traduccion reloaded.docx
TRANSCRIPT
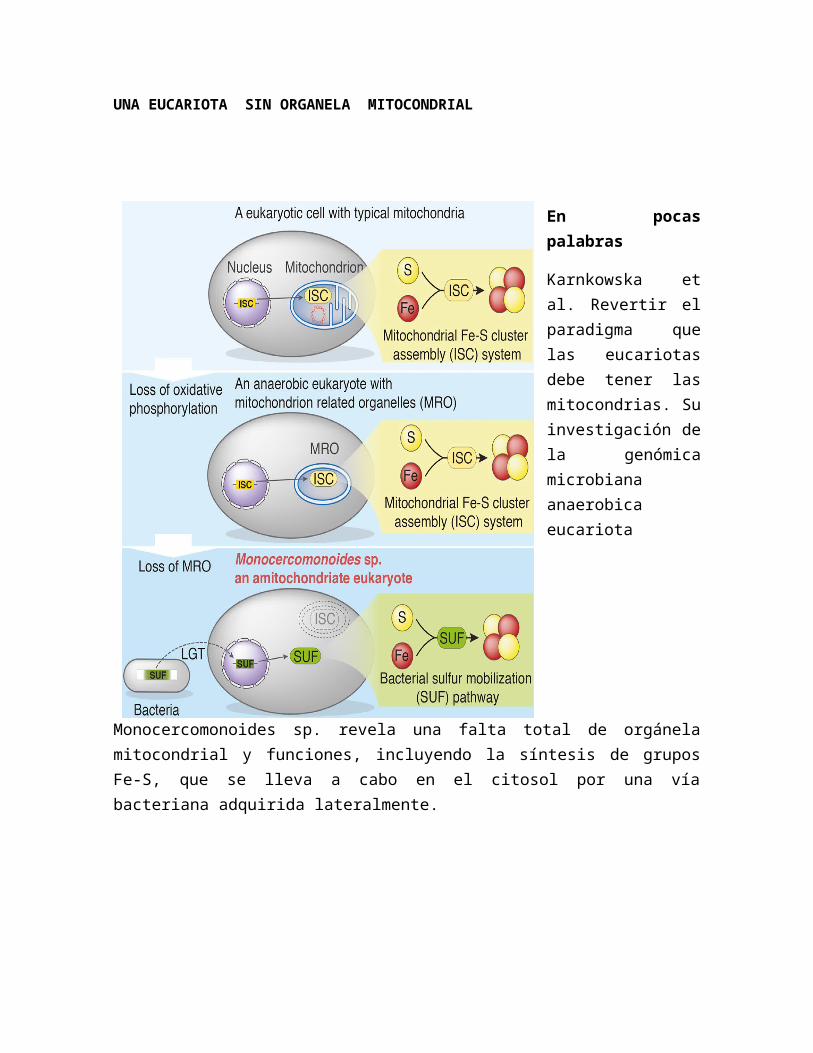
UNA EUCARIOTA SIN ORGANELA MITOCONDRIAL
En pocas palabras
Karnkowska et al. Revertir el paradigma que las eucariotas debe tener las mitocondrias. Su investigación de la genómica microbiana anaerobica eucariota Monocercomonoides sp. revela una falta total de orgánela mitocondrial y funciones, incluyendo la síntesis de grupos Fe-S, que se lleva a cabo en el citosol por una vía bacteriana adquirida lateralmente.
Destaca
Monocercomonoides sp. es un microorganismo eucariotico sin mitocondria La ausencia completa de la mitocondria es una pérdida secundaria, no una
característica ancestral El sendero ISC mitocondrial esencial fue sustituido por un sistema de SUF
bacteriana.

Una eucariota sin una orgánela mitocondrial
La presencia de mitocondrias y orgánelas relacionados en cada estudió eucariota apoya la opinión de que las mitocondrias son componentes celulares esenciales. Aquí presentamos la secuencia del genoma de un eucariota microbiana, la oxymonada Monocercomonoides sp., que reveló que este organismo carece de toda característica proteínas mitocondriales. Fundamentalmente, el hierro-azufre mitocondrial conjunto grupos externos, pensado para ser conservados en casi todas las células eucarióticas, ha sido sustituido por un sistema de movilización de azufre citosólico (SUF) adquirido por la transferencia lateral de genes de bacterias. En el contexto de eucariotas Filogenia, nuestros datos sugieren que no es primitiva la Monocercomonoides amitocondrial pero ha perdido la mitocondria secundaria. Este es el primer ejemplo de un eucariota carente de cualquier forma de mitocondria, demostrando que este orgánela no es absolutamente indispensable para la viabilidad de una célula eucariota.
INTRODUCCIÓN
Las mitocondrias son organelas que surgió a través de la integración endosimbiotica un proteobacterial proto-eucariota endosimbiontico en la célula huésped. Durante el curso de la evolución eucariota, el genoma y proteoma del compartimiento mitocondrial se ha modificado considerablemente, y se ha ganado muchas funciones, perdidas o reubicadas. En casos extremos, los derivados de las mitocondrias de protistas anaerobicas se han modificado que habían sido ignorados o no reconocidos como homólogas de la mitocondria. De hecho, en el decenio de 1980, la hipótesis Arquezoo propuso que algunos microbios eucariotas primitiva carecen de mitocondrias, peroxisomas apilados, aparato de Golgi, intrones spliceosomales, y reproducción sexual. Sin embargo, a lo largo de la siguiente década, orgánelas con doble membrana fueron identificados en todos los investigaciones de Arquezoo putativa.
El último clavo en el ataúd de la hipótesis Arquezoo fue la demostración de que estos orgánelas contienen algún marcador de proteínas mitocondrial, tales como aquellos implicados en el grupo de hierro-azufre (ISC) grupos Fe-S Sistema de Biogénesis, translocasas, maturasas y/o chaperonas moleculares conocidas para facilitar la importación de proteínas en la mitocondria. En la actualidad está ampliamente aceptados que las mitocondrias o organelas mitocondriales relacionados (OPF) compartimentos son esenciales en todas las células eucariotas y que la endosimbiosis mitocondrial tuvo lugar antes de que la radiación de todos las eucariotas existentes.
