we examined theeffect of metformin on oxLDL-induced increases in theexpression of Egr-1 in isolated human monocytes. Intra-cellular levels of Egr-1 increased after oxLDL stimulation,peaking at 120 min after stimulation and decreasing there-after (Fig. 6A). Metformin significantly inhibited the in-creases of Egr-1 expression at 120 min in monocytes stim-ulated with oxLDL (Fig. 6A; p 0.05). To determinewhether metformin inhibits oxLDL-induced activation ofERK1/2, thereby suppressing Egr-1 expression, we exam-ined the effect of metformin on oxLDL-induced phosphor-ylation of ERK1/2 in isolated human monocytes. Intracel-lular levels of phosphorylated ERK1/2 increased afteroxLDL stimulation, peaking at 60 min after stimulation anddecreasing thereafter (Fig. 6B). Metformin significantly inhib-ited oxLDL-induced phosphorylation of ERK1/2 in monocytes at60 min after stimulation (Fig. 6B; p 0.05).
DiscussionIn the present study, we demonstrated that metformin
inhibited the production of TNF and TF in isolated humanmonocytes stimulated with LPS or oxLDL.
The monocytic production of TNF and TF is regulated byvarious transcriptional factors including NF-�B, AP-1, andEgr-1 (Mackman, 1995; Jovinge et al., 1996; Guha et al.,2001). The activity of NF-�B is regulated primarily via itssequestration in the cytosol by anchoring to inhibitor proteinI�B� (Baldwin, 1996). As shown in the present study, pre-treatment with metformin did not affect the degradation ofI�B� or the increase in DNA-binding activity of p65 and p50induced by LPS or oxLDL. These results suggested that theinhibitory effect of metformin on the production of TNF andTF in LPS- or oxLDL-stimulated monocytes might not bemediated by the inhibition of NF-�B pathway.
AP-1 is another important factor regulating the productionof TNF and TF. The activation of p38 and JNK by phosphor-ylation was shown to enhance the transcriptional activity ofAP-1 (Mackman, 1995; Hambleton et al., 1996; Jovinge et al.,1996). In the present study, metformin did not inhibit LPS-or oxLDL-induced phosphorylation of JNK and p38 or theincrease in DNA-binding activity of c-Fos. These results sug-gest that the inhibitory effect of metformin on the productionof TNF and TF in monocytes might not be caused by theinhibition of AP-1 activation.
Metformin, on the other hand, inhibited LPS- or oxLDL-induced increases in intracellular levels of Egr-1 as shown in
Fig. 2. Effects of metformin on increases in TNF and TF mRNA inisolated human monocytes stimulated with LPS. A, TNF mRNA levels inisolated human monocytes preincubated with or without metformin (10�M) for 2 h were determined at 1 h after stimulation with LPS (100ng/ml). TNF mRNA levels in human monocytes were detected by quan-titative RT-PCR as described in Materials and Methods. Data presentedare means S.D. (n � 3) and are representative of three separateexperiments that gave similar results. �, p 0.01 versus LPS (�) withoutmetformin; †, p 0.05 versus LPS (�) without metformin. B, TF mRNAlevels in isolated human monocytes preincubated with or without met-formin (10 �M) for 2 h were determined at 1 h after stimulation with LPS(100 ng/ml). TF mRNA levels in human monocytes were detected byquantitative RT-PCR as described in Materials and Methods. Data pre-sented are means S.D. (n � 3) and are representative of three separateexperiments that gave similar results. �, p 0.01 versus LPS (�) withoutmetformin; †, p 0.05 versus LPS (�) without metformin.