Metamonada, originalmente parte de la Arquezoo, ahora se clasifican como uno de los principales clados de las células eucariotas ''super-grupo'' Excavado y están compuestos

de eucariotas microaerofilias unicelulares o anaeróbicas que son a menudo parásitos o simbiontes especializados. Detallados estudios de biología molecular y celular, incluida la secuenciación del genoma, han sido realizados únicamente por tres especies parasitarias de dos linajes metamonad - Giardia intestinalis y Spiro nucleussalmonicida (Fornicata) y Trichomonas vaginalis (Parabasalia), que han proporcionado información importante acerca de las funciones de sus OPF. El tercer linaje de metamonadas, Preaxostyla, contiene el parafiletico basal de vida libre y los derivados endobioticos trimastigidas oxymonadas. La presencia de homólogos mitocondrial ha sido demostrado convincentemente en Paratrimastix (anteriormente Trimastix) piriformes, aunque las funciones bioquímicas de estos orgánelas son en gran parte desconocidos. Endobioticos oxymonadas pertenecen al menos estudiadas de la ex Arquezoo. Aquí presentamos el primer análisis de la secuencia completa del genoma de un Monocercomonoides oxymonadas, sp. PA203. Encontramos que, a pesar de que este organismo es una célula eucariota estándar en otros aspectos, carece por completo de cualquier rastro de una mitocondria.
RESULTADOS Y DISCUSIÓN
Características del genoma
Usando el 454 todo la secuencia metodológica del genoma rifle, hemos generado un proyecto de la secuencia del genoma de la oxymonada Monocercomonoides sp. PA203, reunidos en 2.095 matrices a ~353 Cobertura (véanse los procedimientos experimentales). El tamaño estimado del genoma (~75 Mb) y el número de genes codificadores de proteínas que predijo (16,629) es intermedio entre lo que se encuentra en diplomonads y T. vaginalis (Tabla 1). Casi el 67% de los genes codificantes de proteínas predicho contienen intrones (~1.9 intronspergenes en promedio; Tabla 1).El conjunto contiene genes que codifican tRNAs para todos los 20 aminoácidos, ADN ribosomal y ~50 unidades fueron identificadas en la pequeña contigs fuera de la parte principal de la Asamblea (Ver procedimientos experimentales complementarios). Para estimar la exhaustividad de la secuencia del genoma del transcriptoma, realizamos el mapeo, en el cual 96,9% de transcripciones correlacionado con el genoma (Ver procedimientos experimentales complementarios), y comprobar la representación del núcleo de los genes eucarióticas. Utilizando el enfoque de mapeo de genes eucarióticas Básico (CEGMA), hemos recuperado el 63,3% del núcleo de los genes eucarióticas, una mayor fracción que en el genoma de G. intestinalis (46,6%). Sin embargo, cuando se excluyeron los genes que codifican proteínas mitocondriales de la CEGMA conjunto de datos y utilizar manualmente Monocercomonoides sp modelos gen curada, el porcentaje de genes recuperados aumentó al 90% (Tabla S1). Por otro conjunto de 163 conservan los genes eucarióticas filogenomico utilizado para analizar el porcentaje de genes recuperadas superó el 95%

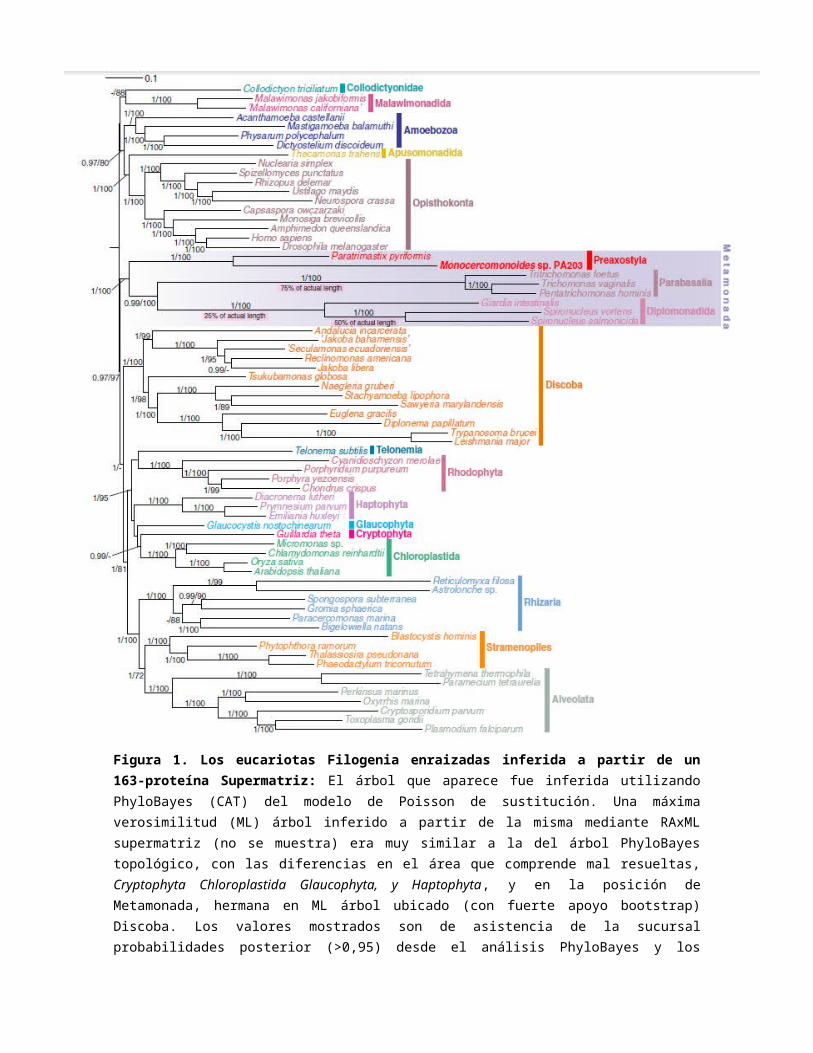
(Tabla S2). Como la última medida exhaustiva, hemos identificado 77 de 78 familias conservados del citosol proteínas ribosomal eucariotas (Tabla S3), con la única excepción de L41e, que es muy corto, difícil de detectar, y no se ha identificado en otros genomas Metamonada. Análisis filogenomico (Figura 1) confirmó la relación de Monocercomonoides sp. P. piriformes y otros Metamonadas y demostró que el linaje Monocercomonoides se forma mucho más cortos en relación con la rama y parabasalidos diplomonados. Todas estas acciones sugieren que los Monocercomonoides sp. reunidos es casi completa de la secuencia del genoma y sus proteínas son codificadas, en promedio, menos que los divergentes de G. intestinalis y T. vaginalis.
Con la primera secuencia en mano de genoma oxymonadas, hemos centrado nuestra atención en uno de los aspectos más desconcertantes de su biología de la naturaleza esquiva de su mitocondria.
Ausencia de proteínas mitocondriales
Sin genes que normalmente están codificados en el genoma mitocondrial (mtDNA) de otros eucariotas fueron encontrados entre las matrices montados, sugiriendo que, como otros, Monocercomonoides metamonadas sp. carece de mtDNA. A continuación, hemos buscado homólogos de proteínas codificadas por el genoma nuclear habitualmente asociadas con la mitocondria o MROs en otros eucariotas. El núcleo de la proteína homóloga de la maquinaria de importación está considerada como fuerte evidencia para el origen común de todas las mitocondrias. Como tal, la presencia de componentes de la translocasas de la membrana externa (TOM) y la membrana interna (TIM), la clasificación y la maquinaria de montaje (SAM) y complejos moleculares mitocondriales (chaperonas Hsp70 y CPN60) en hidrogenosomas, mitosomas y otros MROs demuestra que estos orgánelas están relacionadas con las mitocondrias. Si bien pudimos identificar homologos del citosol en el Monocercomonoides chaperonas sp. secuencia genómica, no pudimos identificar homologos de cualquier componente de la maquinaria de importación mitocondrial (Figura 2A; los procedimientos experimentales; tablas de S3 y S4).
Todas las OPF, con la excepción de G. intestinalis mitosome, son conocidos para la exportación o importación de ATP y otros metabolitos normalmente mediante transportadores de la familia portador mitocondrial (MCF) o, en mitosomas del microsporidium Encephalitozoon cuniculi, por el de tipo bacteriano (NTT) transportadores de nucleótidos. No hemos identificado en el genoma Monocercomonoides sp. cualquier homologo conocido de genes que codifica proteínas de transporte metabolito mitocondrial (Figura 2A; el Cuadro S4).

Grupos Fe-S son cofactores biológicos esenciales asociados con muchas proteínas diferentes y, por lo tanto, son sintetizados de novo en cada organismo a través del árbol de la vida. En Eucariotas, esto se realiza principalmente por el sistema de ensamblaje de ISC mitocondrial y el conjunto de hierro-azufre citosólico (CIA) Sistema. Análisis del genoma Monocercomonoides sp. reveló la presencia de un sistema de la CIA pero una falta completa de los componentes del sistema de ISC (Figura 2A; la tabla S3; los procedimientos experimentales).
No hemos podido identificar cualquiera de las dos enzimas involucradas en la síntesis de un fosfolípido cardiolipina, específicos para la transducción de energía de las membranas. La mayoría de las eucariotas sintetizan cardiolipinas, y el proceso se localiza en las mitocondrias, pero una falta completa de cardiolipina ha sido demostrado experimentalmentepara G. intestinalis, T. vaginalis y E. cuniculi. Además, no pudimos identificar cualquier componente del retículo endoplásmico (RE) Encuentro de estructuras de la mitocondrias (ERMES; en la Figura2A)
Sólo se identificaron dos ortólogos del conjunto de proteínas se suponía localizar a la mitocondria del compartimiento relacionado estrechamente relacionado P. piriformes: aspartato / ornitina carbamoil transferasa familia de proteínas y de nucleótidos de piridina transhidrogenasa. Ni la proteína tiene exclusivamente una localización mitocondrial en eucariotas, y el Monocercomonoides sp. ortólogos no contienen secuencias mitocondriales se suponía la focalización.
Para complementar las búsquedas de homología selectivas basadas también realizamos una búsqueda extensa para homologas de proteínas putativas mitocondriales conocidas mediante una canalización Mitominer basada en la base de datos, el cual fue enriquecido con proteínas mitocondriales identificadas de diversos eucariotas anaerobicas con procedimientos experimentales (OPF). La búsqueda se recuperó 76 Monocercomonoides sp. Las proteínas como candidatos para las funciones en un supuesto mitocondria (Figura 2B; Tabla S5). Análogamente aG. intestinalis, T. vaginalis y E. histolytica, utilizados como controles, los candidatos seleccionados fueron principalmente proteínas que son obviamenteno mitocondrial (por ejemplo, las histonas) o para el que la anotaciónes demasiado general (p. ej. ‘‘Dominio kinasa contienen las proteínas''), lo que indica que la especificidad de la canalización en organismos con mitocondria divergentes es baja. Sin embargo, a diferencia de todos los demásorganismos de control, en el que la búsqueda siempre recuperan al menos unas pocas proteínas distintivo mitocondrial, el conjunto de 76 Monocercomonoides sp. candidatos no contienen tales proteínas. Solamente 11 de los candidatos Monocercomonoides caen en la categoría GO ''metabolización', pero que no reune una evidente vía metabólica. En resumen, este enfoque (Tabla S5) no revelaron

Figura 1. Los eucariotas Filogenia enraizadas inferida a partir de un 163-proteína Supermatriz: El árbol que aparece fue inferida utilizando PhyloBayes (CAT) del modelo de Poisson de sustitución. Una máxima verosimilitud (ML) árbol inferido a partir de la misma mediante RAxML supermatriz (no se muestra) era muy similar a la del árbol PhyloBayes topológico, con las diferencias en el área que comprende mal resueltas, Cryptophyta Chloroplastida Glaucophyta, y Haptophyta, y en la posición de Metamonada, hermana en ML árbol ubicado (con fuerte apoyo bootstrap) Discoba. Los valores mostrados son de asistencia de la sucursal probabilidades posterior (>0,95) desde el análisis PhyloBayes y los valores bootstrap (>50%) a partir de los análisis de ML. Se muestran tres ramas acortó el porcentaje indicado de su longitud real para que quepan en la página. Véase también la tabla S2.