Inhibition of Egr-1 Expression by Metformin 209
at ASPE
T Journals on July 6, 2018
jpet.aspetjournals.orgD
ownloaded from
the present study. Egr-1 is shown to be rapidly and tran-siently expressed in monocytes in response to LPS or oxLDL(Guha et al., 2001; Harja et al., 2004). Because Egr-1 iscritically involved in the production of TNF and TF by mono-cytes (Mackman, 1995; Guha et al., 2001), metformin mightinhibit the production of TNF and TF by inhibiting the acti-vation of Egr-1 in monocytes stimulated with LPS or oxLDL.Egr-1 gene expression is regulated by various transcriptionalfactors, including Elk-1 and Sap-1a. Guha et al. (2001) re-ported that Elk-1, activated by phosphorylated-ERK1/2,plays an important role in the Egr-1 expression induced byLPS. They also reported that an inhibitor of ERK1/2 pathwayreduces LPS-induced production of TNF and TF via inhibi-tion of Egr-1 expression in monocytes. Because metformininhibited LPS- or oxLDL-induced phosphorylation of ERK1/2in the present study, it is probable that metformin mayinhibit the expression of Egr-1 by inhibiting ERK1/2 activa-tion, thereby suppressing the production of TNF and TF inmonocytes stimulated with LPS or oxLDL.
The precise mechanism by which metformin inhibitsphosphorylation of ERK1/2 in monocytes stimulated withLPS or oxLDL remains unclear at present. Metformin isknown as a pharmacological activator of AMP-activatedprotein kinase (AMPK) in various cell types (Zou et al.,
2004; Hattori et al., 2006). However, it still remains con-troversial whether metformin exhibits an anti-inflamma-tory effect through the activation of AMPK. Hattori et al.(2006) demonstrated that high concentrations of met-formin (�1 mM) reduce TNF-induced NF-�B activitythrough the activation of AMPK in human umbilical veinendothelial cells. Isoda et al. (2006) reported that met-formin reduces high-glucose induced proinflammatory sig-naling independent of AMPK activation in human saphe-nous vein endothelial cells. Another AMPK activator,5-aminoimidazole-4-carboxamide riboside, was shown to ex-hibit anti-inflammatory effects independent of the activation ofAMPK in RAW264.7 cells stimulated with LPS (Jhun et al.,2004; Kuo et al., 2008). These observations indicate that theinhibitory effects of metformin on the production of TNF and TFin LPS- or oxLDL-stimulated monocytes might be independentof its activation of AMPK.
TNF plays a critical role in proinflammatory responsesin the development of inflammation observed in the ath-erosclerotic lesions of the vasculature (Tipping and Han-cock, 1993; Libby, 2002). TNF can regulate the variousfunctions of vascular wall cells. TNF induces expression ofadhesion molecules such as vascular cell adhesion mole-cule-1, intercellular adhesion molecule-1, E-selectin, and
Fig. 3. Effects of metformin on the deg-radation of I�B� and increases in intra-cellular levels of phosphorylated p38 andphosphorylated JNK in isolated humanmonocytes stimulated with LPS. Isolatedhuman monocytes pretreated with orwithout metformin (10 �M) for 2 h werestimulated with LPS (100 ng/ml) for theindicated times. Intracellular levels ofI�B� (A), phosphorylated p38 (p-p38) (B),and phosphorylated JNK (p-p54 and p-p46) (C) in human monocytes were deter-mined by Western blot analysis. �-Actin,nonphosphorylated p38 (p38), and non-phosphorylated JNK (p54 and p46) weredetected as the loading control. The re-sults of a densitometric analysis are alsoshown. Data presented are means S.D.of three samples in one experiment rep-resentative of three separate experimentsthat gave similar results. E, LPS withoutmetformin; F, LPS with metformin. �,p 0.01 versus time 0.