ninguna serie creíble de proteína mitocondrial en Monocercomonoides sp. Como una alternativa a las búsquedas de homología, también hemos intentado identificar proteínas mitocondriales buscando varios tipos de secuencias de la firma. La matriz de proteínas de la mitocondria y las MROs se espera para contener conservado la N-terminal enfocar señales necesarias para el objetivo de la importación en MROs. Se ha realizado la predicción in silico ("hecho por computadora o vía simulación computacional") de focalización mitocondrial señales en la predicha Monocercomonoides sp. proteoma e identificaron 107 proteínas candidatas (Figura 2A; los procedimientos experimentales; Tabla S6A). La presencia de una señal de focalización predicho por sí mismo no prueba la focalización, como esas secuencias de aminoácidos puede aparecen también al azar. La anotación funcional reveló que la mayoría de las proteínas recuperadas por la caída de esta búsqueda en Kyoto Encyclopedia of Genes and Genomes (KEGG) categoría ''información genética procesamiento. ‘‘dada la ausencia de un genoma mitocondrial, o maquinaria de traducción de organelas, es improbable que estas proteínas funcionan en un MRO. Sólo ocho candidatos fueron asignados a la categoría de KEGG ''metabolización',y forman parte de diferentes rutas metabólicas.Por último, sólo tres proteínas fueron pronosticados para tener una señal de focalización mitocondrial y homología con una proteína (Mitominer hidrolasa como familia proteína monos_10795, citosólico 1 TCP/CPN60 chaperonina monos_13132 familia de proteínas, y ribonucleasa Z MONOS_6181). Esto también sugiere que ambos ductos no se pudo recuperar conjuntos específicos de proteínas mitocondriales, sino que sólo se detecta baja especificidad ''ruido''.
La membrana mitocondrial externa acomodan dos clases especiales de proteínas, -barril y cola-anclada (TA), proteínas, que están desprovistos de la N-terminal de focalización y en su lugar usar señales específicas señales C-terminal. Hemos identificado 32 candidatos para las proteínas en el TA predijo proteoma, varios de los cuales parecían ser redirigidos de proteínas. Ninguna de ellas tenía la característica características de proteínas destinadas a la membrana externa mitocondrial (Figura 2A; los procedimientos experimentales; Tabla S6B). También hemos fallado para identificar los candidatos plausibles para -barril proteínas de membrana externa (BOMPs) (Figura 2A; los procedimientos experimentales).
En resumen, nuestro análisis amplio del genoma Monocercomonoides sp. sobre la base de búsquedas de homología y búsquedas específicas para N-terminal y C-terminal especificas fallaron al recuperar las proteínas normalmente asociados con las MROs, incluyendo metabolitos translocasas mitocondrial, transportistas y la ISC para el sistema Fe-S, síntesis de grupos, ERMES cardiolipinas y enzimas responsables de la síntesis. A fin de comprobar que nuestra incapacidad para encontrar cualquier proteínas mitocondriales fiables no es

Figura 2. Estrategias de búsqueda para las proteínas relacionados funcionalmente con la mitocondria en Monocercomonoides (A) buscar estrategias para proteínas mitocondriales y de localización de proteínas firmas en una célula eucariota canónica (se proporcionan detalles adicionales de los procedimientos experimentales): (1) la membrana externa mitocondrial (MOM) orientadas cola-anclada (TA), proteínas (Tabla S6B), (2) las proteínas con un señal de focalización mitocondrial (Tabla S6A), (3), -barril proteínas MOM, (4) 41 distintivos proteínas mitocondriales (Tabla S4), los componentes de Tom y TIM translocasas, cpn60, ERMES, ISC vía componentes complejos, cardiolipina sintasa (CL). (B) para recuperar la canalización homologas semiautomática de las proteínas mitocondriales de proteomas. Se utilizó una base de datos personalizada para la búsqueda de homología de proteínas mitocondriales en la predicción de Proteomas Monocercomonoides sp., Entamoeba histolytica, Giardia intestinalis, Trichomonas vaginalis, Blastocystis sp. subtipo 7, Tabla y Saccharomyces cerevisiae (S5). Véase también tablas S4, S5 y S6.
causada por la posible divergencia sin precedentes de proteínasMonocercomonoides sp. o un fracaso de nuestros métodos, hemos buscado proteínas características de otro sistema celular, hasta ahora no se han observado en Monocercomonoides sp. El complejo de Golgi. En este caso, mediante búsquedas basadas en homología, hemos detectado numerosas proteínas asociadas al aparato de Golgi, incluidos los componentes del COPI, AP-1, AP-3 y AP-4, COG, GARP, TRAPPI y complejos incontables y Rab las GTpasas que regulan el transporte hacia y desde el aparato de Golgi (Tabla S3). Esto sugiere la presencia de Golgi como compartimientos en oxymonadas, a pesar de la ausencia de un aparato de Golgi citológicamente discernible.La ausencia específica de proteínas asociadas a la mitocondria en Monocercomonoides sp. implica la ausencia de una legítima compartimiento mitocondrial. Si es así, entonces ¿cómo funciona la función CELDA Monocercomonoides sin este orgánela?
Metabolismo energético sin mitocondria
Para comparar el metabolismo de Monocercomonoides sp. con sujeción protistas anaerobicas compartimentos mitocondriales, realizamos la anotación manual de proteínas del núcleo de vías del metabolismo energético normalmente asociadas con la presencia y la función de un MRO. Como con muchos otros organismos en segundo lugar redujo la mitocondria, la Monocercomonoides sp. genoma no codifica cualquier enzimas para la generación de energía aeróbica (por ejemplo, TCA ciclo o cadena de transporte de electrones proteínas). Hicimos identificar un conjunto completo de enzimas mediante glicólisis, incluida la alternativa de enzimas para la glucolisis anaerobica, así como la fermentación enzimas anaeróbica piruvato: ferredoxin oxidorreductasa (PFOR) y hidrogenasas FeFe-Tabla (S3). [FeFe] hidrogenasa maturasas estaban ausentes, que no es sin precedentes como también están ausentes de G. intestinalis y E. histolytica, parásitos anaeróbicos que son capaces de citosol de producción de H2. Ni (PFOR) ni (FeFe) hydrogenasa ha pronosticado una focalización mitocondrial secuencia y expresión heteróloga de T. vaginalis sugiere una localización citosólico de (PFOR) (Figura S1).En resumen, Monocercomonoides sp. en el metabolismo de la glucosa, parece producir ATP a través del nivel de pasos de sustrato la fosforilación en un largo camino, la glucólisis y la reducción de co-factores son re-oxidado por fermentación,en última instancia, producir acetato y etanol, o por [FeFe] hidrogenasa produciendo gas hidrógeno. La situación en Monocercomonoides sp. es prácticamente idéntica a la de G. intestinalis y E. histolytica, que de forma independiente han reducido sus mitocondrias para mitosomas y toda la producción de ATP se produce en el citosol. Además extendió la glucólisis, Monocercomonoides sp. contiene un conjunto completo de tres genes que codifican para enzimas implicadas en la ruta deiminasa arginina - deiminasa arginina, ornitina carbamoil transferasa deiminasa y carbamato quinasa. Esta ruta también puede ser utilizada para la

producción de ATP por degradación arginina como en T. vaginalis y G. intestinalis. En G. intestinalis, esta vía produce ocho veces más ATP que el metabolismo del azúcar.
Fe-S Conjunto clúster sin mitocondria
Cada célula eucariota contiene una maquinaria de la CIA, que asiste a las etapas finales de la Asamblea de grupos Fe-S en proteínas eucariotas funcionando en el citosol y en el núcleo. Ocho proteínas mostraron ser involucrados en el sendero de la CIA en la levadura y los seres humanos: Cfd1, NUBP1 (PNI35), NARFL (nar1), CIAO1 (CIA1), Dre2, Tah18, Cia2, y MMS19. Cuatro de ellos (por ejemplo, PNI35 Nar1, Cia1 y Cia2) se conservan entre los eucariotas y también presente en el genoma Monocercomonoides sp. Tabla (S3). No detectamos Cfd1 y MMS19, que faltanen muchos otros eucariotas, y Dre2 y Tah18, en e lque faltan de los protistas anaeróbicas que contengan OPF (incluyendo la E. histolytica, Mastigamo ebabalamuthi, T. vaginalis, G. intestinalis y Blastocystis sp.).
A pesar de la presencia de la vía ISC, comúnmente se sugiere que las mitocondrias y las organelas son esenciales para las células eucariotas porque el sistema ISC mitocondrial juega un papel crítico en la fase inicial de la formación del citosol grupos Fe-S. Aunque el sistema ISC es casi universalmente conservada en vía eucariotas y parece ser la característica común de las mitocondrias y las organelas, los genes que codifican las proteínas de la mitocondria sendero ISC no han sido detectadas en el genoma Monocercomonoides sp. La sustitución funcional del sistema ISC ha sido reportada por sólo dos linajes, Pygsuiabiforma (Breviatea) y Archamoebae. Una movilización de azufre metanoarquea (SUF) o bien un sistema de fijación de nitrógeno bacteriano (NIF) Aparentemente ha sustituido el sistema de ISC en la P. biforma y los linajes Archamoebae, respectivamente. Existen datos contradictorios sobre la localización de la NSI sistema en E. histolytica; sin embargo, el inM. balamuthi sistema, el NIF se localiza en el citosol y el MRO.