210 Arai et al.
at ASPE
T Journals on July 6, 2018
jpet.aspetjournals.orgD
ownloaded from

P-selectin (Marui et al., 1993; Collins et al., 1995; Taka-hashi et al., 1996; Young et al., 2002), crucial to the re-cruitment of mononuclear leukocytes to endothelial cells.TNF also increases the production of monocyte chemoat-tractant protein-1 and interleukin-8, which are capable ofinducing the migration of mononuclear leukocytes into theintima (Rollins et al., 1990; Young et al., 2002). TNF aug-ments the production of macrophage colony-stimulating
factor by vascular wall cells, which stimulates the transi-tion of monocytes to lipid-laden macrophages (foam cells).Consequently, it promotes the survival and growth of foamcells and increases production of cytokines and growthfactors by foam cells (Libby, 2002; Young et al., 2002).Therefore, TNF is involved in the accumulation of mono-nuclear phagocytes and amplification of inflammation ofthe lesions leading to the development of atherosclerosis.The degradation of extracellular matrix by matrix metal-loproteinases is thought to be important in plaque rupture(Young et al., 2002). TNF is shown to stimulate the expres-sion of various types of matrix metalloproteinases in en-dothelial cells and smooth muscle cells and macrophages(Young et al., 2002). Because the present study has shownthat metformin inhibits TNF production in LPS- or oxLDL-stimulated human monocytes, it is possible that the inhi-bition of TNF production by metformin in monocytes may
Fig. 4. Effect of metformin on increases in intracellular levels of Egr-1and phosphorylated ERK1/2 in isolated human monocytes stimulatedwith LPS. A, isolated human monocytes pretreated with or withoutmetformin (10 �M) for 2 h were stimulated with LPS (100 ng/ml) for theindicated times. Intracellular levels of Egr-1 in human monocytes weredetermined by Western blot analysis. PU.1 was detected as the loadingcontrol. The results of a densitometric analysis are also shown. Datapresented are means S.D. of three samples in one experiment repre-sentative of three separate experiments that gave similar results. E, LPSwithout metformin; F, LPS with metformin. �, p 0.01 versus time 0; †,p 0.05 versus LPS without metformin. B, isolated human monocytespretreated with or without metformin (10 �M) for 2 h were stimulatedwith LPS (100 ng/ml) for the indicated times. Intracellular levels ofphosphorylated ERK1/2 (p-ERK1/2) in human monocytes were deter-mined by Western blot analysis. Nonphosphorylated ERK1/2 (ERK1/2)was detected as the loading control. The results of a densitometric anal-ysis are also shown. Data presented are means S.D. of three samples inone experiment representative of three separate experiments that gavesimilar results. E, LPS without metformin; F, LPS with metformin. �, p 0.01 versus time 0; †, p 0.05 versus LPS without metformin.
Fig. 5. Effects of metformin on increases in TNF production and TFactivities in isolated human monocytes stimulated with oxLDL.A, human monocytes were preincubated with metformin (10 �M) for2 h and then stimulated with oxLDL (8 �g/ml). Twelve hours after thestimulation, the supernatants were collected, and TNF levels weremeasured by ELISA as described in Materials and Methods. Datapresented are means S.D. (n � 4) and are representative of threeseparate experiments that gave similar results. �, p 0.01 versusoxLDL (�) without metformin; †, p 0.05 versus oxLDL (�) withoutmetformin. B, human monocytes were preincubated with metformin(10 �M) for 2 h and then stimulated with oxLDL (8 �g/ml). Six hoursafter the stimulation, mononuclear cell pellets were collected, and TFactivities were measured by using the clotting assay method as de-scribed in Materials and Methods. Data presented are means S.D.(n � 4) and are representative of three separate experiments that gavesimilar results. �, p 0.01 versus oxLDL (�) without metformin; †,p 0.05 versus oxLDL (�) without metformin.
Inhibition of Egr-1 Expression by Metformin 211
at ASPE
T Journals on July 6, 2018
jpet.aspetjournals.orgD
ownloaded from
at least partly contribute to the prevention of the progres-sion and complications of atherosclerosis.