El principal problema sigue siendo: ¿Cómo Monocercomonoides sp. formar grupos Fe-S? Inesperadamente, hemos identificado los genes que codifican las cuatro subunidades del sistema: SufB SUF, SufC, y fusionados y SufSSufU Tabla (S3). SufS es un ''2'' de componente de sulfurasa cisteínasu actividad podría ser mejorada por SufE o SufU. En Monocercomonoides sp., se fusiona con SufSSufU, que es una característica única. SufCSufB y pueden formar un complejo de matrices en procariotas, y SufB complejo 2C2 es capaz de unirse y transferir 4FE-4S de grupos a un destinatario apoproteína. Todo sistema de SUF identificado proteínas aparentemente conservan todos los importantes sitios catalíticos (Figura S2) y puede realizar de grupos Fe-S biogénesis novo por sí solos o conjuntamente con la maquinaria de la CIA. El sistema SUF para síntesis de grupos Fe S se

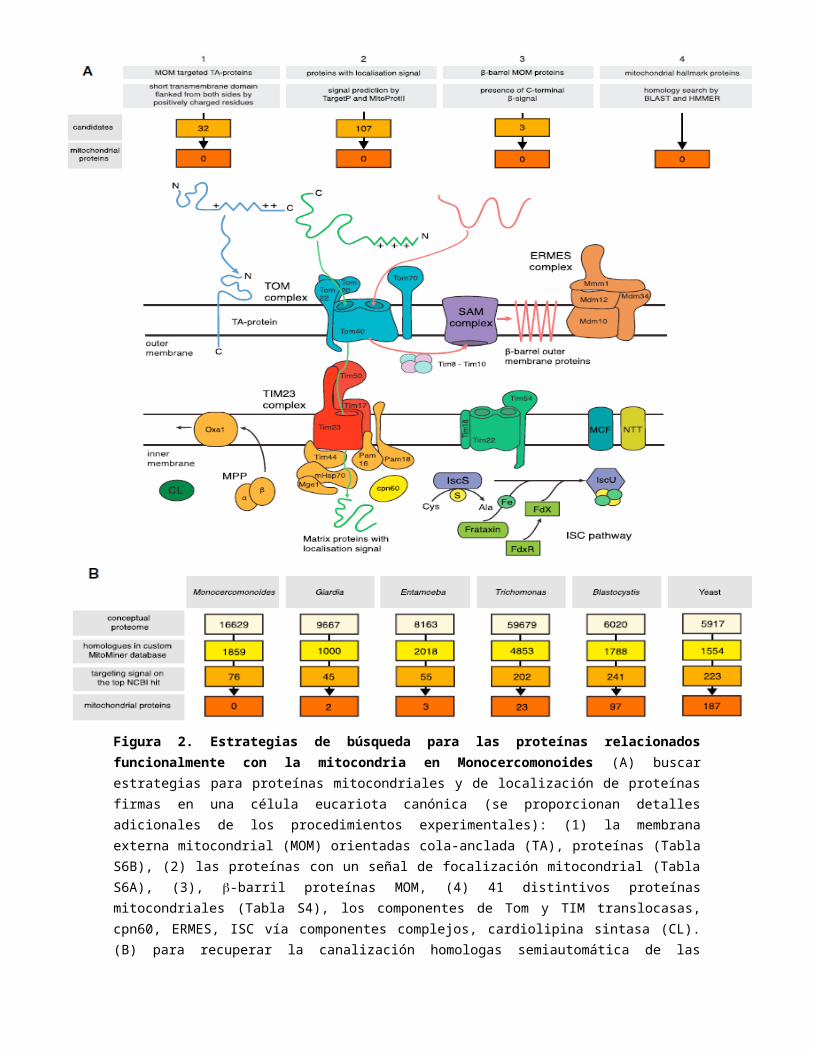
Figura 3. La expresión heteróloga de Monocercomonoides sp. Sistema SUF Proteínas y filogenia de SufB SufC concatenados, y SufS Homologas (A) la expresión heteróloga de Monocercomonoides sp. SufC SufB y proteínas de Trichomonas vaginalis. Monocercomonoides sp. proteínas con una C-terminal de la etiqueta HA fueron expresados en T. vaginalis y visualizados por un anticuerpo anti-HA (verde). La señal del anticuerpo anti-HA no co-localiza con hidrogenosomas teñidos con un anticuerpo anti-enzima málico (rojo). El núcleo teñido con DAPI (azul). Barra de escala, de 10 mm. (B) la expresión heteróloga de Monocercomonoides sp. SufC proteína en Saccharomyces cerevisiae. Monocercomonoides sp. proteínas etiquetadas con GFP se expresaron en S. cerevisiae (verde). La GFP señal no co-localiza con la levadura mitocondrias manchadas por Mitotracker (rojo). Barra de escala, de 10 mm. (C) del árbol enraizado ML SufB SufC concatenados, y SufS secuencias. Apoyo Bootstrap valores superiores a 50 y posterior probabilidades superiores a 0.75 se muestran. Monocercomonoides sp. y Paratrimastix piriformes se muestran en rojo, eukaryotic plástidos y cianobacterias en verde, Blastocystis sp. y Pygsuia biforma en Orange, bacterias y arquea en gris y en azul. Consulte también las figuras S1-S3.
Figura 4. Evolución reductiva de mitocondrias en transición de Metamonadas Hacia un estilo de vida anaeróbico se produjo en un ancestro común de metamonadas y fue seguido por una reducción de las mitocondrias a MROs, acompañado por la pérdida de cristae y genoma, y la transición al metabolismo anaeróbico. La ISC via de grupos fe-S síntesis estuvo presente en un ancestro común metamonada. Otra reducción a un mitosoma tuvo lugar en linaje Giardia intestinalis. Proponemos que en el ancestro común de Paratrimastix piriformes y Monocercomonoides, un sistema SUF adquiridas a través de la LGT desde la bacteria sustituye el sistema ISC-localizada de MRO. Posteriormente, las MROs se perdió completamente en el linaje que llevó a Monocercomonoides sp. Localización de la vía suf en P. piriformes es desconocida.

encuentra en plástidos ,bacterias y arqueas y también se ha encontrado en dos eucariotas microbianos P. biforma y Blastocystis sp.
La presencia de intrones spliceosomalesen la putativa y de Monocercomonoides SufSU SufC confirma que estas proteínas no son procariotas contaminación. Además, hibridación in situ fluorescente (FISH) con sondas gen sufC sufB y demostrado su presencia en el Monocercomonoides sp. núcleo (Figura S3). Importantes, homologos de estas proteínas fueron detectados en la P. piriformes genoma secuenciado, el más cercano a Monocercomonoides relativa. Los componentes de ambos sistemas SUF Monocercomonoides sp. y P. piriformes no contienen señales de focalización mitocondrial reconocible, y nuestros experimentos con la expresión heteróloga de Monocercomonoides sp. SufC SufB y proteínas en T. vaginalis (Figura 3A) y SufC proteína en levaduras (Figura 3b) apoyar una localización citosólico. Los análisis filogenéticos indican que este sistema SUF fue adquirida por un antepasado Monocercomonoides y Paratrimastix por transferencia genética lateral (LGT) de bacterias de forma independiente del resto de SUF-conteniendo células eucariotas (Figura 3C). Proponemos que la adquisición de un sistema SUF citosólico hizo el ancestral sistema de ISC en la mitocondria prescindible, que condujo a la pérdida, y en el linaje Monocercomonoides, a la pérdida completa de la MROs (Figura 4).
Conclusiones
Las mitocondrias y las organelas son actualmente considerada como componentes indispensables de las células eucariotas. La secuencia del genoma de Monocercomonoides sp. informó aquí sugiere que este no es el caso. A pesar de extensas búsquedas, ningún marcador de proteínas mitocondrial como la proteína de membrana translocasas y transportadores de metabolito fueron identificados. Fundamentalmente, la mitocondria ISC específico camino de grupos fe-S Biogénesis está ausente y aparentemente fue reemplazado por un sistema que SUF funciones bacterianas en el citosol. Por otro lado, los genes que codifican para otras características creían estar ausente de estas diferentes células eucariotas, es decir, el cuerpo de Golgi, eran fácilmente identificables. El genoma también contiene genes para trayectos citosólico esenciales del metabolismo energético, aunque sí observamos ejemplos de racionalización metabólicos característicos de otros anaeróbicas o microaerófilos eucariontes.
Reducción de las mitocondrias es conocido desde diversos linajes eucarióticos adaptado al estilo de vida anaeróbica. En Mitosomas Giardia, Entamoeba dispar, y microsporidios representan los casos más extremos de reducción mitocondrial conocida hasta la fecha y, sin embargo, que todavía contienen proteína mitocondrial translocasas reconocibles y generalmente un sistema de ISC. La ausencia específica de todas estas proteínas en el

genoma mitocondrial de Monocercomonoides sp. indica que esta eucariota ha dispensado con el compartimiento mitocondrial completamente. En principio,no podemos excluir la posibilidad de que una mitocondria exista en Monocercomonoides sp., cuya composición proteica hasido alterado por completo. Sin embargo, esa hipotética organela no podía ser reconocida como una mitocondria homóloga por cualquier medio disponible. Sin ningún tipo de pruebas positivas para el último escenario, sugerimos que la completa ausencia de marcadores mitocondriales y vías apunta a la ausencia de buena fe de las mitocondrias. Porque todas las oxymonadas conocidos son simbiontes animales, y homologas mitocondriales están presentes en el cierre de vida libre linaje Paratrimastix hermana, la ausencia de la mitocondria en Monocercomonoides sp. deben ser secundarias. Hipotetizamos que la adquisición del sistema SUF precedió a la pérdida delsistema ISC mitocondrial en el ancestro común de Preaxostyla y permitía la pérdida completa de las mitocondrias en Monocercomonoides sp. estirpe, la primera conocida verdaderamente secundaria eucariota amitocondriaca.
PROCEDIMIENTOS EXPERIMENTALES
Secuenciación de genoma y transcriptoma
Todos los experimentos fueron realizados en el Monocercomonoides sp. PA203 cepa. La cultura (2 l con una densidad celular de aproximadamente 4 3 105 células/ mL) fue filtrada para eliminar la mayoría de las bacterias antes de aislamiento de ADN (cultivo de filtración y detalles adicionales de los procedimientos experimentales). El ADN fue aislado con sangre y tejido DNeasy Kit (QIAGEN). Adn genómico total fue secuenciado utilizando un secuenciador de Genoma 454 GS FLX con reactivos XL. Un total de siete ejecuciones se realizaron de secuenciación, incluidos cuatro escopeta se ejecuta en las bibliotecas con el promedio de la longitud del fragmento de 500 a 800 y tres carreras de 3 kb emparejado a finales de biblioteca. Dos secuencias de RNA (RNA-Seq) se realizaron experimentos utilizando 454 Illumina y plataformas de secuenciación. Los detalles de la secuencia son dados en los procedimientos experimentales complementarios.
Los procedimientos
Ensamblador de Roche Newbler v.2.6 fue utilizado para generar una secuencia de genoma ensamblado desde 454 solo y un par final de lecturas. El montaje final se componía de 2.095 matrices que abarcan casi 75 Mb del genoma. El N50 Tamaño de matrices es de 71,4 KB. Montaje del transcriptoma 454 datos se realizó por Newbler v.2.8 con parámetros predeterminados y Illumina transcriptómica generado datos fueron ensambladas con Trinidad (detalles adicionales de los procedimientos experimentales). El CEGMA fue utilizado para estimar el número de genes eucarióticas conservados en el genoma

Monocercomonoides sp. Tabla general (S1) y la presencia de citosol proteínas eucariotas ribosomal como una medida adicional de exhaustividad (Cuadro S3).
Anotación de genomas y genes
Para la anotación Estructural, Augustus v.2.7, pasa2 y EVM fueron utilizados. Modelos de genes de particular interés fueron evaluados de forma manual con la ayuda de RNA-Seq datos o considerando la conservación con homologas (detalles adicionales de los procedimientos experimentales). Anotación funcional fue asignado a los genes mediante búsquedas por similitud deproteínas predijo usando BLASTP, contra la base de datos de proteínas no redundante NCBI y HMMER3 búsquedas de dominio Pfam hits en la base de datos de familias de proteínas. Anotaciones adicionales se realiza utilizando el servidor de anotación automática KEGG. Los archivos de anotación se encuentran disponibles en la página web http://www.protistologie.cz/hampllab/data.html.
TArn genes fueron pronosticados con tRNAscan-SE; secuencias de rDNA no estaban presentes en el conjunto principal original, pero fueron identificados en no contigs montado en matrices y añadido al conjunto principal. La base de datos del genoma Monocercomonoides sp. fue buscado utilizando el algoritmo TBLASTN y proteoma Monocercomonoides base de datos y seis marcos de traducción de la secuencia genómica fueron registrados utilizando el algoritmo o BLASTP, el perfil modelo ocultos de Markov (HMM) Método de búsqueda desde el HMMER phmmer3 paquetes. Utilizamos una amplia gama de consultas descritas en los procedimientos experimentales complementarios.