TF is a cell membrane-associated protein initiating blood
coagulation. TF expressed by macrophage-derived foam cellsand in the necrotic core of the plaque could come in contactwith the coagulation factors and initiate clotting after plaquedisruption (Ross, 1999; Libby, 2002). TF expression has beenshown to be higher in atheroma from patients with unstableangina in comparison with those with stable angina (Annexet al., 1995). Higher levels of TF expression are observed insymptomatic than in asymptomatic patients with high-gradestenosis of the internal carotid artery (Jander et al., 2001).These observations suggest that high levels of TF exposed onplaque rupture can trigger acute thromboembolic complica-tions, such as myocardial infarction or stroke. Because met-formin inhibited TF expression in LPS- or oxLDL-stimulatedhuman monocytes as presented in this study, it is possiblethat the inhibition of TF expression by metformin in mono-cytes may at least, in part, contribute to a reduction in therisk of atherothrombosis.
The present study demonstrated that metformin inhibitedLPS- or oxLDL-induced TNF production and TF expressionat a concentration of 10 �M in isolated human monocytes invitro. Because the maximum plasma concentrations are 8.7and 13 �M after taking metformin at doses of 1000 and 2000mg, respectively, in the single-dose test of healthy volunteers(Cullen et al., 2004), it is likely that metformin might inhibitthe production of TNF and TF in the actual clinical setting.
In conclusion, the present study suggests that the thera-peutic effects of metformin against atherosclerotic thrombo-sis in patients with type 2 diabetes might at least partly beexplained by the inhibition of the production of TNF and TFthrough the inhibition of Egr-1 expression in monocytes/macrophages.
Acknowledgments
We thank Ms. Natsuki Dohi and Yumi Sakamoto for technicalassistance.
ReferencesAnnex BH, Denning SM, Channon KM, Sketch MH Jr, Stack RS, Morrissey JH, and
Peters KG (1995) Differential expression of tissue factor protein in directionalatherectomy specimens from patients with stable and unstable coronary syn-dromes. Circulation 91:619–622.
Baldwin AS Jr (1996) The NF-�B and I�B proteins: new discoveries and insights.Annu Rev Immunol 14:649–683.
Collins T, Read MA, Neish AS, Whitley MZ, Thanos D, and Maniatis T (1995)Transcriptional regulation of endothelial cell adhesion molecules: NF-�B andcytokine-inducible enhancers. FASEB J 9:899–909.
Cullen E, Liao J, Lukacsko P, Niecestro R, and Friedhoff L (2004) Pharmacokineticsand dose proportionality of extended-release metformin following administrationof 1000, 1500, 2000, and 2500 mg in healthy volunteers. Biopharm Drug Dispos25:261–263.
Edgington TS, Mackman N, Brand K, and Ruf W (1991) The structural biology ofexpression and function of tissue factor. Thromb Haemost 66:67–79.
Guha M, O’Connell MA, Pawlinski R, Hollis A, McGovern P, Yan SF, Stern D, andMackman N (2001) Lipopolysaccharide activation of the MEK-ERK1/2 pathway inhuman monocytic cells mediates tissue factor and tumor necrosis factor � expres-sion by inducing Elk-1 phosphorylation and Egr-1 expression. Blood 98:1429–1439.
Hambleton J, Weinstein SL, Lem L, and DeFranco AL (1996) Activation of c-JunN-terminal kinase in bacterial lipopolysaccharide-stimulated macrophages. ProcNatl Acad Sci USA 93:2774–2778.
Harja E, Bucciarelli LG, Lu Y, Stern DM, Zou YS, Schmidt AM, and Yan SF (2004)Early growth response-1 promotes atherogenesis: mice deficient in early growthresponse-1 and apolipoprotein E display decreased atherosclerosis and vascularinflammation. Circ Res 94:333–339.
Hattori Y, Suzuki K, Hattori S, and Kasai K (2006) Metformin inhibits cytokine-induced nuclear factor �B activation via AMP-activated protein kinase activationin vascular endothelial cells. Hypertension 47:1183–1188.
Isoda K, Young JL, Zirlik A, MacFarlane LA, Tsuboi N, Gerdes N, Schonbeck U, andLibby P (2006) Metformin inhibits proinflammatory responses and nuclear fac-tor-�B in human vascular wall cells. Arterioscler Thromb Vas Biol 26:611–617.
Jander S, Sitzer M, Wendt A, Schroeter M, Buchkremer M, Siebler M, Muller W,Sandmann W, and Stoll G (2001) Expression of tissue factor in high-grade carotidartery stenosis: association with plaque destabilization. Stroke 32:850–854.
Jhun BS, Jin Q, Oh YT, Kim SS, Kong Y, Cho YH, Ha J, Baik HH, and Kang I (2004)
Fig. 6. Effect of metformin on increases in intracellular levels of Egr-1and phosphorylated ERK1/2 in isolated human monocytes stimulatedwith oxLDL. A, isolated human monocytes pretreated with or withoutmetformin (10 �M) for 2 h were stimulated with oxLDL (8 �g/ml) for theindicated times. Intracellular levels of Egr-1 in human monocytes weredetermined by Western blot analysis. PU.1 was detected as the loadingcontrol. The results of a densitometric analysis are also shown. Datapresented are means S.D. of three samples in one experiment repre-sentative of three separate experiments that gave similar results. E,oxLDL without metformin; F, oxLDL with metformin. �, p 0.01 versustime 0; †, p 0.05 versus oxLDL without metformin. B, isolated humanmonocytes pretreated with or without metformin (10 �M) for 2 h werestimulated with oxLDL (8 �g/ml) for the indicated times. Intracellularlevels of phosphorylated ERK1/2 (p-ERK1/2) in human monocytes weredetermined by Western blot analysis. Nonphosphorylated ERK1/2(ERK1/2) was detected as the loading control. The results of a densito-metric analysis are also shown. Data presented are means S.D. of threesamples in one experiment representative of three separate experimentsthat gave similar results. E, oxLDL without metformin; F, oxLDL withmetformin. �, p 0.01 versus time 0; †, p 0.05 versus oxLDL withoutmetformin.
212 Arai et al.
at ASPE
T Journals on July 6, 2018
jpet.aspetjournals.orgD
ownloaded from

5-Aminoimidazole-4-carboxamide riboside suppresses lipopolysaccharide-inducedTNF-� production through inhibition of phosphatidylinositol 3-kinase/Akt activa-tion in RAW 264.7 murine macrophages. Biochem Biophys Res Commun 318:372–380.
Jovinge S, Ares MP, Kallin B, and Nilsson J (1996) Human monocytes/macrophagesrelease TNF-� in response to ox-LDL. Arterioscler Thromb Vasc Biol 16:1573–1579.
Kleemann R, Zadelaar S, and Kooistra T (2008) Cytokines and atherosclerosis: acomprehensive review of studies in mice. Cardiovasc Res 79:360–376.
Komura H, Uchiba M, Mizuochi Y, Arai M, Harada N, Katsuya H, and Okajima K(2008) Antithrombin inhibits lipopolysaccharide-induced tumor necrosis factor-�production by monocytes in vitro through inhibition of Egr-1 expression. J ThrombHaemost 6:499–507.
Kuo CL, Ho FM, Chang MY, Prakash E, and Lin WW (2008) Inhibition of lipo-polysaccharide-induced inducible nitric oxide synthase and cyclooxygenase-2 geneexpression by 5-aminoimidazole-4-carboxamide riboside is independent of AMP-activated protein kinase. J Cell Biochem 103:931–940.
Libby P (2002) Inflammation in atherosclerosis. Nature 420:868–874.Mackman N (1995) Regulation of the tissue factor gene. FASEB J 9:883–889.Marui N, Offermann MK, Swerlick R, Kunsch C, Rosen CA, Ahmad M, Alexander
RW, and Medford RM (1993) Vascular cell adhesion molecule-1 (VCAM-1) genetranscription and expression are regulated through an antioxidant-sensitivemechanism in human vascular endothelial cells. J Clin Invest 92:1866–1874.