Los análisis filogenéticos
Realizamos una serie de máxima probabilidad y análisis filogenético Bayesiana: (1) análisis de filogenomica eucariotas basados en 163 genes y 70 taxones; (2) análisis filogenéticos de genes que codifican para enzimas vía SUF; y (3) gen individual árboles en apoyo de anotación funcional de los genes (detalles adicionales de los procedimientos experimentales).
Predicción de localización subcelular
Para la predicción de localización subcelular Monocercomonoides sp. Proteoma se realizó utilizando TargetP v.1.1 y v.1.101 MitoProt II. TA proteínas fueron identificadas y analizados en función de la presencia de un dominio transmembrana (TMD) de moderada hidrofobicidad, flanqueada por residuos cargados positivamente (detalles adicionales de los procedimientos experimentales). Se identificaron BOMPs, en base a la presencia de una -señal de - C- terminal conservado, usando un conducto descrito previamente.

Proteína mitocondrial Búsqueda utilizando una base de datos Mitominer-Based
Hemos preparado una base de datos personalizada de proteínas mitocondriales para buscar genes que codifican proteínas con localización mitocondrial putativa. La base de datos personalizada se basa en el juego de referencia de la base de datos MitoMiner conteniendo 12 mil 925 proteínas de 11 eucariota proteomas mitocondrial, el cual fue enriquecido por conocidos o previstos de MRO proteínas localizadas de E. histolytica, G. intestinalis, P. biforma, S. salmonicida, T. vaginalis y P. pyriformis. Homologas de proteínas a partir de esta base de datos fueron registrados en la predicción del proteoma de Monocercomonoides sp. y en las previsiones de proteomas de Blastocystis sp., E. histolytica, G. intestinalis, S. cerevisiae, y T. vaginalis,que fueron utilizadas como control base de datos. Mientras busca el control de base datos, las proteínas del organismo buscado fueron eliminadas de la base de datos personalizada. En el último paso, sólo los candidatos eran mantenidos cuyo primer golpe en la base de datos NCBI contiene una señal de focalización mitocondrial predecible (score > 0,5 en TargetP v.1.1 y v.1.101 MitoProtII ) Se dan más detalles adicionales de los procedimientos experimentales.
FISH
Hemos realizado experimentos de FISH con sondas marcadas para determinar si los genes de las proteínas del sistema SUF residen físicamente en los Monocercomonoides sp. genoma o representar la contaminación bacteriana. Los detalles de la preparación de las sondas de etiquetado tendrán en los Procedimientos Experimentales Suplementario. Un litro de Monocercomonoides sp. por cultura se filtró para eliminar las bacterias , y las células se sedimentaron por centrifugación durante 10 min a 2000 g 3 a 4 C. con sondas FISH marcadas con digoxigenina se llevó a cabo de acuerdo con un previamente procedimiento descrito omitiendo el procedimiento de la colchicina. Los núcleos celulares y las sondas se desnaturalizaron bajo un cubreobjetos en un solo paso en 50 ml de 50 % de formamida en 2 3 SSC a 70 C durante 5 min . Se observaron las preparaciones usando un microscopio IX81 (Olympus ) equipado con una cámara IX2- UCB. Las imágenes fueron procesadas utilizando el software Cell ( Olympus ) y 1.42q Image J .
La expresión de proteínas heterólogas en microscopía y Trichomonas vaginalis
El sistema de transfección T. vaginalis fue utilizado para evaluar la localización subcelular de SufB, SufC y obtener PFOR proteínas. Monocercomonoides sp. cDNA de preparación se realiza como se describe para el transcriptoma de secuenciación Procedimiento experimental (suplementarios). Se construye con la etiqueta hemaglutinina (HA) fusionados a 30 Fin de las secuencias codificadoras de los genes estudiados fueron

preparados y expresado en T. vaginalis, un anaerobio Monocercomonoides protistas relacionados con sp. y teniendo un hydrogenosoma (se proporcionan detalles adicionales de los procedimientos experimentales). Monocercomonoides sp. proteínas expresadas en las células T. vaginalis fueron visualizados utilizando técnicas estándar (se proporcionan detalles adicionales de los procedimientos experimentales).
Sistema de expresión de Saccharomyces cerevisiae heterólogo
Este sistema de expresión se utilizó para confirmar los resultados de la T. vaginalis sistema de expresión para la proteína SUFC. El procedimiento fue análogo al se describe en los detalles de los procedimientos experimentales suplementarios.
NÚMEROS DE ACCESO
Los datos de secuencia del genoma leer (número de experimento), SRX1470187 el 454 transcriptoma lee secuencia usando el 454 plataforma (experimento número SRX1453820), y el transcriptoma Illumina leer secuenciados utilizando la plataforma Illumina (número de experimento SRX1453675) han sido depositados a leer archivo de secuencias NCBI bajo el número de SRA: SRP066769. El número de acceso para el Monocercomonoides sp. PA203 genoma informar en este documento es GenBank: LSRY00000000. la adhesión número para el proyecto 454 transcriptoma informa en este documento es GenBank : GEEG00000000 . El número de acceso para el proyecto transcriptoma Illumina informa en este documento es GenBank: GEEL00000000. Las versiones descritas en este documento son versiones LSRY01000000, GEEG01000000, y GEEL01000000. Más informaciones adicionales sobre el análisis del genoma se puede encontrar en http: // www.protistologie.cz/hampllab/data.html.
INFORMACIÓN SUPLEMENTARIA
Información adicional incluye los procedimientos experimentales Suplementarios, tres figuras, tablas y seis y se pueden encontrar con este artículo en línea en http://dx.doi.org/10.1016/j.cub.2016.03.053.