Molor-Erdene P, Okajima K, Isobe H, Uchiba M, Harada N, Shimozawa N, andOkabe H (2005) Inhibition of lipopolysaccharide-induced tissue factor expressionin monocytes by urinary trypsin inhibitor in vitro and in vivo. Thromb Haemost94:136–145.
Petit L, Lesnik P, Dachet C, Moreau M, and Chapman MJ (1999) Tissue factorpathway inhibitor is expressed by human monocyte-derived macrophages: rela-tionship to tissue factor induction by cholesterol and oxidized LDL. ArteriosclerThromb Vasc Biol 19:309–315.
Rollins BJ, Yoshimura T, Leonard EJ, and Pober JS (1990) Cytokine-activatedhuman endothelial cells synthesize and secrete a monocyte chemoattractant,MCP-1/JE. Am J Pathol 136:1229–1233.
Ross R (1999) Atherosclerosis: an inflammatory disease. N Engl J Med 340:115–126.Stumvoll M, Nurjhan N, Perriello G, Dailey G, and Gerich JE (1995) Metabolic
effects of metformin in non-insulin-dependent diabetes mellitus. N Engl J Med333:550–554.
Takahashi M, Ikeda U, Masuyama J, Kitagawa S, Kasahara T, Shimpo M, Kano S,and Shimada K (1996) Monocyte-endothelial cell interaction induces expression ofadhesion molecules on human umbilical cord endothelial cells. Cardiovasc Res32:422–429.
Tipping PG and Hancock WW (1993) Production of tumor necrosis factor and inter-leukin-1 by macrophages from human atheromatous plaques. Am J Pathol 142:1721–1728.
Tipping PG, Malliaros J, and Holdsworth SR (1989) Procoagulant activity expressionby macrophages from atheromatous vascular plaques. Atherosclerosis 79:237–243.
Uchiba M, Okajima K, Murakami K, Okabe H, and Takatsuki K (1997) Effect ofnafamostat mesilate on pulmonary vascular injury induced by lipopolysaccharidein rats. Am J Respir Crit Care Med 155:711–718.
UK Prospective Diabetes Study (UKPDS) Group (1998) Effect of intensive blood-glucose control with metformin on complications in overweight patients with type2 diabetes (UKPDS34). Lancet 352:854–865.
Wilcox JN, Smith KM, Schwartz SM, and Gordon D (1989) Localization of tissuefactor in the normal vessel wall and in the atherosclerotic plaque. Proc Natl AcadSci USA 86:2839–2843.
Young JL, Libby P, and Schonbeck U (2002) Cytokines in the pathogenesis ofatherosclerosis. Thromb Haemost 88:554–567.
Yuksel M, Okajima K, Uchiba M, and Okabe H (2003) Gabexate mesilate, a syntheticprotease inhibitor, inhibits lipopolysaccharide-induced tumor necrosis factor-�production by inhibiting activation of both nuclear factor-�B and activated pro-tein-1 in human monocytes. J Pharmacol Exp Ther 305:298–305.
Zou MH, Kirkpatrick SS, Davis BJ, Nelson JS, Wiles WG 4th, Schlattner U, Neu-mann D, Brownlee M, Freeman MB, and Goldman MH (2004) Activation of theAMP-activated protein kinase by the antidiabetic drug metformin in vivo. Role ofmitochondrial reactive nitrogen species. J BIol Chem 279:43940–43951.
Address correspondence to: Dr. Kenji Okajima, Department of Transla-tional Medical Science Research, Nagoya City University Graduate School ofMedical Sciences, Kawasumi 1, Mizuho-cho, Mizuho-ku, Nagoya City 467-8601, Japan. E-mail: [email protected]
Inhibition of Egr-1 Expression by Metformin 213
at ASPE
T Journals on July 6, 2018
jpet.aspetjournals.orgD
ownloaded from